heterochromatin darkly stained and condensed transcriptionally silent and silences adjacent genes...
TRANSCRIPT
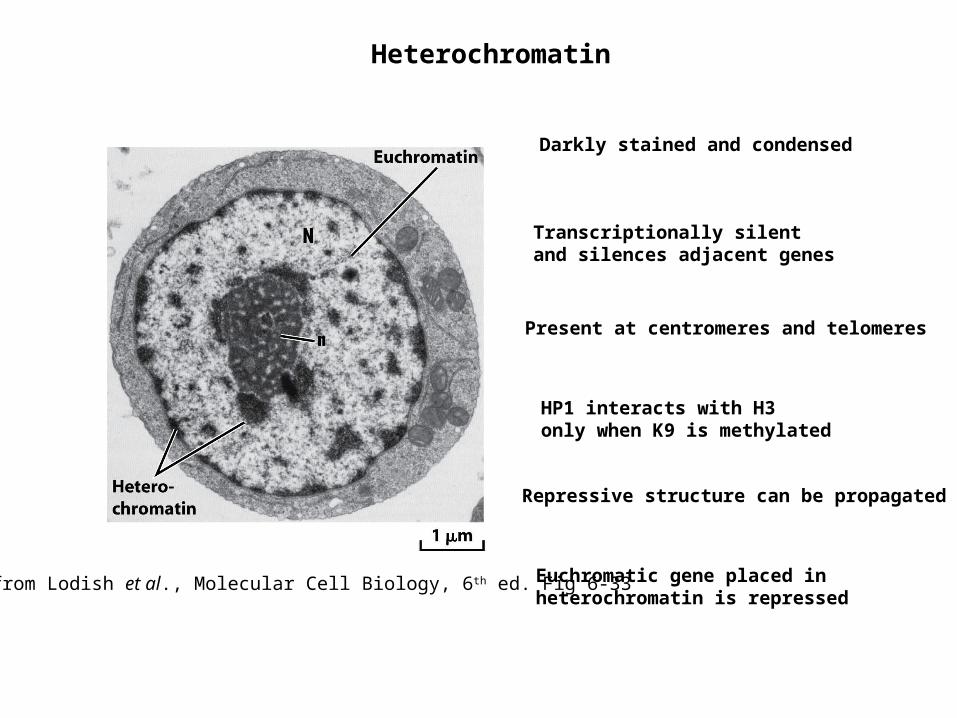
Heterochromatin
Darkly stained and condensed
Transcriptionally silent and silences adjacent genes
Present at centromeres and telomeres
HP1 interacts with H3 only when K9 is methylated
Repressive structure can be propagated
Euchromatic gene placed in heterochromatin is repressed
from Lodish et al., Molecular Cell Biology, 6th ed. Fig 6-33

Histone Modifications Associated with Heterochromatin and Euchromatin
from Lodish et al., Molecular Cell Biology, 6th ed. Fig 6-33
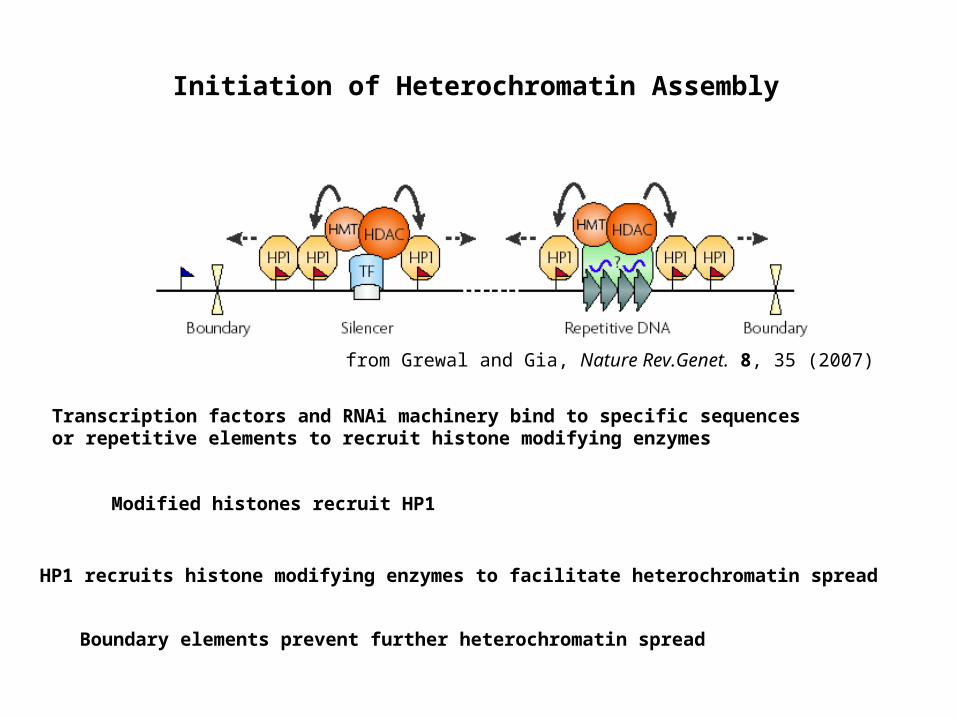
Initiation of Heterochromatin Assembly
from Grewal and Gia, Nature Rev.Genet. 8, 35 (2007)
Transcription factors and RNAi machinery bind to specific sequences or repetitive elements to recruit histone modifying enzymes
Boundary elements prevent further heterochromatin spread
HP1 recruits histone modifying enzymes to facilitate heterochromatin spread
Modified histones recruit HP1
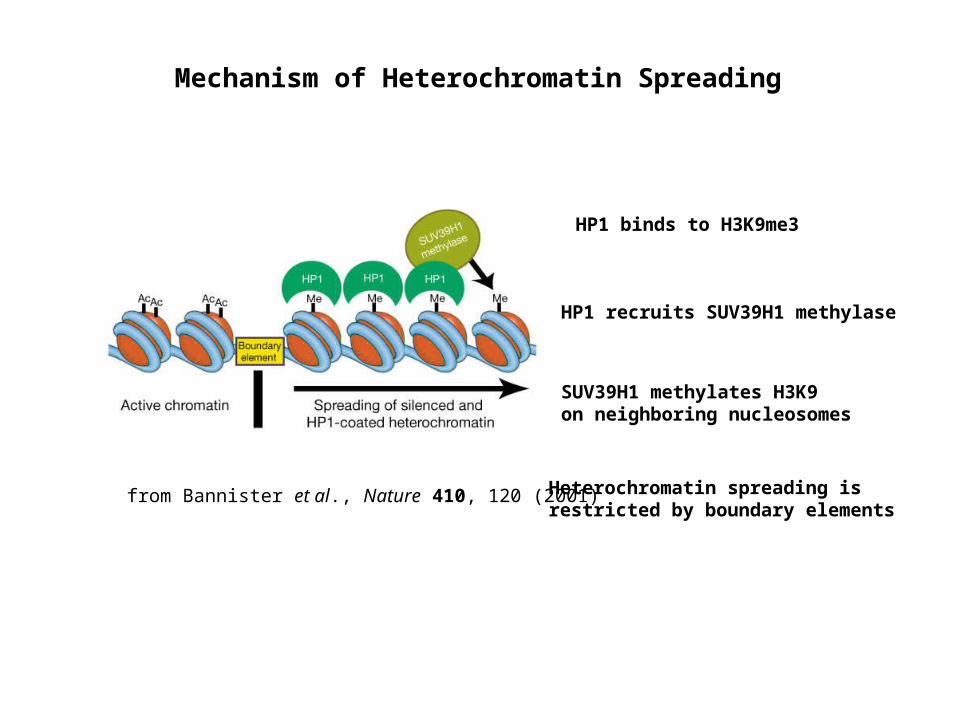
from Bannister et al., Nature 410, 120 (2001)
Mechanism of Heterochromatin Spreading
HP1 binds to H3K9me3
HP1 recruits SUV39H1 methylase
SUV39H1 methylates H3K9 on neighboring nucleosomes
Heterochromatin spreading is restricted by boundary elements
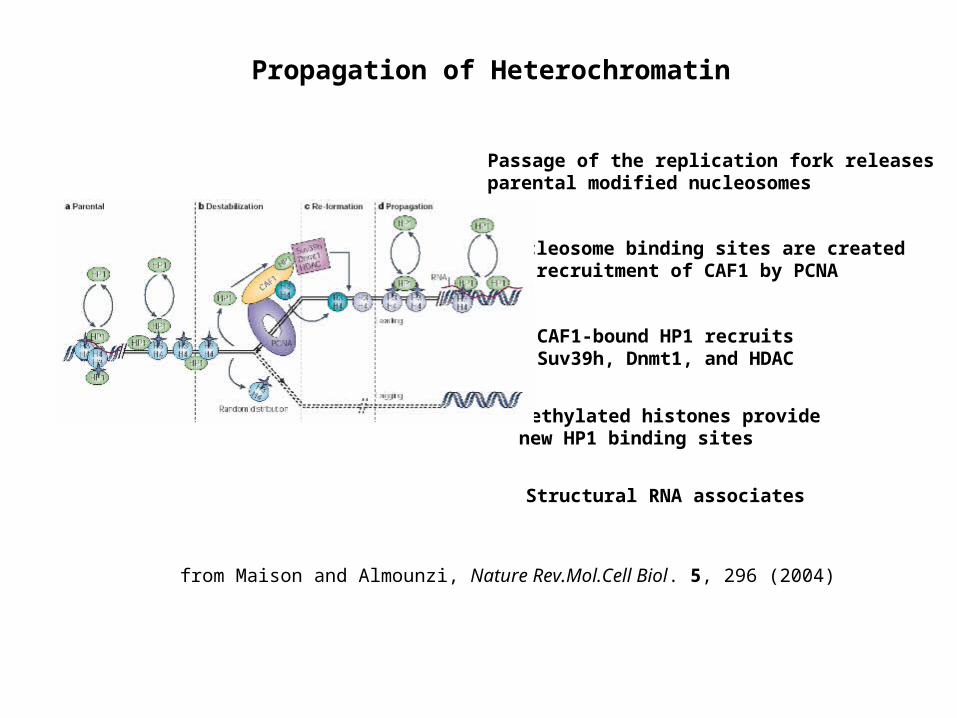
Propagation of Heterochromatin
from Maison and Almounzi, Nature Rev.Mol.Cell Biol. 5, 296 (2004)
Passage of the replication fork releases parental modified nucleosomes
Nucleosome binding sites are created by recruitment of CAF1 by PCNA
CAF1-bound HP1 recruits Suv39h, Dnmt1, and HDAC
Methylated histones provide new HP1 binding sites
Structural RNA associates
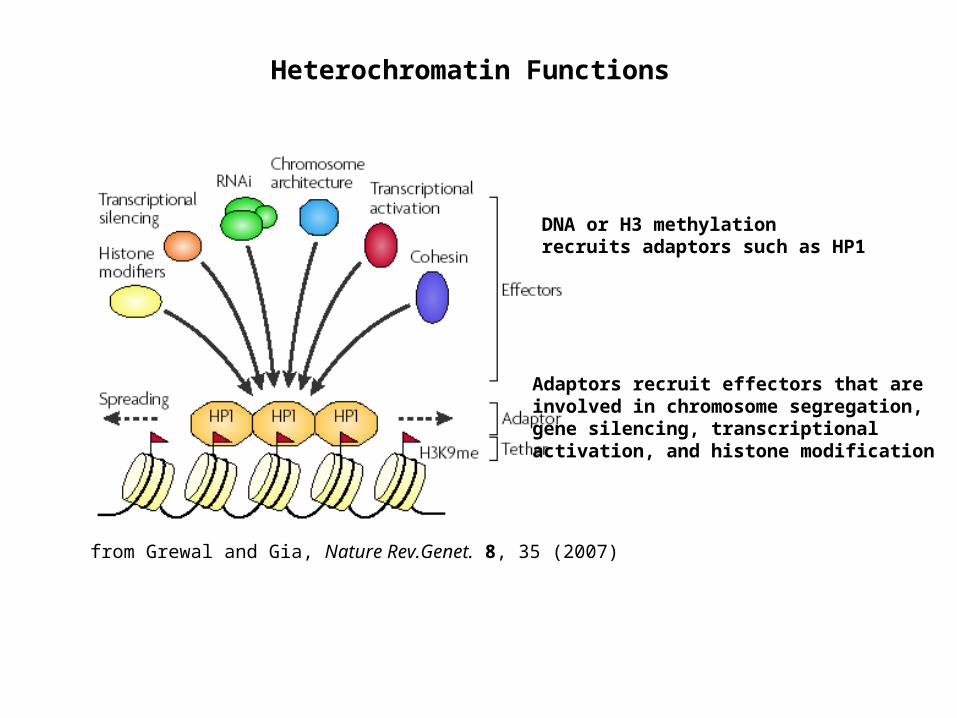
from Grewal and Gia, Nature Rev.Genet. 8, 35 (2007)
Heterochromatin Functions
DNA or H3 methylation recruits adaptors such as HP1
Adaptors recruit effectors that are involved in chromosome segregation, gene silencing, transcriptional activation, and histone modification
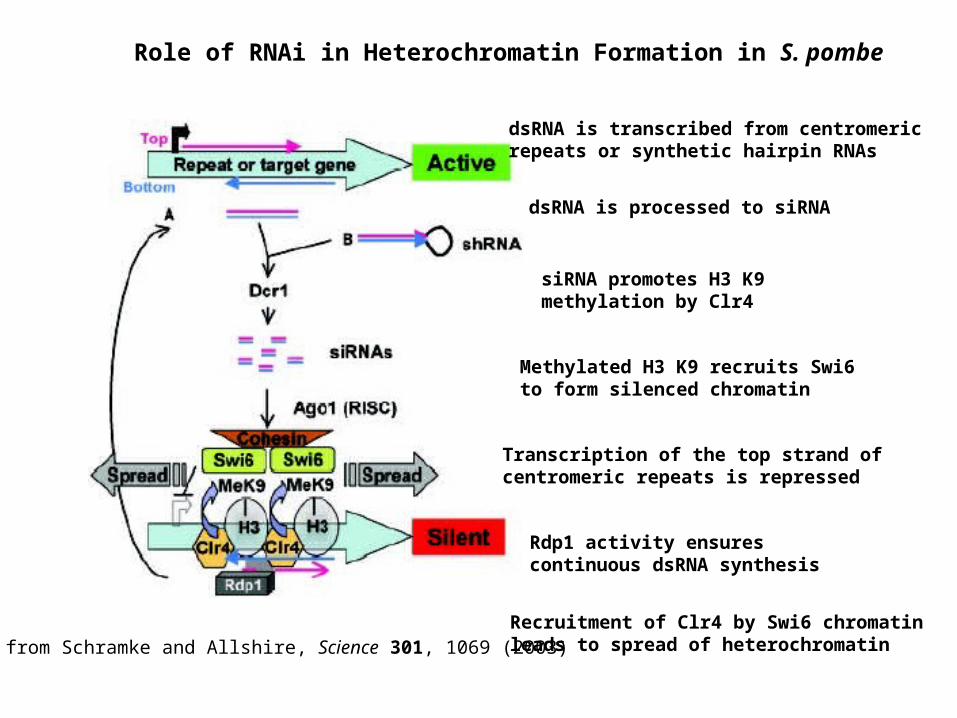
Role of RNAi in Heterochromatin Formation in S. pombe
dsRNA is transcribed from centromeric repeats or synthetic hairpin RNAs
dsRNA is processed to siRNA
siRNA promotes H3 K9 methylation by Clr4
Methylated H3 K9 recruits Swi6 to form silenced chromatin
Transcription of the top strand of centromeric repeats is repressed
Rdp1 activity ensures continuous dsRNA synthesis
Recruitment of Clr4 by Swi6 chromatin leads to spread of heterochromatinfrom Schramke and Allshire, Science 301, 1069 (2003)
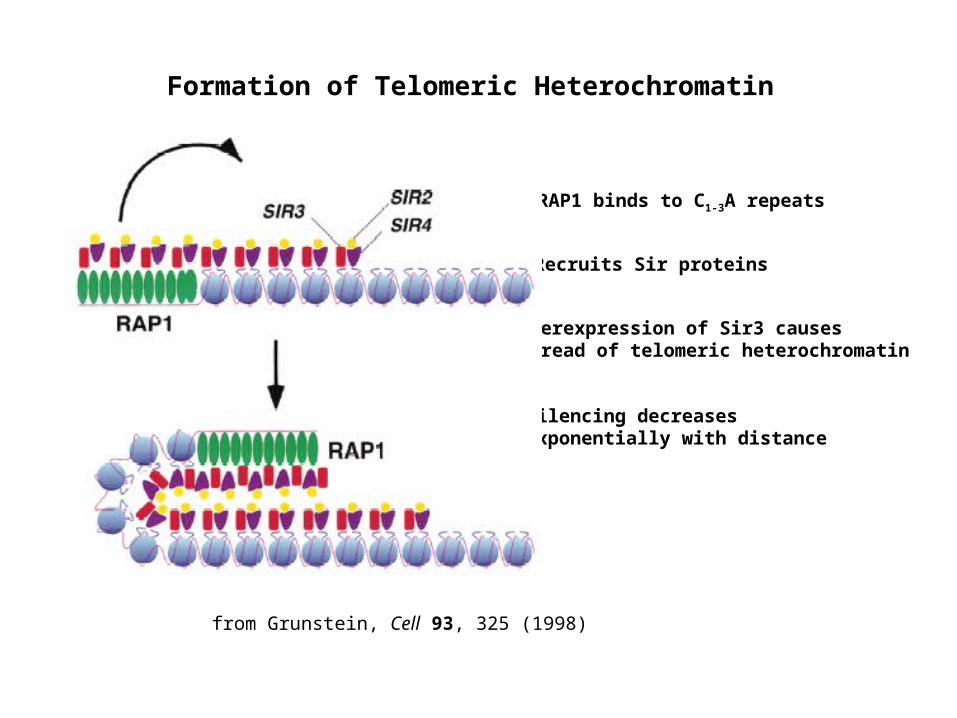
Formation of Telomeric Heterochromatin
from Grunstein, Cell 93, 325 (1998)
RAP1 binds to C1-3A repeats
Recruits Sir proteins
Overexpression of Sir3 causes spread of telomeric heterochromatin
Silencing decreases exponentially with distance
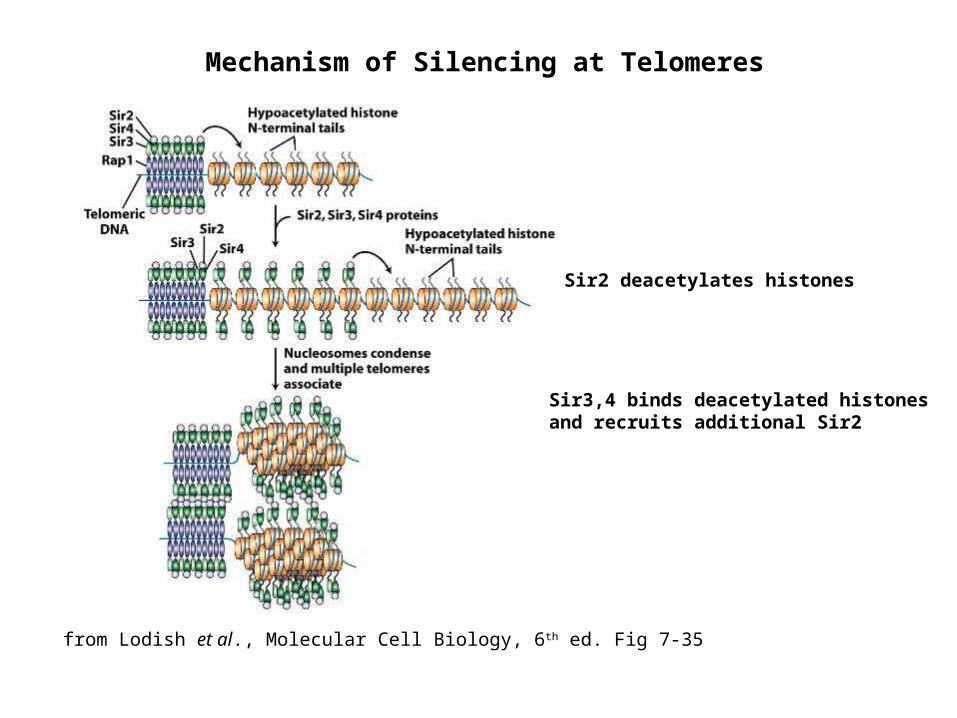
Mechanism of Silencing at Telomeres
Sir2 deacetylates histones
Sir3,4 binds deacetylated histones and recruits additional Sir2
from Lodish et al., Molecular Cell Biology, 6th ed. Fig 7-35

Insulators Prevent the Progression of Condensed Chromatin
from West et al, Genes Dev. 16, 271 (2002)
Insulators protect genes from inappropriate signals
Insulators block the action of distal enhancers
Insulators prevent the spreading of heterochromatin

gypsy Retrotransposon Contains an Insulator
gypsy protects a transgene from position effects
su(Hw) is necessary for enhancer blocking activity
gypsy contains a su(Hw) binding site
su(Hw) blocks the process that brings enhancer and promoter together
Formation of insulator bodies at the nuclear periphery to divide the chromosome into looped domains
Multiple su(Hw) binding sites can inhibit enhancer blocking activity
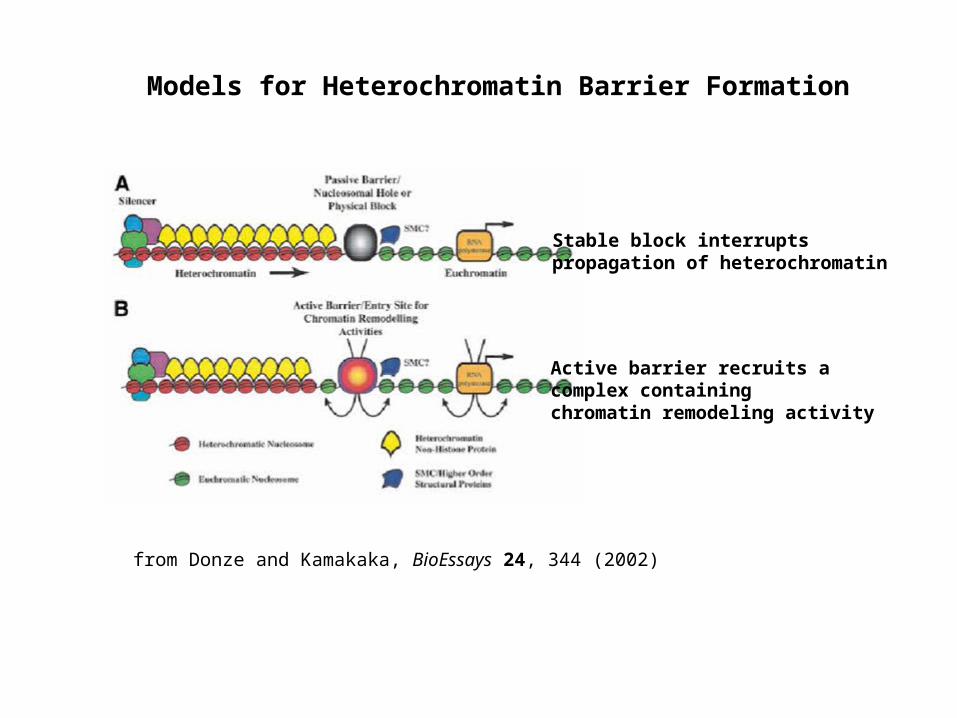
Models for Heterochromatin Barrier Formation
from Donze and Kamakaka, BioEssays 24, 344 (2002)
Stable block interrupts propagation of heterochromatin
Active barrier recruits a complex containing chromatin remodeling activity

Epigenetics
Heritable changes in gene function that cannot be explained by changes in gene sequences
DNA methylation
Nucleosome positioning
Histone variants and modifications

Epigenetic Modifications During Development
Epigenetically imposed restrictions to plasticity are erased in the germ line
Early mammalian development is characterized by progressive restriction of cellular plasticity accompanied by acquisition of epigenetic modifications
Epigenetic modifications impose a cellular memory that accompanies and enables stable differentiation
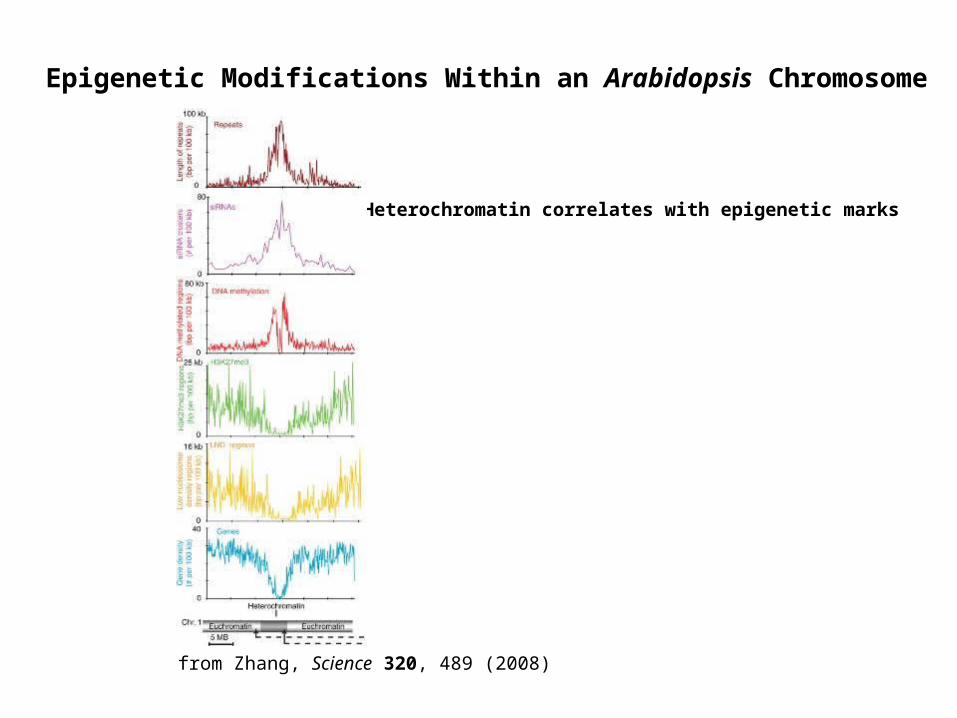
from Zhang, Science 320, 489 (2008)
Epigenetic Modifications Within an Arabidopsis Chromosome
Heterochromatin correlates with epigenetic marks

DNA Methylation
Methylation at CpG residues
Sites of methylation
Inactive X
Imprinted loci
Transposon-derived sequences
CpG islands and CpG island shores
Methylation patterns are reproduced at each round of cell division

Methylated CpG Islands Inhibit Transcription
Promoters are usually unmethylated
More than half of human promoters contain CpG islands
Methylated DNA recruits methyl-CpG-binding domain proteins which recruit histone modifying and chromatin-remodelling complexes
Unmethylated CpG islands recruit Cfp1 which associates with a histone methyltransferase creating H3K4me3
from Portela and Esteller, Nature Biotechnol. 28, 1057 (2010)

Methylated CpG Islands Inhibit Transcription
Promoters are usually unmethylated
More than half of human promoters contain CpG islands
Methylated DNA recruits methyl-CpG-binding domain proteins which recruit histone modifying and chromatin-remodelling complexes
Unmethylated CpG islands recruit Cfp1 which associates with a histone methyltransferase creating H3K4me3
from Portela and Esteller, Nature Biotechnol. 28, 1057 (2010)

Methylation of Repetitive Sequences Stabilize Chromosomes
Unmethylated repetitive sequences cause reactivation of endoparasitic sequences
from Portela and Esteller, Nature Biotechnol. 28, 1057 (2010)
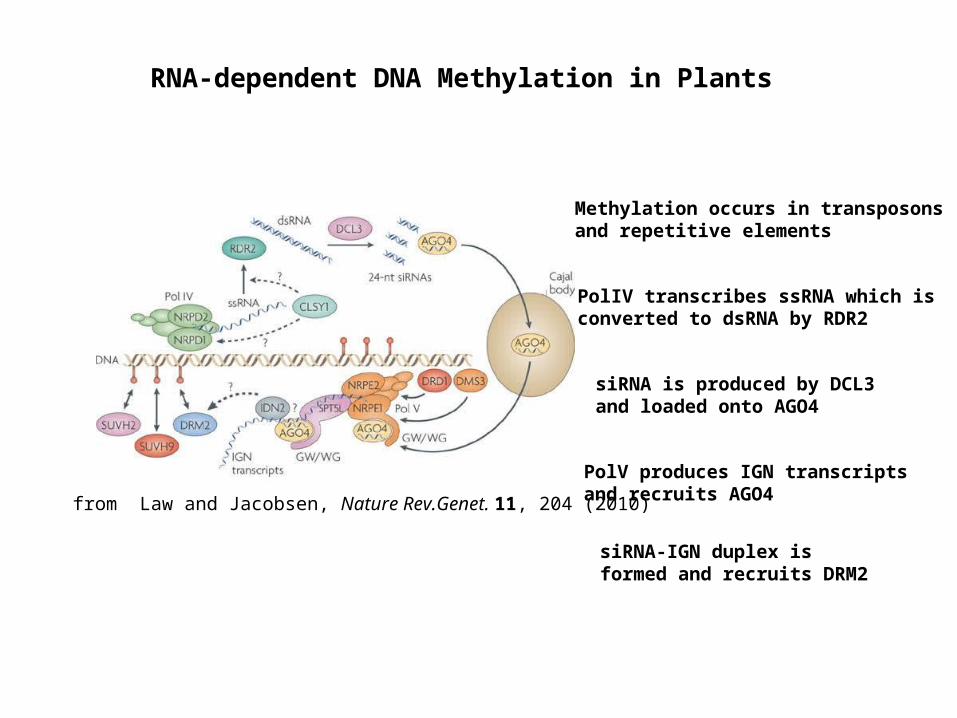
RNA-dependent DNA Methylation in Plants
Methylation occurs in transposons and repetitive elements
PolIV transcribes ssRNA which is converted to dsRNA by RDR2
siRNA is produced by DCL3 and loaded onto AGO4
PolV produces IGN transcripts and recruits AGO4
siRNA-IGN duplex is formed and recruits DRM2
from Law and Jacobsen, Nature Rev.Genet. 11, 204 (2010)

from Law and Jacobsen, Nature Rev.Genet. 11, 204 (2010)
De Novo DNA Methylation in Mammals
DNMT3L interacts with unmethylated H3K4
DNMT3A is recruited and activated and forms a tetrameric complex
Active sites are separated by 8-10 bp and methylates opposite DNA strands
Tetramer oligomerizes and results in 10 bp pattern of methylation on the same strand
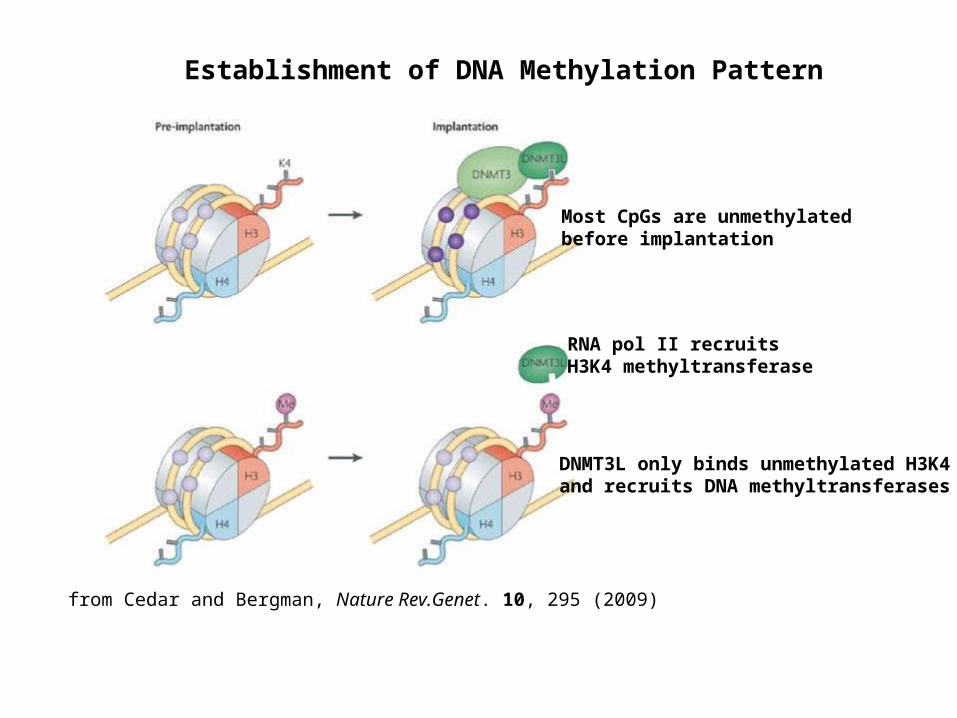
from Cedar and Bergman, Nature Rev.Genet. 10, 295 (2009)
Establishment of DNA Methylation Pattern
Most CpGs are unmethylated before implantation
RNA pol II recruits H3K4 methyltransferase
DNMT3L only binds unmethylated H3K4 and recruits DNA methyltransferases
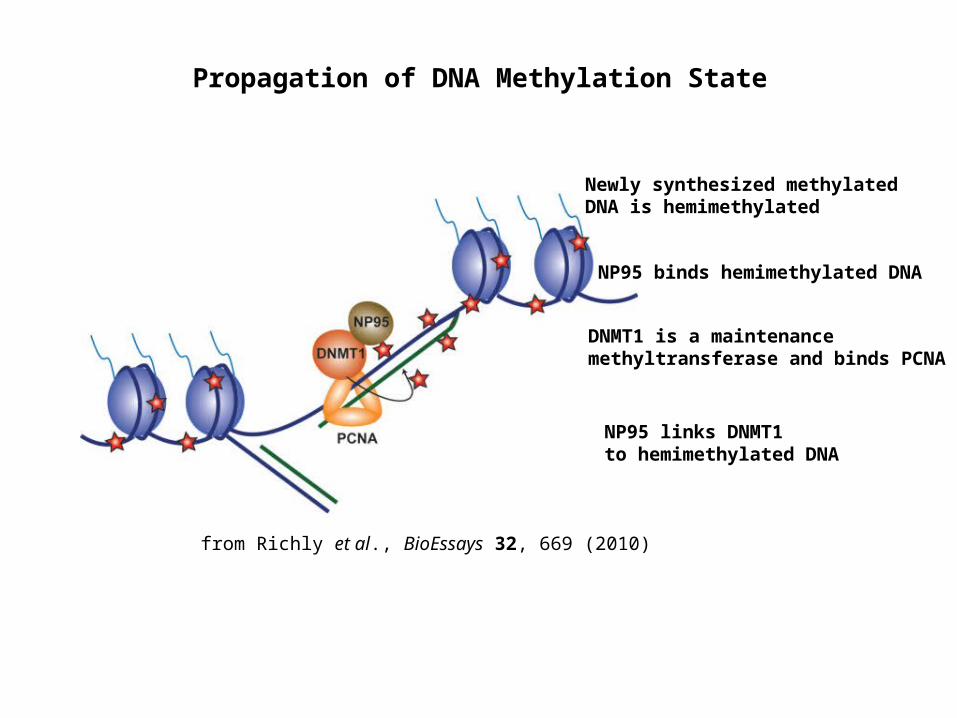
Propagation of DNA Methylation State
Newly synthesized methylated DNA is hemimethylated
NP95 links DNMT1to hemimethylated DNA
DNMT1 is a maintenance methyltransferase and binds PCNA
NP95 binds hemimethylated DNA
from Richly et al., BioEssays 32, 669 (2010)
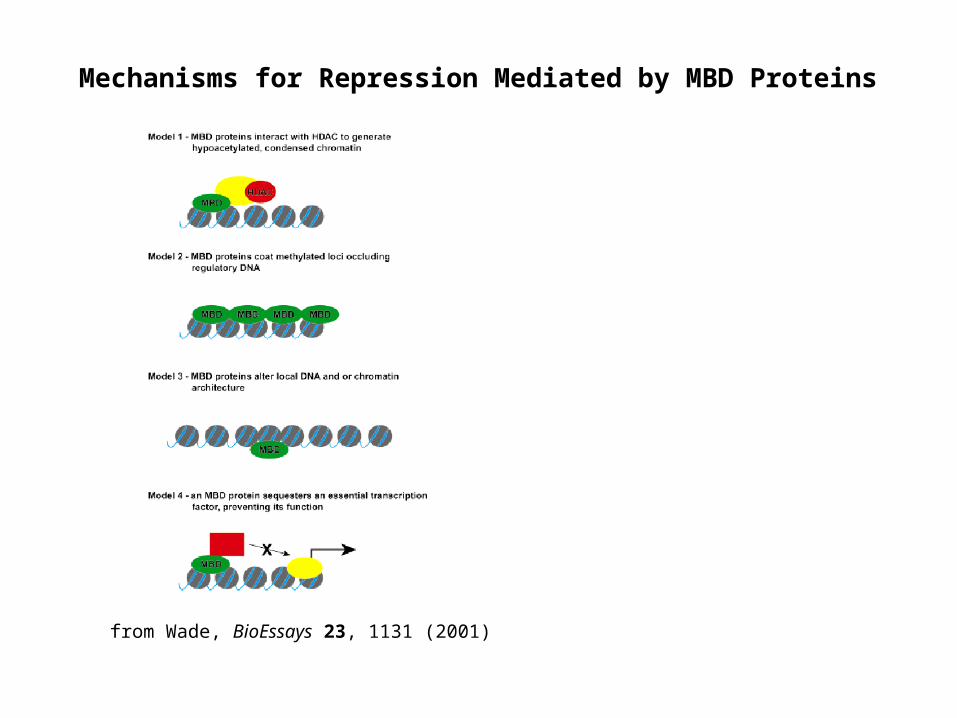
Mechanisms for Repression Mediated by MBD Proteins
from Wade, BioEssays 23, 1131 (2001)
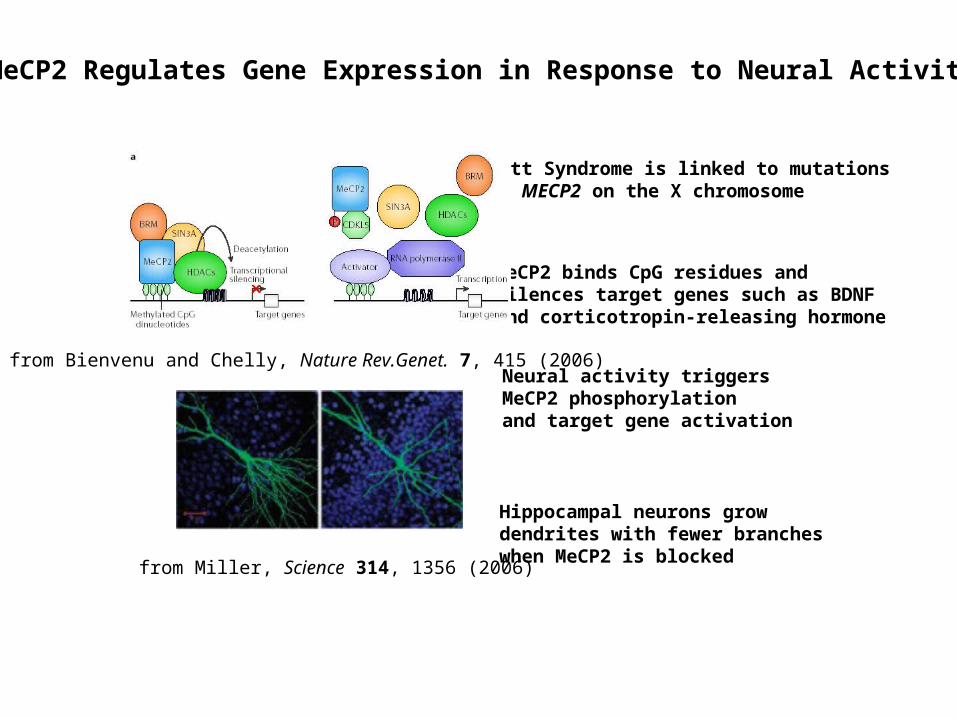
Rett Syndrome is linked to mutations in MECP2 on the X chromosome
MeCP2 binds CpG residues and silences target genes such as BDNF and corticotropin-releasing hormone
Neural activity triggers MeCP2 phosphorylation and target gene activation
MeCP2 Regulates Gene Expression in Response to Neural Activity
Hippocampal neurons grow dendrites with fewer branches when MeCP2 is blocked
from Miller, Science 314, 1356 (2006)
from Bienvenu and Chelly, Nature Rev.Genet. 7, 415 (2006)
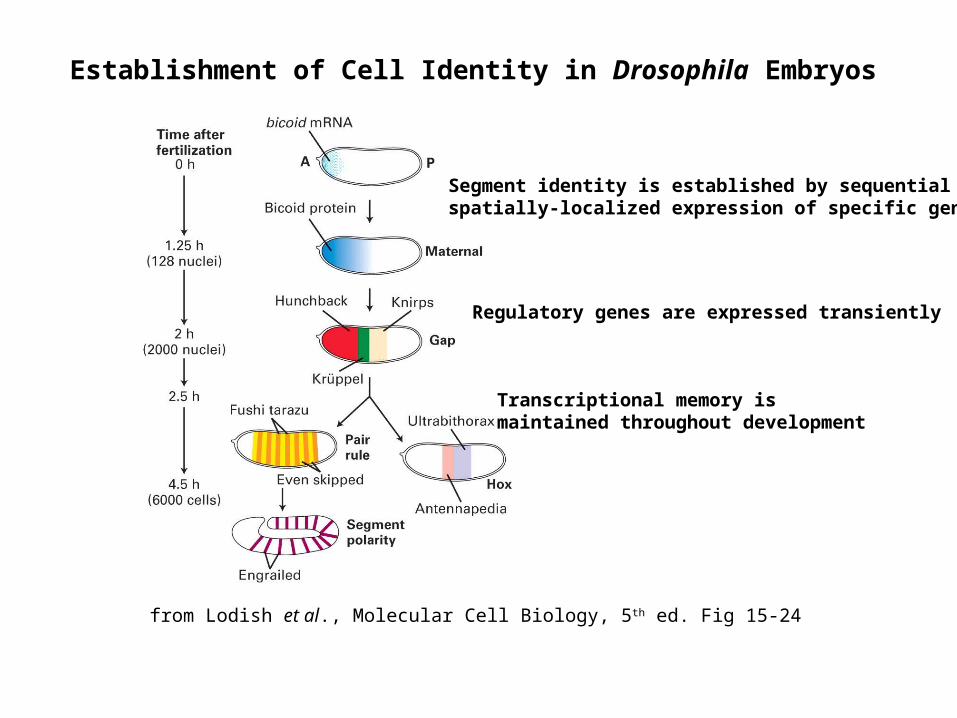
Establishment of Cell Identity in Drosophila Embryos
from Lodish et al., Molecular Cell Biology, 5th ed. Fig 15-24
Segment identity is established by sequential spatially-localized expression of specific genes
Regulatory genes are expressed transiently
Transcriptional memory is maintained throughout development

Misexpression of Homeotic Genes Lead to Morphological Abominations
from Lodish et al., Molecular Cell Biology, 5th ed. Fig 15-25

Polycomb-group Proteins
Maintains a silenced state
Trithorax-group Proteins
Maintains an active state
Counteracts the action of PcG proteins
Memory system composed of PcG and trxG complexes is linked to the histone code
Prevents changes in cell identity by preserving transcription patterns
Polycomb and Trithorax Complexes
Chromatin is altered in a heritable manner
Prevents chromatin remodelling
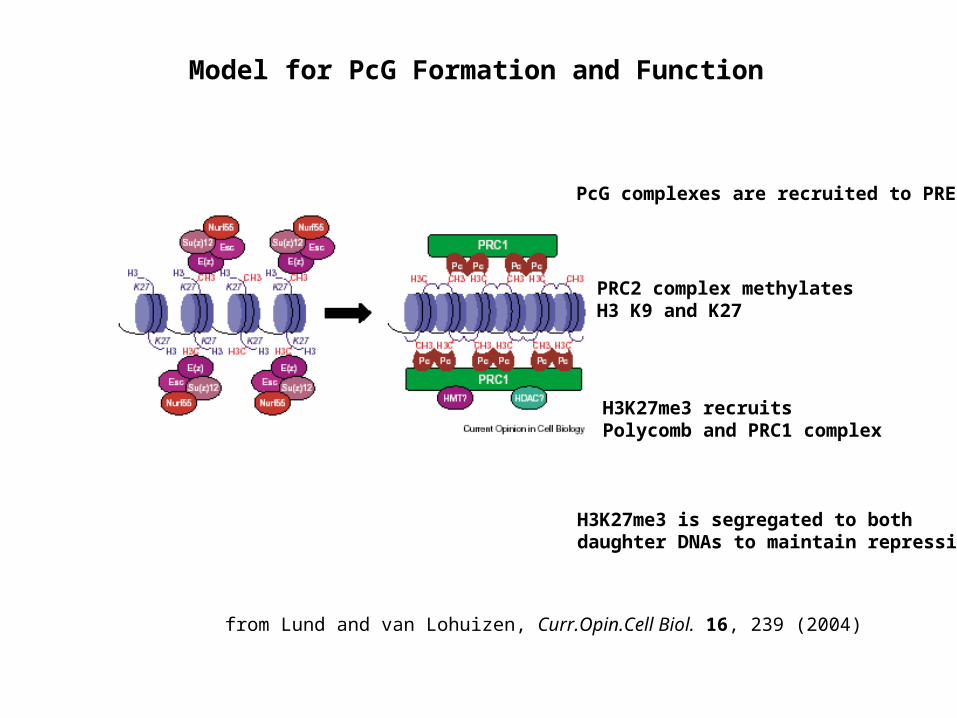
Model for PcG Formation and Function
from Lund and van Lohuizen, Curr.Opin.Cell Biol. 16, 239 (2004)
PRC2 complex methylates H3 K9 and K27
H3K27me3 recruits Polycomb and PRC1 complex
H3K27me3 is segregated to both daughter DNAs to maintain repression
PcG complexes are recruited to PREs
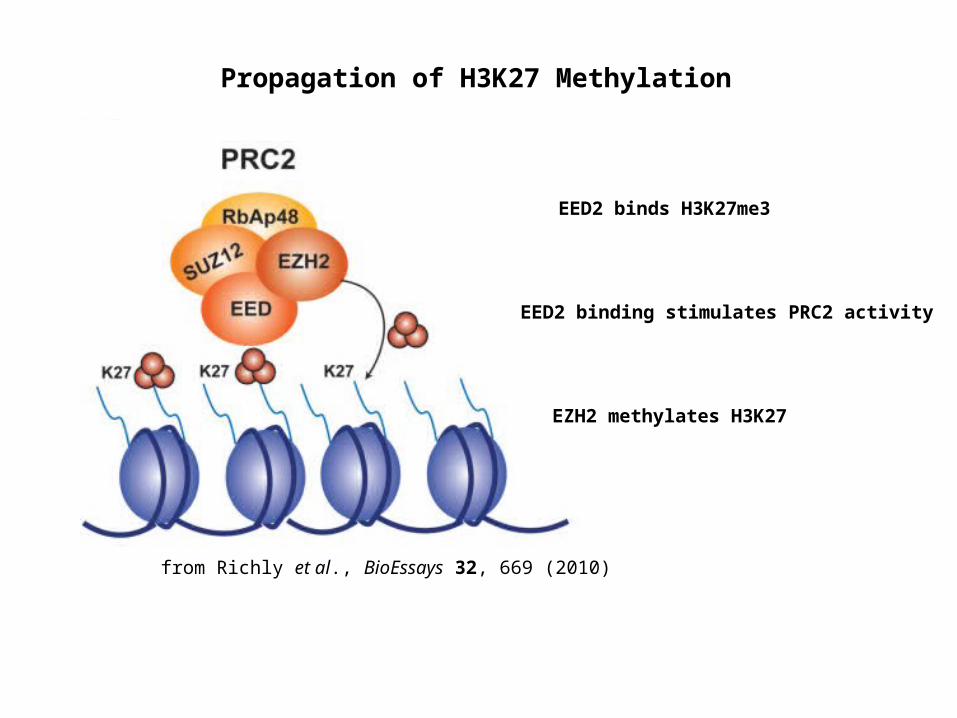
from Richly et al., BioEssays 32, 669 (2010)
Propagation of H3K27 Methylation
EED2 binds H3K27me3
EED2 binding stimulates PRC2 activity
EZH2 methylates H3K27
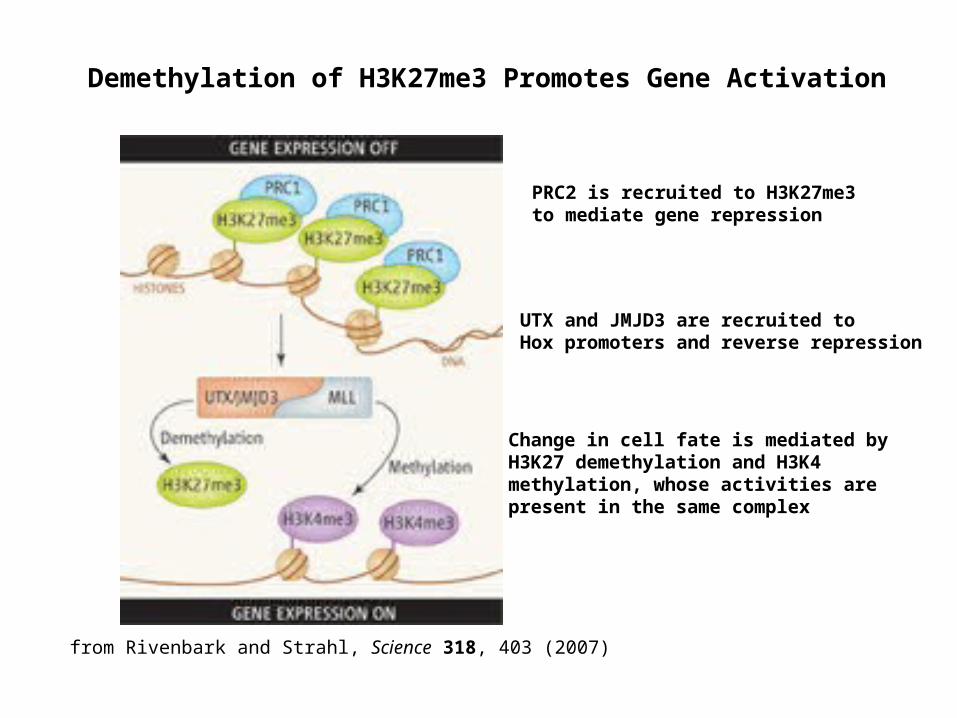
Demethylation of H3K27me3 Promotes Gene Activation
PRC2 is recruited to H3K27me3 to mediate gene repression
UTX and JMJD3 are recruited to Hox promoters and reverse repression
Change in cell fate is mediated by H3K27 demethylation and H3K4 methylation, whose activities are present in the same complex
from Rivenbark and Strahl, Science 318, 403 (2007)

Trithorax Complex Mechanism of Action
TrxC methylates H3K4 and recruits HAT and remodeling complexes
Acetylated H3K9 prevents methylation, and prevents HP1 binding
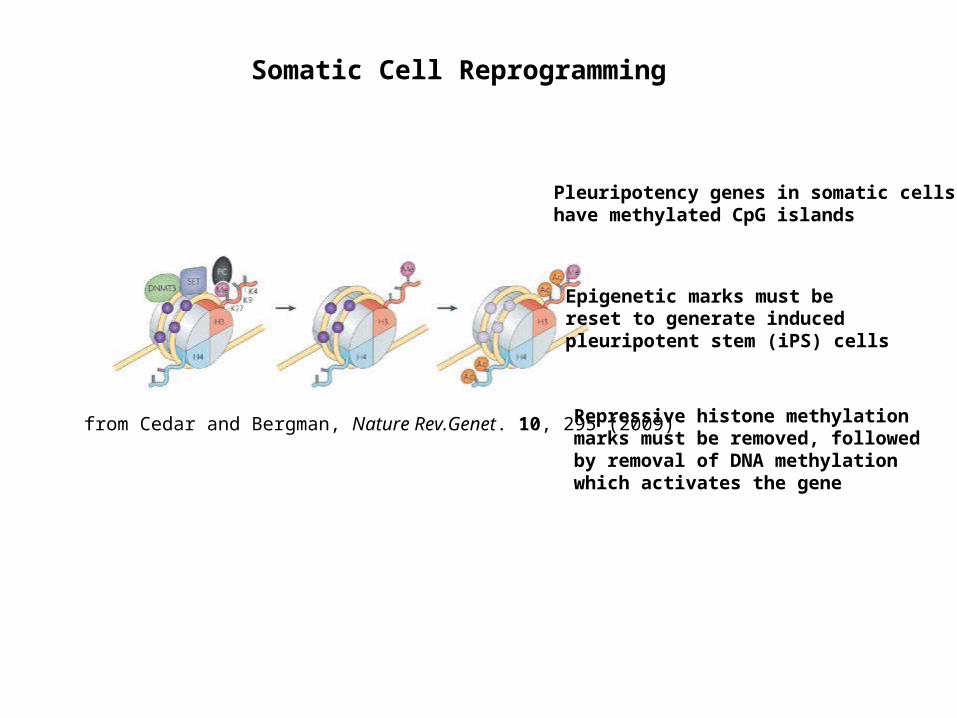
Somatic Cell Reprogramming
from Cedar and Bergman, Nature Rev.Genet. 10, 295 (2009)
Pleuripotency genes in somatic cells have methylated CpG islands
Epigenetic marks must be reset to generate induced pleuripotent stem (iPS) cells
Repressive histone methylation marks must be removed, followed by removal of DNA methylation which activates the gene
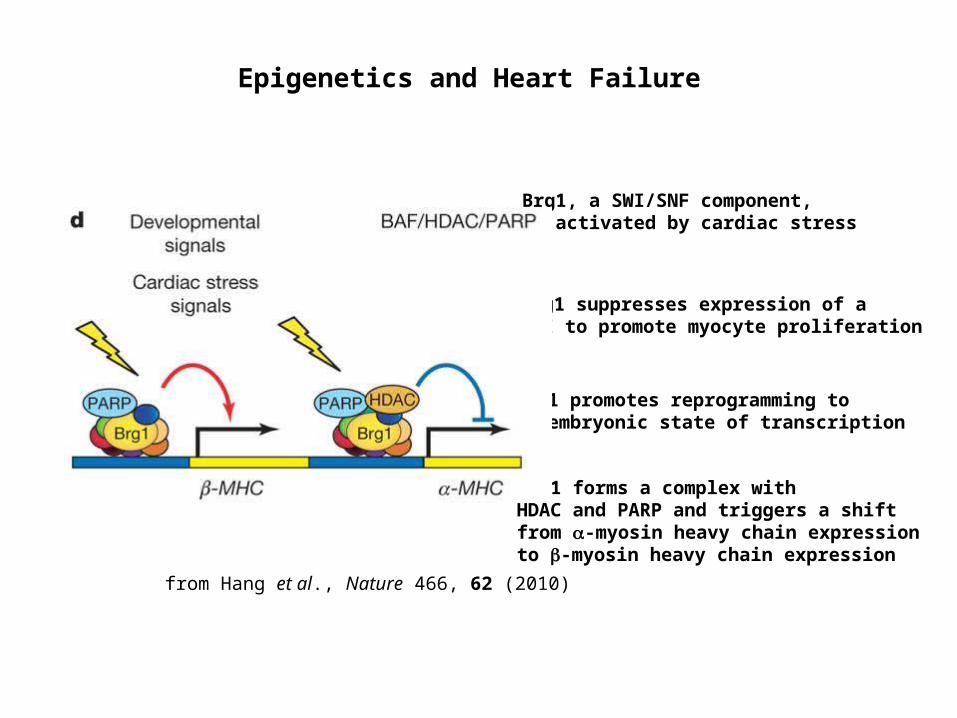
Brg1, a SWI/SNF component, is activated by cardiac stress
Brg1 suppresses expression of a CKI to promote myocyte proliferation
Brg1 forms a complex with HDAC and PARP and triggers a shift from -myosin heavy chain expression to -myosin heavy chain expression
Epigenetics and Heart Failure
from Hang et al., Nature 466, 62 (2010)
Brg1 promotes reprogramming to an embryonic state of transcription
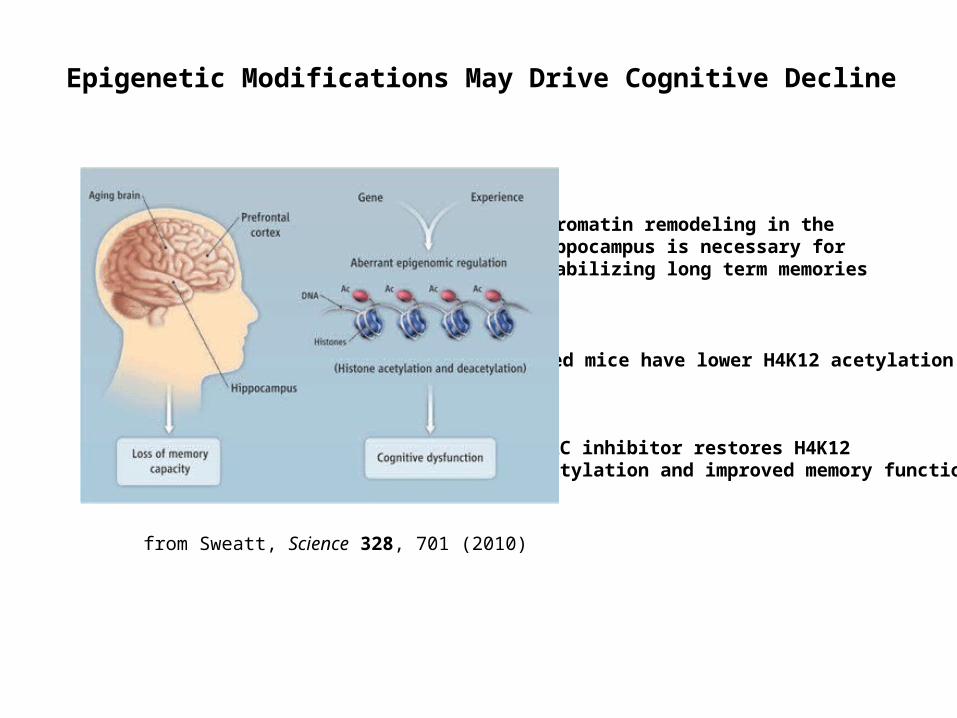
Epigenetic Modifications May Drive Cognitive Decline
from Sweatt, Science 328, 701 (2010)
Chromatin remodeling in the hippocampus is necessary for stabilizing long term memories
Aged mice have lower H4K12 acetylation
HDAC inhibitor restores H4K12 acetylation and improved memory function
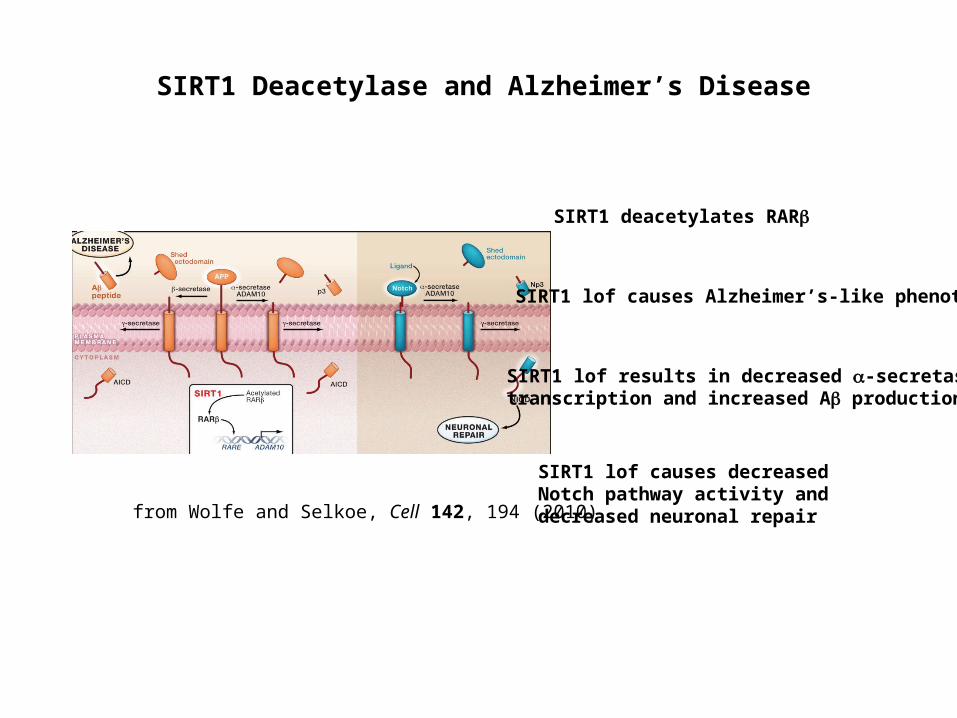
SIRT1 lof causes Alzheimer’s-like phenotype
SIRT1 deacetylates RAR
SIRT1 lof results in decreased -secretase transcription and increased A production
SIRT1 lof causes decreased Notch pathway activity and decreased neuronal repair
SIRT1 Deacetylase and Alzheimer’s Disease
from Wolfe and Selkoe, Cell 142, 194 (2010)
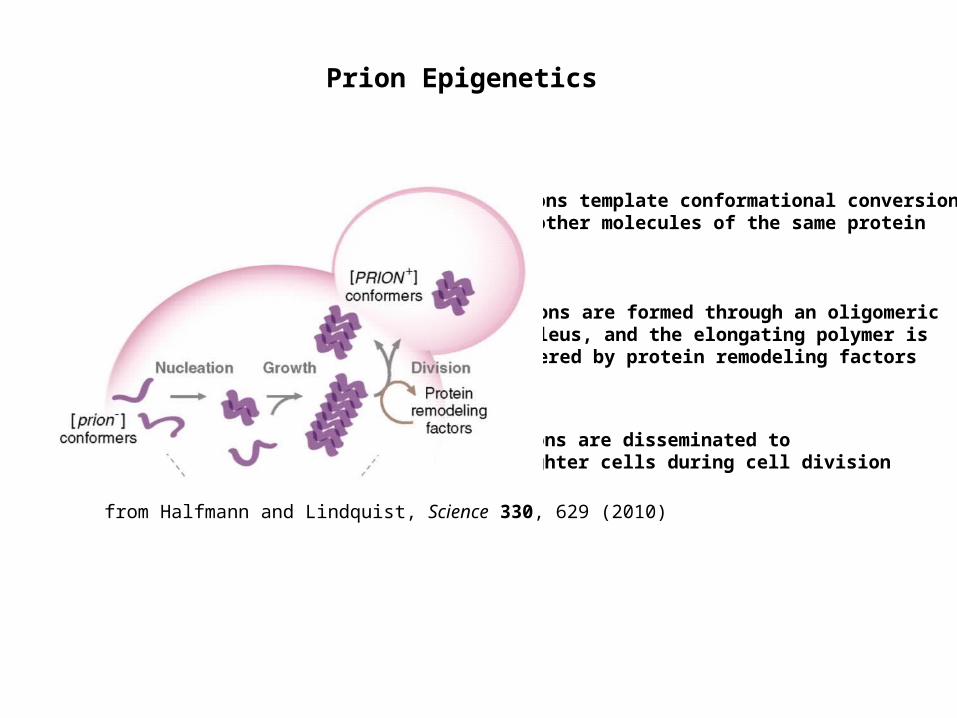
Prion Epigenetics
Prions template conformational conversion of other molecules of the same protein
Prions are disseminated to daughter cells during cell division
Prions are formed through an oligomeric nucleus, and the elongating polymer is severed by protein remodeling factors
from Halfmann and Lindquist, Science 330, 629 (2010)
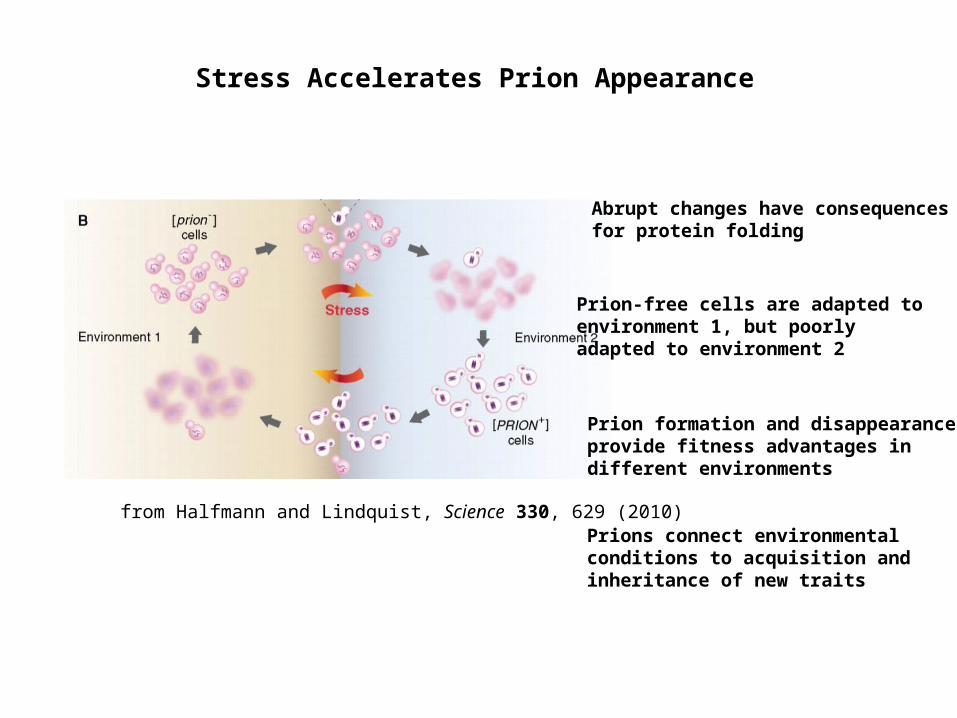
Stress Accelerates Prion Appearance
from Halfmann and Lindquist, Science 330, 629 (2010)
Prion-free cells are adapted to environment 1, but poorly adapted to environment 2
Prion formation and disappearance provide fitness advantages in different environments
Abrupt changes have consequences for protein folding
Prions connect environmental conditions to acquisition and inheritance of new traits
Co-suppression
PcG complexes interact in trans
Increase in gene copy number results in decreased expression
Dependent on PcG genes
from Pirrotta, Cell 93, 333 (1998)

Imprinting
Expression of only one allele of a locus
Only 80 genes in mammals are imprinted
Most imprinted genes are involved in growth control
Imprinted genes involves allele specific methylation and is resistant to genome-wide demethylation
Clusters of imprinted genes contain noncoding RNAs that are involved control allele-specific expression
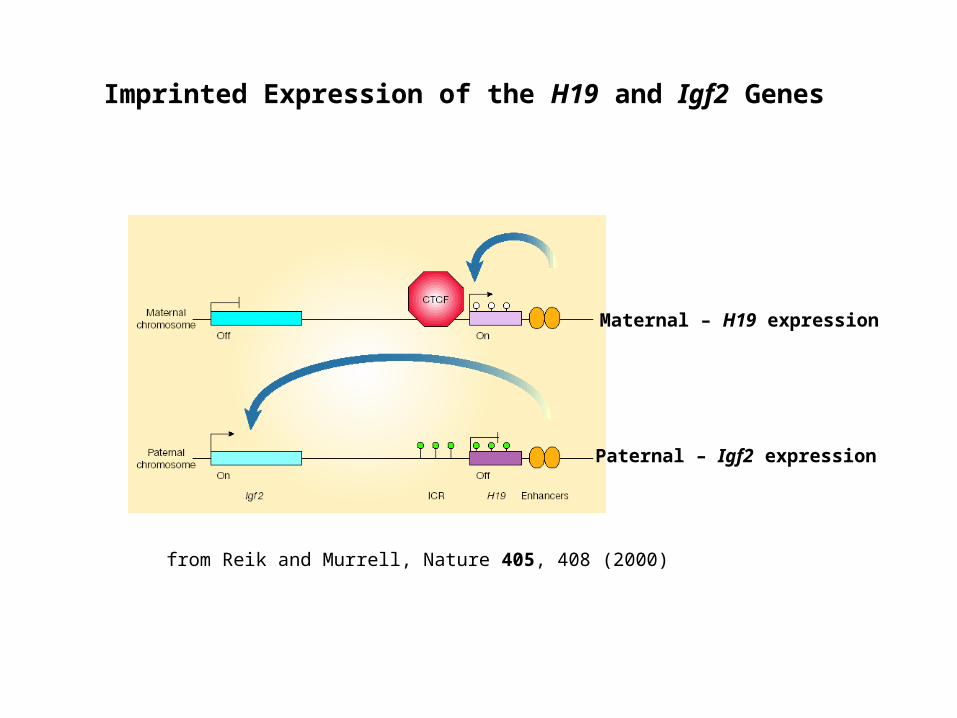
Imprinted Expression of the H19 and Igf2 Genes
from Reik and Murrell, Nature 405, 408 (2000)
Maternal – H19 expression
Paternal – Igf2 expression
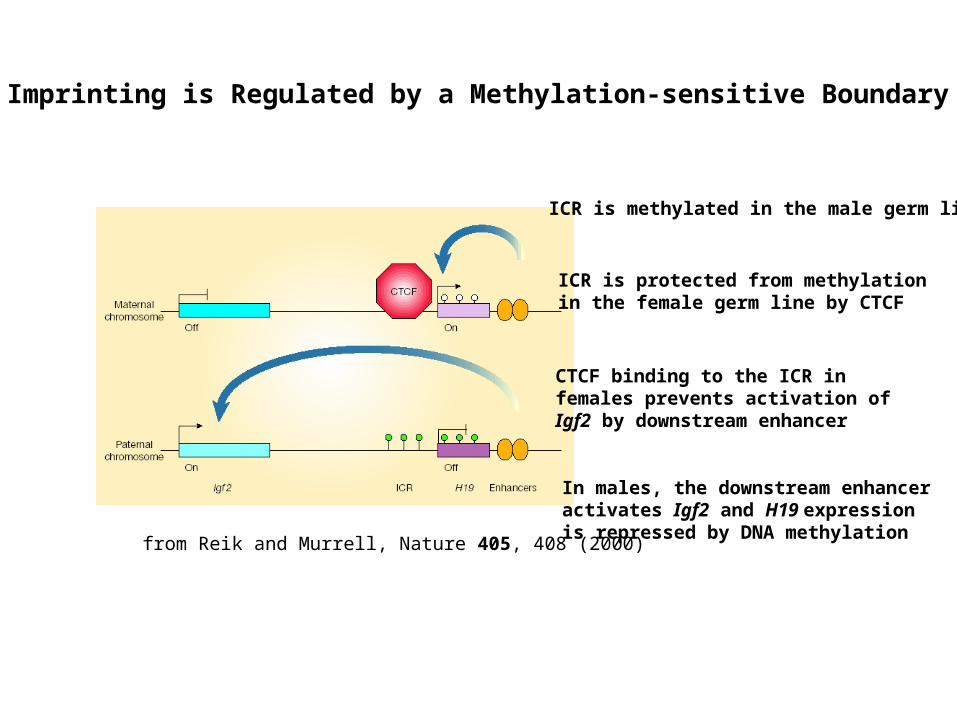
Imprinting is Regulated by a Methylation-sensitive Boundary
from Reik and Murrell, Nature 405, 408 (2000)
ICR is methylated in the male germ line
ICR is protected from methylation in the female germ line by CTCF
CTCF binding to the ICR in females prevents activation of Igf2 by downstream enhancer
In males, the downstream enhancer activates Igf2 and H19 expression is repressed by DNA methylation

from Ferguson-Smith and Surani, Science 293, 1086 (2001)
Imprinting of the PWS-AS Locus
The AS-ICR is required for methylation and inactivation of the PWS-ICR in females to repress nearby genes
The AS-ICR is nonfunctional in males allowing the PWS-ICR to activate nearby genes
The PWS-ICR promotes expression of an antisense Ube3a transcript in males
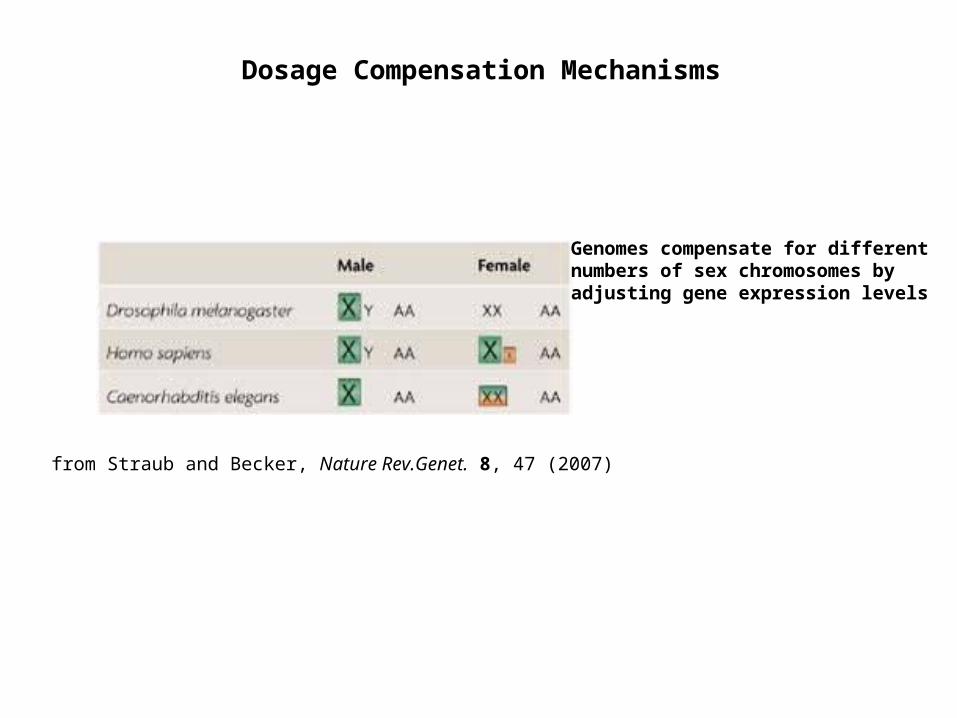
from Straub and Becker, Nature Rev.Genet. 8, 47 (2007)
Dosage Compensation Mechanisms
Genomes compensate for different numbers of sex chromosomes by adjusting gene expression levels

X Inactivation in Mammals
X inactivation is initiated from the Xic
Xist and Tsix partially overlap and are transcribed in opposite directions from the Xic
from Lodish et al., Molecular Cell Biology, 6th ed. Fig 22-7
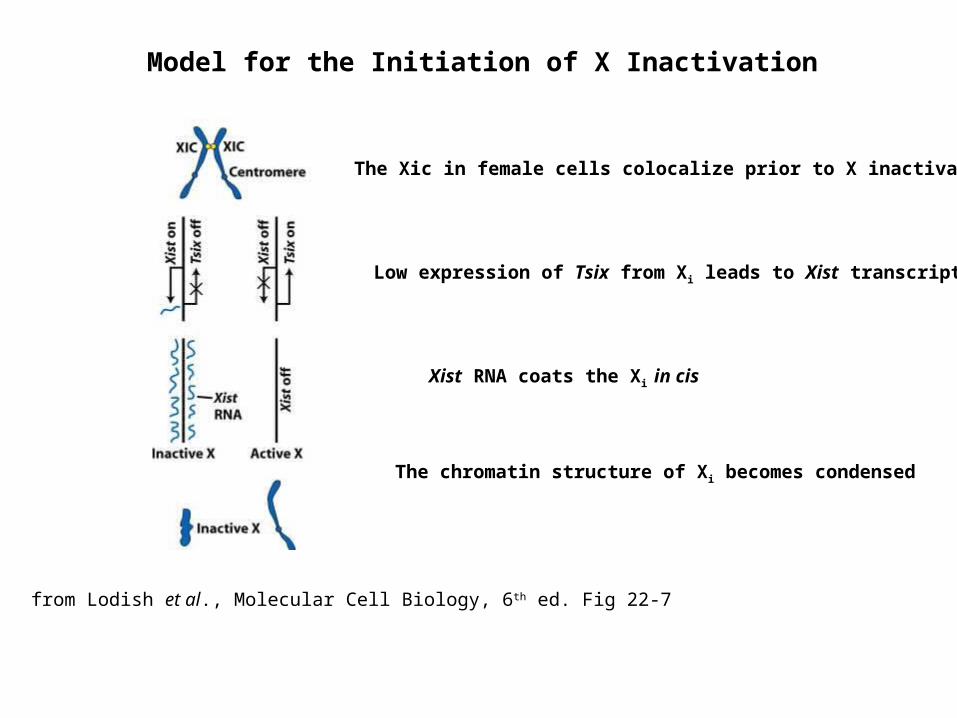
Model for the Initiation of X Inactivation
from Lodish et al., Molecular Cell Biology, 6th ed. Fig 22-7
The Xic in female cells colocalize prior to X inactivation
Low expression of Tsix from Xi leads to Xist transcription
Xist RNA coats the Xi in cis
The chromatin structure of Xi becomes condensed

Stepwise Progression of X Inactivation in Differentiating ES Cells
from Brockdorff, Trends Genet. 18, 352 (2002)
One X chromosome is converted to facultative heterochromatin
Xist transcription off the inactive X initiates chromatin modification events
X inactivation is maintained epigenetically

Calico Cats
One of the genes controlling fur color is on the X chromosome
B – orangeb - black
Random X inactivation early in embryonic development leads to patchworks of skin cells expressing each allele
Female mammals are genetic mosaics

The Dosage Compensation Complex in Drosophila
from Gilfillan et al., FEBS Lett. 567, 8 (2004)
SXL in females prevents MSL2 translation
MSL2 in males stabilizes roX, MSL1, and MSL3
DCC binds to high affinity sites on X chromosome
DCC spreads to nearby sites on active chromatin
H4K16 acetylation impedes formation of condensed chromatin structure

from Straub and Becker, Nature Rev.Genet. 8, 47 (2007)
DCC is Localized to the X Chromosome
DCC localization is determined by staining of polytene chromosomes with anti-MSL1
DCC associates almost exclusively with transcribed regions