heterotrophic flagellates from coastal marine and hypersaline sediments in western australia
TRANSCRIPT

Europ. J. Protisto!' 32,423-448 (1996)December 31, 1996
European Journal of
PROTISTOLOGY
Heterotrophic flagellates from coastal marineand hypersaline sediments in Western Australia
David J. Patterson and A. G. B. SimpsonSchool ofBiological Sciences, ADa, University ofSydney, NSW2006, Australia
SUMMARY
The communities of heterotrophic flagellates from four sites of differing salinity (frommarine to saturated brine) from the Shark Bay region in Western Australia were examined.A total of 46 species were identified, seven of which are described for the first time. Newnames are Ancyromonas melba n. sp. Bodo cygnus n. sp., Colpodella unguis n. sp., Glissandra innuerende n. gen. n. sp., Pleurostomum turgidum n. sp., Ploeotia azurina n. sp.,Rhynchobodo simius n. sp. and Palustrimonas n. gen. The majority of the new specieswere from hypersaline sites. There was a degree of overlap in the communities presentat the different sites, especially between the three lowest salinity sites. Some species, suchas Rhynchomonas nasuta, Bodo saltans and B. designis, have been found over a widerange of salinities. There was little evidence for any endemism in the marine site, withall species observed having been recorded previously from at least one other location geographically remote from Shark Bay.
Introduction Material and Methods
Heterotrophic flagellates form an important and diverse component of the communities living in marinesediments [1, 2, 8, 11, 37]. As a contribution to thestudy of marine heterotrophic flagellates, we havesought to understand their geographical patchiness[8, 22, 38, 54-57]. This work includes an attemptto record the marine heterotrophic flagellate communities from several sites around a continent (Australia). The ultimate aim is to assess whether thesecommunities show any evidence of endemism. This paper, which examines the benthic marine flagellatesfrom sites in Shark Bay, Western Australia, complements both a parallel study of water-column flagellates from the same location [54] and an earlierstudy of benthic marine communities from tropicaleastern Australia [8]. In addition to providing information on the geographical distribution of species, thecommunities from several adjacent sites of differingsalinities are compared.
© 1996 by Gustav Fischer Verlag, Stuttgart
This study was conducted at several sites in the region ofShark Bay, Western Australia. The sites studied were: LittleLagoon (Denham) (25°54' S, 113°32' E); within the vicinityof the stromatolites at the southern end of Hamelin Pool(26°24' S, 113°43' E); and from a land-locked lagoon systemwhich included saturated salt ponds in which salt crystals haddeposited on the floor and around the margins (in the regionof the Taillefer Isthmus, 25°11' S, 113°43' E). In the region ofLittle Lagoon (which is permenantly contiguous with SharkBay itself), the sea has a salinity of about 41roo [24]. HamelinPool has a salinity of between 60roo and 65%0 [24]. Sampleswere taken from two sites in the land-locked lagoon. One sitehad a salinity of about 150%0 (our measurement made using arefractometer) and is referred to as the hypersaline pond. Theother site was a small depression thickly encrusted with saltoverlain with a few centimetres of saturated brine, and is referred to as the saturated puddle.
Samples of sandy sediments were taken from regions withshallow waters (intertidal or with < 10 cm of overlyingwater). Only the oxygenated (non grey/non-black) layerwas taken using a flat scoop. Water was taken from each siteas wel!. Sediment with water was transferred to trays suchthat the sediment settled to form an even layer about 1 cm
0932-4739-96-0032-0423$3.50-0

424 . D. J. Patterson and A. G. B. Simpson
thick. After an hour or more settling time the overlying waterwas removed and a sheet of lens cleaning paper was placed onthe surface of the sediment. No.1 coverslips were carefullyplaced on the lens cleaning paper. Lids were placed on thetrays to reduce evaporation. Over the next four days coverslips were removed from the trays, placed on slides and theliving communities associated with them were examinedusing a Zeiss Axioplan or Zeiss Standard microscope fittedwith phase contrast and DIC optics. No material was fixedor stained. Both microscopes were configured for photomicrography with Olympus OM-2N cameras linked with Olympus T32 flashgun tubes, which were positioned in the lightpaths [32]. The Zeiss Axioplan microscope was also fittedwith video facilities including a Sony video graphic printer.
Data were tabulated for subsequent analysis by PRIMER[see 8].
Results
Kinetoplastida Honigberg, 1963
Bodo cygnus Patterson and Simpson n. sp. Figs. 1 ac, 2a, 2 b
Diagnosis: Elliptical body 8-12 11m long, two heterodynamic flagella, inserting into a subapical pocket, short rostrum with ingestion apparatus, movingby swimming and skidding, body with spiral groove.Type micrograph: Fig. 1 a.
Description: Flagellate with an elliptical body measuring 8-12 11m long. The anterior flagellum is aboutthe same length as the body, moves with a sweepingmotion when the cell is swimming and may wraparound the anterior end of the cell. The posterior flagellum is about twice the length of the body and is acronematic. Cell moves by swimming or skidding, andthe front end of the cell describes a spiral. The flagellainsert in a distinct pocket, and the cell has a spiralgroove that extends from the pocket to the posteriorend of the cell. A fine linear mouth may be seen atthe anterior end of the cell (Fig. 1 c). Observed onlyfrom the hypersaline pond.
Remarks: B. cygnus is very similar to B. designis, theonly feature that reliably separates the species being thespiral groove. Although there is considerable overlap insize, B. cygnus tends to be larger. This species wasfound only in the hypersaline environment, and we observed no cells that could be identified as B. designis inthe hypersaline habitats. There is some similarity between B. cygnus and Rhynchobodo simius, but theycan be distinguished because of the absence of skidding behaviour and more pronounced rostral regionin R. simius. We cannot, at this stage, be absolutely
confident that this species is assignable to Bodo, aswe did not observe a kinetoplast. We have assignedthe species to Bodo because of the large number of features (body shape/size, motility, feeding apparatus, acronematic posterior flagellum, the anterior flagellumwrapping around the body) held in common withB. designis. We note that while this combination of features may be regarded as diagnostic, none of these features alone are exclusive to Bodo.
Bodo designis Skuja, 1948. Fig. 2 c.
Description: length 5-11 11m, mostly 5 - 9 11m. Cellelliptical, usually slightly wider posteriorly, with a subapical indentation from which two unequal flagellaemerge. The cell is plastic. The anterior flagellum isabout the length of the cell or slightly shorter, andpoints forward with the tip curved. The acronematicposterior flagellum is about twice the cell length. Swimming and skidding cells rotate around their longitudinal axis. Only observed from Little Lagoon andHamelin Pool, where it was common in most samples.
Remarks: The cells observed were largely consistentwith those described from other marine habitats [8,22]. Individuals have been identified largely becauseof the rotating motion when they swim. The rotatingmotion is also evident when the cells skid in close proximity to the substrate. Cells have been reported withsizes from 5-15 11m [8]. They may feed with the anterior flagellum wrapped around the anterior end of thecell, a characteristic shared with B. saliens. However, inthis study we report on Rhynchobodo simius n. sp., andon B. cygnus n. sp. which have similar aspects, overlapin size, also swim with a rotating motion, and may alsowrap the anterior flagellum around the front of the cell.R. simius was never observed skidding. When at rest,R. simius may also be distinguished by the more posterior insertion of the flagella, by the more prominent rostral region and the spiral groove. Bodo cygnus n. sp. isdistinguished from B. designis by a spiral groove in thebody surface and has only been observed in hypersalinehabitats. It is possible that some of the references tolarger individuals of B. designis in the literature [e.g.22, 55] might refer to either B. cygnus or R. simius.
Bodo saliens Larsen and Patterson, 1990, Figs. 1 h,1 i, 2 d.
Description: Observed occasionally, length 6-9 11m.Cell body somewhat lanceolate in form and inflexible.Anterior flagellum directed forwards with a singlecurve, posterior flagellum typically directed straight
Fig. 1. All micrographs to the same scale and are DIC images unless otherwise specified. (a)-(c) Bodo cygnus, (a) cell showing ~
flagellar pocket and groove, (b) detail of groove, (c) cell showing groove and ingestion device. (d) Bodo saltans-cell fromHamelin Pool. (e)-(g) Bodo saltans-cells from the saturated puddle, (e) attached cell, (f) swimming cell showing rostrumand short flagella, (g) swimming cell with food vacuoles. (h), (i) Bodo saliens, (j)-(o) Rhynchobodo simius, (j) general appearance of cell-wide aspect, (k) general appearance-narrow aspect, (I) detail of rostrum with ingestion device, (m) large (16 l!m) cellfrom the hypersaline pond, (n) ventral view with detail of flagellar insertion, note acronematic posterior flagellum, (0) same cellas in (n) showing groove detail. (p), (q) Rhynchomonas nasuta, (p) phase contrast. (r)-(t) Dinema validum, (r) general appearance of cell, (s) ventral view showing flagellar lengths, (t) dorsal view. Scale bar (bottom right) represents 10 l!m.
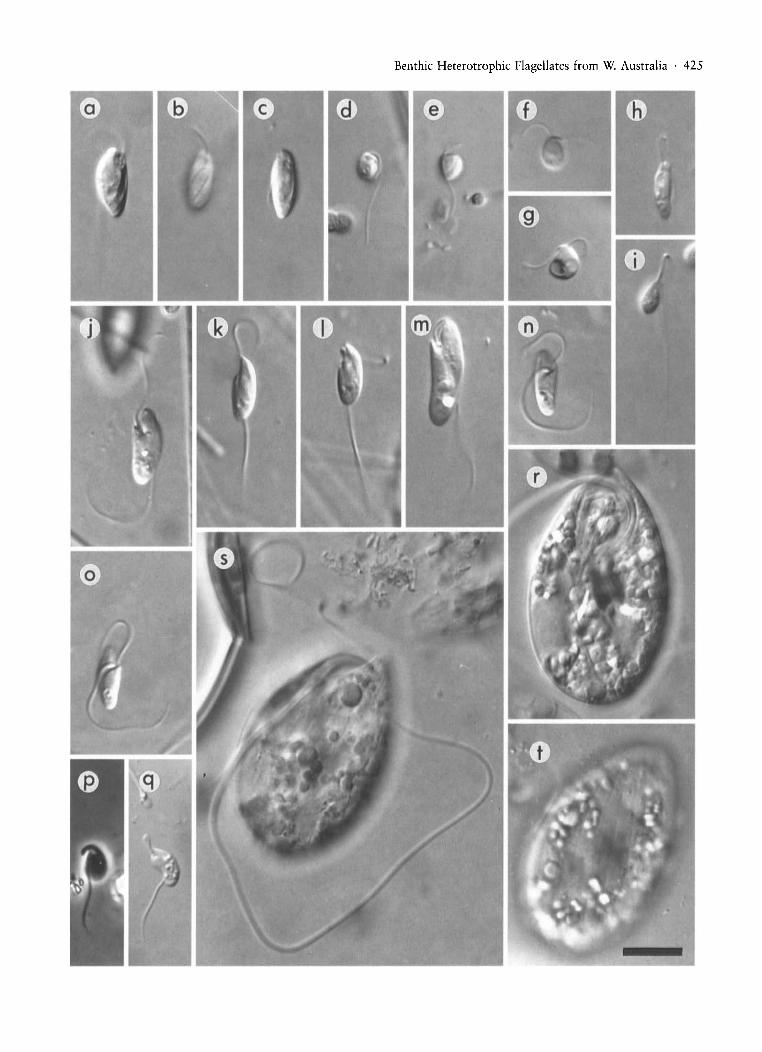
Benthic Heterotrophic Flagellates from W. Australia . 425
ba

426 . D.]. Patterson and A. G. B. Simpson
behind the cell. Swimming motion frequently in rapiddarts in straight lines. Observed from Little Lagoon,Hamelin Pool, and the hypersaline pond.
Remarks: Appearance entirely consistent with descriptions elsewhere in the recent literature [6, 22,55]. B. saliens can be distinguished from all other flagellates by the rapid darting motion with the front flagellum held in a crook (see also B. designis above).
Bodo saltans Ehrenberg, 1832. Figs. 1 d-g, 2 e.
Description: A roundish to bean-shaped flagellate,measuring 4-5 J.lm, was observed from Hamelin Poolon a few occasions (Figs. 1 d, 2 e). Cells typically adhereto the substrate by means of the tip of the long (2-3times the body length) recurrent flagellum. The cellsoccasionally jumped or flicked, presumably becauseof the bending of the recurrent flagellum. The otherflagellum was about the same length as the body butwas inserted anteriorly and curved over the body.The beating of the shorter flagellum drew a currentof water passing anterior to posterior over the cell.The anterior portion of the cell may appear as a laterally directed rostrum. The cell spiralled during swimming, which was rapid.
A flagellate 4-9 J.lm long was the most common colourless protist in the samples from the saturated puddle. The cell had two heterodynamic flagella emergingclose together near the anterior end. One flagellum wasabout body length. The other, "recurrent", flagellumwas 1-2.5 times the body length. The extreme anterior region took the form of a short laterally directedrostrum. Large cells usually had posterior food vacuoles which often distended the cell. The flagellatewas most often observed swimming with both flagellabeating. Some cells attached to surfaces by the tip of anon-beating recurrent flagellum. In these attached cellsthe other flagellum beat with an oaring motion andpassed a current across the cell. None of the cells observed from the saturated puddle displayed jumping orflicking behaviour.
Remarks: We regard all these observations as referable to Bodo saltans. This well known species is common in freshwater habitats but has only occasionallybeen observed from marine habitats [36, 54, 55].The jumpinglflicking behaviour observed in the Hamelin pool material is distinctive for B. saltans [3]. Therest of the appearance of the Hamelin pool cells is alsoconsistent with Hanel's [16] concept of the species.
Once allowances for the viscosity of the media weremade, the appearance of most cells from the saturatedpuddle was consistent with Hanel's [16] concept ofB. saltans in respect of size, form and flagellar characters, but the distinctive jumping/flicking behaviour wasnot observed. We have, however, observed this sameorganism performing the jumping behaviour in samples from an artificial hypersaline lagoon in South Australia (salinity 180-260%0 NaCl). A previousobservation from salt saturated samples of a flagellateunder the name Bodo sp. 1 [40] includes the jumping
behaviour and is regarded by us as B. saltans. Namyslowski [30] inadequately described several halophilicspecies of Amphimonas, most or all of which are probably the organism that we observed from the saturatedpuddle.
Rhynchobodo simius Patterson and Simpson n. sp.Figs. 1 j-o, 2 g, h.
Diagnosis: Rhynchobodo sp. 9-16 J.lm long withprominent, immovable rostrum a quarter or more ofthe length of the cell. Flagella longer than the cell withposterior flagellum longer than the anterior one. Cellnot noticeably deformable. With groove extendingfrom the flagellar pocket to the posterior of the cell.Type micrograph: Fig. 1 j.
Description: Elliptical slightly compressed cell measuring 9- 16 J.lm long (all cells except one were 912 J.lm long - see below), with two flagella insertingsubapically, at the head of a distinct spiral groove inthe cell body. Anterior flagellum 1-1112 times celllength, posterior (recurrent) flagellum 1V2-2V2 timescell length. The most anterior portion of the recurrentflagellum usually lies in the groove. Flagella appearthick and the recurrent flagellum at least is acronematico The cell body anterior to the flagellar insertionforms a rostrum. The rostrum is a quarter or more ofthe total length of the cell and contains a tubular ingestion apparatus that opens at the tip. Cells move with aspiral swimming motion. Cells were not seen to glide,skid, deform or squirm. When cells swam into contactwith objects, the anterior flagellum frequently wrappedaround the cell and motion was either arrested or thecell spun on its longitudinal axis. The cells measuring9-12 J.lm long were observed from Hamelin Pool. Thecell measuring 16 J.lm was observed from the hypersaline pond (Fig. 1 m).
Remarks: We believe that this species has been described several times under different names from bothfreshwater and marine sites. Phyllomitus apiculatussensu Mylnikov [26, 27] and Steinberg et al. [49]and Cryptaulax vulgaris sensu Novarino et al. [31]both appear to refer to this organism. We believe thatthe organism differs from Skuja's [45] original description of Phyllomitus apiculatus because R. simius doesnot have a deep subapical gullet. The original description of Cryptaulax vulgaris [45] refers to a very metabolic (i.e., highly pliable) protist. The organism alsodiffers from all other nominal species of Cryptaulax[22, 44, 45, 46, 51] by its lack of pliability and/orby the length of the rostrum.
We therefore consider all previous identifications ofthis organism to be incorrect. As none of these studiesintroduced a species name, a novel one is required. Theultrastructure of the organism as described by Mylnikov [27] is virtually identical to that of Rhynchobodoarmata and R. taeniata [4, 55]. The name Rhynchobodo appears to now be legitimate, despite being originally introduced in error by Lackey [20] andmisleading statements made by Burzell [6]. It is un-

clear which of the three previously described nominalspecies is the correct type for the genus and whetherVers' [55] formal amendment was necessary considering Brugerolle's [4] study. We hope to provide a definitive resolution for this problem in a later paper.R. simius differs from R. armata and R. agilis becausethe latter two species do not have a spiral groove [4,20]. R. simius may be distinguished from R. taeniatabecause the latter is a much narrower cell with a shorter, thinner rostrum [44, 55].
R. simius is similar to Bodo cygnus, B. designis andBordnamonas tropicana but may be distinguished fromthem by the longer, anteriorly directed rostrum andlack of skidding type movement. The groove is moreobvious than in Bodo cygnus while B. designis andBordnamonas tropicana lack grooves altogether.
Although we have identified the 16 11m long cell fromthe hypersaline puddle as R. simius, we note that it hasa significantly larger body, shorter rostrum and morespiral groove than all the other cells observed thatwere assignable to the species. This cell shows manysimilarities with Cryptaulax marina sensu Vars [55,57]. Vars' [55, 57] observations do not appear to beof the same organism as in Throndsen's [51] originaldescription of C. marina [Tong, pers. comm.], whichis figured with the flagella inserting about a fifth ofthe way down the cell, with a narrow rostrum anteriorto the flagellar insertion. Vars' [55] micrographs of theorganism she described as C. marina show a broad rostrum and the flagella inserting more than a third of theway down the cell. We are unsure as to where theboundary between 'c. marina sensu V"m [55, 57]'and R. simius lies and whether two species should bedistinguished. Further research is required.
Rhynchomonas nasuta Klebs, 1892, Figs. 1 p, 1 q, 2 f.Description: Gliding cells,S - 7 /lm long, with bul
bous rostral region that moves from side to side andwith a sweeping motion. Body plastic and flattened.Cells may detach from the substrate and move withrapid vibrations. Found at Little Lagoon, HamelinPool and hypersaline pond. Common in all locationswhere it was seen.
Remarks: Observed cells in good agreement withdecriptions elsewhere [8, 22, 50]. Distinguishablefrom all other gliding flagellates by the flexible and bulbous rostrum.
Euglenida, Biitschli 1884
Dinema validum Larsen and Patterson, 1990, Figs.1 r-t, 2 i.
Description: Cell 27-38 11m long. Elliptical, withabout 20 evenly-spaced, almost longitudinal striations. Spherical nucleus located in the posterior partof the cell. Ingestion apparatus visible, rods thick,but easily obscured by other cytoplasmic elements.Anterior flagellum slightly longer than cell lengthand beating with a sweeping motion. Posterior flagellum thick, emerging from the flagellar pocket in a
Benthic Heterotrophic Flagellates from W. Australia . 427
hook, extending almost three times the cell length.Cells gliding and have a capacity to contract. With adiscrete cortex. Food includes diatoms. Observed occasionally from Little Lagoon.
Remarks: In agreement with the original description[22] except that the largest cells observed were somewhat larger than has been previously reported. Theseobservations extend the size range reported for this species to 26-38 11m. This species is distinguished fromothers in the genus by the length and thickness ofthe recurrent flagellum [22].
Dolium sedentarium Larsen and Patterson, 1990,Figs. 2 j, 3 a-c.
Description: Cell length 50-60 11m, width 1825 /lm. Cells rigid, slightly flattened, 6 grooves, twodorsal, two ventral and two marginal. Cell sessile, attached by pointed posterior tip, anterior end somewhatflared, but hollow centrally. Two flagella, one emergentfrom a flagellar pocket through a narrow flagellar canal and usually projecting from cell. May eat diatoms.Cell not observed to swim or glide and appears to besessile. From Little Lagoon.
Remarks: Cell length longer than reported by [22]. Incontrast with Ekebom et al. [8] and in keeping withLarsen and Patterson [22], 6 longitudinal ridges wereseen. Some cells seen from Little Lagoon were moreslender than has been previously observed. Dolium isthe only known sedentary phagotrophic euglenid [22].
The name Dolium Larsen and Patterson is a juniorhomonym of Dolium Linnaeus (a mollusc). A changeis, at this time, required by the rules of the [CZN. Thiswould lead to different names under the Botanical andZoological codes of nomenclature. Consequently, wechoose not to introduce a new name, now holdingthe view that the context of the application of namesof protists that are homonymous with those of plantsor animals renders their meaning unambiguous [36]and thereby promotes stability.
Heteronema exaratum Larsen and Patterson, 1990,Figs. 2 k, 3 k.
Description: One cell observed from Little Lagoon,length 15 Jlm. Cell profile roundish, and flattened, posteriorly acute, body surface with about 20 grooves oneach of the dorsal and ventral surface. Grooves are obliquely angled, and more prominent on dorsal surface.Moves rapidly, swimming near or touching the substrate, front flagellum held in curve in front of cell.The two flagella are of similar width and both slightlylonger than the cell. Able to contract somewhat. Ingestion apparatus well developed.
Remarks: In agreement with original description byLarsen and Patterson [22]. H. exaratum and H. ovaleare the only species in the genus that are oval shapedwith flagella both about body length. H. exaratumcan be distinguished from H. ovale by being smaller,lacking a pointed posterior and lacking strong bandsof pellicular warts [22].
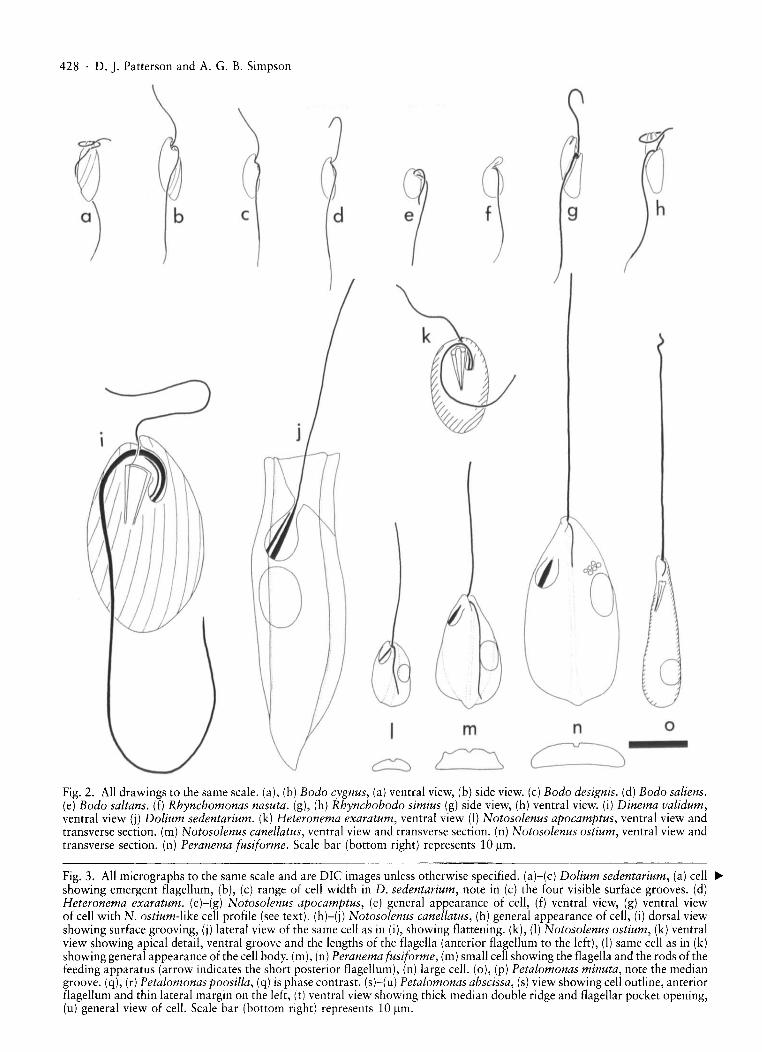
428 . D.]. Patterson and A. G. B. Simpson
c d 9
on
~
m
~c::::::::J
, - I
Fig. 2. All drawings to the same scale. (a), (b) Bodo cygnus, (a) ventral view, (b) side view. (c) Bodo designis. (d) Bodo saliens.(e) Bodo saltans. (f) Rhynchomonas nasuta. (g), (h) Rhynchobodo simius (g) side view, (h) ventral view. (i) Dinema validum,ventral view (j) Dolium sedentarium. (k) Heteronema exaratum, ventral view (I) Notosolenus apocamptus, ventral view andtransverse section. (m) Notosolenus canellatus, ventral view and transverse section. (n) Notosolenus ostium, ventral view andtransverse section. (n) Peranema fusiforme. Scale bar (bottom right) represents 10 ~m.
Fig. 3. All micrographs to the same scale and are DIe images unless otherwise specified. (a)-(c) Dolium sedentarium, (a) cell •showing emergent flagellum, (b), (c) range of cell width in D. sedentarium, note in (c) the four visible surface grooves. (d)Heteronema exaratum. (e)-(g) Notosolenus apocamptus, (e) general appearance of cell, (f) ventral view, (g) ventral viewof cell with N. ostium-like cell profile (see text). (h)-(j) Notosolenus canellatus, (h) general appearance of cell, (i) dorsal viewshowing surface grooving, (i) lateral view of the same cell as in (i), showing flattening. (k), (I) Notosolenus ostium, (k) ventralview showing apical detail, ventral groove and the lengths of the flagella (anterior flagellum to the left), (I) same cell as in (k)showing general appearance of the cell body. (m), (n) Peranema fusiforme, (m) small cell showing the flagella and the rods of thefeeding apparatus (arrow indicates the short posterior flagellum), (n) large cell. (0), (p) Petalomonas minuta, note the mediangroove. (q), (r) Petalomonas poosilla, (q) is phase contrast. (sHu) Petalomonas abscissa, (s) view showing cell outline, anteriorflagellum and thin lateral margin on the left, (t) ventral view showing thick median double ridge and flagellar pocket opening,(u) general view of cell. Scale bar (bottom right) represents 10 ~m.
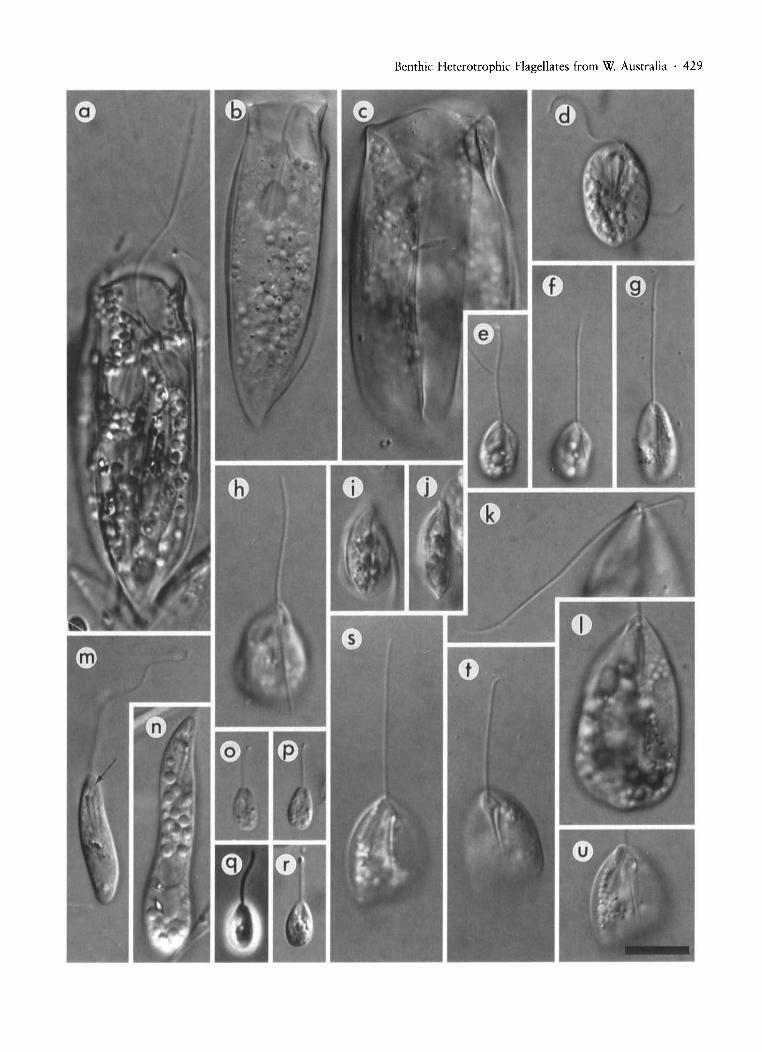
Benthic Heterotrophic Flagellates from W. Australia . 429

430 . D. J. Patterson and A. G. B. Simpson
Notosolenus apocamptus Stokes, 1884. Figs. 2 t 3 e-g.
Description: Cell length 7-18 /lm, mostly 8-12/lm.Cell ovate, flattened, anterior end narrowed, posteriorend rounded. A broad longitudinal dorsal groove runsalong the entire length of the cell. Cell convex bothsides of the dorsal groove, ventral side fairly flat.Two flagella of unequal length emerge from an ovateflagellar pocket. Anterior flagellum up to 2 times celllength and posterior flagellum about 3/4 of the celllength. Cells move by smooth gliding with ventral sideand anterior flagellum against surface. One cell(Fig. 3 g) was observed with a body profile resemblingthat of N. ostium, and measuring 16/lm, although thecell lacked the collar of N. ostium and, unlikeN. ostium, the posterior flagellum was about 3/4 thecell length. Observed from Little Lagoon and HamelinPool.
Remarks: This species was common and widespread,and generally consistent with the text of the observations of Larsen and Patterson [22] but we note thattheir Fig. 19 a has the recurrent flagellum drawn asbeing shorter than in photographs (Fig. 21 b, [22]) oras described in the text. See Larsen and Patterson[22] for detailed comments.
Notosolenus canellatus Skuja 1948, Figs. 2 m, 3 h-j.
Description: Gliding euglenid, measuring 1518/lm, anterior flagellum about 1V2 times celllength, posterior flagellum about cell length. Anteriorend with small but obvious collar, posterior end slightlypointed, dorsal face with one median and two lateralgrooves but basically convex. Edges of the cell, especially at the posterior end, are thin flanges. Ventrallyflattened, occasionally with one fine median ventralgroove (possible to overlook). Observed from Little Lagoon.
Discussion: The overall profile and grooving of thisorganism is as described by Skuja for an isolate withcontractile vacuoles from freshwater [45], but Skuja's isolates measured 20-24/lm. However, Larsen[21] identified isolates measuring 12-25 /lm and without contractile vacuoles from the Danish Wadden Seawith this species. Adopting Larsen's approach, we regard the organisms observed by us as also belongingto this species. N. canellatus is the only Notosolenuswith all of the following features; posterior flagellumabout cell length, a pointed posterior end and withthree dorsal grooves.
Notosolenus ostium Larsen and Patterson, 1990,Figs. 2 n, 3 k, 3 l.
Description: One cell observed, 30 /lm. Cell elongateovate with a prominent dorsal longitudinal groove. Theflagellar pocket is on the right side, the nucleus on theleft. Two flagella of unequal length. Anterior flagellum1V2 times cell length, held straight forward. Posteriorflagellum about one quarter of cell length. Anteriorpole of cell with small collar, and posterior end of cell
with blunt nipple. Cell moved by smooth gliding. Cellcontained ingested eukaryotic algae. Observed fromLittle Lagoon only.
Remarks: Our observations were consistent with thedescription in Larsen and Patterson [22]. Ventralgrooves and ingestion apparatus, both of which are difficult to see [22] were not observed. Notosolenus ostium is distinguishable by its large size and elongatecell body with simple surface sculpting (i.e. a single prominent median groove).
Peranema fusiforme Larsen, 1987, Figs. 2 0, 3 m, 3 n.
Description: Cell length 20-40 /lm, mostly at lowerend of range. Cell usually narrow spindle-shaped whengliding, but metabolic and more expanded whensquirming. About 40 delicate pellicular grooves. Sausage shaped bodies sometimes aligned along the striations. Anterior flagellum as long as the cell or slightlylonger, posterior flagellum short, curved and does notextend as far as the ingestion apparatus. Flagellar pocket is located in the first half of the cell on the left side.Ingestion apparatus with two rods which narrow towards their posterior ends. Anterior part of the rodswith cap-like swellings. Nucleus in the posterior partof the cell. 0 bserved from Little Lagoon.
Remarks: Shape, size, striations, location of nucleiand short recurrent flagellum correspond well with previous observations of Peranema fusiforme [8, 21, 22].We note that, with most cells being 25 -30 /lm, the organisms observed here are a little smaller than thosereported earlier. Occasional large cells were observedby Larsen and Patterson [22] and by Ekebom et al.[8]. P. fusiforme and P. macrostoma [8] are the onlyspecies in the genus that have been described with avery short recurrent flagella. The two species can bedistinguished by the smaller size and less well developed ingestion rods of P. fusiforme.
Petalomonas abscissa (Dujardin, 1841) Stein, 1859,Figs. 3 s-u, 4 a.
Description: Cell length 15-16 /lm. Emarginate posterior end. With two dorsal discontinuities runningparallel to lateral margins, left one well-developed,right one weakly developed. The lateral margins arethin flanges. One narrow curving groove ventrally,with raised strengthened margins that form a doublekeel. The support for the ventral grooves arise as anextension of a strengthened collar around the openingof the flagellar pocket. One flagellum, 1V2-2 times celllength, pointing anteriorly. Flagellar pocket to the leftventral side. Observed from Little Lagoon.
Remarks: This organism can easily be identified withthe taxon referred to as P. abscissa by Larsen [21], Larsen and Patterson [22], and Ekebom et al. [8]. Larsen[21] discusses the difficulties associated with the identity/name of this species, and we further note the overlap in size and form with P. mira var. bicarinata Skuja.

Benthic Heterotrophic Flagellates from W. Australia . 431
n
h
a
,
o
9
m
f
o
o
k
d~
00
cc::::>
j
b
o~
a
~
Fig. 4. All drawings to the same scale and are ventral views and transverse sections unless otherwise specified. (a) Petalomonasabscissa. (b) Petalomonas raiula, ventral view only. (c) Petalomonas minuta. (d) Petalomonas poosilla. (e)-(h) Ploeotia azurina,(e) "large morph", ventral view, (f) "large morph", dorsal view and transverse section, (g) "small morph", ventral view, (h)"small morph", dorsal view and transverse section. (i) Ploeotia corrugata. (i) Ploeotia costata. (k) Ploeotia oblonga. (I) Ploeotiaplumosa. (m) Ploeotia pseudanisonema. (n) Ploeotia punctata. Scale bar (bottom right) represents 10 J.lm.

432 . D. J. Patterson and A. G. B. Simpson
Petalomonas minuta Hollande, 1942, Figs. 30, 3 p,4c.
Description: Gliding, flattened euglenid, Cell ovate,length 6 - 9 IJ.m. With median groove down the lengthof the cell, probably on dorsal surface. One flagellumabout same length as body and pointing anteriorly. Observed from Little Lagoon.
Remarks: This species has been described from marine habitats by Larsen [21] and by Larsen and Patterson [22]. Larsen [21] mentioned a median groove whileLarsen and Patterson, like Hollande [18], specify thatthis groove is located ventrally. Our present observations suggest that the groove may be located dorsally, and this matter warrants further attention.Similar in size to P. minor which can be distinguishedby a prominent dorsal ridge, and to P. poosilla whichhas no well-marked ridges or grooves.
Petalomonas poosilla (Skuja, 1948) Larsen andPatterson, 1990, Figs. 3 q, 3 r, 4 d.
Description: Gliding, somewhat flattened, euglenid.Cell ovate, measuring about 8 IJ.m. May have veryslight dorsal ridge. Flagellum about cell body length,observed occasionally from Little Lagoon and oncefrom Hamelin Pool.
Remarks: Similar to P. minuta but lacks the wellmarked median groove. Also similar to P. minor butlacks well-marked dorsal ridge and with different dorso-ventral profile.
Petalomonas raiula Larsen and Patterson, 1990.Fig.4b.
Description: One vaguely diamond-shaped cell observed measuring 12 IJ.m, distinguished by 3 backwardly directed points, one median, two lateral.Single flagellum is about as long as body and directedanteriorly. From Little Lagoon.
Remarks: In accord with original and only previousdescription by Larsen and Patterson [22].
Ploeotia azurina Patterson and Simpson, n. sp. Figs.4e-h, 5 a-h.
Diagnosis: Ploeotia measuring 10-16 IJ.m long, with7 dorsal ridges, two forming the lateral margins, notvery flattened ventrally, ovate with pointed posteriorend, and recurrent flagellum twice to more than three
times the cell length. Type micrograph: Fig. 5 a (Notethat Figs. 5 a and 5 d are of the same individual).
Description: Ploeotia measuring 10-16IJ.m, average13.5 IJ.m. Profile ovate, dorsally convex with 7 ridges (2forming the margins) running longitudinally or at aslightly oblique angle. Usually only 6 ridges can beseen at any time. Cells may be flattened ventrally witha mid-ventral ridge arising anteriorly at the opening ofthe flagellar pocket. In some cells two lateral ventralridges may also be seen and these cells appear less ventrally flattened. Posterior flagellum 2- 3V2 times celllength and appears thick, anterior flagellum aboutthe same length as the cell. Posterior end of cell tapers. Cell moves by gliding, without Anisonema-likereversals. Observed from Little Lagoon, moderatelycommon.
Remarks: This species has a high level of similaritywith P. corrugata, which usually seems to have 6 dorsalridges and a mid-ventral ridge [see 8]. Although previous descriptions [22] of P. corrugata refer to 6 dorsalridges, a more careful scrutiny of cells led to the observation of 7 dorsal ridges in at least some cells [8].P. azurina can be distinguished from P. corrugata bythe less flattened dorso-ventral profile. P. azurina is significantly more slender than P. corrugata and widest inthe anterior half of the cell, whereas P. corrugata iswidest in the middle. P. azurina also differs fromP. corrugata by being slightly larger in size with apointed rather than emarginate posterior end, with amore visible median ventral ridge, and, usually, witha longer and thicker recurrent flagellum. There is alsosome similarity with P. decipiens and with P. adhaerens,from both of which P. azurina differs in being smaller,less flattened ventrally, and by having a slimmer profilewhen viewed dorso-ventrally. All other described species of Ploeotia not mentioned above differ fromP. azurina either by having dorsal grooves rather thanridges or by having less than six dorsal ridges.
We have noted two common morphs. The first("small morph") is noticeably flattened ventrally witha single ventral ridge and a relatively thick recurrentflagellum up to 3V2 times body length (as in the typemicrograph). The other ("large morph") has slightlyspiral grooves, with a shorter (maximally 2112 timescell body length) posterior flagellum, more roundedin cross section and, typically, with two additionalridges, one on each side of mid-ventral ridge. Cells intermediate between these two forms were observed,
Fig. 5. All micrographs to the same scale and are DIe images unless otherwise specified. (a)-(h) Ploeotia azurina, (a) general ~appearance of cell (phase contrast), (b) cell with long posterior flagellum and single conspicuous ventral ridge, (c) ingestionapparatus detail, (d), (e) micrographs showing the variability of size and dorsal grooving, (f) cell with short posterior flagellum,(g) end on aspect showing corrugated dorsal ridges, inconspicuous ventral ridges, (h) lateral view of only slightly flattened cellwith stronger ventral discontinuities. (i)-(k) Ploeotia corrugata, (i) general appearance, note seven dorsal grooves, (i) ventralview, (k) dorsal view. (lHm) Ploeotia costata, (I) general appearance of cell, (m) dorsal view showing furcate ridges. (n)-(o)Ploeotia oblonga, (n) General appearance less anterior flagellum, note furcate ridges, (0) ventral view showing prominentmedial ventral ridge with furcate edges. (p)-(r) Ploeotia pseudanisonema, (p) dorsal view, (q) ventral view, (r) general appearance, note length of posterior flagellum. (sHu) Ploeotia plumosa, (s) general appearance, (t) dorsal view showing fine ridgesand abutting curved inclusions, (u) flagellar pocket and mouth detail. Scale bar (bottom right) represents 10 /lm.

MM'<t"
.~
-;;;.....</)
;j
<~a.....,
0J::</)
(I)...-'2<)0.0
«l
ii:u:..a0.,
0.....0..(I)...(I)
::r:u:..a...c(I)
i=Q

434 . D. J. Patterson and A. G. B. Simpson
and for this reason we regard all individuals in this category as being from a single species.
The single cell described under the name P. decipiensby Ekebom et al. [8], appears to be a large representative of P. azurina, and the reference to corrugated cellsunder the name P. punctata in Ekebom et al. [8] mayalso refer to P. azurina. More work in the area of smallindividuals of the genus Ploeotia is required.
Ploeotia corrugata Larsen and Patterson, 1990. Figs.4i, 5 i-k.
Description: Cell length 10-15 /lm. Cell flattened,with a ovate profile but posteriorly with a slight indentation. Dorsal side with 6 or 7 dorsal grooves, two ofwhich form the lateral margins of the cell, ventrallywith single ridge to left of centre arising at openingof flagellar pocket. Observed occasionally from LittleLagoon.
Remarks: The cells described here are largely inagreement with the original description [22] but weconfirm the presence of a ventral ridge noted for thefrist time by Ekebom et al. [8] and the presence of 7dorsal ridges in some cells. We assume that previousdescriptions of 6 dorsal ridges were in error. In Ekebom et al. [8] similarities with Lentomonas applanatum [10] are discussed and we here confirm thatdescriptions of P. corrugata by Larsen and Patterson[22] and of 1. applanatum by Farmer and Triemer[10] are of the same species. This is not the same asEntosiphon applanatum as described by Preisig [41]because the Entosiphon has a protrusible siphonwhereas Ploeotia does not. There are similarities withthe new species. P. azurina (see above). P. corrugatamay be easily distinguished from all Ploeotia spp. except P. azurina by the corrugated appearance of thedorsal face.
Ploeotia costata (Triemer, 1986) Farmer andTriemer, 1988. Figs. 4 j, 51, 5 m.
Description: Cells 15 - 23 /lm long, ovate, sometimesslightly dorso-ventrally flattened. A total of fivegrooves, two dorsal and three ventral with the groovesappearing slightly twisted. Two flagella of unequallength, anterior flagellum about cell length, posterior
flagellum thicker and 2 - 2.5 times cell length. An ingestion apparatus with two rods visible, right hand rodwith slight lateral protrusion. Cells glide. From LittleLagoon.
Remarks: This organism is broadly consistent withthe various descriptions in the literature [8, 9, 23,52]. The most similar species is P. oblonga (see below). The species have been separated on the basisof a number of criteria (cell size, strength of mid-ventral ridge complex, width of this complex relative towidth of the recurrent flagellum, lengths of flagella relative to body length and cell shape [22]). We have relied on the strength of the mid-ventral ridge complex(groove plus associated ridges) to separate the two species [8]. This is stronger in P. oblonga than in P. costata.We note under our remarks on P. oblonga that theridges in both species are furcate. With the acquisitionof more data, it has become increasingly difficult todetermine where the boundary between the speciesmight lie. Indeed, we are faced with a problem of determining how many species should be recognised inthis area.
Ploeotia oblonga Larsen and Patterson, 1990. Figs.4 k, 5 n, 5 o.
Description: Cell 16-36 11m long, oblong to ovate,slightly flattened dorso-ventrally, with two majorgrooves dorsally and three major grooves ventrally.Very fine grooves line the major grooves on both sidesindicating furcate nature. Collectively groove plus furcate ridges may be regarded as a "ridge complex". Themedian ventral ridge complex is more prominent thanthe two lateral ones. Two flagella of unequal length,anterior flagellum almost cell length, posterior flagellum, 2-2112 times cell length, and thicker than the anterior flagellum. Ingestion apparatus with a knob-shapedprotrusion. Moves by gliding. Found in Little Lagoononly.
Remarks: In agreement with the original description[22] although we have extended the size range. Largerindividuals are easy to distinguish from other species,but there is considerable overlap of size and appearanceof smaller cells of this species with P. costata (seeabove). P. costata and P. oblonga differ from all otherspecies by having 5 furcate "ridge complexes".
Fig. 6. All micrographs are DIC images unless otherwise specified. Figures (b)-(z), (is) are all to the same scale. (a)-(d) Ploeotia ~
robusta, (a) cell showing flagellar length (phase contrast, scale bar represents 20 /lm), (b) general appearance of cell, (c) dorsalview, (d) ventral view. Note in (c) and (d) the fine grooves. (e), (f) Ploeotia vitre, (e) dorsal view showing ridges with adherentparticles, (f) ventral view, note closely spaced median ventral ridges. (g)-(i) Ploeotia cf. punctata showing range of sizes andappearances. (j), (k) Anehmia exotica (j) is a more lateral view than (k) is. (I) Colpodella pugnax, 4-way division cyst. (m)Cafeteria ligulifera, note only part of the recurrent flagellum is visible in this micrograph. (n)-(p) Cafeteria roenbergensis,(n) swimming cell, (0) attached cell showing anterior flagellum, (p) same cell as in (0) showing recurrent flagellum. (q) Developayella elegans, swimming cell (phase contrast). (r) Bordnamonas tropicana. (s)-(v) Ancyromonas melba, (s) general appearance of cell, (t) view showing anterior flagellar depression and similarity in thickness of the flagella, (u) ventral view showingflagellar lengths, (v) swimming cell, note length of anterior flagellum. Swimming is "backwards (i.e. in the direction indicatedby the arrow). (w), (x) Ancyromonas sigmoides, (w) cell without anterior flagellum (phase contrast), (x) small cell with thinanterior flagellum. (y) Colpodella unguis. (z), (is) Metopion fluens. Scale bar for (b)-(z), (is) (bottom right) represents 10 /lm.
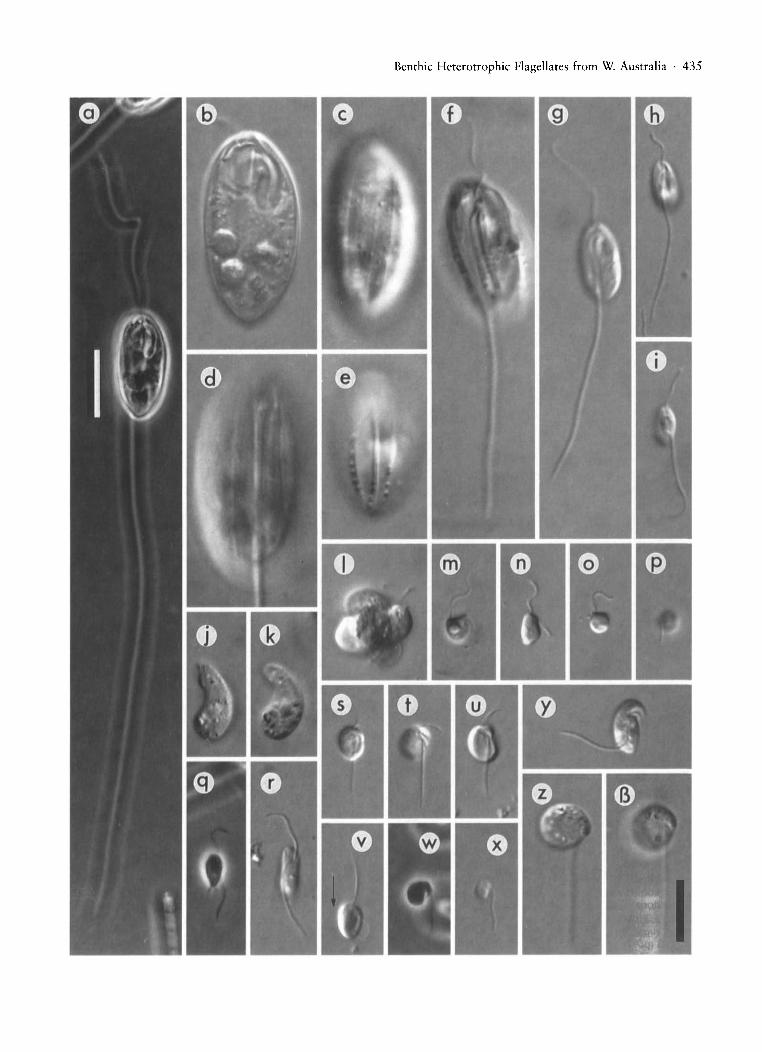
Benthic Heterotrophic Flagellates from W. Australia . 435

436 . D. J. Patterson and A. G. B. Simpson
Ploeotia plumosa Ekebom et al., 1996. Fig. 41, 5 s-u.
Description: Cell 20-22 f.lm long, oblong to ovate,slightly flattened, with 4 grooves dorsally and 5grooves ventrally. The dorsal grooves are more widelyspaced than the ventral ones. Slightly curved inclusionsabut on the grooves (Fig. 41). Anterior flagellum aboutcell length. Posterior flagellum 2-3 times cell lengthand thicker than the anterior flagellum. Ingestion apparatus with two well-developed rods, may be prominentat anterior end. Often with one large or several smallerhomogeneous inclusions at the posterior end of the cell.Cells glide smoothly in straight lines, and may jerkbackwards. Two cells encountered at Little Lagoon.
Remarks: The observations made of these cells areconsistent with those of cells found on the Great Barrier Reef [8]. We note that many species of Ploeotiahave 10 grooves in total, and that we may have overlooked one groove in this species. The prominent inclusions abutting the grooves are distinctive to thisPloeotia.
Ploeotia pseudanisonema Larsen and Patterson,1990. Fig. 4 m, 5 p-r.
Description: Two cells observed, 18 and 19 f.lm long,with approximately 10 longitudinal grooves fairlyevenly spaced, median ventral furrow only set off atits anterior end. Anterior flagellum about the samelength as the cell. Posterior flagellum very long 3-4times the length of the cell. Glides quickly, and capableof Anisonema-like retractions. Ingestion apparatus extending almost the length of the cell, with slight prominence on one rod. One cell from Little Lagoon, onefrom Hamelin Pool.
Remarks: Generally in agreement with the observations by Larsen and Patterson [22] and Ekebom et al.[8]. See Larsen and Patterson [22] for detailed comments on the distinctiveness of the species.
Ploeotia efr. punetata Larsen and Patterson, 1990.Figs. 4 n, 6g-i.
Description: We observed a variety of small glidingcells resembling the organisms assigned to Ploeotiapunetata by Ekebom et al. [8]. The identity of theseorganisms is unclear and this is discussed below. Cellsmeasured from 7-13 f.lm, elliptical to ovate, withoutclearly visible grooves in any cell, but some cells havea ventral furrow/shelf. No punctae seen. Anterior flagellum 11;2-2 times cell length. Posterior flagellum2V2-3V2 times cell length but never with proximal section noticeably more thick than distal section. Glidingmotion slow (normal) or fast, with Anisonema like reversals in slow and fast cells, no "A. trepidum-like arrests" noted [see 22]. Ingestion apparatus visible inmost cells, but not strongly developed; not recordedin some cells. Anterior part of cell may be truncate.The posterior flagellum trails along the ventral ridgeif present when cell is moving. All cells observed fromLittle Lagoon.
Remarks: In adding a variety of species to the genus Ploeotia Larsen and Patterson [22] included anumber of small species (P. punetata, P. longifilum,P. pseudanisonema). Ekebom et al. [8] observed manysmall cells and noted that the characteristics used byLarsen and Patterson did not always apply consistently. They consequently placed an array of small gliding individuals in the taxon "P. punetata" even thoughthese cells did not have some of the features held byLarsen and Patterson [22] to be diagnostic; cell shapewas not consistently oblong, longitudinal ridges andpunctae were not always observed, low amplitude vibrations were never observed, and a ventral groove notmentioned for this species by Larsen and Patterson wasobserved. The observations from Little Lagoon are similar to those of Ekebom et al. [8] and confirm that smallgliding euglenids exhibit a variety of form, and thatcharacters do not distribute in a concordant fashion.In the absence of clear-cut demarcations within this"territory", we feel that we have no alternative butto adopt a "lumping" strategy even though this maydraw together unrelated taxa. Further efforts are required to establish useful diagnostic characters in thisterritory.
Ploeotia robusta Larsen and Patterson, 1990. Figs.6a-d, 7a.
Description: Cells elliptical, 28-32 f.lm long, posterior end slightly pointed. Anterior flagellum almost 2times cell length, posterior flagellum 4 - 5 times celllength and tapering from a very thick base. Capableof Anisonema-like reversals [22]. Single ventral discontinuity arising from the ventral opening of the flagellarcanal at the anterior end and with at least eight delicategrooves arranged evenly around the body. From littleLagoon.
Remarks: Largely consistent with original and onlydescription to date [22] excepting that we have nowobserved fine surface grooves. These could have beenoverlooked in the original description. The cells fromWestern Australia were at the low end of size rangenoted by Larsen and Patterson [22].
Ploeotia vitrea Dujardin, 1841 emend. Farmer andTriemer; 1988. Figs. 6e, 6(' 7b.
Description: Two cells observed, 22 and 24 f.lm long,respectively. Anterior flagellum same length as the cell.Posterior flagellum 2 times cell length and thick. Celloval or slightly flattened in cross-section with 4 dorsal, 2 lateral and 4 ventral ridges. The ridges are prominent and, with the exception of the two middlemostventral ridges, Widely spaced. These mid-ventral ridgesare closer to each other than the remainder, separatedby about a ridge-width. Spherical particles may adherealong the posterior half of each ridge. Feeding apparatus visible for 3/4 of the length of the cell with a slightprotrusion at the top of the right hand rod. Observedfrom Little Lagoon.
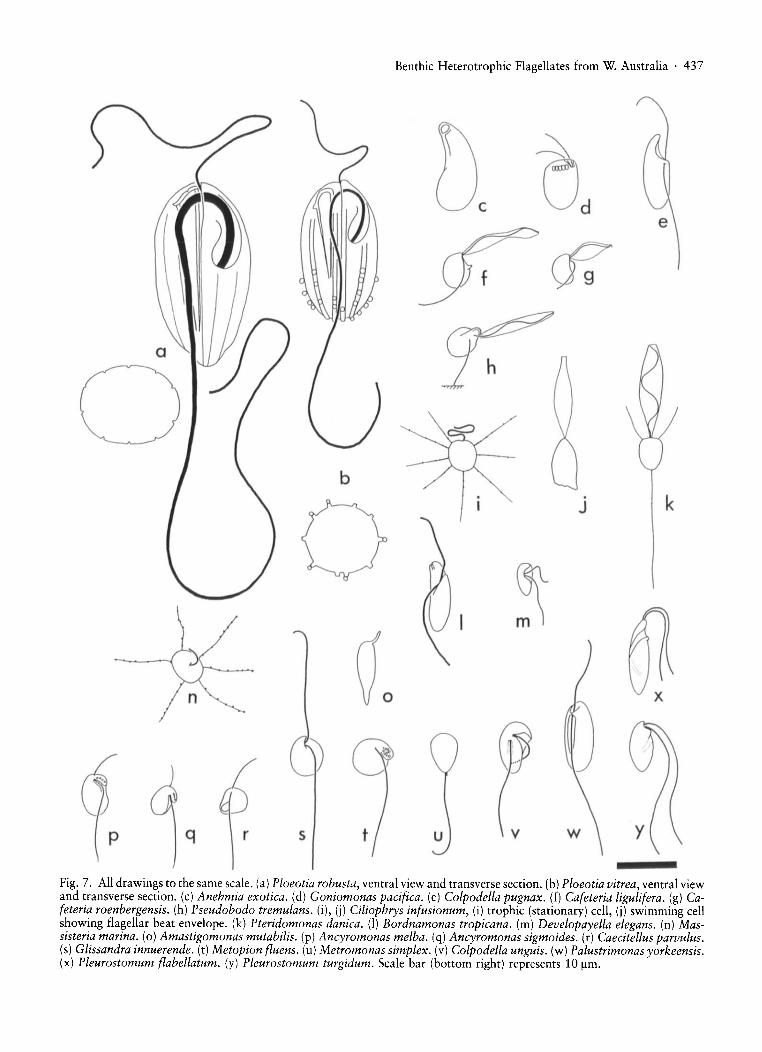
Benthic Heterotrophic Flagellates from W. Australia· 437
k.J
J
v9
~d
~
v
@
c
Vo
b
srqp
Fig. 7. All drawings to the same scale. (a) Ploeotia robusta, ventral view and transverse section. (b) Ploeotia vitrea, ventral viewand transverse section. (c) Anehmia exotica. (d) Goniomonas pacifica. (e) Colpodella pugnax. (f) Cafeteria ligulifera. (g) Cafeteria roenbergensis. (h) Pseudobodo tremulans. (i), (j) Ciliophrys infusionum, (i) trophic (stationary) cell, (i) swimming cellshowing flagellar beat envelope. (k) Pteridomonas danica. (I) Bordnamonas tropicana. (m) Developayella elegans. (n) Massisteria marina. (0) Amastigomonas mutabilis. (p) Ancyromonas melba. (q) Ancyromonas sigmoides. (r) Caecitellus parvulus.(s) Glissandra innuerende. (t) Metopion f/uens. (u) Metromonas simplex. (v) Colpodella unguis. (w) Palustrimonas yorkeensis.(x) Pleurostomum f/abellatum. (y) Pleurostomum turgidum. Scale bar (bottom right) represents 10 /lm.

438 . D.]. Patterson and A. G. B. Simpson
Remarks: Agrees well with detailed observations byFarmer and Triemer [9] and Larsen and Patterson [22].Cells observed fall inside reported size range of 1625 /lm. P. vitrea is distinguishable from all other species in the genus by the prominence and similarity ofthe ten ridges.
Euglenozoa Incertae SedisAnehmia exotica Ekebom et al. 1996. Figs. 6 j, 6 k,7 c.
Description: Cell length 12 /lm. Cell sausage shapedand without flagella. Anterior end with an subapicaltriangular depression. Immediately below is a smallbulge with an underlying fibrous structure. The cellbody is curved and slightly twisted. Refractile dropletsof different sizes and food vacuoles suggestive of detritivory or protistivory are present in the sac-shapedposterior end. Cell glides slowly with anterior endpressed against the surface. The cell is held away fromthe substrate with the concave (ventral) side of the cellfacing the substrate. Observed occasionally from LittleLagoon.
Remarks: This organism corresponds closely to thedescription by Ekebom et al. [8]. We note here however a bulge underlying the mouth and an apparent underlying skeletal structure, which were overlooked inthe original description. The reasons for tentatively assigning this genus to the Euglenozoa are given in Ekeborn et aI., [8]. Anehmia exotica is the only aflagellate,non-metabolic, non-amoeboid heterotrophic glidingprotist known.
Cryptomonadida Senn, 1900.Goniomonas pacifica Larsen and Patterson, 1990.Fig. 7d.
Description: Size 4-8/lm, mostly between 4 and6 /lm. Cell ovate in profile, truncated anteriorly, flattened. Two flagella less than cell length emerge fromanteriorly directed pocket on one side and beat in frontof the cell. A row of extrusomes runs across the cellclose to anterior end. Surface grooves not observed.Observed from Little Lagoon and one larger cell fromHamelin Pool.
Remarks: Observations largely in agreement withoriginal and subsequent descriptions [e.g. 8, 22]. Thisspecies has now been reported with a more extendedrange of sizes (4-15/lm; this paper; [55]) in contrastto the original description as 8-10 /lm [22]. Surfacegrooving can be difficult to see. See Patterson and Larsen [22] for detailed comments about the distinctiveness of the different nominal species of Goniomonas.
Alveolates
This is probably a monophyletic grouping of tubulocristate protists in which the cell surface is underlain bya layer of abutting alveoli. Includes ciliates, dinoflagellates, apicomplexa and Colponema [31]. Monophylyindicated by 18S-like RNA sequence analyses [14].
Colpodella pugnax Cienkowski, 1865. Figs. 61(division cyst), e.
Description: Size 12-15/lm, 12-13 /lm in cellswithout food vacuoles. In swimming cells the cell bodyis basically elliptical but with a prominent curved anterior rostrum. The heterodynamic flagella are 1lf4-1V2times cell length, insert at the base of the rostrum andare almost directly opposed. They insert in separatetiny flagellar depressions or grooves. The cells swimmore rapidly than other flagellates observed in the saturated brine samples and rotate while they move. Thecells do not glide nor show plasticity. There is no prominent gutter or groove on the cell body. Several speciesof the chlorophyte genus Dunaliella were also present,and one swimming Colpodella was observed with aningested D. salina in a posterior food vacuole. Severalcysts with ingested algal material (Dunaliella spp.)were observed, and in various stages of formation oftwo or four daughter cells (Fig. 61). Observed onlyfrom the saturated brine puddle.
Remarks: This species could not be distinguished byits morphology from Cienkowski's [7] description ofColpodella pugnax, which he observed attacking chlorophyte alga. Cienkowski's observations were of an organism from a freshwater environment.
The light microscopical appearance of this isolateagrees with more extensive observations reported bySimpson and Patterson [43]. In common with othermembers of the genus, this species divides after feedingwithin a cyst which may contain either two or fourdaughter cells [7, 43].
Colpodella is a genus of free-living flagellates whichattack eukaryotic prey by sucking up the cytoplasm viaa rostrum. Several species of Colpodella, includingC. pugnax, have been examined at the ultrastructurallevel and been shown to possess in apical complex, micropores, multiple cell membranes (i.e. cortical alveoli)and tubular mitochondrial cristae [5, 13,29,43] as dothe apicomplexan sporozoa. We regard Colpodella asthe sister taxon to the parasitic apicomplexa (inclusiveof Perkinsus).
Despite the distinctive ultrastructural appearance ofColpodella, light microscopical identification can bedifficult, and confusion has been made in the past withthe genus Bodo (for example, Colpodella pugnax wasincorrectly identified as "Bodo sp. 3" in a recent study[40]). Some of these accounts have reported a "kinetoplast" [17, 40] although ultrastructural studies confirm that the staining structure is not a kinetoplast. Inlieu of ultrastructural observations, we have relied onthe distinctive cyst development and the insertion of theflagella into separate depressions to distinguish this organism from the genus Bodo.
Colpodella pugnax has previously been reportedfrom an artificial hypersaline lagoon in South Australia [43] and from the hypersaline Hutt Lagoon in Western Australia [40]. It may be distinguished from allother Colpodella sp. (and all Bodo sp.) by the size ofthe single-pointed, broad and curved rostrum while

lacking a ventral gutter [43] and while swimming witha spiralling motion rather than with one face heldagainst the substrate (see C. unguis).
Colpodella unguis: Patterson and Simpson, n. sp.Figs. 6 y, 7 v.
Diagnosis: Cell with two orthogonal flagella inserting close together into separate pockets/grooves. Anterior portion of cell forms a prominent, curved rostrum,the inside edge of which is connected back to the cellbody by a thin flap of cytoplasm, forming a deep,wedge-shaped channel with the anterior flagellum inserting at the "tip" of the wedge. Cell somewhat flattened and swims in close association with the substrate.7 -10 J..lm long. Type micrograph: Fig. 6 y.
Description: The cell is 7 -10 J..lm long, bean shapedand is somewhat flattened dorso-ventrally. The two flagella insert on the side about a third of the way downthe cell. The posterior flagellum inserts into a posteriorly directed fine groove and is about 11/2 times celllength. The anterior flagellum is shorter and emergesat right angles to the posterior flagella into a deep channel open on one side. The channel is wedge- or sharkfin (dorsal)-shaped with the flagellum inserting at thepoint/tip. Due to the channel, the cell anterior to theflagella insertion appears as heavy, claw-like rostrumwhich curves round such that the end points almostposteriorly. The inside curve of the "rostrum" formsthe anterior-most edge of the channel.
The cell swims very rapidly with one side in closeassociation with the substrate. The posterior flagellumactively beats during swimming. It is unclear where theanterior flagellum is situated during swimming but itmay be held close to the body. The body vibrates whileswimming. No other locomotion, such as gliding orsquirming, was observed. Observed occasionally fromHamelin Pool and the hypersaline pond.
Remarks: We assign this species to Colpodella because of the presence of a suite of features found insome species in this genus and not in other genera.These features are the orthogonal or opposed flagellainserting into separate pockets or (very) shallowgrooves and a prominent, curved rostrum with a thinflap of cytoplasm connecting the inside curve of therostrum back to the cell body (see Colpodella pugnax). In C. unguis this flap forms the dorsal face ofthe wedgeshaped channel. However, we did not observe the feeding behaviour nor the division cyst heldto be distinctive for Colpodella [43] (although we notethat we did not observe any food vacuoles containingbacteria, either). The generic assignation is currentlyconsidered tentative. Further research on the ultrastructure and life-cycle of this organism is requiredto test the assignation of this species to Colpodella.
C. unguis is distinguishable from all Colpodella species accepted in the recent review of the genus [43].C. vorax, C. angusta, and C. gonderi have much shorter and less curved rostra than C. unguis (amongst otherdifferences). C. unguis may be distinguished from
Benthic Heterotrophic Flagellates from W. Australia· 439
C. pugnax and C turpis because the latter two have flagella of almost equal length which insert at an obtuseangle (with one flagellum pointing almost anteriorly)and they both swim with a spiralling motion. C. edaxhas a prominent gutter posterior to the flagellar insertion [19, 29] which is absent from C unguis.C. perforans may be the established species most similar to C. unguis, but apepars to lack a flap connectingthe rostrum to the cell body. In addition, the drawingsand micrographs of Brugerolle and Mignot [5] suggestthat the rostrum of C. perforans is fairly straight whenthe cell is not feeding, in contrast to the extremelycurved nature of the structure in C. unguis.
Stramenopiles Patterson, 1989, Bicosoecida Grasseand Deflandre 1952
Cafeteria ligulifera Larsen and Patterson, 1990. Figs.6 m, 7(.
Description: Small biflagellated cells measuring 56 J..lm, attached by tip of posterior flagellum when feeding. Posterior flagellum about 2 times the length of thecell, anterior flagellum about 3 times cell length. Flagella insert subapically, and there is a small projectionfrom the ventral surface posterior to the flagellar insertion. An extremely fine hoop-like extension of the projection was observed in one cell. Observed occasionallyfrom Hamelin Pool.
Remarks: Largely consistent with the description inLarsen and Patterson [22] although these observationsextend the size range to 3-6 J..lm. Can be easily distinguished from Cafeteria roenbergensis and other speciesin the genus by the length of both flagella and fromPseudobodo tremulans by the prominent anterior collar in the latter.
Cafeteria roenbergensis Fenchel and Patterson 1988.Figs. 6 n-p, 7g.
Description: Small D-shaped flagellate, length 3.55 J..lm. When feeding, attaching by tip of posterior flagellum, and creating a current of water with the anterior flagellum. Anterior flagellum is usually as long asthe body, occasionally longer, but never more than 2times cell length. When swimming anterior flagellumis directed forwards. The posterior flagellum is aboutas long as the body and trails in swimming cells. Observed from Hamelin Pool and hypersaline pond.
Remarks: Appearance consistent with descriptionsin Fenchel and Patterson [12] and Larsen and Patterson [22]. Distinguishable from all other naked bicosoecids except C. marsupialis by the shortness of theanterior flagellum. Lacks the long ventral furrow ofC. marsupialis [22]. C. roenbergensis was unexpectedly not observed from Little Lagoon.
Pseudobodo tremulans Griessman 1913. Fig. 7 h.
Description: Biflagellated stramenopile, body 5 J..lm,with two flagella inserting subapically, and with a collar curving partly around region of flagellar insertion.

440 . D. J. Patterson and A. G. B. Simpson
Attaching to substrate directly or indirectly (by strandof mucus) by posterior flagellum. From Little Lagoon.
Remarks: Largely consistent with descriptions byGriessmann [15], Larsen and Patterson [22] interalia. Like C. ligulifera the anterior flagellum is morethan twice cell length. Pseudobodo tremulans can bedistinguished from all Cafeteria species by its anteriorcollar and because, on occasions, it is attached indirectly to the substrate by a mucoid extension of theposterior flagellum.
Ciliophryida Febre-Chevalier, 1985.
Ciliophrys infusionum Cienkowski, 1876. Figs. 7 i,7 j.
Description: Two cells observed, 4 and 6.5 /lm wide.When in heliozoan form, cell is more or less spherical,with delicate arms, bearing extrusomes, arising fromthe cell surface. With a single flagellum held in figure-of-eight in the heiozoan form. Swims with flagellum directed anteriorly and pseudopodia retracted.Observed from Hamelin Pool.
Remarks: Cells in agreement with observations byLarsen and Patterson [22]. See Larsen and Patterson[22] for detailed discussion on the distinctiveness ofC. infusionum and its junior synonyms.
Pteridomonas danica Patterson and Fenchel, 1985.Fig. 7k.
Description: Two cells observed, both 5 /lm long,from Little Lagoon. Cells are apple shaped (pomiform) and attached to substrate by a long contractilestalk, with a single apical flagellum surrounded by awreath of fine arms.
Remarks: Cells in agreement with description by Patterson and Fenchel [34]. We concur with Patterson andFenchel [34] and Larsen and Patterson [22] on how thisspecies may be distinguished from other species of Pteridomonas and from Actinomonas.
Stramenopiles Incertae Sedis
Bordnamonas tropicana Larsen and Patterson,1990. Figs. 6 r, 7l.
Description: 8-12 f.!m long, body oval, often quiteslender. Two heterodynamic flagella insert into a subapical depression. The region anterior to the flagellardepression forms a short blunt rostrum with an anteriorly directed ingestion apparatus. The cell movesby skidding rapidly, photographs show both flagellain focus at the same time indicating that both flagellaare in contact with the substrate. The anterior flagellum is thick, slightly longer than the cell and held ina stiff irregular curve in front of the cell. The posteriorflagellum is thick, about 1-1112 times cell length andcurves upon emergence to trail behind the cell, passingfirst along a depression in the body surface. Neitherflagellum is acronematic and the anterior flagellum
has a particularly blunt end. Observed from Little Lagoon and Hamelin Pool.
Remarks: Patterson and Larsen [22] described thisorganism from several marine sites, and it has sincebeen observed more widely [8, 38, 55]. Larsen and Patterson [22] tentatively assigned this species to the stramenopiles, on the basis that the movement with theflagellum directed in front of the cell suggested the presence of flagellar hairs. We note here, however a number of features that are not consistent with placement inthe stramenopiles. They are; the flagella are thick; theflagella insert into common subapical depression; thecells glide/skid and there is an apical mouth visibleby light microscopy. These suggest that the affinitiesmay lie elsewhere (such as with the kinetoplastids),and further work is required.
The appearance and behaviour of the front flagellumduring movement combined are distinctive for Bordnamonas.
Developayella elagans Tong, 1995. Figs. 6 q, 7m.
Description: A single cell 5 f.!m long, ovoid with atapering posterior end, was observed. Two flagella insert into a deep lateral depression in the anterior half ofthe cell. Swimming similar to other stramenopiles withthe anterior flagellum beating sinusoidally and generating the swimming current. When the cell is stationary (feeding?) the cell is anchored to the substrate bythe posterior flagellum which also beats with a narrowenvelope. The anterior flagellum, meanwhile, has aslow, distinctive beat alongside the cell. Observedfrom Hamelin Pool.
Remarks: The appearance and behaviour of the cellwere as decribed in detail by Tong [53]. The flagellarbeat of non-swimming cells is said to be distinctivefor this monospecific genus.
Cercomonadida, Vickerman, 1983
Massisteria marina Larsen and Patterson, 1990.Fig.7n.
Description: Single cell observed. Globular flagellate6 /lm across with several thin, granulated pseudopods.The appearance corresponded well with the descriptionof the trophic cell by Larsen and Patterson [22] exceptthat only one curved shorted flagellum was observed.Observed from Little Lagoon.
Remarks: Massisteria marina does not appear to usethe flagella while in the stationary feeding state [22,35]. When the cells swim the pseudopodia are retracted and the flagella lengthens substantially [35].We have no evidence that the lack of a second flagellum is a consistent or significant feature and have nogrounds for distinguishing the observed organismfrom Massisteria marina. The size of the cell falls within the previously reported range of between 2.5 f.!m[38] and 9/lm [22].
M. marina is the only aloricate organism that hasbeen described that simultaneously bears branching,

extrusome covered pseudopodia and emergent flagellavisible with light microscopy.
Apusomonadidae Karpov and Mylnikov, 1989
Amastigomonas mutabilis (Griesman, 1913) Molinaand Nerad, 1991. Fig. 7 o.
Description: A single cell was observed from LittleLagoon. The cell was 10 ~m long, plastic and glidingwith a flexible snout and pointed rear end but no visible flagella.
Remarks: Cell observed was similar in profile to cellsassigned to this species by Larsen and Patterson [22]under the name of Thecamonas mutabilis, and by Patterson et aI., [38]. The genus Amastigomonas is distinguished by the flexible, but non-bulbous snout.A. debruneyi, the similar, but smaller, usually morewidespread species was not observed from Shark Baybenthic samples although it was observed duringwater column sampling [54].
Protista Incertae Sedis
Ancyromonas melba Patterson and Simpson, n. sp.Figs. 6 s-v. 7 p.
Diagnosis: Ancyromonas measuring 5- 7 ~m withanterior flagellum of thickness similar to that of recurrent flagellum. Ventral groove continues as a longitudinal ventral crease rather than an emarginatedepression. Type micrograph: Fig. 6 u.
Description: Biflagellated cells 5 - 7 ~m long, ovalshaped and dorsa-ventrally flattened with ventral sidehollowed. The anterior end of the cell is set off by aventral groove running from the left lateral marginto the right ventral aprt of the cell. The resulting"snout" has a few indistinct granules running alongit. The ventral groove continues longitudinally as aslight ventral crease (Fig. 6 s). The cell usually glideson a posterior flagellum which emerges from the nearthe anterior-most termination of the ventral groove.This flagellum is usually held under the cell during gliding. The posterior flagellum is 11/2-2 times cell lengthand tapers at its end rather than being sharply acronematico The anterior flagellum emerges from a slight depression at the right anterior end of the cell and sweepsin front of the cell as it glides. This flagellum is about 3J4of the length of the cell and is of similar thickness to theposterior flagellum.
One cell was observed swimming. The cell movedwith the posterior end directed forwards, the posteriorflagellum producing on oaring beat. The anterior flagellum trailed somewhat stiffly and passively behindmaking occasional contact with the substrate. In thiscell the anterior flagellum was 11/2 times cell lengthbut otherwise the morphology was the same as forthe gliding cells.
Observed from the hypersaline pond and once fromHamelin Pool.
Remarks: This species is strongly reminiscent of Ancyromonas sigmoides in the following features: small
Benthic Heterotrophic Flagellates from W. Australia . 441
size, dorso-ventrally flattening, gliding, presence of agroove from left margin to right ventral side of cell,granules along the snout, posterior flagellum arisingfrom ventral depression, anterior flagellum if presentarising from small depression near the base of thesnout. We regard these characters as justifying theplacement of both species in the same genus.
The genus is reviewed briefly by Patterson and Zo1£fel [39], and now contains A. melba, A. sigmoides, anda species observed by Lemmermann [23] under thename A. contorta [16]. Species added by Skvortzkov[47] are poorly described and because their generic affinities are highly uncertain, they are not included here[see 33]. We do not regard Ancyromonas magna ofZhue et al. [58] as a member of this genus. A. melbamay be distinguished from A. sigmoides by a numberof features. In the latter species the anterior flagellumis short (see Fig. 2 b in [28]), thin (Fig. 2 fin [38]) orabsent. The groove in A. melba is located exclusivelyon the ventral surface, while the posterior part of thegroove of A. sigmaides runs along the margin of thecell. In gliding A. melba cells, the recurrent flagellumlies closer to the mid-line of the body, whereas inA. sigmoides it is located marginally or projects fromthe side of the cell. A. melba is usually larger thanA. sigmoides and appears more ventrally hollowed.Backwards swimming has not been observed inA. sigmoides. It is possible that some of the larger cellsreported to date in the literature as A. sigmoides are ofA. melba. The contorted body of A. contorta distinguishes it from both of these other species.
The prominent anterior flagellum of A. melba makesit easy to distinguish this species from Metopion {luens.Caecitellus parvulus has a prominent anterior flagellum like A. melba but differs becaue the two flagellaappear to emerge from the anterio-posterior axis ofthe cell. Caecitellus has an ingestion apparatus supported by a hoop of cytoskeletal material [38] andno rostrum.
This species has now been observed in material fromAntarctica (Tong, V0rs and Patterson, in prep.).
AncyromonassigmoidesKent, 1880.Figs. 6 w,6x, 7 q.
Description: Gliding flagellate 3-5 ~m long. Onetrailing acronematic flagellum emerging from lateroventral groove. Anterior portion of cell forms short,laterally directed snout with granules. A second, verythin anterior flagellum emerges from a slight anteriordepression in many cells. Observed from Little Lagoonand Hamelin pool samples.
Remarks: Previously reported length ranges 2.57.6 ~m [16, 22, 55, 56]. Can most easily be distinguished from Ancyromonas melba (see above) becauseA. melba has an anterior flagellum of similar thicknessto the posterior flagellum for almost all of its lengthand the posterior flagellum is held more under the cellduring gliding. A. sigmoides resembles Metopion{luens but can be distinguished by the presence ofthe thin forwardly directed filamentous anterior flagel-

442 . D. J. Patterson and A. G. B. Simpson
Fig. 8. All micrographs DIe and to scale. (a)-(c) Glissandra innuerende, (a) general appearance of large cell, (b) small cellshowing flagellar insertion, (c) same cell as in (b) showing flagella. (d)-(g) Palustrimonas yorkeensis, (d), (e) general appearance of cell, note insertion of flagella into separate pocket/grooves, (f) ventral view showing shape of groove, (g) cell with largefood vacuole, note length of posterior flagellum. (h)-(k) Pleurostomum flabellatum, (h) general appearance of cell showingflagella, (i) cell showing both ends of the feeding apparatus, ending posteriorly at a food vacuole (arrow), (j) same cell as in (i)showing slightly raised edges of feeding apparatus, (k) flattened cell with short flagella. (I)-(n) Pleurostomum turgidum, (I)general appearance of cell showing flagellar insertion, (m) detail of flagella beating into groove, (n) small cell showing flagella.Scale bar (bottom right) represents 10 11m.
lum instead of a thicker, backwardly directed secondflagellum in M. {luens [39]. The anterior flagellummay easily be overlooked or may be absent inA. sigmoides [22], and the second flagellum may alsobe absent from M. {luens [38]. See Larsen and Patterson [22] for further comments on the distinctiveness ofthis species (under the name Bodo cephaloporus).
Caecitellus parvulus (Griessman, 1913) Patterson etaI., 1993. Fig7r.
Description: Length 4 /lm. Biflagellated cell widerthan long and with hoop of cytoskeletal material visible on one side of the flagella on the ventral surface.The anterior flagellum sweeps in front of the cell asit glides along on its posterior flagellum. Observedtwice from Hamelin Pool.
Remarks: Observations consistent with previous accounts by Griessmann [15] and Larsen and Patterson[22] under the name Bodo parvulus. The hoop wasnot observed in these studies, but was first identified
in an ultrastructural examination [38]. The hoop is relatively easy to observe and is distinctive.
Glissandra Patterson and Simpson, n. gen. Figs. 7 s,8 a-c.
Diagnosis: Gliding biflagellated protist of uncertainaffinities. Cell round in outline, measuring 5 - 8 /lm andwith two flagella inserting laterally into an anteriorventral groove and extending posteriorly and anteriorly. Glides with most of both flagella held againstthe substrate. Type species G. innuerende.
Glissandra innuerende Patterson and Simpson, n. sp.Figs. 7 s, a-c.
Diagnosis: Glissandra species 5 - 8 /lm long. Typemicrograph: Fig. 8 a.
Description: Gliding protist with almost sphericalbody measuring 5- 8 /lm in diameter, with two flagella, inserting laterally, side by side and subapically into asmall ventral groove. The groove extends to the front

margin of the cell being occasionally evident as a slightdiscontinuity. Flagella about equal in length and about2lj2 times body length. Both flagella lie against the substrate in gliding cells, with one directed anteriorly andthe other posteriorly. Only the tip of the anterior flagellum moves back and forward during gliding, with theremainder of the flagellum pointing stiffly in the direction of motion (similar to behaviour of the flagellumduring gliding in the euglenid genus Petalomonas). Cytoplasm often filled with small granules (Fig. 8 a) butno large food vacuoles were observed. Observed occasionally from Little Lagoon and Hamelin Pool.
Remarks: This species has certain similarities withgliding heterotrophic euglenids, cercomonads, andthaumatomonads. It can be distinguished from euglenids by the lack of a flagellar pocket and pellicle. Itis distinguishable from cercomonads by the presenceof the ventral groove into which the flagella insert,and it is distinguishable from Protaspis and similargenera (i.e. thaumatomonads) by the nature of thegroove into which the flagella insert, by the beat pattern of the anterior flagella, and by the side-by-side(and lateral) rather than anterio-posterior insertionof the flagella. Thaumatomonads also have a very prominent anterior nucleus [ego 22] while we were unableto observe the nucleus in Glissandra.
A single individual assignable to this species has previously been observed by one of us from Rio de Janeiro,Brazil (Patterson, unpubl.).
Metopion fluens (Larsen and Patterson, 1990). Figs.6z, 6~, 7t.
Description: Cell 6-8 /lm long, oval shaped and flattened. Close to one end of the cell is a crease on theventral side which offsets a small bulbous snout region. A single flagellum 11/2 - 2 times cell length insertsat the inside end of the crease. This flagellum appearsthick and terminates abruptly rather than tapering orending in a hair tip. The cell glides on this flagellumwith the snout pointing almost backwards and thebulk of the cell sticking out sideways on the oppositeside of the flagellum. The snout region may appearto contain many fine granules. Large inclusions andcoloured granules were observed in the posterior (i.e.opposite the snout) end in some cells. Observed fromLittle Lagoon.
Remarks: Metopion fluens appears similar to Ancyromonas sigmoides but is distinguished by the lack of aprominent ventral groove, the abrupt-ending ratherthan acronematic posterior flagellum, generally largersize and lack of any anterior flagellum (or apical flagellar depression) in all cells. When a second flagellum ispresent in Metopion it is directed posteriorly [22]. Nocells with a second flagellum were observed in thisstudy.
The presence of large inclusions and coloured fragments suggests that Metopion fluens is predatory andconsumes other protists. Actual feeding was not observed.
Benthic Heterotrophic Flagellates from W. Australia . 443
Metromonas simplex (Griessmann, 1913) Larsenand Patterson, 1990. Fig. 7u.
Description: Cell 5- 7 /lm long and dorso-ventrallyflattened. Cell in the broad aspect is round to pearshaped with the pointed end posterior-most. Anteriormargin of the cell usually thicker and more refractilethan the rest of the cell. Usually a single flagellum inserting at the posterior of the cell approximately 2-3xcell length. Cells are usually observed attached to thesubstrate by the distal portion of this flagellum. Thecell swings from side to side from this attachment likea pendulum. The cells may also be observed gliding onthe long flagellum with the cell body being held clear ofthe substrate. Sometimes the stub of a second flagellummay be observed right next the point of emergence ofthe major flagellum. Observed from Little Lagoon.
Remarks: Some cells were observed with colouredparticles inside which we regard as the remains of plastids from eukaryotic prey. This confirms other unpublished observations that this organism is a predator, inother words a consumer of protists. It typically appearsin preparations late in a succession and the increase innumbers is associated with a decline in the numbers ofother flagellates (see Ekebom et al. [8] for further comments). Metromonas simplex is one of the smallest flagellates for which predatory behaviour is known.
The pendulum behaviour of Metromonas distinguishes this genus from all other flagellates. Metromonas simplex may be distinguished from Metromonasgrandis (the only other species in the genus) by its smaller size and rounded rather than pointed (nippled) apical end in the former [22].
Palustrimonas Patterson and Simpson, n. gen. Figs.7w,8d-g.
Diagnosis: Heterotrophic, free-living, solitary protistwith two flagella inserting sub-apically into separateprominent grooves or pockets, one directed anteriorly, one posteriorly. Neither flagellum generates"reverse" thrust. Cell not highly flexible nor trulymetabolic. Cell does not glide. No prominent, permanent gullet but capable of consuming large items offood (i.e. other eukaryotes).
Palustrimonas yorkeensis (Ruinen, 1938) Pattersonand Simpson, n. gen., n. comb. Figs. 7 w, 8 d-g.
Description: Cell 8-14 /lm long, body not plasticand ovate to slender elliptical in profile. Anteriorend may be somewhat pointed. The flagella insert subapically and opposed. The anterior flagellum insertsinto a short anteriorly directed pocket. The posteriorflagellum inserts into a groove that extends for mostof the length of the cell. The groove is often an elongated tear-shape with the point posterior-most(Fig. 9 f). the anterior flagellum is approximatelyequal to the cell body in length and flails around theanterior end of the cell. The posterior flagellum is2-2.5 times cell length and may be held fairly stiffly

444 . D. J. Patterson and A. G. B. Simpson
or beats in slow waves. The posterior flagellum generally appears thicker than the anterior flagellum. Insome cells the posterior flagellum is missing. Swimming is often erratic and tumbling with progress in aconsistent direction quite slow. The cells do not glide.
Many cells had large colourless inclusions in the posterior end of the cell suggesting that P. yorkeensis ispredatory. In some cells the inclusions distended thecell sufficiently to obliterate the posterior portion ofthe groove. One cell was observed with an inclusionin the apical region which suggests that food may beingested at the anterior end of the cell. Prey capture/ingestion was not observed nor was any structure interpretable as any form of ingestion device.
Observed from the saturated puddle where it wascommon.
Remarks: The flagellate showed a certain similaritywith Ruinen's [42] description of Phyllomitus yorkeensis from Australian hypersaline waters. Her descriptionof cells from Bunbunga closely matches the organismwe observed in general cell shape (particularly withslender cells), relative lengths of the flagella, positionsand shape of the anterior flagellar pocket and bodygroove and the frequent presence of large, obvious posterior food vacuoles. Ruinen [42], however, indicatesthat the cells are deformable and that the posterior flagellum inserts into the anterior flagellar pocket. Ruinengives the size range as 14 - 22 11m, somewhat largerthan in our observations. We regard Ruinen's placement of the posterior flagellum as an error, noting thatthe ends of the groove and the anterior pocket are veryclose together. The other differences (body plasticity,body size) are potentially minor and we do not consider that we have grounds for separating the species.
Phyllomitus is a poorly defined and poorly circumscribed genus which, historically, has encompassed several very different types of flagellates [39]. The typespecies has apically inserting flagella which adhere together [48] while other more recently added species arehighly plastic sack-shaped with non-adhering flagella[e.g. 19,22,45]. Neither Ruinen's or our descriptionbears a particular resemblance to the type species,nor to any other species placed in Phyllomitus [39].Therefore the species should not remain in Phyllomitus. We have erected a new genus. Palustrimonas,for heterotrophic flagellates that are not highly plasticor amoeboid that have two opposed flagella that insertsubapically and close together into separate, distinctgrooves. Palustrimonas yorkeensis is the type and, currently, only species in the genus.
Palustrimonas shows some similarity to Colpodellaand Colponema. Both of these genera include predatory flagellates with 2 flagella inserting close together, but at a wide angle and not into the samedepression or groove. Colpodella has a dinstinctive division stage where the cell forms two or four daughtercells inside a cyst [7,43]. Although Colpodella divisioncysts were present in the same samples as Palustrimonas they all contained the coloured remains of algalprey and were assignable to Colpodella pugnax. The
separate flagellar pockets in all known species of Colpodella are very small to nonexistent compared tothose of Palustrimonas. Palustrimonas may be distinguished from Colponema because the latter is highlymetabolic and has a much more substantial groovethat extends to the extreme posterior of the cell [25].
Pleurostomum flab ellatum Ruinen, 1938. Figs. 7 x,8h-k.
Description: Cell 11-1411m long, spindle-shapedand round in transverse section with two parallel flagella inserting apically. The flagella are between 1112and 112 times body length. They are often, but not always, the same length. The flagella were usually observed stuck to the coverslip or slide. When free theflagella beat homodynamically. Subapically there is awide and deep opening or groove that runs half tothree quarters the way down the body in a half spiraland usually ends in close association with a foodvacuole. The body was not observed to bend or flexexcept in one individual where the posterior third ofthe cell was stuck to the coverslip. Observed occasionally from the saturated puddle.
Remarks: The genus Pleurostomum includes thoseheterotrophic protists with two parallel, homodynamic, apically or subapically inserting flagella, anda cytostome (cytostomal groove) with a broad, subapical opening [30,42]. Pleurostomum has only been observed from hypersaline environments [42].Namyslowski, when erecting the genus, describedthe species P. caudatum, P. salinum, P. parvulum andP. gracile [30]. Ruinen [42] added P. flabellatum basedon observations from India and Australia.
The species we observed displayed a mixture of characteristics from several of the previously describednominal species. The size range was similar toP. gracile but this species is described as being quiteflexible and always having flagella of different length[30, 42]. The rounded cross-section and stiff natureof the cell are similar to P. salinum but this specieshas been previously described as being about 20 11mlong and much more rod-shaped than most individualswe observed [30,42]. The body form in profile and thespiral cytostomal groove closely match Ruinen's drawing of P. flabellatum but this species is described as flattened and is distinguished by a distinctive spirallingmovement that we did not observe [42].
We regard our species is being most similar to andnot distinguishable from P. flabellatum. All the cellswe observed were stuck to glass surfaces and locomoting so we do not regard our lack of observation ofspiralling during motion as critical. Most of the cellswe observed were not noticeably flattened, howeverRuinen's drawings indicate that there is variation inthis feature [42]. Flatness may be linked to nutrition.We interpret the spiral fins observed by Ruinen [42]as the edges of the cytostomal groove, which may appear slightly raised (Fig. 9 j). Ruinen [42] states that thecytostome runs immediately beneath the fins. Ruinen

Table 1. Summary of the species observed from each location(darker shading represents species described for the first time)
... i:r ,g- ~(Il:ij r >l:W ~ Ea ~.
~ ~o ,
~ 2!>< - ?' (;l. '" -~ ft ~~r ~5" VI _.0" O-"E
~8 ~"O '"~ 8 ~"8 !:l 0-
~o=:0-5- .. " ~i>SD.
t- I-Alllas/igQIllO~IaSmtllabilis
f---AI/e/I/I/ia exotica
..' , ,Dillellla l'OlidulII ~ •• 1..... - '., .- - __ a
Dolilllll sedel//(Iriwll '!I~'l ,".'i.
-_. ----He/erOl/elllll e.tara/1I111 l •.."" ~ ., -'\-"
-- - -Massis/eria lIIari/1a l
I .. - ------ .. - - -Me/opioll f/llells ','".. " - -- ---Me/romollas silllp/ex • .~ (j
",' -t- .-N%so/eIlIlS apocalllp/lis
~ • LIII 1'· - -
N%so/elll/s ClIllella/IiS - . l~ -_.- '- --NOlosolellllS os/il/III f..:: ~ "I- -----
• _ ~"9 t •Peral/elllll fusi/onlle 'J, ._-- .~--
Pe/alolllo/las abscissa "~_i •,. ' ' --Pe/a/olllo/IaS mil/II/a .. :-,.".
-- ---- ---1"'-"·1
Pe/alomo/las raiu/a .£ l, ... ·
1---1-.
P/oeo/ia aZl/rillo II. sp. --P/oeo/ia cfr. p,mc/ala ,r. r ..
Ploeolia corruga/o "-
_ l>.:::....- ---.
P/oeOlia cosla/o '. - -',. ... .
P/oeo/la ob/ol/go ,. , .- -- -_. - -~/ia p/lll11osa - /.
-. ==t-- - -Ploeo/ia robus/ll -Ploeo/io \'il rell • r i ~ ~~. I .Psel/dobodo /remu/al/s ~ , I.
,';-' l~. - --Allcyrolllol/as sigllloides '~. ~4 _
Bodo desigl/is " • .. ---Bordl/lIl11o/1aS /ropicaJlll - .I l
Glissalldra il/lll/erel/de 1/. sp.- ~ -GOlllomol/as pacifica I ..
- - - -1---Pe/a/olllollas poosilla .-Ploeo/ia pseucUlllisol/ellla .~ .-Bodo saliells " ,I" •• -RhYlle/lOlllo/IaS 1I0Sllta .. l, I
Caeci/elhlS parvulus ;, Ci;. ~"
Cofe/eria ligl/li/era -Cafe/erio roellbergensis "
Developoyella elega/IS ,--- ----
Al/c)'rOlI/OIIlIS lIIelba II. sp.
Co/potlella IIl/gl/is 1/. sp.
RhYllcl/obodo simills II. sp. IBodo ra//(lIIs
..J ..•
,.Bodo eyglllls /I .sp. -Co/podellll p"gl/ax . ..
~illlol/as .yorkeel/sls -Plel/ros/OIIl/lIl/jlabel/a!!!!!!..... t-.- - . .. ...Pleuros/olllulII /lJrJlidllllll/. SIJ·
Benthic Heterotrophic Flagellates from W. Australia . 445
describes P. flabellatum as 16-30 /-lm long, larger thancells we observed but not sufficiently so, in our opinion,to warrant species level separation [see 22]. The longercells observed by Ruinen may refer to individualswhere the end was drawn out to a long, thin thread[42]. Further work is required to determine speciesboundaries within this poorly known genus.
Pleurostomum turgidum Patterson and Simpson, n.sp. Figs. 7 y, 81-n.
Diagnosis: Pleurostomum 6-9/-lm long with ovalshaped body. Flagella insert subapically, are laterallydirected and are both more than two times bodylength. Type micrograph: Fig. 81.
Description: Cell 6 - 9 /-lm long. The cell is more thanhalf as wide as long with a somewhat pointed anteriorend. The broad, shallow, spiral body groove extendsfrom an immediately subapical beginning to abouthalf way down the cell and may end in close association with a food vacuole. One edge of the groove projects out from the cell somewhat to give the appearanceof a rounded bulge about a quarter of the way downfrom the anterior end. The flagella insert subapicallyin a slight depression and are laterally directed. Bothflagella are 2112 to 3112 times body length with one somewhat longer than the other. The ends of the flagellausually stuck to the coverslip during observation.The flagella beat isodynamically down the body andinto the groove. The beat is slow and pronounced.When subjected to the flash of the camera system,the flagella may reverse to point ahead of the cell. Position of the nucleus not known. Observed occasionallyfrom the saturated puddle.
Remarks: Previously described species of Pleurostomum are all slender organisms with apically insertingflagella up to two times body length [30,42]. Pleurostomum turgidum may be distinguished from all otherspecies in the genus (see above) by the very long flagella, the subapical insertion of the flagella, the lateraldirection of the flagella and the fatness of the cell.The anterior end of the cytostomal groove is not asdeep and obvious as in P. flabellatum.
Unidentified flagellates
The species list given above is not exhaustive. Wehave observed a number of other heterotrophic flagellates, but sufficiently infrequently as to emerge with aclear concept of their identities. These include: i) a colourless species of Plagiodinium-like dinoflagellateabout 45 /-lm long with a small peg-like epicone anda fine theca, from Little Lagoon, ii) an Amphidiniopsis-like dinoflagellate measuring 50/-lm with adimpled theca with pores and with bluegreen algalsymbionts, from Little Lagoon, iii) several specimensof a fast moving, tiny dinoflagellate, 3 - 5 /-lm longwith a spiral girdle, from Hamelin Pool and the hypersaline pond, iv) two individuals of a small, non-loricatechoanoflagellate from Hamelin Pool, v) an individualfrom a species of Bicoscoeca from Hamelin Pool, vi) a

446 . D. J. Patterson and A. G. B. Simpson
swimming stramenopile 6- 8 f.lm long with flagella inserting a third of the way down the cell and, vii) a singleslowly gliding flagellate 10 f.lm long, somewhat reminiscent of Glissandra but with a thinly oval body, fromLittle Lagoon.
Discussion
This study is part of a series directed at understanding large scale patchiness of marine heterotrophic flagellates in Australia [8,22, 54] and elsewhere [22, 36,55,56,57].
Comparisons of communities for biogeographicalpurposes rely on a taxonomic base. We have made observations during this study which extend the range ofcharacteristics attributable to various species of flagellates. We have chosen not to emend formally descriptions for two reasons. The first is that emendeddescriptions are better left to a dedicated taxonomicrevision - and such a revision is not the objective ofthis study. Secondly, the taxonomy of free-living heterotrophic flagellates is very incomplete and furtherchanges are highly probable. A revision should occurafter more stability has been achieved.
This series of studies is designed to segregate intrinsicand extrinsic factors which may contribute to conclusions about biogeography. The exclusion of taxa willtend to exaggerate the perceived cosmopolitanism offlagellate communities. We have therefore included organisms with a distinctive identity even if that identityis based on relatively few observations, and we havesought to be explicit in our reasoning for lumping orsplitting.
The significant majority of the species observed inthis study are not exclusive to Australia, and thus wehave little evidence for endemism. All species observedfrom Little Lagoon have been recorded previously fromat least one other site in the world [8,22]. This statement embraces the newly described species Ploeotiaazurina and Glissandra innuerende which have previously been noted from the Great Barrier Reef andBrazil (respectively) but descriptions have not previously been published (see species descriptionsabove). Three species (Ploeotia azurina, Ploeotia plumosa and Anehmia exotica) have, to date, only beenrecorded from Australian sites. As none of the catalogues of heterotrophic flagellates from any site may beguaranteed as comprehensive, further studies elsewhere may well reveal that the three species have amore extensive geographic distribution. Consequently, we are not confident that these species are endemic to Australia.
When the communities from different sites withinthe Shark Bay area are compared, there is an apparentreduction in diversity of flagellates with each incrementof salinity. The community from Hamelin Pool (aboutdouble the salinity of seawater) was largely a subset ofthe organisms observed from marine sites using thesame techniques with 12 out of the 17 species observed
from here also seen from "normal" marine sites [thisstudy, 8,22]. By contrast, only two of the seven speciesrecorded from the hypersaline pond (over four timesthe salinity of seawater) have been recorded from nor:mal marine benthic sites [this study, 8,22]. None of thefive species found in the saturated puddle have beenpreviously recorded from marine sediments, and two(Pleurostomum flabellatum, Palustrimonas yorkeensis) have been observed previously exclusively from hypersaline habitats [30, 42]. In addition, Ancyromonasmelba, Bodo cygnus, Colpodella unguis and Pleurostomum turgidum are described for the first time in thisstudy and have yet to be reported from marine or freshwater habitats. As one of these species (A. melba) hassince been observed in Antarctic material (see "remarks" to A. melba), only some of these species maybe restricted to hypersaline environments.
The apparently extreme halotolerance of some flagellate species observed in this study is remarkablewhen compared to that displayed by even the most euryhalilne metazoa or metaphyta. Some species (rhynchomonas nasuta, Bodo saliens) observed from themarine Little Lagoon site were also present in the150%0 salt (i.e. more than 4 times as concentrated asnormal sea water) environment of the hypersalinepond. Assuming that the organisms have been correctly identified. Bodo saltans exhibits the greatest possible range of halotolerance, occurring in freshwater tosaturated brine habitats. Further research on specieslevel taxonomy is desirable in order to confirm this because the viscosity and other attributes of concentratedbrine may affect the organisms and lead to inaccurateidentifications.
Acknowledgements
The authors would like to thank Dr J. Larsen, Dr S. Tong, F.Ekelund, the subscribers to Taxacom and in particular twoanonymous reviewers for helpful discussion and comments,B. Dicks for darkroom work and expertise, the DenhamCALM office, Dr B. Burne and the Denham police for adviceand assistance. This study was supported by grants from theAustralian Biological Resources Study (ABRS) and from theAustralian Research Council (ARC).
References
1 Alongi D. M. (1991): Flagellates of benthic communities:characteristics and methods of study. In: Patterson D. J.and Larsen J. (eds.) the biology of free-living-heterotrophic flagellates, PP' 57- 75. Systematics Associationspecial volume No. 45, Oxford.
2 Bak R. P. M. and Nieuwland G. (1989): Seasonal fluctuations in benthic protozoan populations at different depthsin marine sediments. Neth. J. Sea Res., 24, 37-44.
3 Brooker B. E. (1971): Fine structure of Bodo saltans andBodo caudatus (Zoomastigophora: Protozoa) and theiraffinities with the Trypanosomatidae. Bull. Brit. Mus.Nat. Hist. (Zoo!.), 22, 89-102.

4 Brugerolle G. (1985): Des trichocystes chez les bodonides,un charactere phylogimetique supplementaire entre Kinetoplastida et Euglenida. Protistologica, 21, 339-348.
5 Brugerolle G. and Mignot].-P. (1979): Observations sur Iecycle, I'ultrastructure et la position systematic de Spiromonas perforans (Bodo perforans Hollande, 1938), flagelle parasite de Chilomonas paramecium: ses relationsavec les dinoflagelles et sporozoaires. Protistologica,15, 183-196.
6 Burzell L. A. (1973): Observations on the proboscis-cytopharanx complex and flagella of Rhynchomonas metabolita Pshenin, 1964 (Zoomastigophora: Bodonidae). ].Protozoo!., 20, 385 -393.
7 Cienkowski L. (1965): Beitrage wr Kenntnis der Monaden. Arch. Mikroscop Anatom., 1, 202-232.
8 Ekebom]., Patterson D.]., and Vars N. (1996): Heterotrophic flagellates from coral reef sediments (Great Barrier Reef, Australia). Arch. Protistenkd. 146, 251-272.
9 Farmer M. and Triemer R. E. (1988): A redescription ofthe genus Ploeotia Duj. (Euglenophyceae). Taxon, 37,319-325.
10 Farmer M. and Triemer R. E. (1994): An ultrastructuralstudy of Lentomonas applanatum (Preisig) N. G. (Euglenidal. J. Euk. Microbio!., 41,112-119.
11 Fenchel T. (1986): The ecology of protozoa. Springer-Verlag, Berlin.
12 Fenchel T. and Patterson D. ]. (1988): Cafeteria roenbergensis nov. gen., nov. sp., a heterotrophic microflagellatefrom marine plankton. Mar. Microbial Food Webs, 3,919.
13 Foissner W. and Foissner I. (1984): First record of an ectoparasitic flagellate on ciliates: an ultrastructural investigation of the morphology of Spiromonas gonderi nov.spec. (Zoomastigophora, Spiromonadidae) invading thepellicle of ciliates of the genus Colpoda (Ciliophora, Colpodidae). Protistologica, 20, 635-648.
14 Gajadar A. A., Marquardt W. c., Hall R., Gunderson J.,Ariztia-Carmona E. V. and Sogin M. L. (1991): Ribosomal RNA sequences of Sarcocystis muris. Theileria annulata and Crypthecodinium cohnii reveal evolutionaryrelationships among apicomplexans, dinoflagellates,and ciliates. Mo!. Biochem. Parasito!., 45, 147-154.
15 Griessmann K. (1913): Ober marine Flagellaten. Arch.Protistenk., 32, (year 1914), 1-78.
16 Hanel K. (1979): Systematik und Okologie der farblosenFlagellaten des Abwassers. Arch. Protistenkd., 121, 73137.
17 Hollande A. (1938); Bodo perforans n. sp., flagelle nouveau parasite externe du Chilomonas paramecium Ehrenb. Arch. Zoo!' Exp. Gen., 79, 75-81.
18 Hollande A. (1942): Etude cytologique et biologique dequelques flagelles libres. Volvocales, Cryptomonadines,Eugleniens, Promastigines. Arch. Zoo!' Exp. Gen., 83,1-268.
19 Klebs G. (1892): Flagellatenstudien. Zeitschr. Wiss. Zoo!.,55, 265 -445.
20 Lackey J. B. (1940); Some new flagellates from the WoodsHole area. Am. Mid!. Nat., 23, 463-471.
21 Larsen]. (1987): Algal studies of the Danish Wadden Sea.IY. A taxonomic study of the interstitial euglenoid flagellates. Nord. ]. Bot., 7,589-607.
22 Larsen J. and Patterson D. ]. (1990): Some flagellates(Protista) from tropical marine sediments. ]. Nat. Hist.,24, 801-937.
23 Lemmermann E. (1913): Pantostomatinae, Promastiginae, Distomatinae. In: Pascher A (ed.): Die
Benthic Heterotrophic Flagellates from W. Australia· 447
SiiBwasserflora Deutschlands, Osterreich und derSchweiz, I, pp. 115 -174. Jena.
24 Logan B. W. and Cebulski E. (1970): Sedimentary environments of Shark Bay, Western Australia. Am. Assoc.Pet. Geo!. Mem., 13, 1-37.
25 Mignot ].-P. and Brugerolle G. (1975): Etude ultrastructurale du flagelle phagotrophe Colponema loxodes Stein.Protistologica, 11, 429-444.
26 Mylnikov A. P. (1983): [Feeding of predatory zooflageilates.] Information Bulletin Biologiya Vnutrennikh Vod,62, 16-19 (In Russian).
27 Mylnikov A. P. (1986): Ultrastructure of a colourless flagellate, Phyllomitus apiculatus Skuja 1948 (Kinetoplastidal. Arch. Protistenkd., 132, 1-10.
28 Mylnikov A. P. (1990): [Characteristic features of the ultrastructure of the colourless flagellate, Heteromita sp.]Tsitologiya, 32, 567-571 (In Russian).
29 Mylnikov A. P. (1991): [Ultrastructure and biology of certain representatives of the order Spiromonadida (Protozoa)]. Zoologicheskiy Zhurnal, 7, 5-15 (In Russian).
30 Namyslowski B. (1913): Ober unbekannte halophileMikroorganismen aus dem Innern des SalzbergwerkesWieliczka. Bull. Int. Aced. Sci. Krakow, Series B, 3/4,88-104.
31 Novarino G., Warren A., Kinner N. E., and Harvey R. W.(1994): Protists from a sewage contaminated aquifer onCape Cod, Massachusetts. Geomicrobio!., 12, 23-36.
32 Patterson D. ]. (1982): Photomicrography using a dedicated electronic flash. Microscopy 34,437-442.
33 Patterson D. ]. (1994): Protozoa: evolution and systematics. In: Hausmann, K. and Hiilsmann N. (eds.): Progressin Protozoology (Proceedings of the IX International congress of Protozoology, Berlin, 1993), pp. 1-14. GustavFischer Verlag, Stuttgart.
34 Patterson D. ]. and Fenchel T. (1985): Insights into theevolution of heliozoa (Protozoa, Sarcodina) as providedby ultrastructural studies on a new species of flagellatefrom the genus Pteridomonas. Bio!. ]. Linn. Soc., 34,381-403.
35 Patterson D.]. and Fenchel T. (1990): Massisteria marinaLarsen and Patterson 1990, a widespread and abundantbacterivorous protist associated with marine detritus.Mar. Eco!. Prog. Ser., 62, 11-19.
36 Patterson D.]. and Larsen]. (1991): Nomenclatural problems with protists. Regnum Vegetabile, 123, 197-208.
37 Patterson D. ]., Larsen]. and Corliss J. O. (1989): Theecology of heterotrophic ciliates and flagellates living inmarine sediments. Progress in Protistology, 3, 185-277.
38 Patterson D. ]., Nygaard K., Steinberg G., and Turley C.(1993): Heterotrophic flagellates and other protists associated with oceanic detritus throughout the water columnin the mid North Atlantic. ]. Mar. Bio!. Ass. UK, 73, 6795.
39 Patterson D.]. and Z6lffel M. (1991): Heterotrophic flagellates of uncertain taxonomic position. In: Patterson D.]. and Larsen]. (eds.): The biology of free-living-heterotrophic flagellates, pp. 427-475. Syst. Ass. special vol.No. 45, Oxford.
40 Post F. ]., Borowitzka L. ]., Borowitzka M. A., Mackay B.and Moulton T. (1983): The protozoa of a Western Australian hypersaline lagoon. Hydrobiologia, 105, 95 -114.
41 Preisig H. (1979): Zwei neue Vertreter der farblosen Euglenophyta. Schweiz. Z. Hydro!., 41, 155 -160.
42 Ruinen]. (1938): Notizen iiber Salzflagellaten. II. Oberdie Verbereitung der Salzflagellaten. Arch. Protistenkd.,90, 210-258.

448 . D. J. Patterson and A. G. B. Simpson
43 Simpson A. G. B. and Patterson D. J. 1996: Ultrastructureand identification of the predatory flagellate Colpodellapugnax Cienkowski (Apicomplexa) with a descriptionof Colpodella turpis (n. sp.) and a review of the genus.Syst. Parasito!' 33, 181-198
44 Skuja H. (1939): Beitrag zur Algenflora Lettlands II. ActaHorti Bot. Univ. Latviensis, 11-12,41-169.
45 Skuja H. (1948): Taxonomie des Phytoplanktons einigerSeen in Uppland, Schweden. Symbolae Botanicae Upsalienses, 9, 5-399.
46 Skuja H. (1956): Taxonomische und Biologische Studiendas Phytoplankton Schwedischer Binnengewasser. NovaActa Regiae Societatis Scientiarum Uppsaiensis, 4th series, 16, 1-403.
47 Skvortzkov B. W. (1957): New and rare Flagellata fromManchuria, Eastern Asia. Phillipine Journal of Science,86, 139-202.
48 Stein F. R. (1978): Der Organismus der Infusionsthiere.III. Der Organismus der Flagellaten I. Leipzig.
49 Steinberg c., Lenhart B. and Klee R. (1983): Bemerkungen zur Okologie eines farblosen Phytoflagellaten Phyllomitus apiculatus Skuja (1948), (Cryptophyceae). Arch.Protistenk., 127, 307-317.
50 Swale E. M. S. (1973): A study of the colourless flagellateRhynchomonas nasuta (Stokes) Klebs. Bio!. J. Linn. Soc.,5,255-264.
51 Throndsen J. (1969): Flagellates of Norwegian coastalwaters. Nytt Magasin for Botanikk, 16, 161-216.
52 Triemer R. E. (1986): Light and electron microscopic observation of a colourless euglenoid, Serpenomonas costatan. g. n. sp. J. Protozoo!., 33, 412-415.
53 Tong S. M. (1995): Developayella elegans nov. gen., nov.spec., a new type of heterotrophic flagellate from marineplankton. Europ. J. Protisto!., 31,24-31.
54 Tong S. M. (in press): Heterotrophic flagellates from thewater column in Shark Bay, Western Australia. Mar. Bio!.
55 V0rs N. (1992a): Heterotrophic amoebae, flagellates andheliozoa from the Tvarminne area. Gulf of Finland, in1988-1990. Ophelia, 36, 1-109.
56 V0rs N. (1992b): Heterotrophic protists (excl. dinoflagellates, loricate choanoflagellates, and ciliates). In: Thomsen H. A. (ed.): Plankton from inner Danish Waters.An analysis of the autotrophic and heterotrophic plankton in Kattegat. HAV 90 Rapport. Danish NationalAgency for Environmental Protection [in Danish, withspecies lists and illustrations]. Havsforskning fraMilj0styrelsen, 11, 195-246.
57 V0rs N. (1993): Heterotrophic amoebae, flagellates andheliozoa from arctic marine waters (North West Territories Canada and West Greenland). Polar Bio!., 13,113-126.
58 Zhang, Y., Yang G.-T. and Mu L.-Q. (1993): New flagellatae from Harbin, China. Bul!. Bot. Res., 13, 113-117.
Key words: Amastigomonas - Ancyromonas - Ancyromonas melba n. sp. - Anehmia - Bodo - Bodo cygnus n. sp.- Bordnamonas - Caecitellus - Cafeteria - Ciliophrys - Colpodella - Colpodella unguis n. sp. - Developayella Dinema - Dolium - flagellate - Glissandra n. gen. - Glissandra innuerende n. sp. - Goniomonas - Heteronema hypersalinity - marine microbial communities - Massisteria - Metopion - Metromonas - Notosolenus - Palustrimonas n. gen. - Peranema - Petalomonas - Pleurostomum - Pleurostomum turgidum n. sp. - Ploeotia - Ploetiaazurina n. sp. - Pseudobodo - PRIMER - Pteridomonas - Rhynchobodo simius n. sp. - Rhynchomonas - sediments - taxonomy
D. J. Patterson and A. G. B. Simpson, School of Biological Sciences, A08, University of Sydney, NSW 2006, Australia