host range of the violet root rot fungus, helicobasidium purpureum pat
TRANSCRIPT

[ 488 J
Trans. Brit. mycol. Soc. 45 (4), 488-494 (1962).
HOST RANGE OF THE VIOLET ROOT ROT FUNGUS,HELICOBASIDIUM PURPUREUM PAT.
By T. F. HERING*
Botany School, University of Cambridge
The host range of Helicobasidium purpureum was studied in a glasshouse experiment. Seventeen species, in eight families, developed infection cushions ofviolet root rot. Over 10 weeks, no greater losses in viability of sclerotia wereassociat ed with susceptible roots than with resistant ones. In a study on theeffect of root systems on germination of sclerotia, Lotus comiculatus, a resistantspecies, induced a significant increase in germination in 10 weeks. Trifoliumpratense, a susceptible species, significantly reduced the percentage of germination , compared with a control series in fallow soil.
The aim of the experiments here described was twofold. First, theyconstituted an attempt to study the host range of H. purpureum. Over 120species, of forty families, have been recorded as subject to attack by H.purpureum (Duggar, 1915; Viennot-Bourgin, 1949). Although Eriksson(1913) and others have claimed that parasitic races, adapted to particularhost species, exist, the majority of evidence from cross-inoculation andfield observation does not support this view (Buddin & Wakefield, 1924,1927; Baudys, 1929; Dana & Wolff, 1931). A study of the range of hostsattacked by a pure strain ofthe fungus was therefore planned as part of thepresent investigation.
Secondly, an attempt was mad e to observe the effect of roots of higherplants on the survival of sclerotia in soil. It is now known that restingbodies of many parasitic organisms are stimulated to germinate by substances diffusing from susceptible hosts (Brown, 1946) and this effect hasbeen demonstrated for fungus sclerotia by Coley-Smith & Hickman (1957).Of particular interest would be the discovery of a resistant species capableof inducing germination of sclerotia. Hull & Wilson (1946) showed thatthe amount of H. purpureum in an infested soil can be greatly reduced bylifting a susceptible crop early, but such measures are more safely carriedout with a resistant species. MacFarlane (1952a, b) showed that thedegree of infestation of soil by Plasmodiophora brassicae Woron. could bereduced by a resistant trap-crop.
In the first experiment to be described below, forty-five species, including both crop plants and common weeds, were tested for susceptibility toH. purpureum, and an attempt was made to trace the effect of roots onsurvival of sclerotia, by means of viability tests. Five species were selectedfor a more intensive study in the second experiment to be described. Theexperimental work was performed with a strain of H. purpureum isolated byDr P. G. Valder from infected sugar beet, being his isolate B (Valder,
* Present address : The Nature Conservancy, Merlewood R esearch Station, Grangeover-Sands, Lanes.

Helicobasidium purpureum. T. F. Hering 489
1958). It was maintained by culturing on 5 %malt agar, and it remainedconstant in cultural characters during the period of the experiments.
GENERAL METHODS
Production of sclerotia in pure culture
The method was that devised by Valder (1958). Samples of pre-soakedrye grains weighing 5 g. were placed in 250 ml. Erlenmeyer flasks with250 g. sand, the mixture being shaken before the addition of 15 ml. water.The flasks were autoclaved for I hr. at 15 lb. pressure and then shaken toloosen the contents; they were inoculated with a 3 mm. disk cut submarginally from a 2-4-week-old colony of H. purpureum on 5 %malt agar.The inocula were left central in the flasks, which were incubated for 10weeks at 22'5° C. in the dark. Two weeks after inoculation, the flasks werecapped with moisture-proof cellophane and they were shaken fortnightlyto break up the mycelium. At the end of IO weeks nearly all the rye grainshad been converted into dark brown sclerotia of H. purpureum, often withexternal reddish brown incrustations. The sclerotia were of fairly uniformsize; a previous test had shown that viability at IO weeks was IOO %.
Testing sclerotia for viability
The following technique, devised by Valder (1958), proved suitable fortesting viability of sclerotia recovered from soil. Slices, 1-1'2 ern. thick,were cut from washed sound carrots and placed in pairs in previouslyautoclaved Petri dishes, containing 50 g. sand and IO ml. water. Foursclerotia were placed in each dish, touching the intact carrot epidermis.The dishes were incubated in polythene bags for up to 5 weeks at 22'5 °C.,and within this time viable sclerotia produced visible lesions, with characteristic mycelial strands and infection cushions of H. purpureum.
THE FIRST GLASSHOUSE EXPERIMENT
Crop plants and weeds offorty-five species were sown or transplanted inseedling boxes (14 x 8t in.) filled to a depth of 2t in. with sieved soil fromthe University of Cambridge Botanic Garden. This soil is a light loam ofneutral reaction. Three boxes were allotted to each species; the numberof plants per box varied with the size of the plants. When the plants hadbecome established, ten sclerotia of H. purpureum were buried in each boxIt in. deep, spaced out and at least I in. from the nearest rootstock. At6 and 10 weeks, four sclerotia were taken from each box and tested forviability. With annuals, roots were examined for disease at the time offruiting; roots of perennials were examined 15 weeks after the introductionof the sclerotia. Temperatures in the glasshouse were recorded daily(Table I).
Results of sclerotial viability tests and of examination of plants fordisease lesions are presented in Table 2. In some cases results of viabilitytests are incomplete, because sclerotia were destroyed in the test dishes byfly larvae. With susceptible species, the percentage of individuals attacked
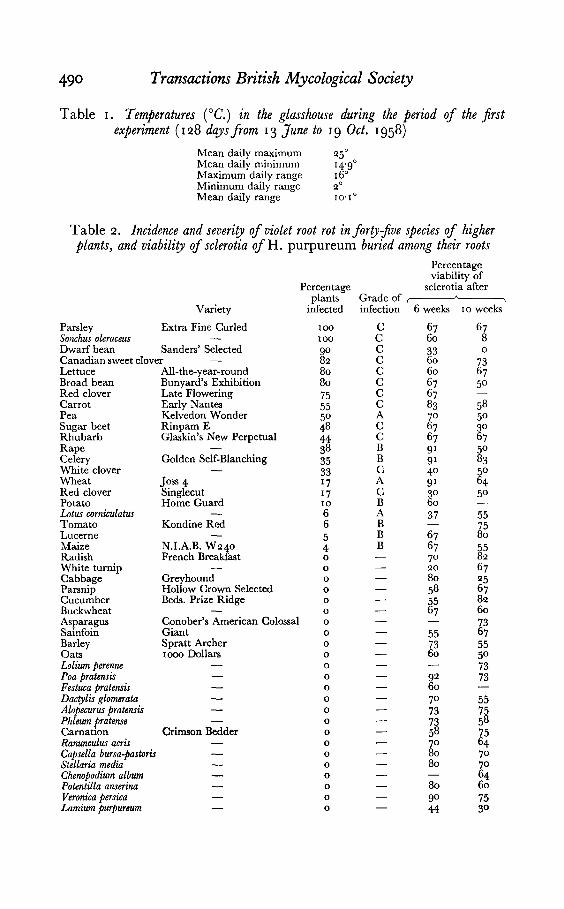
490 Transactions British Mycological Society
Table I. Temperatures (Ce.) in the glasshouse during the period of the firstexperiment (128 days from 13 June to 19 Oct. 1958)
Mean daily maximum 25°Mean daily minimum 14'9°Maximum daily range 16°Minimum daily range 2°Mean daily range 10'1°
Table 2. Incidence and severiry ofviolet root rot inforty-fioe species of higherplants, and oiability of sclerotia ofH. purpureum buried among their roots
Percentageviability of
Percentage sclerotia afterplants Grade of (
Variety infected infection 6 weeks 10 weeks
Parsley Extra Fine Curled 100 C 67 67Sonchus oleraceus 100 C 60 8Dwarf bean Sanders' Selected 90 C 33 0Canadian sweet clover 82 C 60 73Lettuce All-the-year-round 80 C 60 67Broad bean Bunyard's Exhibition 80 C 67 50Red clover Late Flowering 75 C 67Carrot Early Nantes 55 C 83 58Pea Kelvedon Wonder 50 A 70 50Sugar beet Rinpam E 48 C 67 30Rhubarb Glaskin's New Perpetual 44 C 67 67Rape 38 B 91 50Celery Golden Self-Blanching 35 B 91 83White clover 33 C 40 50Wheat Joss 4 17 A 91 64Red clover Singlecut 17 C 30 50Potato Home Guard 10 B 60Lotuscorniculatus 6 A 37 55Tomato Kondine Red 6 B 75Lucerne 5 B 67 80Maize N.LA.B. W240 4 B 67 55Radish French Breakfast 0 70 82White turnip 0 20 67Cabbage Greyhound 0 80 25Parsnip Hollow Crown Selected 0 58 67Cucumber Beds. Prize Ridge 0 55 82Buckwheat 0 67 60Asparagus Conober's American Colossal 0 73Sainfoin Giant 0 55 67Barley Spratt Archer 0 73 55Oats 1000 Dollars 0 60 50Loliumperenne 0 73Poa pratensis 0 92 73Festuca pratensis 0 60Dactylis glomerata 0 70 55Alopecurus pratensis 0 73 75Phleum pretense 0 73 58Carnation Crimson Bedder 0 58 75Ranunculus acris 0 70 64Capsella bursa-pastoris 0 80 70Stellaria media 0 80 70Chenopodium album 0 64Potentilla anserina 0 80 60Veronica persica 0 90 75Lamium purpureum 0 44 30

Helicobasidium purpureum. T. F. Hering 491
was noted, and the severity of infection of individual plants was gradedas follows:
A. Hyphae have grown along roots, forming an investing layer, butthere are no infection cushions.
B. Infection cushions are present, but the root tissue appears healthy.C. Infection cushions are surrounded by violet or brown patches of
discoloured host tissue; a rot extends into the root cortex.Infection cushions were found in this experiment on seventeen species,
of eight families, and severe violet root rot on members of five families(Leguminosae, Umbelliferae, Chenopodiaceae, Polygonaceae, Compositae). It is therefore clear that the wide host range of H. purpureum is acharacteristic of individual pure strains, and is not due to the presence of alarge number of distinct parasitic races. However, at least one feature ofthese results suggests that this strain of H. purpureum is in some degreespecialized. Neither lucerne nor asparagus became diseased to any extent,although violet root rot is a well-known disease of both. This result recallsEriksson's (19 I 3) observation that his strain of H. purpureum, isolated fromcarrot, was only weakly parasitic on clover and lucerne. Other speciespreviously described as susceptible, but showing complete resistance inthis experiment, were several species of grasses. Disease escape by annualssuch as Stellaria media and Chenopodium album may be attributable to thevery rapid flowering and death of these plants under glasshouse conditions.One new host, maize, was added to the list of known host plants; theattack, however, was confined to a single individual and caused no discernible damage.
THE SECOND GLASSHOUSE EXPERIMENT
Of the species which had proved resistant to H. purpureum in the firstexperiment, two were associated with poor survival among sclerotia. Thesetwo, Lotus corniculatus and Lamium purpureum, were selected for further study.For comparison it was planned to include two susceptible species, carrotand red clover; and one resistant species, Ranunculus aeris, that hadexercised no apparent effect on viability of sclerotia. When the experimentwas set up, R. aeris was not available in sufficient quantity, and R. repenswas substituted. R. repens proved an unsatisfactory substitute, since itbecame infected.
The technique used for this experiment was based on one described byColey-Smith & Hickman (1957). The soil containers were glass jam jars(diam. 8'5 cm., height 12 cm.). Each received 700 g. sieved soil from theCambridge Botanic Garden. Before being filled, the jars were lined withwashed nylon mesh (40 meshjcm.; thread thickness 0'1 mm.). Ten pureculture sclerotia were lodged in each jar, between the nylon mesh and theglass, I in. below the neck. This position made it easy to obtain visualestimates of germination. The soil was brought to 60 %moisture-holdingcapacity with distilled water, and thereafter made up to weight every2 days. At the time of setting-up, red clover (Late Flowering) and carrot(Early Nantes), grown from seed and then 6 weeks old, were transplantedinto the jars; the other species similarly transplanted were wild plants.Three plants were transplanted in each jar (two with R. repens), and five

492 Transactions British Mycological Societyjars were allotted to each species, with five unplanted jars as controls. Todiscourage growth ofalgae, the jars were enclosed in black polythene bags,and the soil was kept free of weeds. The temperatures in the glasshousewere recorded daily (Table 3).
Estimates of sclerotium germination were made at 6 and 10 weeks.Germinating sclerotia were readily distinguished by a vigorous outgrowthof strands and single hyphae. After 10 weeks the plants were lifted andexamined for disease (Table 4).
Table 3. Temperatures (Ce.) in the glasshouse during the period of thesecond experiment (70 days from 9 May to 19 July 1959)
Mean daily maximum 28'1 0
Mean daily minimum 13'7 0
Maximum daily range 23°Minimum daily range 3°Mean daily range 14'4
,6 weeks 10 weeks
Percentageplants
infected
Table 4. Incidence of violet root rot infive species of higher plants, andgermination of sclerotia of H. purpureum buried among their roots
Percentage germinationof sclerotia after
CarrotRed cloverLotus corniculatusRanunculus repensLamiumpurpureumControl
(unplanted)
8753
o60
o
o50186
24
444
72
3210
44
6 weeks sample10 weeks sampleTotal
6 weeks sample10 weeks sampleTotal
A comparison of the viabiliry of sclerotia incubated among theroots of susceptible andresistant species
Totalsclerotia Percentage
recovered viability Probability*
188 63'3 >0'1169 54'4 >0'1357 59'1 >0'1
240 65'8275 63'65 15 64'7
Resistant(28 spp.)
Susceptible](17 spp.)
Table 5.
* The probability that sclerotia recovered from contact with susceptible and resistant rootsare members of a single population,
t In this table, species listed as susceptible are those which developed infection cushions inthis experiment ; resistant species are all others ,
EFFECT OF HOST ROOTS ON GERMINATION
One of the questions which it was hoped to answer in this investigationwas whether susceptible roots stimulated sclerotia to germinate and thusto lose viability. To answer this question, the results ofthe first experimenthave been analysed in Table 5.

Helicobasidium purpureum. T. F. Hering 493
From this analysis, it can be concluded that contact either with susceptible roots or with substances diffusing from them causes no significantlowering of viability of sclerotia over a period of 10 weeks. It would besurprising if, in the long run, sclerotia did not germinate and hence loseviability in the presence of susceptible roots, but it is possible that in thisexperiment such an effect was being masked by the response of thesclerotia to other factors. Two aspects of the results of the second experiment (T able 5) suggest this:
(I) Although not susceptible, Lotus corniculatus was associated with ahigh level of sclerotium germination; the level found with two susceptiblecrops, carrot and Ranunculus repens, and in the control soil, was significantlyless than this. It is interesting in this connexion to note Viennot-Bourgin's(1949) empirical recommendation of L. corniculatus as a suitable crop forland heavily infested with H. purpureum.
(2) The effect of red clover in the second experiment was the reverse ofthe effect of L. corniculatus. Here eight of the fifteen plants became infected,but there were no visible changes in the sclerotia. This shows that visualassessment is not sufficiently sensitive to reveal a very slight degree ofsclerotium germination. Clearly some of these sclerotia must have germinated to a very limited extent, sufficient to cause infection, but thisgermination was less noticeable than that occurring in the control. Noexplanation can be offered for the retardation of germination which isapparently induced by red clover. The second experiment failed to substantiate the indication that Lamium purpureum stimulated germination ofsclerotia, and the low viability associated with this species in the firstexperiment must have had some other cause.
If the extremes of behaviour of sclerotia shown in the presence of redclover and of Lotus corniculatus, respectively, are taken into account, it isnot surprising that the first experiment failed to show a simple correlationbetween the susceptibility of a species to H. purpureum and loss of viabilityin associated sclerotia of the fungus.
I am indebted to Dr S. D. Garrett, who suggested this study, for guidance and encouragement. The work was carried out during the tenure ofa Studentship from the Agricultural Research Council.
REFERENCES
BAUDYS, E. (1929). Studium houby Rhizoctonia violacea. Bull. czechosl. Acad. Agric. 5,693-697. (Abstr . in Rev. appl. Mycol. 9, 219).
BROWN, R. (1946). Biological stimulation in germination. Nature, Lond., 157,64-69.BVDDIN, W. & WAKEFIELD, E. M. (1924). Some observations on th e growth ofRhizoctonia
crocorum (Pers.) DC. in pure culture. Ann. appl. Biol. II, 292-3°9.BUDDIN, W. & WAKEFIELD, E. M. (1927). Studies on Rhizoctonia crocorum (Pers.) DC. and
Helicobasidiumpurpureum (T ul.) Pat. Trans. Brit. mycol. Soc. I~, 11 6-14°.COLEy-SMITH, J . R. & HI CKMAN, C . J. (1957). Stimulation ofsclerotiu m germination in
Sclerotiumcepiuorum Berk. N ature, Lond., 180, 445.DANA, B. F. & \VOLFF, S. E. (1931). The occurrence of violet root rot in central Texas.
Phytopathology, IU, 557-558.D UGGAR, B. M. (1915). Rhizoctonia crocorum (Pers.) DC. and R. solani Kuhn (Cortu ium
vagum B. & C.) with notes on other species. Ann. 11'10. bot. Gdn, 2, 403-458.

494 Transactions British Mycological SocietyERIKSSON, J. (1913)' Etudes sur la maladie produite par la Rhizoctone violacee, Rev.
gen. Bot. 25, 14-30.HULL, R. & WILSON, A. R. (1946). Distribution of violet root rot (Helicobasidium
purpureum Pat.) of sugar beet and preliminary experiments on factors affecting thedisease. Ann. appl. Biol. 33, 420-433.
MACFARLANE, I. (1952 a). Factors affecting the survival of Plasmodiophora brassicae Wor.in the soil and its assessment by a host test. Ann. appl. Biol. 39, 239-256.
MACFARLANE, I. (1952 b). Clubroot of cruciferous plants. Rep. Rothamsted expo Sta.,pp.89-90.
VALDER, P. G. (1958). The biology of Helicobasidium purpureum Pat. Trans. Brit. mycol.Soc. 41, 283-308.
VIENNOT-BoURGIN, G. (1949). Les champignons parasites, pp. II79-1 186. Paris: Massonet Cie.
(Accepted for publication I November 196 I )