hiv-1 tat protein increases the permeability of brain endothelial cells by both inhibiting occludin...
TRANSCRIPT

B R A I N R E S E A R C H 1 4 3 6 ( 2 0 1 2 ) 1 3 – 1 9
Ava i l ab l e on l i ne a t www.sc i enced i r ec t . com
www.e l sev i e r . com/ loca te /b ra i n res
Research Report
HIV-1 Tat protein increases the permeability of brainendothelial cells by both inhibiting occludin expression andcleaving occludin via matrix metalloproteinase-9
Ruifen Xua, b, 1, Xuyang Fengc, 1, Xin Xied, 1, Jin Zhangb, Daocheng Wua,⁎, Lixian Xub,⁎aKey Laboratory of Biomedical Information Engineering of Education Ministry, School of Life Science and Technology,Xi'an Jiaotong University, Xi'an, ChinabDepartment of Anesthesiology, School of Stomatology, Fourth Military Medical University, Xi'an, ChinacDepartment of Cardiology, Xijing Hospital, Fourth Military Medical University, Xi'an, ChinadRege Lab of Tissue Engineering, Department of Bioscience, Faculty of Life Science, Northwest University, Xi'an, China
A R T I C L E I N F O
⁎ Corresponding authors. Fax: +86 28 8477611E-mail addresses: [email protected]
1 Ruifen Xu, Xuyang Feng, and Xin Xie con
0006-8993/$ – see front matter © 2011 Elseviedoi:10.1016/j.brainres.2011.11.052
A B S T R A C T
Article history:Accepted 28 November 2011Available online 7 December 2011
Brain homeostasis is maintained by the blood–brain barrier (BBB), which prevents theentrance of circulating molecules and immune cells into the central nervous system. TheBBB is formed by specialized brain endothelial cells that are connected by tight junctions(TJ). Previous studies have proven that the Tat protein of human immunodeficiency virustype 1 (HIV-1) alters TJ protein expression. However, the mechanisms by which thealterations occur have not been characterized in detail. In this study, primary humanbrain microvascular endothelial cells (HBMEC) were exposed to recombinant HIV-1 Tat pro-tein, and the effects on occludin were observed. Tat treatment decreased occludin mRNAand protein levels. This effect was partially abrogated by addition of the RhoA inhibitorC3 exoenzyme and the p160-Rho-associated coiled kinase (ROCK) inhibitor Y-27632. Mean-while, Tat also induced MMP-9 expression. RNA interference targeting MMP-9 reduced boththe paracellular permeability of Tat-treated HBMEC and the concentration of soluble occlu-din in supernatants from the cells. Taken together, these results show that the HIV-1 Tatprotein disrupts BBB integrity, at least in part by decreasing the production of occludin.
© 2011 Elsevier B.V. All rights reserved.
Keywords:HIV-1 TatBlood–brain barrierTight junctionMMP-9Occludin
1. Introduction
The blood–brain barrier (BBB) acts as an interface between thebrain and other tissues. The BBB can protect the brain fromcirculating molecules, lymphocytes and harmful compound(Correale and Villa, 2009). The BBB is an essential part of theneurovascular unit (NVU), and far from being a simple physi-cal barrier, is a heterogeneous, complex and dynamic tissue
5..edu.cn (D. Wu), xulixiantributed equally to this w
r B.V. All rights reserved.
(Nag et al., 2011). The integrity of the BBB is determined bythe cerebral microvasculature, tight junction (TJ) proteins,brain microvascular endothelial cells (BMECs), cellular trans-port pathways and enzymatic machinery. Tight junction pro-teins in particular play a key role in restricting paracellularpermeability (Vorbrodt and Dobrogowska, 2004).
As the most apical structure within the intercellular cleft,TJ limit the paracellular flux of hydrophilic molecules across
[email protected] (L. Xu).ork.

14 B R A I N R E S E A R C H 1 4 3 6 ( 2 0 1 2 ) 1 3 – 1 9
the BBB. TJ and adherens junctions form a circumferentialzipper-like seal between adjacent endothelial cells. Normally,no substances with a molecular weight greater than 180 Dacan gain access through the TJ (Sandoval and Witt, 2008).Three essential transmembrane proteins, including claudins,occludin and junction adhesion molecules (JAMs), determinethe integrity of the TJ. TJ protein expression and regulationare dependent on cellular localization, post-translationalmodification, and interactions between proteins (Vorbrodtand Dobrogowska, 2003). Occludin, a tetraspanning mem-brane protein, is expressed at high concentrations at BBB TJ.Occludin is a phosphoprotein with a molecular weight of60–65 kDa. Occludin has two extracellular loops separated bya short cytosolic loop with both NH2- and COOH-terminal do-mains being cytosolic. The COOH-terminal domain of occlu-din binds to zonula occludens proteins (ZO-1, ZO-2, and ZO-3), which in turn bind to the actin cytoskeleton, localizingoccludin mostly to the cellular membrane. Tyrosine phos-phorylation of occludin has been shown to occur with its dis-association from intracellular proteins and to result inincreased TJ permeability (Kirschner et al., 2010).
HIV-1 can cause neuroAIDS, which involves multiple com-plicated processes. HIV-1-infected cells in both the peripheryand the CNS produce viral proteins, including gp120, Tat andNef, as well as host inflammatory mediators such as cyto-kines and chemokines (Strazza et al., 2011). Disruption oftight junctions is commonly observed in HIV-1-infected pa-tients (Dallasta et al., 1999). The HIV-1 virus and its proteinsmay act alone or in cooperation with host cytokines and che-mokines to affect the integrity of the BBB (Mahajan et al.,2008). Fragmentation and decreased immunoreactivity ofoccludin and ZO-1 were observed after HIV infection; in addi-tion, HIV-1 Tat induced alterations in claudin-5 (Dallasta et al.,1999; Andras and Toborek, 2011).
Tat protein vastly increases the level of HIV dsRNA tran-scription (Naryshkin et al., 1998). Tat is a strong pro-inflammatory agent that can recruit monocytes and inducetheir transendothelial migration (Albini et al., 1998). Severalof the vascular effects caused by HIV-1 and observed in theCNS can be reproduced by treatment with Tat. Tat has beenfound to affect endothelial cell actin microfilament dynamics,causing actin cytoskeletal rearrangements and disassembly(Wu et al., 2004). Meanwhile, Tat canmodulate the permeabilityof the blood–brain barrier (Banks et al., 2005; Banerjee et al.,2010; Gandhi et al., 2010). However, the mechanisms by whichthe alteration occurs have not been characterized in detail. Inthis study, we found that Tat protein disrupts occludin by bothinhibiting its expression and cleaving it via matrix metallopro-teinase (MMP)-9 in brain endothelial cells; this finding mayhelp us understand how HIV-1 factors act during neuroAIDS.
2. Results
2.1. Tat treatment decreases the levels of occludin mRNAand protein
There is controversy in the literature regarding the effect ofTat on occludin. In our study, recombinant Tat reduced thelevels of occludin gene transcription and protein expression.
Occludin mRNA and total protein levels were determined byreal-time RT-PCR and Western blot, respectively. Exposure ofcells to Tat resulted in a decrease in their occludin mRNAlevels after 30 min-24 h (Fig. 1A). Total occludin protein levelsdid not change within the first 6 h of Tat treatment andchanged only after 12–24 h (Fig. 1B). Using fluorescence mi-croscopy, alterations in occludin immunoreactivity werealso observed. For the untreated group, occludin stainingwas strong and uniform. When cells were treated with Tatfor 24 h, occludin immunoreactivity became weaker(Fig. 1C).
2.2. The RhoA/ROCK pathway is involved in the regulationof occludin by Tat
A previous study proved that Tat upregulated the expressionof P-glycoprotein via Rho-mediated signaling (Zhong et al.,2010). We wanted to explore whether the RhoA/ROCK signalingcascade is involved in the downregulation of occludin by Tat.Traditionally, RhoA activity has been assessed by a RhoAGTPase pull-down assay. Here, we applied the RhoA inhibitorC3 transferase and the ROCK inhibitor Y-27632 to evaluatetheir effects. The inhibitors were added to the culture system1 h before Tat treatment. We found that they partially blockedthe downregulation of occludin at the mRNA level after 60minof Tat exposure when compared with the Tat-treated group(P<0.05, Fig. 2A). Because exposure of cells to Tat could decreasethe cells’ total occludin levels after 12 h, we chose to examinethe effect of RhoA/ROCK inhibition on occludin protein expres-sion. The resultswere similar to those for occludinmRNA levelswith both inhibitors, the occludin protein levels were signifi-cantly different from that in the Tat-treated group (P<0.05,Fig. 2B).
2.3. Tat treatment increases MMP-9 expression and activity
Recent studies have indicated that matrix metalloprotei-nases (MMPs), especially MMP-9, play a critical role inblood–brain barrier (BBB) disruption. Tat could induce MMP-9 expression in several cell types including monocytes,breast cancer cells and astrocytes (Kumar et al., 1999; Leeet al., 2005; Ju et al., 2009). To confirm whether Tat could in-duce MMP-9 production in HBMEC, we treated the cells withTat and found MMP-9 mRNA levels to be dramatically higher(10 times, P<0.01) at 60 min when compared with untreatedcells; subsequently, the levels declined gradually (Fig. 3A).As further studies using semi-quantitative gelatin zymogra-phy showed, MMP-9 was not present at detectable levels inan untreated control, but could be induced by Tat treatment(Fig. 3B).
2.4. SiRNA targeting MMP-9 reduces the concentration ofsoluble occludin
To confirmwhetherMMP-9 coulddirectly disassemble occludin,we explored the relationship between MMP-9 expression andthe concentration of soluble occludin. In the untreated group,the concentration of soluble occludin was below the detec-tion range of the purchased sandwich ELISA kit. After Tattreatment of cells for 48 h, the concentration of soluble

Fig. 2 – Role of the RhoA/ROCK signaling pathway in theregulation of occludin by Tat. Inhibitors targeting RhoA orROCK were applied, and occludin mRNA (A) and protein (B)were detected by real-time RT-PCR and Western blot,respectively. The data are expressed as the ratio of occludinto β-actin mRNA and the ratio of occludin to the β-actinprotein band density, respectively. *P<0.05 and **P<0.01compared to the controls.
Fig. 1 – Alterations in occludin mRNA and protein levelsinduced by recombinant HIV-1 Tat clade B protein. Isolatedhuman brain microvascular endothelial cells were treatedwith recombinantHIV-1Tat protein (200 ng/ml) for the indicatedtime. (A) Occludin mRNA was detected by real-time RT-PCR.OccludinmRNA levels are expressed as the ratio of occludin toβ-actin mRNA. (B) Occludin protein was detected byWesternblot. Occludin protein levels are expressed as the ratio ofoccludin to β-actin protein band density. (C) Intercellularoccludin distributionwas observed by fluorescencemicroscopy(400×). *P<0.05 compared to the controls.
15B R A I N R E S E A R C H 1 4 3 6 ( 2 0 1 2 ) 1 3 – 1 9
occludin reached approximately 400 pg/ml. To determinewhether MMP-9 was responsible for the production of solubleoccludin, we transfected HBMEC with 50 nM MMP-9 siRNA.Transfection caused a dramatic reduction in soluble occludinlevels of up to 73% (Fig. 4A). In addition, the permeability ofHBMEC monolayers incubated with Tat for 48 h was signifi-cantly higher than their basal permeability. MMP-9 silencingpartially reduced the increase in permeability resulting fromTat treatment (Fig. 4B).
3. Discussion
Since the last century, the acquired immunodeficiency syn-drome (HIV/AIDS) epidemic has been a major global publichealth threat. During the late stages of AIDS, HIV-1–infectedpatients commonly suffer from a wide range of neurologicaldisorders collectively known as HIV-1-associated neurocogni-tive disorders (HAND). HAND is caused by virus particlesthemselves and secreted viral components. In HAND, the in-tegrity of the blood–brain barrier (BBB) is compromised, andits permeability is increased. HIV-1 proteins including Tatand gp120 are known to play vital roles in causing BBB dys-function. Previous studies have proven that HIV-1 proteinscan increase endothelial permeability by modulating the ex-pression and distribution of different tight junction proteins.Although HIV-1 clade C has spread throughout the UnitedStates and other western countries, previous studies havesuggested that clade C is less neuropathogenic than clade
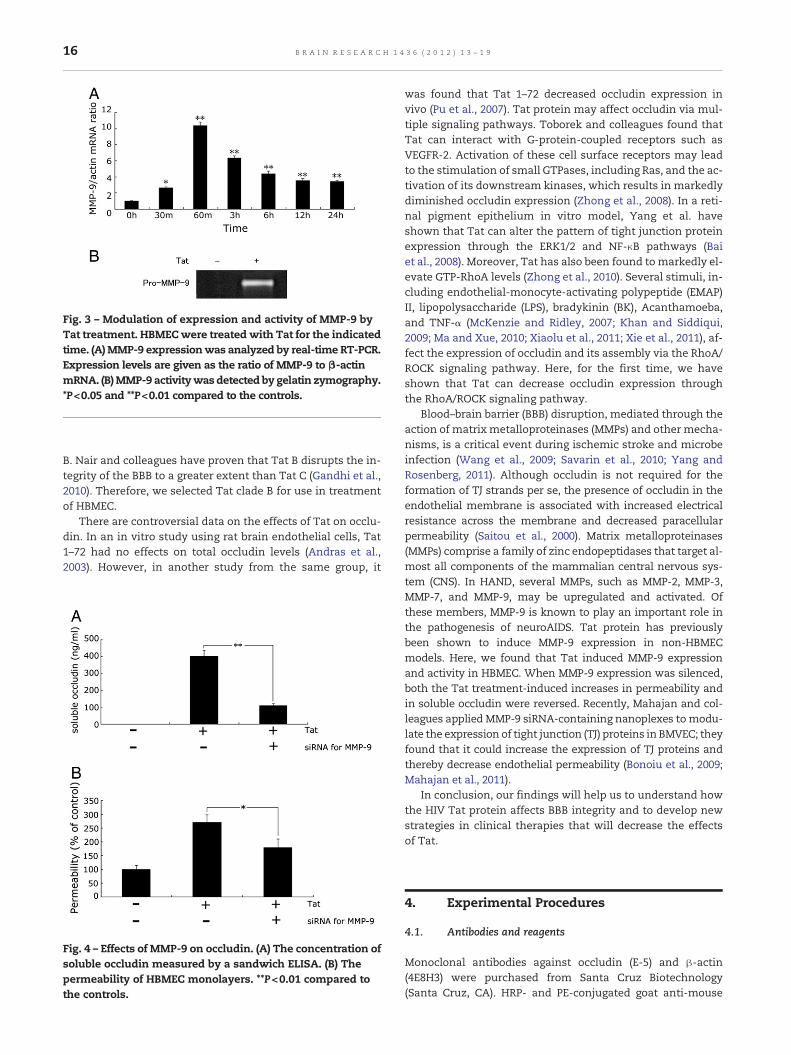
Fig. 3 – Modulation of expression and activity of MMP-9 byTat treatment. HBMECwere treated with Tat for the indicatedtime. (A)MMP-9 expressionwas analyzed by real-time RT-PCR.Expression levels are given as the ratio of MMP-9 to β-actinmRNA. (B)MMP-9 activitywas detected by gelatin zymography.*P<0.05 and **P<0.01 compared to the controls.
16 B R A I N R E S E A R C H 1 4 3 6 ( 2 0 1 2 ) 1 3 – 1 9
B. Nair and colleagues have proven that Tat B disrupts the in-tegrity of the BBB to a greater extent than Tat C (Gandhi et al.,2010). Therefore, we selected Tat clade B for use in treatmentof HBMEC.
There are controversial data on the effects of Tat on occlu-din. In an in vitro study using rat brain endothelial cells, Tat1–72 had no effects on total occludin levels (Andras et al.,2003). However, in another study from the same group, it
Fig. 4 – Effects of MMP-9 on occludin. (A) The concentration ofsoluble occludin measured by a sandwich ELISA. (B) Thepermeability of HBMEC monolayers. **P<0.01 compared tothe controls.
was found that Tat 1–72 decreased occludin expression invivo (Pu et al., 2007). Tat protein may affect occludin via mul-tiple signaling pathways. Toborek and colleagues found thatTat can interact with G-protein-coupled receptors such asVEGFR-2. Activation of these cell surface receptors may leadto the stimulation of small GTPases, including Ras, and the ac-tivation of its downstream kinases, which results in markedlydiminished occludin expression (Zhong et al., 2008). In a reti-nal pigment epithelium in vitro model, Yang et al. haveshown that Tat can alter the pattern of tight junction proteinexpression through the ERK1/2 and NF-κB pathways (Baiet al., 2008). Moreover, Tat has also been found tomarkedly el-evate GTP-RhoA levels (Zhong et al., 2010). Several stimuli, in-cluding endothelial-monocyte-activating polypeptide (EMAP)II, lipopolysaccharide (LPS), bradykinin (BK), Acanthamoeba,and TNF-α (McKenzie and Ridley, 2007; Khan and Siddiqui,2009; Ma and Xue, 2010; Xiaolu et al., 2011; Xie et al., 2011), af-fect the expression of occludin and its assembly via the RhoA/ROCK signaling pathway. Here, for the first time, we haveshown that Tat can decrease occludin expression throughthe RhoA/ROCK signaling pathway.
Blood–brain barrier (BBB) disruption, mediated through theaction of matrix metalloproteinases (MMPs) and other mecha-nisms, is a critical event during ischemic stroke and microbeinfection (Wang et al., 2009; Savarin et al., 2010; Yang andRosenberg, 2011). Although occludin is not required for theformation of TJ strands per se, the presence of occludin in theendothelial membrane is associated with increased electricalresistance across the membrane and decreased paracellularpermeability (Saitou et al., 2000). Matrix metalloproteinases(MMPs) comprise a family of zinc endopeptidases that target al-most all components of the mammalian central nervous sys-tem (CNS). In HAND, several MMPs, such as MMP-2, MMP-3,MMP-7, and MMP-9, may be upregulated and activated. Ofthese members, MMP-9 is known to play an important role inthe pathogenesis of neuroAIDS. Tat protein has previouslybeen shown to induce MMP-9 expression in non-HBMECmodels. Here, we found that Tat induced MMP-9 expressionand activity in HBMEC. When MMP-9 expression was silenced,both the Tat treatment-induced increases in permeability andin soluble occludin were reversed. Recently, Mahajan and col-leagues applied MMP-9 siRNA-containing nanoplexes tomodu-late the expression of tight junction (TJ) proteins in BMVEC; theyfound that it could increase the expression of TJ proteins andthereby decrease endothelial permeability (Bonoiu et al., 2009;Mahajan et al., 2011).
In conclusion, our findings will help us to understand howthe HIV Tat protein affects BBB integrity and to develop newstrategies in clinical therapies that will decrease the effectsof Tat.
4. Experimental Procedures
4.1. Antibodies and reagents
Monoclonal antibodies against occludin (E-5) and β-actin(4E8H3) were purchased from Santa Cruz Biotechnology(Santa Cruz, CA). HRP- and PE-conjugated goat anti-mouse

17B R A I N R E S E A R C H 1 4 3 6 ( 2 0 1 2 ) 1 3 – 1 9
secondary antibodies were from Invitrogen (Carlsbad, CA).Recombinant HIV-1 Tat clade B protein was purchasedfrom Prospec (Rehovort, Israel). The RhoA inhibitor C3 exo-enzyme from Clostridium botulinum (C3 transferase) and theROCK inhibitor Y-27632 were from Calbiochem (La Jolla,CA). FITC-dextran (Mr 42,000) was from Sigma-Aldrich (St.Louis, MO).
4.2. Cell culture and treatment
Human brain microvascular endothelial cells (HBMEC) wereisolated according to Weksler's method (Weksler et al., 2005).Briefly, HBMEC were isolated from human brain tissues,which were obtained at Xijing Hospital by surgical excisionof an area of the temporal lobe from adult patients with epi-lepsy, in accordance with the research governance guidelinesof the Local Ethics Committee. HBMEC were cultured in Endo-thelial Growth Medium-2 (EGM-2) (Lonza, Walkersville, MD)supplemented with EGM-2 SingleQuots (Lonza) and 2% FBS(HyClone, Logan, UT). Cells were seeded onto 1 mg/ml colla-gen type I-coated (Organogenesis, Canton, Mass) flasks,glass coverslips or filters at a density of approximately5,000 cells/cm2. In all experiments, cells were starved in 1%FCS in EGM-2 basal medium without EGM-2 SingleQuots for2 h.
Tat was used in these studies at a concentration of 200 ng/ml according to Buch's method (Bethel-Brown et al., 2011). AnMTT assay showed that the HIV Tat protein at 200 ng/ml hadno effect on cell viability. The ROCK inhibitor Y-27632 andRhoA inhibitor C3 transferase were used at 5 μmol/L, and15 μg/ml concentrations, respectively.
4.3. Real-time reverse transcription-PCR (RT-PCR)
Real-time RT-PCR was applied as in our previous report, withsome modifications (Feng et al., 2010). Briefly, total RNA wasextracted with TRIzol reagent (Invitrogen), purified with theQiagen RNeasy kit according to the manufacturer's instruc-tions, and treated with DNase (Qiagen). The levels of mRNAtranscripts were analyzed by one-step quantitative reversetranscriptase–polymerase chain reaction (qRT-PCR) using spe-cific primers, along with the SYBR® Green PCRMaster Mix andRT-PCR kit according to the manufacturer's instructions, onan ABI Prism 7700 Sequence Detection System (Applied Bio-systems). The sequences of the primers were as follows:5'-ctccatcctggcctcgctgt-3' and 5'-gctgtcaccttcaccgttcc-3' forβ-actin; 5'-acaagtccaatattttgtgg-3' and 5'-tattgatctcatcaagtt-3'for human occludin (NM_002538.2); and 5'-cgctaccacctc-gaactttg-3' and 5'-gccattcacgtcgtccttat-3' for human MMP-9(NM_004994.2). Reactions of 20 μl containing 50 ng of totalRNA, 10 μl 2x SYBR Green PCR Master Mix, 6.25 U Multi-Scribe reverse transcriptase, 10 U RNase inhibitor and0.1 mM primers were subjected to one cycle of 95 °C for10 min and then 40 cycles of 95 °C for 15 s, 56 °C for 30 s and72 °C for 45 s. For relative quantification, the levels of individ-ual mRNA transcripts were first normalized to the level of thecontrol β-actin mRNA. The differential expression of thesegenes was then analyzed by the ΔCt method and expressedas fold change. The amplified products were analyzed by1.5% agarose gel electrophoresis.
4.4. Western blot
Occludin expression was detected by Western blot accordingto Reyes's method (Gonzalez-Mariscal et al., 2000). The har-vested cells were lysed with 1% NP-40 lysis buffer (1% NP-40,25 mM Hepes at pH 7.5, 150 mmol/L NaCl, 1.5 mmol/L EDTA,1 mmol/L PMSF, 10 μg/ml leupeptin, 10 μg/ml aprotonin,1 mmol/L sodium orthovanadate and 25 mmol/L NaF) andtheir protein concentrations were determined by BCA assay(Pierce). The protein lysates (40 μg/lane) were fractionated on8% SDS–polyacrylamide gels (SDS–PAGE), electrophoreticallytransferred (Trans-Blot-Cell; Bio-Rad Laboratories, Richmond,CA) to nitrocellulose membranes, and subjected to Westernblot analysis. Non-specific protein binding was blocked with0.5% (w/v) non-fat dry milk in PBS/0.05% Tween-20 (PBS-T).Blots were incubated with an antibody to occludin (1:1,000)overnight at 4 °C and then incubated with HRP-conjugatedanti-mouse antibodies at room temperature for 1 h. Thebound antibodies were visualized using LumiGLo reagent(Pierce), and the level of each protein relative to β-actin wasdetermined.
4.5. Immunofluorescence staining
Occludin staining was performed as previously described(McKenzie and Ridley, 2007). Cells were pre-extracted in 0.2%pre-extraction buffer (0.2% Triton X-100, 100 mmol/L KCl,3 mmol/L MgCl2, 1 mmol/L CaCl2, 300 mmol/L sucrose, and10 mmol/L Hepes) for 2 min and then fixed in 95% ethanolfor 30 min at 4 °C, treated with 100% ice-cold acetone for1 min, air dried and re-hydrated in PBS. Cells were blocked in3% BSA (in PBS) for 1 h, incubated with the primary antibodyin 3% BSA overnight at 4 °C and then incubated with secondaryantibody for 1 h at room temperature. Images were collectedwith a fluorescence microscope.
4.6. Silencing of MMP-9 with small interfering RNA (siRNA)
An siRNA specific for MMP-9 and a Scramble II control weredesigned against the reported MMP-9 sequence. The se-quences were 5'-UACCUGUACCGCUAUGGUU-3' (19 nt) forthe MMP-9 siRNA and 5'-UUGGUAUCGCCAUGUCCAU-3' (21 nt)for the Scramble II control (Turner et al., 2007). HBMEC weretransfected with 50 nM of the MMP-9 siRNA or Scramble IIsiRNA using GeneSilencer siRNA transfection reagent (Gen-eTherapy System) according to themanufacturer's instructions.
4.7. Gelatin zymography
The activity of MMP-9 was tested by gelatin zymography.Zymography was performed on each sample with nonredu-cing electrophoresis through a 10% sodium dodecylsulfate–polyacrylamide gel containing 1 mg/mL gelatin. As a positivecontrol, conditioned medium from a human fibrosarcomacell line (HT-1080) was also loaded onto each gel. After separa-tion was completed, sodium dodecylsulfate was removedfrom the gels by incubation in 2.5% Triton X-100 (3×15 min)before immersion for 24 h in incubation buffer (50 mmol/LTris–HCl, 10 mmol/L CaCl2, and 0.05% Brij-35; pH 7.4) at 37 °C.Gels were then fixed and stained for 2–3 h in 0.1% Coomassie

18 B R A I N R E S E A R C H 1 4 3 6 ( 2 0 1 2 ) 1 3 – 1 9
brilliant blue in amediumconsisting of 50%methanol, 20%aceticacid and 30% double-distilled water. MMP-9 was visualized asclear bands of lysis against a darkbluebackgroundof intact sub-strate. The molecular weight of each band was compared withknown molecular weight standards (Takara, Dalian, China).The relative densities of gelatinolytic bands were determinedfrom negative scanned images of gels using ImageQuant soft-ware (Amersham).
4.8. Transendothelial permeability assays
HBMEC were grown on 0.4-μm pore Transwell filters (Nunc,Roskilde, Denmark) until confluent. After the cells werestarved for 2 h, FITC-dextran was applied apically at 0.1 μg/ml and allowed to equilibrate for 30 min. Next, a sample ofthe medium in the lower chamber was collected to measurethe basal permeability. Monolayers were then either leftuntreated (control) or were stimulated in triplicate with re-combinant HIV-1 Tat. Samples were removed from the lowerchamber for fluorescence measurements and compared tocontrol monolayers. Fluorescence was measured using aPerkin-Elmer Fusion Universal Microplate Analyzer with anexcitation wavelength of 492 nm and detection wavelengthof 520 nm.
4.9. ELISA
Levels of soluble occludin were assessed using the appropriateELISA kits (Uscn Life Science) according to the manufacturer'sinstructions. Final results were read at a wavelength of 450 nm.
4.10. Statistics
Results are expressed as the mean±S.E.M. with n representingthenumber of experiments. To compare values amongmultiplegroups, one-way analysis of variance (ANOVA) was applied. Forcomparison of the mean values between two groups, the un-paired t-testwasused.Differenceswere acceptedas statisticallysignificant at P<0.05.
Acknowledgments
This work was supported by the National Natural ScienceFund (No. 81000597 & 81070248), the Critical Project Founda-tion on Social Development of ShaanXi province(No.2010 K16-04-05) and Natural Science Fund of ShaanXiprovince (No. 2009JM4023).
R E F E R E N C E S
Albini, A., et al., 1998. HIV-1 Tat protein mimicry of chemokines.Proc. Natl. Acad. Sci. U. S. A. 95, 13153–13158.
Andras, I.E., et al., 2003. HIV-1 Tat protein alters tight junctionprotein expression and distribution in cultured brainendothelial cells. J. Neurosci. Res. 74, 255–265.
Andras, I.E., Toborek, M., 2011. HIV-1-Induced Alterations ofClaudin-5 Expression at the Blood–brain Barrier Level. MethodsMol. Biol. 762, 355–370.
Bai, L., et al., 2008. HIV-1 Tat protein alter the tight junctionintegrity and function of retinal pigment epithelium: an invitro study. BMC Infect. Dis. 8, 77.
Banerjee, A., et al., 2010. HIV proteins (gp120 and Tat) andmethamphetamine in oxidative stress-induced damage in thebrain: potential role of the thiol antioxidant N-acetylcysteineamide. Free Radic. Biol. Med. 48, 1388–1398.
Banks, W.A., et al., 2005. Permeability of the blood–brain barrier toHIV-1 Tat. Exp. Neurol. 193, 218–227.
Bethel-Brown, C., et al., 2011. HIV-1 Tat-mediated induction ofplatelet-derived growth factor in astrocytes: role of earlygrowth response gene 1. J. Immunol. 186, 4119–4129.
Bonoiu, A., et al., 2009. MMP-9 gene silencing by a quantumdot-siRNA nanoplex delivery to maintain the integrity of theblood brain barrier. Brain Res. 1282, 142–155.
Correale, J., Villa, A., 2009. Cellular elements of the blood–brainbarrier. Neurochem. Res. 34, 2067–2077.
Dallasta, L.M., et al., 1999. Blood–brain barrier tight junctiondisruption in human immunodeficiency virus-1 encephalitis.Am. J. Pathol. 155, 1915–1927.
Feng, X., et al., 2010. Lipopolysaccharideup-regulates the expressionof Fcalpha/mu receptor and promotes the binding of oxidizedlow-density lipoprotein and its IgM antibody complex toactivated human macrophages. Atherosclerosis 208, 396–405.
Gandhi, N., et al., 2010. Interactive role of humanimmunodeficiency virus type 1 (HIV-1) clade-specific Tat proteinand cocaine in blood–brain barrier dysfunction: implications forHIV-1-associated neurocognitive disorder. J. Neurovirol. 16,294–305.
Gonzalez-Mariscal, L., et al., 2000. Tight junction proteins ZO-1,ZO-2, and occludin along isolated renal tubules. Kidney Int. 57,2386–2402.
Ju, S.M., et al., 2009. Extracellular HIV-1 Tat up-regulates expressionofmatrixmetalloproteinase-9 via aMAPK-NF-kappaBdependentpathway in human astrocytes. Exp. Mol. Med. 41, 86–93.
Khan, N.A., Siddiqui, R., 2009. Acanthamoeba affects the integrityof human brain microvascular endothelial cells and degradesthe tight junction proteins. Int. J. Parasitol. 39, 1611–1616.
Kirschner, N., et al., 2010. Tight junctions: is there a role indermatology? Arch. Dermatol. Res. 302, 483–493.
Kumar, A., et al., 1999. Human immunodeficiency virus-1-tatinduces matrix metalloproteinase-9 in monocytes throughprotein tyrosine phosphatase-mediated activation of nucleartranscription factor NF-kappaB. FEBS Lett. 462, 140–144.
Lee, Y.W., et al., 2005. Human immunodeficiency virus-1 Tatprotein up-regulates interleukin-6 and interleukin-8expression in human breast cancer cells. Inflamm. Res. 54,380–389.
Ma, T., Xue, Y., 2010. RhoA-mediated potential regulation ofblood-tumor barrier permeability by bradykinin. J. Mol.Neurosci. 42, 67–73.
Mahajan, S.D., et al., 2011. Suppression of MMP-9 Expression inBrain Microvascular Endothelial Cells (BMVEC) Using a GoldNanorod (GNR)-siRNA Nanoplex. Immunol. Invest.
Mahajan, S.D., et al., 2008. Tight junction regulation by morphineand HIV-1 tat modulates blood–brain barrier permeability. J.Clin. Immunol. 28, 528–541.
McKenzie, J.A., Ridley, A.J., 2007. Roles of Rho/ROCK and MLCK inTNF-alpha-induced changes in endothelial morphology andpermeability. J. Cell. Physiol. 213, 221–228.
Nag, S., et al., 2011. Review: molecular pathogenesis of blood–brainbarrier breakdown in acute brain injury. Neuropathol. Appl.Neurobiol. 37, 3–23.
Naryshkin, N.A., et al., 1998. RNA recognition and regulation ofHIV-1 gene expression by viral factor Tat. Biochemistry (Mosc.)63, 489–503.
Pu, H., et al., 2007. Limited role of COX-2 in HIV Tat-inducedalterations of tight junction protein expression and disruption ofthe blood–brain barrier. Brain Res. 1184, 333–344.

19B R A I N R E S E A R C H 1 4 3 6 ( 2 0 1 2 ) 1 3 – 1 9
Saitou, M., et al., 2000. Complex phenotype of mice lackingoccludin, a component of tight junction strands. Mol. Biol. Cell11, 4131–4142.
Sandoval, K.E., Witt, K.A., 2008. Blood–brain barrier tight junctionpermeability and ischemic stroke. Neurobiol. Dis. 32, 200–219.
Savarin, C., et al., 2010. Monocytes regulate T cell migrationthrough the glia limitans during acute viral encephalitis. J.Virol. 84, 4878–4888.
Strazza, M., et al., 2011. Breaking down the barrier: The effects ofHIV-1 on the blood–brain barrier. Brain Res. 1399, 96–115.
Turner, N.A., et al., 2007. Selective gene silencing of either MMP-2or MMP-9 inhibits invasion of human saphenous vein smoothmuscle cells. Atherosclerosis 193, 36–43.
Vorbrodt, A.W., Dobrogowska, D.H., 2003. Molecular anatomy ofintercellular junctions in brain endothelial and epithelialbarriers: electron microscopist's view. Brain Res. Brain Res.Rev. 42, 221–242.
Vorbrodt, A.W., Dobrogowska, D.H., 2004. Molecular anatomy ofinterendothelial junctions in human blood–brain barriermicrovessels. Folia Histochem. Cytobiol. 42, 67–75.
Wang, G., et al., 2009. Bone marrow-derived cells are the majorsource of MMP-9 contributing to blood–brain barrier dysfunctionand infarct formation after ischemic stroke in mice. Brain Res.1294, 183–192.
Weksler, B.B., et al., 2005. Blood–brain barrier-specific propertiesof a human adult brain endothelial cell line. FASEB J. 19,1872–1874.
Wu, R.F., et al., 2004. Human immunodeficiency virus type 1 Tatregulates endothelial cell actin cytoskeletal dynamics throughPAK1 activation and oxidant production. J. Virol. 78, 779–789.
Xiaolu, D., et al., 2011. Role of p115RhoGEF inlipopolysaccharide-induced mouse brain microvascularendothelial barrier dysfunction. Brain Res. 1387, 1–7.
Xie, H., et al., 2011. Role of RhoA/ROCK Signaling inEndothelial-Monocyte-Activating Polypeptide II Opening ofthe Blood-Tumor Barrier : Role of RhoA/ROCK Signaling inEMAP II Opening of the BTB. J. Mol. Neurosci.
Yang, Y., Rosenberg, G.A., 2011. MMP-Mediated Disruption ofClaudin-5 in the Blood–brain Barrier of Rat Brain After CerebralIschemia. Methods Mol. Biol. 762, 333–345.
Zhong, Y., et al., 2010. Intact lipid rafts regulate HIV-1 Tatprotein-induced activation of the Rho signaling and upregulationof P-glycoprotein in brain endothelial cells. J. Cereb. Blood FlowMetab. 30, 522–533.
Zhong, Y., et al., 2008. Caveolin-1 regulates humanimmunodeficiency virus-1 Tat-induced alterations of tightjunction protein expression viamodulation of the Ras signaling.J. Neurosci. 28, 7788–7796.