horseradish peroxidase localization of the mylohyoid ... · experiments were performed with...
TRANSCRIPT

Horseradish peroxidase localization of the mylohyoid motoneurones in the rabbit
Henryk Gromysz, Witold A. Karczewski, Anna ~ o s m a l ' and Andrzej Kukwa
Department of Neurophysiology, Medical Research Centre, Polish Academy of Sciences, 3 Dworkowa St., 00-784 Warsaw, Poland; ' ~ e ~ a r t m e n t of Neurophysiology, Nencki Institute of Experimental Biology, 3 Pasteur St., 02-093 Warsaw, Poland
Abstract. Experiments were performed with anaesthetized (urethan and chloralose) spontaneously breathing rabbits. Horseradish peroxidase (HRP) was injected (1 mg) with a Hamilton syringe into the end of the sectioned mylohyoid nerve. The HRP- labelled neurones could be localized in the motor nucleus of the trigeminal nerve (N.V.mt.). It was found that the labelled neurones are present in the caudal and intermediate part of the nucleus. However, their largest agglomerations were noted in the caudal part, where they occupied the entire frontal section of the nucleus. It is assumed that the N.V.mt. directing their axons to the muscles of the upper respiratory airways, simultaneously send collaterals to the central respiratory neurones participating in the regulation of the respiratory rhythm. According to this hypothesis the neurones of the N.V.mt. may ensure synchronisation between the upper and lower respiratory system.
Key words: mylohyoid nerve, horseradish peroxidase, control of breathing, respiratory pattern, motor nucleus of the trigeminal nerve

422 H. Gromysz et al.
The mylohyoid nerve is one of the branches of the trigeminal nerve. It is generally accepted that this nerve by activating the mylohyoid muscles, takes part in controlling the lumen of the upper res- piratory system.
It was previously found that expiratory activity of the mylohyoid nerve is stimulated by the activity of pulmonary stretch receptors (Kukwa et al. 1989) and, on the other hand, activity of this nerve dim- inishes due to intravenous injection of ketamine and low percentage solution of alcohol (Kukwa et al. 1989, Gromysz et al. 1990).
Expiratory neurons of the pons, whose phasic ac- tivity depended on information from pulmonary stretch receptors was first described in rabbits (Gromysz and Karczewski 1973) and cats (Feldman et al. 1976a, b). In recent years studies on rabbits re- vealed a group of expiratory neurones in the motor nucleus of the trigeminal nerve (N.V.mt.). As dem- onstrated by histological control, the largest cluster of these neurones is found in the caudal part of the nucleus. These neurones are excited by the lung mechanoreceptors, and their electric stimulation with a single stimulus elicits a short-latency inhibi- tion of the phrenic nerve activity (Gromysz et al. 1988). Inversely, pharmacological (lignocaine) elimination of these neurones results in apneustic respiration and abolition of the Breuer-Hering infla- tion reflex (Gromysz and Karczewski 1990).
The expiratory activity recorded from the N.V.mt. and that of the mylohyoid nerve respond to the excitatory and inhibitory stimuli in a similar way (Gromysz et al. 1988, Kukwa et al. 1989, Gromysz and Karczewski 1990). The above quoted data seem to indicate that the activity of the mylohyoid nerve and those recorded from the N.V.mt. should have a common driving source. Knowing the localization of the expiratory neurones of the N.V.mt. (Gromysz et al. 1988, Gromysz and Karczewski 1990) we de- cided to check the localization of the mylohyoid nerve motoneurones. For this purpose HRP was ap- plied to the mylohyoid nerve. The results were ex- pected to eventually confirm the hypothesis of Gromysz and Karczewski (1990) that the N.V.mt. as the anatomic substrate of the pneumotaxic centre
(beside the role clasically ascribed to it) has an im- portant function in the central and reflex regulation of the respiratory rhythm.
The experiments were performed on four rabbits (3-3.5 kg) under urethane (800 mglkg) and chlo- ralose (30 mglkg) anaesthesia, breathing sponta- neously. After dissecting the phrenic and mylohyoid nerves their bioelectrical activity was recorded under control conditions. The mylohyoid nerve ex- hibits an expiratory activity. It is characterized by the highest number of action potentials appearing at the beginning of expiration and decrementally de- creasing as the expiratory phase develops (Fig. 1).
N.V. V kmbd
Fig. 1 . Control recording of expiratory activity of the mylo- hyoid nerve (N.V.) and the phrenic nerve (N.Phr.).
After recording, the mylohyoid nerve was taken off the electrode and 1 mg of HRP was injected into its ending with a Hamilton syringe in 3 ym physiologi- cal saline. After 45 h the animals were sacrificed and perfused (under deep anaeshesia) with McE- vans solution and a fixing mixture (I % paraformal- dehyde and I % glutaraldehyde in 0.1 M phosphate buffer, pH 7.2-7.4). In the end phase of fixation 10% sucrose solution in phosphate buffer was intro- duced. Blocks from the level of the studied structure were stored in 10% and next in 30% sucrose solution for about 24 h. Histological sections 50 ym thick were cut in the frontal plane on a freezing microtome. The sections were incubated in tetramethylbenzidine solu- tion according to Mesulam procedure ( 1982).
In all cases of HRP injection into the mylohyoid nerve, the localization of the labelled neurones is very similar and will be presented globally.
Mylohyoid motoneurones 423
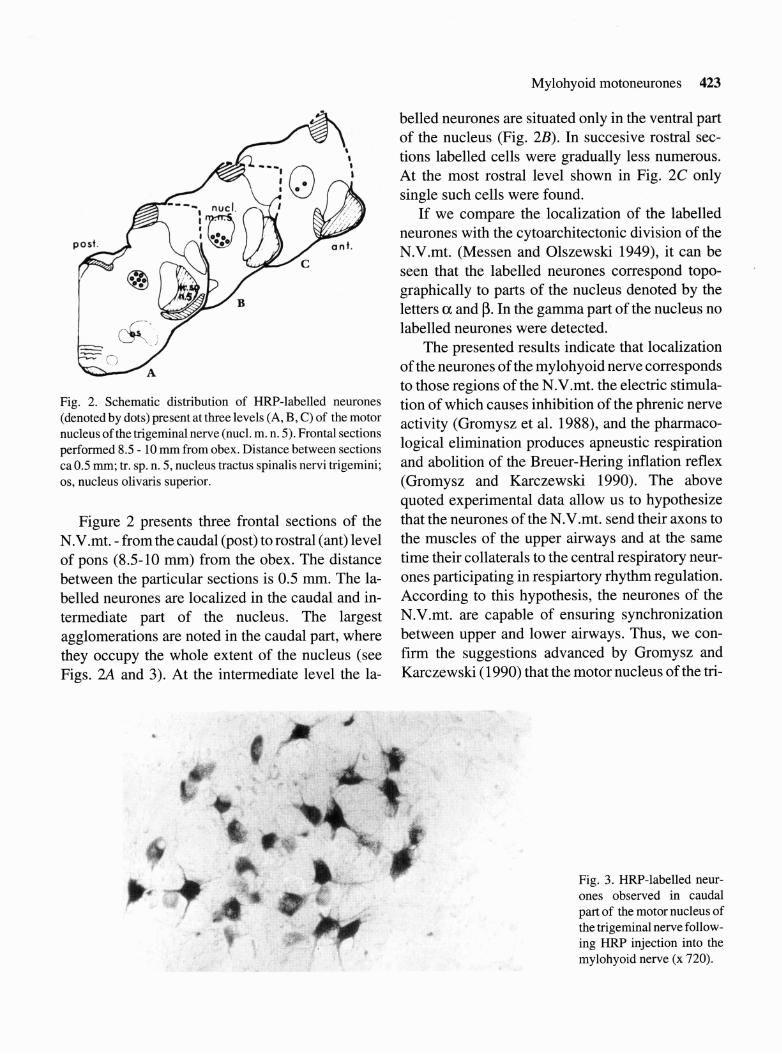
Fig. 2. Schematic distribution of HRP-labelled neurones (denoted by dots) present at three levels (A, B, C) of the motor nucleus of the trigeminal nerve (nucl. m. n. 5). Frontal sections performed 8.5 - 10 mm from obex. Distance between sections ca0.5 mm; tr. sp. n. 5, nucleus tractus spinalis nervi trigemini; os, nucleus olivaris superior.
Figure 2 presents three frontal sections of the N.V.mt. - from the caudal (post) to rostral (ant) level of pons (8.5-10 mm) from the obex. The distance between the particular sections is 0.5 mm. The la- belled neurones are localized in the caudal and in- termediate part of the nucleus. The largest agglomerations are noted in the caudal part, where they occupy the whole extent of the nucleus (see Figs. 2A and 3). At the intermediate level the la-
belled neurones are situated only in the ventral part of the nucleus (Fig. 2B). In succesive rostral sec- tions labelled cells were gradually less numerous. At the most rostral level shown in Fig. 2C only single such cells were found.
If we compare the localization of the labelled neurones with the cytoarchitectonic division of the N.V.mt. (Messen and Olszewski 1949), it can be seen that the labelled neurones correspond topo- graphically to parts of the nucleus denoted by the letters a and p. In the gamma part of the nucleus no labelled neurones were detected.
The presented results indicate that localization of the neurones of the mylohyoid nerve corresponds to those regions of the N.V.mt. the electric stimula- tion of which causes inhibition of the phrenic nerve activity (Gromysz et al. 1988), and the pharmaco- logical elimination produces apneustic respiration and abolition of the Breuer-Hering inflation reflex (Gromysz and Karczewski 1990). The above quoted experimental data allow us to hypothesize that the neurones of the N.V.mt. send their axons to the muscles of the upper airways and at the same time their collaterals to the central respiratory neur- ones participating in respiartory rhythm regulation. According to this hypothesis, the neurones of the N.V.mt. are capable of ensuring synchronization between upper and lower airways. Thus, we con- firm the suggestions advanced by Gromysz and Karczewski (1990) that the motor nucleus of the tri-
Fig. 3. HRP-labelled neur- ones observed in caudal part of the motor nucleus of the trigeminal nerve follow- ing HRP injection into the mylohyoid nerve (x 720).

424 H. Gromysz et al.
geminal nerve (Pontine Intermediate Respiratory Karczewski, P. Grieb, J. Kulesza and G. Bonsingnore).
Group) is the anatomical substrate of the pneumo- 'lenum New York, p.183-186. Gromysz H., Karczewski W.A. (1990) Motor nucleus of the taxic centre. This structure (being under the excita- v-L nerve and con&ol of breathing. preuer-Henng
tory influence the vagal plays a and apneustic breathing). ActaPhysiol. Pol. 41: 147-155. and important role in the inspiratory "off switch" Gromysz H., Karczewski W.A., Kukwa A. (1990) Breuer- mechanism. Herning reflexes in ketamine - induced apneustic brea-
thing in the rabbit. Acta Physiol.Pol.41: 157-163.
Feldman J.L., Cohen M.I., Wolotsky P. (1976a) Powerful in- hibition of pontine respiratory neurons by pulmonary af- ferent activity. Brain Res. 104: 341-346.
Feldman J.L., Cohen I., Wolotsky P. (1976b) Phasic pulmon- ary afferent activity drastically alters the respiratory modulation on neurons in the rostra1 pontine pneumotaxic center. In: Respiratory centers and afferent system (Ed. B. Duron). INSERM, Paris, 59: 95-105.
Gromysz,H., Karczewski W.A. (1973) Responses of respir- atory neurons of the rabbit to some excitatory and inhibi- tory stimuli. Acta Neurobiol. Exp. 33: 245-261.
Kukwa A., Gromysz H., Jernajczyk U., Karczewski W.A. (1989) Studies on the mechanisms of obstructive sleep apnea. Acta Physiol. Pol. 40: 473-478.
Messen H., Olszewski J. (1949) A cytoarchitectonic atlas of the rhombocephalon of the rabbit. Karger S., New York, p. 1-52.
Mesulam M.M. (1982) Principles of horseradish peroxidase neurohistochemistry and their applications for tracing neuronal pathways - transport, enzyme histochemistry and ligth microscopic analysis. In: Tracing neural connections with horseradish peroxidase. (Ed. M.M. Musulam). John Willey and Sons, New York, p. 1-152.
Gromysz H., Kukwa A., Jernajczyk U., Karczewski W.A. (1988) Trigeminal nerve breathing and sleep apnea. In: Control of breathing during sleep and anesthesia (Ed. W.A. Received 9 February 1993, accepted 20 April 1993