gonad development in billfish (istiophoridae) from the indian ocean
TRANSCRIPT

J . Zool., Lond. (1970) 160, 355-370
Gonad development in billfish (Istiophoridae) from the Indian Ocean*
N. R. MERRETT
East African Marine Fisheries Research Organization, Zanzibar:
(Accepted 19 September 1969)
(With 8 plates and 4 figures in the text)
Gonad examinations were made on billfish caught during a tuna longline survey in the equatorial western Indian Ocean. The species considered were Zstiophorus platypterus, Tetrapterus a h , Tetrapterus angustirostris, Makaira nigricans, and Makaira indica. The morphology of the urino-genital system is described for all species. It was found to be similar in all except T. angustirostris in which the gonads are Y-shaped rather than bilaterally symmetrical. The histology of the gonads is described, and the changes within the testis and ovary are traced throughout the spermatogenic and oogenic cycles. These are shown to be similar in all species examined.
Contents Introduction . . . . . . . . Materials and methods . . . . Morphology . . . . . . . . Histology . . . . . . . .
The testis . . . . . . The ovary . . . . . .
Discussion . . . . . . .. summary . . . . . . . . References . . . . . . . . Abbreviations used in plates and figures Stains used . . . . . . . .
. . . . . . . . . . . . . . 355
. . . . . . . . . . . . . . 356 .. 351
. . . . . . . . . . . . . . 361
. . . . . . . . . . . . . . 361
. . . . . . . . . . . . . . 363
. . . . . . . . . . . . 368
. . . . . . . . . . . . . . 368
. . . . . . . . . . . . . . 369 .. 370
.. .. 370
. . . . . . . . . . . .
..
. . . . . . . . . . . .
. . . . . . . . . .
Introduction Knowledge of the breeding biology of commercially important species is one of the
major aims of fisheries research. The billfishes (family Istiophoridae) are prominent constituents of the pelagic oceanic catches of the world, yet little is known of their repro- ductive cycles. The relation of the gonads to the other visceral organs in billfish has been briefly described by Nakamura (1938, 1949), La Monte (1958), and de Sylva & Davis (1963). The latter authors also comment upon the histology of the ripening ovary and testis, but made no detailed examination of changes within the gonads throughout their development. Thus there is a need for a detailed description of the morphology of the gonads, with the closely linked urinary system of these fish, and of the histological changes
* This study constituted part of a thesis presented in fulfilment of the requirements for the degree of Master of
2 Present address: National Institute of Oceanography, Wormley, Godalming, Surrey, England. Science in the University of East Africa.
27 355

356 N. R . M E R R E T T
occurring during oogenesis and spermatogenesis. As a result of this study, an index of maturity relating the macroscopic appearance of the gonad to recognizable stages in its histology has been established. The index is used, in a subsequent paper, to elucidate the breeding biology of billfish in the equatorial western Indian Ocean (Merrett, in prep.).
Materials and methods Specimens of the following species were examined. Sailfish, Istiophorus platypterus (Shaw & Nodder), 1791 : 12$ (39-92 lb), 169 (30-107 lb). Striped marlin, Tetrapterus audax Philippi, 1887: 15$ (63-151 Ib), 319 (43-163 Ib). Shortbill spearfish, Tetrapterus angustirostris Tanaka, 1914: 1 i3 (30 lb), 39 (41-44 lb). Blue marlin, Makaira nigricans LacCp2de, 1803. O$ (-), 2 9 (164-500 Ib). Black marlin, Maikaira indica (Cuvier), 1831 : 2 8 (172-189 Ib), 59 (107-785 Ib). The fish were captured in the equatorial western Indian Ocean during a tuna longline fisheries
survey by the East African Marine Fisheries Research Organization, from September 1964 to December 1967. Preliminary investigation and removal of the urino-genital system from the specimens was carried out on capture aboard the research vessel. The body cavity was opened by a slit along the mid-ventral line, from the origin of the pectoral fins posteriorly to the anus. The gonads were removed by severing the mesenteric connections with the body wall from their anterior ends. Posteriorly, at the junction of the gonad pair, the ureters were followed dorsally and dissected out with a portion of the kidney. The whole was finally removed by severing the rectum but retaining a small section of the ventral body wall bearing the anus and urino-genital groove.
FIG. 1. Diagram of the gonad indicating the regions sampled for histological examination.
Three 1 cm cube samples for histological examination were removed from different levels of the gonads (Fig. 1). These were fixed in Bouin 15 (Allen, 1916) for 2 h and, after washing in running sea-water for 24 h, were transferred to 5 % formol-saline. The tissues were dehydrated in graded alcohols, cleared, embedded in wax, and sectioned at 8 pm. The sections were examined histologically after staining with haematoxylin and Orange G ; and histochemically for carbo- hydrates and nucleic acids with haematoxylin, Orange G and Alcian blue, and haematoxylin, 0range.G and periodic acid Schiff, and finally with a combination of all four stains. The Feulgen

G O N A D D E V E L O P M E N T I N BILLFISH 357
nuclear reaction (Feulgen & Rossenbeck, 1924) was also used on the sections. The complete failure of this technique is thought to be due to insufficient buffering of the formol-saline.
Ova diameter measurements in the maturing ovary were made from the slides used for histo- logical examination, with a tacit realization that a shrinkage factor was inevitably involved (see below). Sixty ova, 30 each from two separate slides were measured. Only the largest ova were considered, and their maximum length and breadth were measured with a micrometer eye piece. The product of the two diameters was then taken to represent the surface area of the ovum. The mean ovum diameter of the sample was calculated as the square root of the mean area. It was reasoned that, by selecting only the largest ova and by taking the maximum measurements, errors introduced by fixation-shrinkage artefacts would be corrected. Further, the fact that the area is calculated on the basis of a square rather than a circle, would also correct for this shrinkage (approximately 20%, ROSS, 1953). Similarly it was assumed that by taking a large sample from each fish, anomalies, due to the fact that few ova would be sectioned at their maximum diameter, would also be corrected in the results.
Measurements of the diameters of eggs shed on capture by running ripe female I. platypterus and T. angustirostris were made with a micrometer eyepiece. All examinations were made in formol-saline.
Morphology
The gonads are paired organs lying in the posterior half of the body cavity, on each side of the stomach and intestine. They are suspended from the lateral edges of the cham- bered air bladder by mesenteries (Plate I(a)). In all istiophorid species examined from this area, with the exception of T. angustirostris, the gonads are almost bilaterally symmetrical and both terminate at their point of discharge to the exterior in the urino-genital papilla. In T. angustirostris the gonads are Y-shaped, with the right organ shorter and its posterior end joined to the left. The junction is at the level of the urino-genital aperture. The left gonad is prolonged posteriorly (Plate I(b)).
The products of the gonads are discharged from the urino-genital papilla which lies posterior to, and in a common groove with, the anus (Plates I(c) and II(a),(b)). The urinary and genital systems are closely linked. The kidneys lie against the vertebral column, in a dorsal position in the body cavity. They run from the anterior end of the body cavity to a point approximately perpendicular to the posterior end of the groove containing the anus and urino-genital papilla. At the posterior end the kidneys are united, and the ureters arise from this at a point vertically above to the middle of the common groove.
The relation between the urinary and genital systems is similar in the species examined. It is shown in situ in Plate I(a) for female T. audax, and dissected from the body cavity in ventral view for female T. angustirostris in Plate I(b). Further dissection of the urinary system is shown in Plate I(c) for female T. audax. The ureters arise as separate tubes which unite at a short distance from the kidney. They are invested by a single muscular sheath which continues over the combined ureter. The single tract passes through the chambered air bladder. It then becomes dilated and curves over the right gonad, in the length of which the dilation increases to form a urinary bladder terminating at the base of the wino-genital papilla. The bladder discharges through a short duct in the urine-genital papilla.
The ovaries are elongate sausage-shaped organs, which taper at both ends, and joined only at their posterior ends (see Plate I(a),(b)). A strong muscular sheath binds the ovaries

358 N. R . M E R R E T T
to the wino-genital papilla and the basal part of the intestine. They are invested in thick layers of connective tissue which sometimes contain deposits of fat ; fat is also occasionally found in the mesovarium. Beneath the connective tissue the ovaries are pale flesh-pink to wine red in colour, depending upon the stage of maturity. Internally, at certain stages, a central lumen runs the length of the ovary. In all species investigated in this study, the ovaries have been found to be unequal in length, which is in agreement with the findings of La Monte (1958). Measurements of the ovary lengths of specimens of I . platypterus, T. augustirostris, and T. audax are given in Table I. The results for the last named species may be compared with similar measurements given by Williams (1968). These measurements confirm that in different individuals either ovary can be the longer of a pair.
TABLE I
Gonad length measurements on certain istiophorids
Length Weight Testis Ovary Species Date (cm) (Ib) Left Right Left Right
I. platypterus 9 24/4/67 22/4/67 22/4/67 16/9/67
d 23/4/67 29/5/66
T. angustirostris ? 2/7/65 24/7/67
T, audax 9 15/2/67 23/4/66 16/2/67 23/4/66 11/7/66 11 110167
? 15/2/67 21 14/66 15/2/67 21 14/66 24/4/66
M. indica 0" 29/5/66
45.5+129.5 47-5+ 142.0 53*5+159*5 59.5+173*0
45.0+ 172.0 58-0+ 181 -0
30*0+139*0 22*5+ 139.0
47*0+140*5 51 *O+ 154.0 55*5+170.5 56*5+177*5 55-5 + 170.0 53 -5 + 168.0
47-5+ 147-5 45*0+142*5 48.5+148*0 55-0+153.0 50-5f158.5
60.0+181.5
30 40 56 84
39 92
- 41
53 81
112 113 116 126
63 64 72 76 82
1 89
13.3 14.0 18.0 19.0 21.0 25.0 31.5 26-3
- - - -
43.7 27.0 53.5 34.0
6.1 9.2 15.0 15.5 14.2 21.5 14-0 17.0 13-5 18.6 22.0 15.0
- - - - - - - - - -
- -
The relations between the urino-genital papilla and the anus of resting female T. audax and M . indica is shown in Plate I(c) and Fig. 2 respectively. In each case the common groove enclosing these structures has been bisected longitudinally. It will be noticed that within the groove, the anal papilla is the most anterior and ventrally placed. Immediately

G O N A D DEVELOPMENT IN BILLFISH 359
posterior to its base is the urino-genital papilla. In the female this carries only the urinary duct. The point of discharge of the ovaries is situated between the bases of the anal papilla and the urino-genital papilla. In the resting female T. uudux (Plate I(c)), a duct is visible but occluded at its dorsal end where it joins the inactive ovary. In an immature specimen no duct is yet visible (Fig. 2).
1.0
b I
UP a
FIG. 2. Sagittal section of the urino-genital groove and associated structures of a female M. indicu. (Explanation to abbreviations see p. 370.)
The running ripe stage of the ovary and its relations with the structures discussed above is shown for T. ungustirostris in Fig. 3, which shows the two ovaries at the level of their point of discharge. The anterior portions of both ovaries have been removed and the posterior end of the left ovary. Both ovaries have been cut away to show the oviducts in their ventral walls. It will be noticed that the lumen of the ovary is much enlarged. This is caused by differential ripening and discharge of the eggs. Those eggs situated close to the lumen ripen first and in this case have been discharged and make way for the successive ripening of eggs outwards to the periphery. The posterior end of the intestine is shown ventral to the common wall of the ovaries. The bladder is shown in its natural position posterior to the end of the right gonad. This curves towards the mid-line, narrow- ing into the ureter, which then runs vertically through the air bladder to enter the ventral surface of the kidney. Plate I(d) shows a transverse section of the two ovaries of a specimen of T. ungustirostris cut at the level of the point of discharge of the oviducts. This specimen is less mature than that described above, at a stage prior to ovulation.
The urinary system of male specimens of all species is directly similar to that already described. The testes, however, vary greatly in appearance from the ovaries, aIthough

3 60 N. R. MERRETT
their relation with the other internal organs is the same. The testes are elongated organs, triangular in cross section, and much thinner than the ovaries. Unlike the ovaries they are not invested by a muscular tunic. The surface of the testis is convolute, deeply so in mature fish, giving a lobular appearance in all species. The colour of the testis varies
I.0-
‘ k
.re
r.od
FIG. 3. Dissection of the ovaries of T. angustirostris at ovulation, shown in relation totheurinarysystem.(Explana- tion to abbreviations see p. 370.)
according to its maturity, ranging from pink, in the inactive stage, to pinkish-white when it is ripe. As in the case of the ovaries, the testes are usually subequal in length. Testis measurements for I. platypterus, T. audax and M . indica are shown in Table I. A vas deferens runs the length of each testis. The vasa deferentia converge at the posterior end of the testes, dilating to form a seminal vesicle. A short canal leads from the seminal vesicle to the genital pore at the tip of the urino-genital papilla, just anterior to the urinary pore. The relations of these structures in male T. audux and T. angustirostris are shown in longitudinal section in Plate II(a),(b).

G O N A D D E V E L O P M E N T I N B I L L F I S H 361
Histology
The testis
The immature testis is a slender ribbon-like structure. The surface bears slight convolu- tions, which increase in size and number considerably as maturation proceeds. Several thin sheets of connective tissue invest the entire surface of the organ. No specimen of the most immature stage of the testis has been obtained for histological examinations; the least advanced examined shows the beginning of tubule formation.
Plate III(a) shows this stage in the testis of M . indica, in transverse section, where separate lobes formed by the convolutions of the wall are visible. Each lobe is comprised of fibrous connective tissue surrounding lumina of diverse shapes and sizes. The fibres vary in size, but in shape they are elongated with spindle-shaped or ovoid nuclei. Small spherules of chromatin are visible in the basophilic nuclei. One or two nucleoli may be present. Primary spermatogonia occur within the lumina. They are arranged peripherally, and lie in a syncytium of Sertoli cells. The latter fill the central portion of the smaller lumina. The nuclei of the Sertoli cells are round to oval and 8-10 pm in diameter, and are a similar size in all species. Within the nuclei, strands of chromatin lie against the nuclear membrane. The primary spermatogonia are strongly basophilic, the cell membrane being readily stained with iron-haematoxylin. Granular chromatin occurs close to the mem- branes of the large spherical nuclei. The nuclei are similar in size: 5-6 pm in diameter in those species in which they were observed; i.e. I . platypterus, T. audax and M. indica. With development, the lumina containing the primary spermatogonia increase in size to form the tubules of the maturing testis.
At this stage in development the gonad blood supply, which previously consisted of a pair of unbranched blood vessels running through the testis, develop further. Each lobular convolution of the testis contains its own blood vessels, supplied by an afferent artery and an efferent vein situated towards the dorsal side of the testis. However, extra- vascular blood corpuscles are very common and are widely distributed through the con- nective tissue of the cell walls.
As maturation progresses the immature testis increases in size. Tubule formation is soon completed, and well defined spermatic ducts differentiate to channel the spermatozoa from the lobules of each convolution to the vas deferens. During the early stages the spermatic ducts act as spermatogenic tubules, as does the vas deferens. The latter runs close to the testicular blood vessels, towards the dorsal side. The diameters of the sperma- togenic tubules in early development are approximately 140-170 pm, with walls of 5 pm in thickness. With continued maturation the diameters of the tubules increase and the walls become thinner. The connective tissue described above forms the walls of the tubules.
The maturation of primary spermatogonia within the tubules is apparently rapid as, apart from the specimen of M. indica described above, all testes examined have contained cells in all stages of spermatogenesis (Plate III(b),(c)). The primary spermatogonia present in the connective tissue during early development decrease rapidly in number with maturation. Also during early development secondary spermatogonia are numerous. The cell outlines of the secondary spermatogonia are difficult to distinguish. Their nuclei are, however, dense and measure 3.5-4.5 pm in all species examined (no testis of

362 N. R. M E R R E T T
M. nigricans was available for histological examination). As maturation proceeds the second- ary spermatogonia divide to form primary spermatocytes, and soon these become more numerous than the former. Primary spermatocytes may be recognized by the dense chromatin within the nucleus, and their smaller size. They are about 3 pm in diameter in all the species examined. Bowers & Holliday (1961) showed that, in the spermatogenesis of the herring, the organization of the nucleus of the spermatocytes varied between groups of these cells. This has also been observed in the testes of the Istiophoridae. Certain groups of cells show chromatin appearing as a dense meshwork of threads, presumably in the late prophase of meiotic division, and these are the most numerous. In other groups, the chromatin is distributed around the periphery of the nuclei; while in still others it is mainly concentrated to one side, but with some chromosomes still visible towards the centre of the nuclei. These changes in nuclear organization are regarded as due to maturation. The phases observed in all the istiophorids species are similar to those described for the perch by Turner (1919), and for the herring by Bowers & Holliday (1 961).
Primary spermatocytes give rise to secondary spermatocytes. The nuclei are less dis- persed in the secondary phase than in the earlier generation and are often kidney-shaped, measuring 2-0-2.6 pm in diameter in all species examined. No nucleoli have been observed. Secondary spermatocytes appear less common than the primary phase, which suggests that they exist for a shorter period of time.
Each successive generation of cells formed in the process of spermatogenesis has been observed to be situated more centrally in the lumen of the tubule than the previous genera- tion. The spermatids, therefore, are found close to the centre of the lumen of the tubule. Their nuclei are spherical, stain densely, and are approximately half the size of those of the secondary spermatocytes, about 1 -3 pm in all species examined. Most centrally placed in the lumen of the tubule are the spermatozoa. Their head-pieces vary little in size from the nuclei of the spermatids, and they are not easily distinguished from the latter as, in prepared sections of testis, their tails are not readily apparent.
The morphology of the spermatozoa of the istiophorid species is more clearly studied from a smear of the fresh milt. The spermatozoa of I . platypterus, T. audas and M . irtdica were examined and found to be similar. Measurements showed the heads to be 1 -3-1 *9 pm in diameter; in length, the middle pieces measured 0.8-1.3 pm, and the tails up to 30 pm. The only difference observed between the species was that the heads of the spermatozoa in I . plurypterus tended to be in the upper part of the diameter range, and the middle pieces were found generally to be shorter than in the other two species.
In the course of this study it was observed that spermatozoa were present in testes of all sizes to a greater or lesser extent. However, no fish was caught with a swollen, white testis turgid with milt, contrary to observations by de Sylva & Davis (1963) on the white marlin, Terrprerus albidus. Yet, differential maturation of spermatozoa was observed in cer- tain lobules along the length of the testis. It will be recalled from the description of the mor- phology of the testis that the species examined have a substantial seminal vesicle. This vesicle has muscular walls, which are thought to assist in the expulsion of milt with ensuing passive replenishment of the vesicle by suction from the testis, without the need for the latter to be in a turgid condition. Since, during maturation, the production of spermatozoa is a continuous process such a mechanism seems feasible, and would accord with the observed differential ripening of the testicular lobes.

- cy 3 b
PLA
TE I. (a
) B
ody
cavi
ty o
f a
fem
ale T. uu
ci'u
x, w
ith o
varie
s in
a r
ecov
erin
g sp
ent c
ondi
tion.
(b)
Urin
o-ge
nita
l sys
tem
of a
fem
ale
T. a
ngus
tiros
rris
, with
ova
ries
in a
con
ditio
n ju
st p
rior
to o
vula
tion.
Ven
tral
vicw
, with
kid
ney
and
uret
er d
efle
cted
to t
he r
ight
. (c
) U
rino-
gcni
tal
syst
em o
f a
fem
ale
T. a
rrdu
x. S
agitt
al s
ectio
n. (
d) T
rans
vers
e se
ctio
n of
the
ova
ry p
air
of th
e T
. anE
ustir
osfr
is s
how
n in
Pla
te 1
1, at
the
leve
l of
the
ovid
ucts
.
3 8
h - W
4 0
v
LL
o\ 0

J. Zool., Lonrl. (1970) 160
PLATE 11. (a) Sagittal section through thc urino-genital groovc of a male T. nrtdu.~. (b) Sagittal scction through the urino-genital groove of a male T. artgusfirostrts.

h
h
e
PLATE 1
11. (
a) S
perm
atog
onia
deve
lopm
ent i
n an
imm
atur
e virg
in te
stis
of M
. irt
dicu
. Tra
nsve
rse s
ectio
n. (b
) Ger
mce
ll m
atur
atio
n in
the
test
is of
Z.p
/ufj
pter
us. T
rans
vers
e sec
tion.
(c)
Ger
m c
ell m
atur
atio
n in
the
test
is of
T. u
udux
. Tr
ansv
erse
sect
ion.
(d)
Med
ian
W 4
0
Y
r
QI
0
sect
ion
of th
e ant
erio
r end
of t
he r
ecov
erin
g spe
nt o
vary
of M
. ind
icu.

J. Zool., Lond. (1970) 160

J . Zool., Lotrrl. (1970) 160
PLATE V. (a) Early stage in chorion formation in thc egg of T. arrdus, showing a peripheral ring of dense cyto- plasm surrounding the nucleus, and cytoplasmic strands from the thcca externa to the connective tissue. Transverse section. (b) Section of the chorion of a maturing egg of I . plutypterrus.

PLAT
E VI.
(a),(
b),(c
) Su
cces
sive
stag
es o
f nu
clea
r en
croa
chm
ent
in t
he m
atur
ing
egg
of T
. ang
usfir
osfr
is. T
rans
vers
e se
ctio
ns.
(d) H
omog
enou
s dis
pers
al o
f yol
k un
its th
roug
h th
e cy
topl
asm
of
the
mat
urin
g eg
g of
I. p
/u/y
pter
us. T
rans
vers
e se
ctio
n.

G O N A D D E V E L O P M E N T IN B I L L F I S H 363
The ovary This organ is invested by a tunic of fibrous connective tissue, through which run groups
of smooth muscle fibres. The external surface is covered by a thin sheet of peritoneum. External to this, the multi-layered mesentery suspends the ovary from the dorsal wall of the body cavity. Internally, many branched and incomplete fibrous connective tissue septa arise. The main septa run approximately radially towards the centre of the ovary and, at maturity, discharge into the oviduct which is effectively an intra-ovarian cavity. Within the septa run the branches of the ovarian blood vessels. Plate III(d) show the arrange- ment of the septa in longitudinal section in the anterior end of the ripening ovary of a female M . indica. A more advanced stage of ovary ripeness is shown in Plate IV(a) (anterior end, longitudinal section) T. audux. The septa are at this stage thinner and a lumen has formed in the core of the ovary. The ovarian arteries and veins can also be seen in this sect ion.
In the immature ovary, in T. audax and M . indica, the tunic has been found to vary from 120 pm to 340 pm in thickness. The septa vary greatly in thickness, arising from a broad base at the tunic and becoming rapidly thinner as they run into the lumen. Within the separate lumina formed between the septa are the regions of non-fibrous connective tissue stroma in which the oogenic cells develop (Plate IV(b)). Blood capillaries ramify through this stroma.
The earliest stages in the immature ovary were not available for histological examination. The most immature stages observed were from the specimens mentioned above. In these, development of oocytes within the non-fibrous connective tissue stroma was nearly complete. However, in certain areas oogonia are visible which give rise, by mitotic division, to the oocytes. The oogonia are 10-23 pm in diameter in these species, with large rounded nuclei, 7-14 pm in diameter. Within the nuclei, centrally placed nucleoli occur from which chromatin threads radiate to the nuclear membranes. The oogonia are not prominent as they do not take up stain readily (Plate IV(c)). These cells are embedded in the non-fibrous connective tissue. The cells of the latter are much smaller and have spherical nuclei 2.5-3.0 pm in diameter in all species. In contrast, the nuclei of the fibrous connective tissue comprising the septa, may be distinguished by their spindle-shaped appearance and larger size, 3-7 pm in all species.
Primary oocytes are readily distinguished from the oogonia by the strongly basophilic cytoplasm which stains darkly with haematoxylin. Changes can also be observed in the nuclear contents. The nucleoli of the oocytes become situated around the periphery of the nuclei, and chromatin threads are observed centrally. The smallest oocytes observed in the species considered, ranged from 15 to 30 pm in cell diameter, and from 9 to 15 pm in nucleus diameter. The growth of the primary oocytes results in their increased pro- trusion into the lumen of the ovary. Their arrangement becomes more regular and each lies in a well-defined vacuole, the walls of which are formed by the fibrous connective tissue of the septum. At this early stage, when the primary oocytes are between 20 and 30 pm in diameter, they are seen to be surrounded by a layer of connective tissue in which a few small flattened nuclei occur ( 4 5 pm in all species). From this connective tissue coat, the theca externa of the future follicle, several cytoplasmic strands can be seen linking the oocyte with the connective tissue septa. These are considered to form the means whereby the developing oocytes obtain their blood supply (Bowers & Holliday, 1961).
28

364 N. R . M E R R E T T
Growth of the oocytes continues until they are 40-50 pm in diameter. At this stage a certain proportion of the oocytes are seen to have developed a second enveloping layer. This consists of a single layer of cells internal to the connective tissue theca. The cells are similar in appearance to the non-fibrous connective tissue, with spherical nuclei 2.5- 3.0 pm in diameter. Chromatin can be seen situated peripherally within the nuclear membranes of these cells. In surface view the cells are polygonal in shape, 10-17 pm in length and have nuclei of 3-6 pm in diameter. In tangential view they are rectangular. Their cytoplasm is strongly basophilic. In their account of oogenesis in the herring, Bowers & Holliday (1961) suggests that the polygonal cells are, in fact, non-fibrous con- nective tissue cells, which have migrated through the strand of connective tissue. This latter is now seen to be a very thin double layer of anatomically similar cells comprising the theca interna and the theca externa of the mature follicle.
Those oocytes in which the granulosa layer has been formed are observed at this stage to lie in septa1 vacuoles larger than those containing oocytes in which the granulosa layer is absent (Plate IV(d)). According to Bowers & Holliday (1961) the former are destined to develop further to mature follicles, whereas the latter increase only a little more in size during the rest of the ovarian cycle and are the resting oocytes of the mature ovary.
During this period of development more oocytes are being formed, and growth of the whole ovary continues. The ovary is pink in colour due to the high concentration of blood vessels. The formation of yolk granules begins in the cytoplasm of oocytes that have reached a diameter of 100-110 pm in all species. The nuclear diameter at this stage is about 40 pm. The spherical to ovoid yolk globules gradually enlarge to form yolk vesicles. All increase in number. When the largest is about 5 pm in diameter they lie in a ring around the nucleus, closer to the plasma membrane than to the nuclear membrane. However, even at this stage there is always a ring of very small yolk globules around the periphery. With yolk formation the developing oocytes can be clearly distinguished by their larger size from the resting oocytes. Since yolk formation only occurs in oocytes in which a granulosa layer has been laid down, it has been suggested (Bowers & Holliday, 1961) that the presence of granulosa cells is responsible for the formation of yolk and, subsequently, of the chorion.
At this stage in yolk formation the chorion makes its first appearance. Initially, the peripheral cytoplasm of the oocytes is more dense than the remainder. A thin wall can then be distinguished external to the dense layer, inside which a second and a third layer are soon laid down to complete the chorion structure. All three layers thicken as develop- ment of the oocyte proceeds and all have a laminar structure traversed by faint radial striations.
The follicle of the egg is now fully formed. Internally, the nucleus, which is centrally placed, is seen to have a dense layer of cytoplasm around it, which itself is surrounded by large yolk vesicles (Plate V(a)). The eggs at this stage are visible to the naked eye.
Subsequent to the full differentiation of the follicle, a phase of growth occurs resulting in the ovaries occupying an increasing volume of the body cavity. In contrast to the continued production of spermatozoa during the maturation of the testis, this and subse- quent enlargement of the ovary is due solely to the growth of existing oocytes. At this stage, secondary yolk formation takes place in the developing oocytes. The development of the resting oocytes has been arrested prior to primary yolk formation and consequently

G O N A D D E V E L O P M E N T I N B I L L F I S H 365
the mean diameter of these never exceeds 100 pm. They remain at this stage until the next oogenic cycle begins.
As secondary yolk formation occurs in all regions of the cytoplasm in the developing oocytes, the dense layer of cytoplasm around the nucleus disperses. With the growth of the egg, the follicle increases in size, by increase in numbers of granulosa cells and the connective tissue cells of the theca externa. The chorion (Plate V(b)) also continues to thicken, and by the time the eggs reach a diameter of 500-600 pm it is between 8 pm and 14 pm thick. At this stage, when the chorion is examined in surface view, the radial striae are seen to consist of pores traversing it. (Observations on the surface of the ovulated eggs show these pores to be from 0.66 pm to I .98 pm apart.) Also at this point of develop- ment, changes appear to take place in the nature of the nuclear membrane. Small yolk granules accumulate between the many yolk vesicles which have now appeared in the cytoplasm. Some of these granules lie close to the nuclear membrane and seem partly to penetrate it. This phenomenon, also observed in the herring (Polder, 1961) and in the goldfish (Beach, 1959), has been seen in all istiophorid species with this size of egg, i.e. T. audax, I. plarypterus and T. angustirostris. It will be noted from Plate VI(a),(b),(c) that in the case of T. angustirostris this nuclear encroachment has, in some instances, reached such a point that the nucleus is recognized only by the nucleoli which now lie among the yolk granules.
The yolk vesicles have, by now, enlarged considerably. Vesicles of up to 100 pm in diameter have been observed in the eggs of I. platypterus, and up to 70 pm in T. angusti- rostris. However, in both these species and also T. audax, the majority of the yolk globules present measure only about 20 pm in diameter. In I . platypterus the yolk units are homo- geneously disposed throughout the cytoplasm (Plate VI(d)). In T. angustirostris, however, a perinuclear U-shaped vesicle is present and the distribution of the yolk granules is restricted to the periphery of the vesicular membrane (Plate VII(a)). Internally this vesicle appears to have a micro-granular structure, which stains intensely with haematoxy- lin, but negatively with periodic acid Schiff and Alcian blue. It is, therefore, inferred that both neutral and acid polysaccharides are absent from the vesicle, and that it probably has a high protein content. This distinction between the ovarian eggs of the two species no longer exists in the post-ovulated egg, by which time the perinuclear vesicle can be observed in I. platyptertrs, also.
Although a continuous pattern of development was traced from the immature ovary to a point at which the eggs measure 750 pm the next ovarian stage represented in the collections was ovulation. At capture, eggs shed by T. angustirostris and I. platypterus were collected and measured. Figure 4 shows the distribution curves of the diameters of the eggs of each. The mean diameter of the eggs of T. angustirostris, I -442 mm (S = 0-051), was found to be larger than that of the eggs of I. platypterus, 1 +304 mm ( S = 0.043). In comparison to the mean diameter of the ovulated eggs of T. angustirostris, a sample of 103 unovulated eggs from the periphery of the gonad was measured and found to have a mean diameter of 845 pm. This may be compared with the measurement of 800-850 pm for the unovulated eggs from this species by Nakamura (1949). The gap in the data thus represents about 100 pm of growth in the maturing eggs.
Ovulation marks the final phase in the development of the egg. Immediately prior to this, the ovary becomes more vascular and this can be observed macroscopically. The follicles, both theca externa and granulosa layers, then burst. On bursting, the released
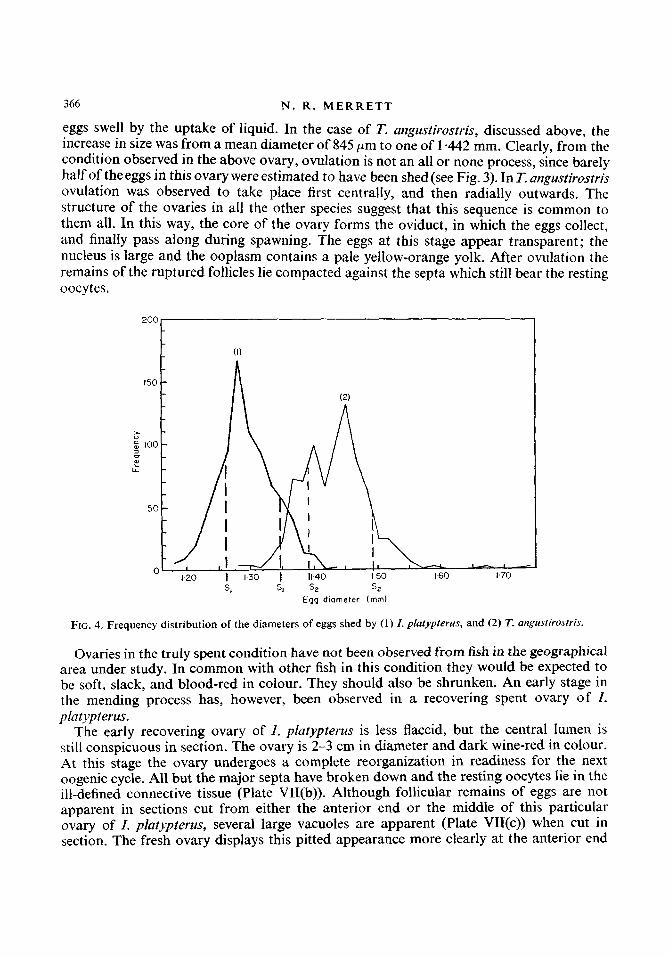
366 N . R . M E R R E T T
eggs swell by the uptake of liquid. In the case of T. angustirostris, discussed above, the increase in size was from a mean diameter of 845 pm to one of 1 -442 mm. Clearly, from the condition observed in the above ovary, ovulation is not an all or none process, since barely half of the eggs in this ovary were estimated to have been shed (see Fig. 3). In T. angustirostris ovulation was observed to take place first centrally, and then radially outwards. The structure of the ovaries in all the other species suggest that this sequence is common to them all. In this way, the core of the ovary forms the oviduct, in which the eggs collect, and finally pass along during spawning. The eggs at this stage appear transparent; the nucleus is large and the ooplasm contains a pale yellow-orange yolk. After ovulation the remains of the ruptured follicles lie compacted against the septa which still bear the resting oocytes.
FIG. 4. Frequency distribution of the diameters of eggs shed by (1) I . platypterris, and (2) T. artgrcstirostris.
Ovaries in the truly spent condition have not been observed from fish in the geographical area under study. In common with other fish in this condition they would be expected to be soft, slack, and blood-red in colour. They should also be shrunken. An early stage in the mending process has, however, been observed in a recovering spent ovary of I . platypterus.
The early recovering ovary of 1. platypterus is less flaccid, but the central lumen is still conspicuous in section. The ovary is 2-3 cm in diameter and dark wine-red in colour. At this stage the ovary undergoes a complete reorganization in readiness for the next oogenic cycle. All but the major septa have broken down and the resting oocytes lie in the ill-defined connective tissue (Plate VII(b)). Although follicular remains of eggs are not apparent in sections cut from either the anterior end or the middle of this particular ovary of I. platypterus, several large vacuoles are apparent (Plate VII(c)) when cut in section. The fresh ovary displays this pitted appearance more clearly at the anterior end

G O N A D D E V E L O P M E N T I N B I L L F I S H 367
than at the middle. The diameter of the vacuoles in the prepared sections measure up to 1.4 mm. On the basis of their shape and general appearance, it is concluded that these spaces were once occupied by ripe eggs. An opaque substance, staining puplish with haematoxylin, lines the vacuoles. A similar, if not identical, substance pervades the loose tissue of the ovary. enveloping many of the resting oocytes. Equally, this may represent another stage in the fate of the structure which Nakamura (1949) somewhat non-com- mittally referred to as the “nimbus”. Quite clearly, this is a point in the oocytology of these fish which requires more extended and careful investigation.
This particular ovary is the only one in which developmental differences have been observed in the eggs from the two levels of the ovary at which sections were cut. At the anterior end, the tissue of the ovary appears more compact, and several collapsed septa are visible (Plate VlI(b)). Within some of these septa, areas of activity can be observed, involving concentrations of nuclei. It would seem that these are about to form new septa, as has been described for the herring (Polder, 1961). The oocytes in this part of the ovary are rounded and have a mean diameter of 58 pm. Few nuclei can be observed within the cytoplasm which appears to be in a state of degeneration.
Sections from the centre of this ovary (Plate VII(d)) show more intense activity. The septa are less buckled and the connective tissue is seen to be in the form of a loose mesh- work, within which many nuclei and phagocytic cells occur. The majority of the resting oocytes do not appear rounded in outline. They are angular, with cytoplasmic connections to the connective tissue. Many are devoid of nuclei; others have their cytoplasm extending into the connective tissue (Plate VIII(a)). The basophilic purple staining substance (“nim- bus”?) is still visible around certain oocytes. The developing germ cells in this region are larger in mean diameter (64 pm) than those of the anterior end, the difference, however, is not great, and there is a suggestion that at least a partial resorption of the resting oocytes, as well as the residuum of the recently ovulated eggs, is now occurring. This proposition i s supported by the fact that the mean diameter of the largest resting oocytes in the later stages of oogenesis is between 90 and 100 pm, whereas the mean diameter of the oocytes in this spent ovary is considerably smaller, and their actual sizes more uniform. This suggests that regressive changes are already in train. The structural differences at the two levels of this ovary are considered to reflect differences in ovulatory stages. As the central core of the ovary forms the oviduct, ovulation must proceed both centrifugally and postero- anteriorly.
A further stage in the recovery of the ovary is shown in Plate VlII(b) for T. nudax. The structure of the cytoplasm has reorganized and no longer gives the purple basophilic stain reaction. However, the undeveloped oocytes of the previous spawning remain deeply stained. Many are surrounded or capped by equally basophilic material, with interconnecting cytoplasmic strands.
Egg follicles in the process of being resorbed have been observed in sections of the ovary of T. angustirostris (Plate VIII(c)). Here the mean egg diameter is 121 pm, and yolk formation is already proceeding. However, although reorganization of the ovary is almost complete, certain cells with resorbed material are still visible.
The thickness of the ovary wall during the oogenic cycle is extremely variable, and throughout the cycle, in all species, has been found to measure between 35 and 1700 pm. Variations have also been observed within a single ovary. It would appear that develop- ment of the ovary wall from the immature state. involves an initiaI thickening, but that

368 N. R. M E R R E T T
this growth then stops and the subsequent fate of the wall involves a progressive attenuation due to tension exerted by the expanding, developing germ cells. This tension, of course, is released at spawning, as a result of which the wall simulates thickening. The subsequent egg cycles involve a repetition of this process.
Discussion The relation of the gonads to the other internal organs in billfish follows the same pattern
as in other teleosts. The urinary tract is closely connected with the posterior part of the gonads. Although the products of excretion and reproduction leave the body through separate ducts, they do so into a common groove with the anus. The external appearance of the billfish testis is characteristically convoluted; but the appearance of the ovary (excepting T. angustirostris) is similar to other teleosts.
The course of gametogenesis has not been described in detail for many teleosts. How- ever, this process in billfish is similar in general to those descriptions available. The gametes arise from germ cells that are similar in both sexes. They are invested by similar connective tissue which differentiates in both males and females to form tubules and follicles, con- taining spermatozoa and ova respectively. There is a continual production of spermatozoa within the tubules of the testis during maturation, while in the ovary maturation is con- cerned with the growth of existing oocytes. The observations in the present study further suggest a continuous availability of spermatozoa in mature males by the differential maturation of the testicular lobules in conjunction with the possession of a muscular seminal vesicle. The male cycle appears, therefore, to contrast with the case in other teleosts. The female cycle is similar to other teleosts however, as maturation is synchronous through- out the ovary.
Summary The relation of the reproductive and excretory organs is discussed, and the morphology
The histological changes within the gonads during gametogenesis is described in detail. The immature testis is lobular, consisting of connective tissue in which lie primary
spermatogonia. The latter surround lumina of diverse shapes and sizes. Differentiation of the lumina occurs to form a complex of tubules opening into a vas deferens within each testis. Spermatogenesis occurs within these ducts.
Some spermatozoa are present early in the maturity cycle; and although there is a small overall increase in size in the testis, differential maturation of the lobules occurs.
Spermatozoa are stored in a muscular seminal vesicle which is thought to be capable of the active expulsion of milt.
The immature ovary contains oogonia and oocytes lying within connective tissue septa projecting radially into the centre of the ovary. As development takes place the oocytes are encapsulated by follicles consisting of an outer layer of connective tissue and an inner layer of granulcsa cells.
After the formation of the follicle, the deposition of yolk begins and the chorion is laid down. At this stage the fate of the oocytes is decided, as growth continues in the maturing oocytes only. Resting oocytes are the more numerous but develop no further. The follow- ing increase in size of the ovary is due solely to the growth of the existing oocytes destined for maturation.
of the gonads and urinary system described for five species of billfish.

G O N A D D E V E L O P M E N T I N B I L L F I S H 369
Ovulation occurs by the bursting of the follicles. This apparently takes place at first centrally and then radially outwards in the ovary, with the central lumen so formed acting as an oviduct.
After spawning the ovary undergoes reorganization in readiness for the next oogenic cycle. Evidence is put forward to suggest a partial resorption of the resting oocytes as well as the residuum of the recently ovulated eggs.
The thickness of the ovary wall throughout the cycle is variable, but is considered to be related to the phases undergone beneath it.
Professor 1. Griffiths of the University College, Dar es Salaam, Tanzania, kindly afforded me facilities in the Zoological Department. I am also indebted to him for his encouragement and helpful criticism throughout the study. My thanks are due to the Director and Staff of the East African Marine Fisheries Organization, and to the Captain and crew of the F.R.V. “Manihine” for facilitating the collection of the data and other aspects of the work. I am grateful, also, to the Malindi Sea Fishing Club for permitting me to collect data and samples from specimens caught during the 1967 Billfish Tournament. Finally, I wish to thank Dr P. H. Greenwood, of the British Museum (Natural History), for kindly reading the manuscript.
R E F E R E N C E S
Allen, E. (1916). Studies on cell division in the albino rat (Mus norvegicus, var. alba). I1 Experiments on technique, with description of a method for demonstrating the cytological details of dividing cells in brain and testis. Anat. Rec. 10: 565-589.
Beach, A. W. (1959). Seasonal changes in the cytology of the ovary and of the pituitary gland of the goldfish.
Bowers, A. B. & Holliday, F.G.T. (1961). Histological changes in the gonad associated with the reproductive cycle in the herring (Clupea harengus L.). Mar. Res. 5: 1-16.
de Sylva, D. P. & Davis, W. P. (1963). White marlin, Terrapterus albidus, in the middle Atlantic Bight, with observa- tions on the hydrography of the fishing grounds. Copeia 1963: 81-99.
Feulgen, R. & Rossenbeck, R. (1924). Mikroskopisch-chemischer Nachweis einer Nucleinsaure von Typus der Thymonucleinsaure und auf die darauf beruhende elective Farbung von Zellkernen im mikroskopischen Praparaten. Z. physiol. Chem. 135: 203.
La Monte. F. R. (1958). On the biology of the Atlantic marlins, Ilfukaira anipla (Poey) and Afukaira albida (Poey): notes on the alimentary, excretory, and reproductive organs of Atlantic Makaira. BUN. Am. MUS. tzar. Hist. 114: 396401
Merrett, N. R. (In prep.). Aspects ofthe biology of billfish (Isfiophoridae) froriz the equatorial Western Indian Ocean. Nakamura, H. (1938). Report of an investigation of the spearfishes of Formosan waters. Rep. Tuiwun Govf-Genl.
Fhh. exp. sin. 1937 No. 10: 1-34. Translated from the Japanese language by W. G. Van Campen, Spec. Sci. Rep. US. Fish. Wildl. Serv.-Fish. No. 153: 1 4 6 1955
Nakamura, H. (1949). The I N ~ U S and rheirfisheries. Tokyo: Takeuchi shobo. Translated from Japanese by W. G . Van Campen, Spec. Sci. Rep. U S . Fish. Wild/. Serv.-Fi.sli. No. 82: 1-115 1952.
Polder, J. J. W. (1961). Cyclical changes in testis and ovary related to maturity stages in the North Sea herring, Clrrpea Irareirgus L. Arch n2erl. Zoo/. 14: 45-60.
Ross. K. F. A. (1953). Cell shrinkage caused by fixatives and paraffin-wax embedding in ordinary cytological preparations. Q . J I tnicrosc. Sci. 94: 125-139.
Turner, C. L. (1919). The seasonal cycle in the perch sperrnary. J. Morph. 32: 681-711. Williams, F. (1968). Longline fishing for tuna off the coast of East Africa. 1959-1960. Indian J. Fish. 10: 233-390.
CUT?. J. 2001. 37: 615-625.

3 70
a ab af b
ch cn CY CYS e fct g k 1.0 I.od lu m n nfct nl
C
nm 0 oc od og
N. R . M E R R E T T
Abbreviations used in plates and figures
anus chambered air bladder atretic follicle bladder capillary chorion chromatin cytoplasm cytoplasmic strand eggs fibrous connective tissue granulosa kidney left ovary left oviducal funnel lumen mesovarium nucleus non-fibrous connective tissue nucleolus 'nimbus' ovary oocyte oviducal funnel oogonium
PV re r.0 r.od ro
sd sg SP SPV
S sc
sv sz t
tce U UiS2 U g P UP
vd Yg YV
ta
V
perinuclear vesicle rectum right ovary right oviducal funnel resting oocyte stomach spermatocyt e spermatid primary spermatogonium septum septa1 vacuole seminal vesicle spermatozoa testis tunica theca externa ureter urino-genital groove urjno-genital papilla urpary pore vein vas deferens yolk globule yolk vesicle
Stains used in g o d sections illustrated
The sections shown in Plates III(a)-(c) and IV(a)-VIII(c) were stained with a combination of haematoxylin, Orange G, Alcian blue, and periodic acid Schiff-exept for sections shown in:
Plate III(b)--stained in haematoxylin and Orange G only. Plates 4(b), VI(a),(b )-stained in haematoxylin, Orange G, and Alcian blue. Plate VIII(a)-stained in haematoxylin, Orange G, and periodic acid Schiff.