growth inhibition of soybean callus tissues by glutathione
TRANSCRIPT
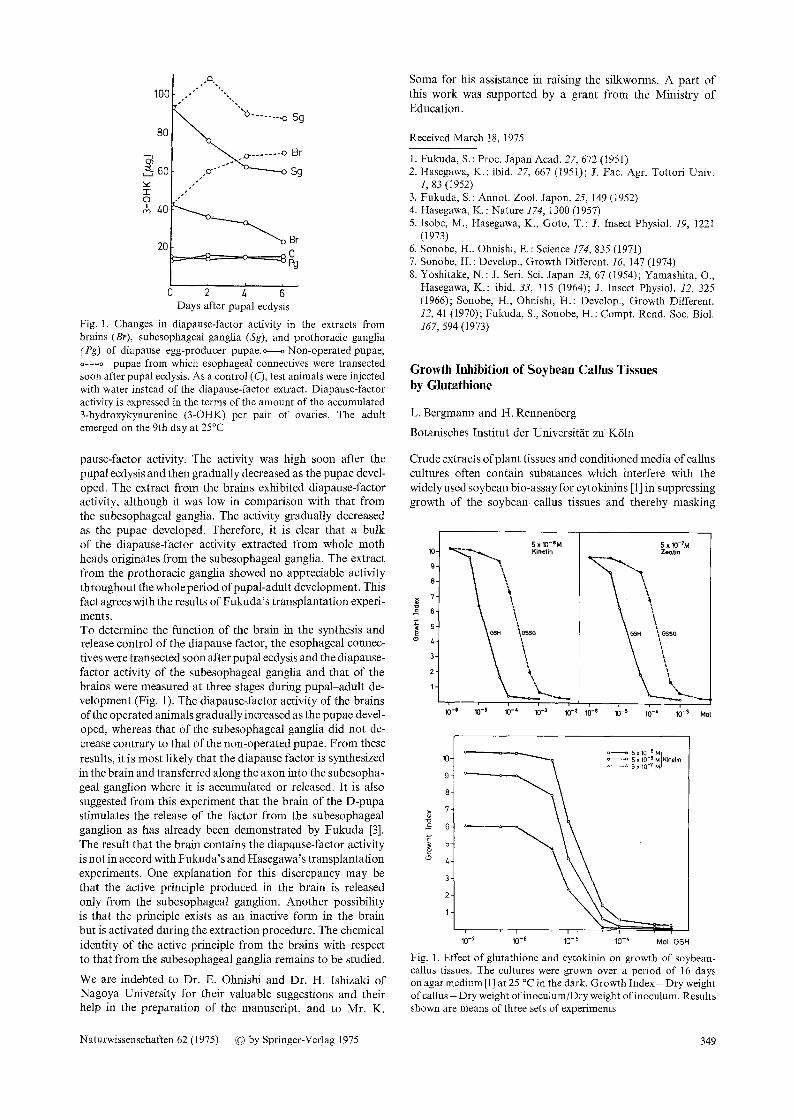
13 100 ,.."" "'"-.,.
' "'% . . . . . . . -o Sg
Br ::~60 .o- o
Sg
"1- 0 ""
" 40
Br 20
,, C F~
o 2 ~ Days after pupal ecdysis
Fig. 1. Changes in diapause-factor activity in the extracts from brains (Br), subesophageal ganglia (Sg), and prothoracic ganglia (Pg) of diapause egg-producer pupae.o---oNon-operatedpupae, o---o pupae from which esophageal connectives were transected soon after pupal ecdysis. As a control (C), test animals were injected with water instead of the diapause-factor extract. Diapause-factor activity is expressed in the terms of the amount of the accumulated 3-hydroxykynurenine (3-OHK) per pair of ovaries. The adult emerged on the 9th day at 25~
pause-factor activity. The activity was high soon after the pupal ecdysis and then gradually decreased as the pupae devel- oped. The extract from the brains exhibited diapause-factor activity, although it was low in comparison with that from the subesophageal ganglia. The activity gradually decreased as the pupae developed. Therefore, it is clear that a bulk of the diapause-factor activity extracted from whole moth heads originates from the subesophageal ganglia. The extract from the prothoracic ganglia showed no appreciable activity throughout the whole period of pupal-adult development. This fact agrees with the results of Fukuda's transplantation experi- ments. To determine the function of the brain in the synthesis and release control of the diapause factor, the esophageal connec- tives were transected soon after pupal ecdysis and the diapause- factor activity of the subesophageal ganglia and that of the brains were measured at three stages during pupal-adult de- velopment (Fig. 1). The diapause~factor activity of the brains of the operated animals gradually increased as the pupae devel- oped, whereas that of the subesophageal ganglia did not de- crease contrary to that of the non-operated pupae. From these results, it is most likely that the diapause factor is synthesized in the brain and transferred along the axon into the subesopha- geal ganglion where it is accumulated or released. It is also suggested from this experiment that the brain of the D-pupa stimulates the release of the factor from the subesophageal ganglion as has already been demonstrated by Fukuda [3]. The result that the brain contains the diapause-factor activity is not in accord with Fukuda 's and Hasegawa's transplantation experiments. One explanation for this discrepancy may be that the active principle produced in the brain is released only from the subesophageal ganglion. Another possibility is that the principle exists as an inactive form in the brain but is activated during the extraction procedure. The chemical identity of the active principle from the brains with respect to that from the subesophageal ganglia remains to be studied.
We are indebted to Dr. E. Ohnishi and Dr. H. Ishizaki of Nagoya University for their valuable suggestions and their help in the preparation of the manuscript, and to Mr. K.
Soma for his assistance in raising the silkworms. A part of this work was supported by a grant from the Ministry of Education.
Received March 18, 1975
1. Fukuda, S. : Proc. Japan Acad. 27, 672 (1951) 2. Hasegawa, K.: ibid. 27, 667 (1951); J. Fac. Agr. Tottori Univ.
1, 83 (I952) 3. Fukuda, S. : Annot. Zool. Japon. 25, 149 (1952) 4. Hasegawa, K. : Nature 174, 1300 (1957) 5. Isobe, M., Hasegawa, K., Goto, T.: J. Insect Physiol. 19, 1221
(1973) 6. Sonobe, H., Ohnishi, E. : Science 174, 835 (1971) 7. Sonobe, H. : Develop., Growth Different. 16, 147 (1974) 8. Yoshitake, N. :J. Seri. Sci. Japan 23, 67 (1954); Yamashita, O.,
Hasegawa, K.: ibid. 33, 115 (1964); J. Insect Physiol. 12, 325 (1966); Sonobe, H., Ohnishi, H.: Develop., Growth Different. 12, 41 (1970); Fukuda, S., Sonobe, H. : Compt. Rend. Soc. Biol. 167, 594 (1973)
Growth Inhibition of Soybean Callus Tissues by Glutathione
L. Bergmann and H. Rennenberg
Botanisches Institut der Universitfit zu K61n
Crude extracts of plant tissues and conditioned media of callus cultures often contain substances which interfere with the widely used soybean bio-assay for cytokinins [1] in suppressing growth of the soybean callus tissues and thereby masking
10
g,
8
,~ "7,
7 6. ~: 5" s o g.
3-
5 x 10-6M " - - ' ~ . . ~ Kinetin
"\ 10-6
5 x IO-'/M Zeatin
Cr 1\6SSG
,o-' ; - ' ,o-, ";-' , ;- ' ,o-' ,o-' , ; - ' ,,o,
o ~ 5X10-6 M 10" ~ 5 x 10 -5 MIKinetln
& - - & 5x 10-7 M /
9-
8-
~, 7-
O L-
3-
2-
1-
i u t i 10 -7 10 -e 10 -5 10 -4 Mo[ GSH
Fig. 1. Effect of glutathione and cytokinin on growth of soybean- callus tissues. The cultures were grown over a period of 16 days on agar medium [1] at 25 ~ in the dark. Growth Index = Dry weight of callus - Dry weight of inoculum/Dry weight of inoculum. Results shown are means of three sets of experiments
Naturwissenschaften 62 (1975) �9 by Springer-Verlag 1975 349

the cytokinin effect [2]. In this communication evidence is presented that glutathione is such an inhibitory substance. The cytokinin-dependent growth of callus cultures isolated from the cotyledons of Glycine max. L. var. Corsoy is inhibited by glutathione which has been purchased from Boehringer Mannheim sterilized by filtration and added aseptically to the autoclaved media. As shown in Fig. 1 the growth is reduced to about 50% by the addition of 10-5M reduced glutathione (GSH) and it is completely inhibited by concentrations of 10-4M and higher. Oxidized glutathione (GSSG) also inhibits the growth of soybean tissues, but for comparable effects con- centrations about ten times those necessary with GSH are needed. The growth inhibition by glutathione is observed with soybean cultures grown on media supplemented with either kinetin or zeatin as cytokinin. It cannot be compensated by higher kinetin concentrations in the medium (Fig. 1), nor do higher levels of auxin (10- 6 _ 10 4 MNAA [naphthalene acetic acid]) counteract the inhibition. The growth inhibition seems to depend on the whole peptide, since an addition of filter-sterilized glutamic acid, glycine, and cysteine alone as well as together has no inhibitory effect on the growth of the soybean tissues (Table 1).
Table 1. Dry-weight yields (mg/cuIture) of soybean callus on media with and without amino acids. The cultures were grown over a period of 16 days on media [1] supplemented with 5 x 10- 6 M kinetin and 10-SM NAA at 25 ~ in the dark
Control 21.8 • 1.2 Glutamic acid (5 x 10-4M) 20.4_+ 1.5 Cysteine (5 x 10-4N0 18.4• 1.1. Glycine (5 x 10 4M) 26.4+2.0 Glutamic acid, cysteine,
glycine (5 • 10-4M each) 20.0 + 1.2
Glutathione is present in substancial amounts in plant cells [3] the concentrations reported resembling those shown to be inhibitory to soybean-callus growth; it can also be found in conditioned media from suspension cultures of tobacco cells (Rennenberg, unpublished results). Since glutathione is extracted by most of the solvents used for the extraction of cytokinins it is important for tests with the soybean No-assay to separate glutathione and cytokinins by ion-exchange chro- matography or other methods.
Received March 14, 1975
1. Miller, C.O., in: Modern Methods of Plant Analysis, Vol. 6 (ed. H.F. Linskens and M.V. Tracey). Berlin: Springer 1963
2. Fox, J.E., in: The Physiology of Plant Growth and Development (ed. M.B. Wilkins). London: McGraw-Hill 1969
3. Jocelyn, P.C. : The Biochemistry of the SH-Group. London: Aca- demic Press 1972
Nitrogenase Activity in Associations of Rhizobium lupini and Lupinus polyphyUus Tissue Cultures
D. Werner and G. Oberlies
Fachbereich Biologie der Botanisches Institut
Universitfit Marburg/Lahn,
The symbiosis between legumes and rhizobia is distinguished by the ecological importance of the symbiotic nitrogen fixa- tion, which is in the same order of magnitude as the whole
industrial nitrogen fertilizer production (about 40 mill. t/ year) [1]. A new approach to the study of the development of the symbiosis between rhizobia and legumes and of the genetics of rhizobia and of the symbiotic nitrogen fixation was the first successful demonstration of an in-vitro symbiosis in tissue cultures with Rhizobiumjaponicum strains as effective bacteria [2, 3]. We report here on what is, as far as we are aware, the first association with nitrogenase activity of tissue cultures and Rhizobium lupini. Rhizobium lupini has a much more discrete infection range in legumes than Rhizobium japonicum [4]. The genetics have been studied chiefly by Heumann and coworkers [5], and are much better understood than those of Rhizobium japonicum. The results given in Fig. 1 are from experiments with two
13001
4~
,~ ~ooo
E 700
~z"
~00 (b
0
E t00
O
tl i t
, , ' / \ ! I 1 i o
o
0 15 2'5 3'5 4'5 5'5 55 d af ter infect ion
Fig. 1. Development of nitrogenase activity (acetylene-reduction assay) in associations of Rhizobium lupini strain 7-1 (--�9 and strain 7-2 ( - o - ) with Lupinus polyphyllus tissue cultures at 28 ~
Rhizobium lupini strains (N 9. 7-1 and No. 7-2) which we received by courtesy of Dr. Heumann, University of Erlangen. The strains were isolated from the same source (Lupinus hart- wegii), but they differed in their sensitivity to streptomycin. The tissue cultures of Lupinus polyphyllus were transferred into liquid suspension in our laboratory from a root callus and cultivated over many passages. The usual microbiological tests were performed to assure axenic cultures. The liquid- grown plant cells were transferred to so-called tissue chambers on a solid medium [6]. After 14 days the plant-cell layer was infected with 0.3 ml bacterial suspension (about 1 x 10 9 cells/ ml). After the times indicated in Fig. 1, the acetylene-reduction activity was determined by incubating 3 tissue chambers in an atmosphere of 8 % v/v acetylene for 24 h under sterile condi- tions. The ethylene produced was determined by gaschromato- graphy [7]. Endogenously formed ethylene was not detect- able. The data show that the development of nitrogenase activity in the associations starts between 4 and 5 weeks after infection. After 6 to 7 weeks the activity reaches a value of 900 to 1200 nmoles CZH4/24 h. Calculated per g dry weight of the association, the activity is 56 to 86 nmoles C 2 H 4 x h - t • dry weight -~. This is between 10 and 20% of the activity obtained with associations between soybean tissue cultures and Rhizobiumjaponicum [6]. The development of nitrogenase activity with soybean tissue cultures and Rhizobium japonieum lasts only 10 to 14 days after infection by the same method,
350 Naturwissenschaften 62 (1975) �9 by Springer-Verlag 1975