germination preventing mechanisms in iris seeds
TRANSCRIPT

Annals of Botany 58, 551-561, 1986 551
Germination Preventing Mechanisms in Iris Seeds
A. BLUMENTHAL, H. R. LERNER, ELLA WERKER and ALEXANDRA POLJAKOFF-MA YBER
Department of Botany, The Hebrew University of Jerusalem, Jerusalem, 91904, Israel
Accepted: 2 April 1986
ABSTRACT
The different germination behaviour of the seeds of two irises, Iris lorteli and I. atropurpurea was found to be due to the different mechanical resistance of the integument, at the micropylar end, to radicle protrusion. A pressure of 135 atm was necessary in l. lorteli seeds for radicle protrusion, while in I. atropurpurea 77 atm was sufficient. In contrast Pancratium maritimum required a pressure of only 10 atm. The outer integument of seeds of l. lorteti was found to contain a compound which was toxic to the germinated embryo but did not act as a germination inhibitor. Extracts of the endosperm also had a slight germination inhibiting effect. An interaction between this weak inhibitor and the effect of the testa could not be ruled out completely. A test assay for germination using excised embryos was developed. A method for germination of Iris seeds, by cutting off the outer integument at the micropylar end, was developed and is being exploited commercially.
Key words: Iris lorteti, Iris atropurpurea, germination, germination inhibition, embryo culture, seed coat mechanical resistance.
INTRODUCTION
Seed dormancy is a common feature of the irises belonging to the Oncocyclus section. The germination of seeds ripening in a particular year is spread over very long periods. Even seeds ripening in the same capsule show great variability in their ability to germinate. In many of the irises only a small percentage of seeds germinate during the first year after ripening; most seeds germinate during the second and third year, while the rest of the seeds germinate intermittently over a period lasting up to 20 years after ripening (Randolph and Cox, 1943; Kidd, 1964; Jorgensen, 1965; Arditti and Pray, 1966, 1969; Avishai, 1977). As this section of irises grows mainly in the Mediterranean and the Irano-Turanian phytogeographic regions it seems possible that this strategy of germination may be adaptively advantageous in the climates of these regions.
The irises of the Oncocyclus section have large beautiful flowers and have high economic potential if they could be domesticated and bred for selection of desired characters. The widespread seed dormancy of these species is a barrier in breeding procedures. It would therefore be of great interest to understand the causes of their dormancy and to find ways to overcome it.
Amongst the Oncocyclus irises of Israel, Iris atropurpurea and I. lorteti represent opposite extremes of seed dormancy. Observations in the field have shown that while 60 per cent of I. atropurpurea seeds germinate in the first year after ripening, only one per cent of I. lorteti seeds germinate in the first year, and this only if the first year is very rainy. These two irises therefore provide interesting material for comparative investigations.
It has often been suggested that iris seeds contain an endogenous factor determining seed dormancy (Crocker, 1906; Randolph and Cox, 1943; Kartaschoff, 1958; Jorgensen, 1965; Arditti and Pray, 1969), however the mode of action of this dormancy factor is not clear. On the basis of information present in the literature two mechanisms appear
0305-7364/86/100551 + 11 $03.00/0 © 1986 Annals of Botany Company
at Frankfurt Univesity L
ibrary on August 22, 2014
http://aob.oxfordjournals.org/D
ownloaded from

552 Blumenthal et al.-Germination of Iris Seeds
to be more feasible than others: (a) the presence of germination inhibitors located in the endosperm (Randolph and Cox, 1943; Kartaschoff, 1958; Jorgensen, 1965; Arditti and Pray, 1969), (b) the mechanical resistance of a hard seed coat to the protrusion of the radicle (Crocker, 1906). It is also possible that both of these mechanisms may contribute to seed dormancy, a view that is supported by the work of Stolz (1968).
The possible role of germination inhibitors and a hard seed coat were therefore investigated in the seeds of the two iris species.
MATERIALS AND METHODS
Iris seeds were obtained from plants collected from the wild in Israel and grown in cultivation; l. atropurpurea (supplied by M. Deshe) originated from the Chefela and l. lorteti (supplied by 'Ampil Nurseries ') from Upper Galilee. All seeds were collected in May 1982 and stored in paper bags at room temperature (approx. 23 0q.
Isolation and germination of embryos
Seeds were imbibed for 24 h at room temperature aerated with an aquarium pump. They were then surface sterilized by immersion for 30 min in a solution containing 10 per cent H20 2, 10-4 M CuS04 and 100 mg 1-1 Na ascorbate, and then rinsed five times with a large volume of sterile distilled water.
The embryo was excised under sterile conditions by cutting the seed longitudinally and removing the embryo located in a groove in one half of the cut seed (Randolph and Cox, 1943; Randolph, 1945; Randolph and Randolph, 1955). The embryos were placed in petri dishes on solid Gamborg Bs mineral medium (Gamborg, Miller and Ojima, 1968) and incubated for 4 d at 25°C in the dark. Germination was defined as an increase in length of embryo by more than 5 mm. Two petri dishes were used in each treatment, with ten embryos per dish. The effects of seed extracts were tested after incorporation at various concentrations in the solid medium.
Treatments testing the effect of the seed coat on germination
Seeds were imbibed for 24 h, as described above, then the outer integument (including the ariI) was peeled off and the seeds dried to constant weight at 30°C. Measurements of imbibition were made both on intact seeds and without the outer integument. After surface sterilization, seeds without the outer integument with and without further manipulations were tested for germination on Gamborg Bs solid medium for 7 d at 25 o~ in the dark. A seed was considered germinated when the protruding rootlet was longer ~2~. .
The effect of oxygen was determined on intact seeds set to germinate on 40 ml solid Gamborg medium in 500 ml Erlenmeyer flasks. The flasks were covered with aluminium foil, and a stream of humidified oxygen, sterilized by passage through a 0·45 Jlm filter, was passed continuously through them.
Method for measuring the mechanical strength of the micropylar region
Seeds of the two irises were imbibed, the aril was removed and each seed was cut in half perpendicularly to its long axis. The embryo was removed and replaced by a cylindrical steel rod whose tip was 0·2 mm2 • The other end of the rod was fixed to the middle of a plate placed on one pan of a balance (Fig. 1). A plate with a hole in its middle was fixed above the half seed (see inset Fig. 1). The hole was observed through a stereoscopic microscope. On the other pan of the balance a beaker with just enough
at Frankfurt Univesity L
ibrary on August 22, 2014
http://aob.oxfordjournals.org/D
ownloaded from

Blumenthal et al.-Germination of Iris Seeds
-\-----7
1-----6
II II
3 ---+-+--+--~
FIG. I. Apparatus for measuring the inner integument resistance, by simulating the pressure' exerted by root protrusion. 1, burette; 2, loading pan; 3, pressure building pan; 4, stereoscopic microscope; 5, supporting device for seed. In inset: 6, cylindrical steel rod; 7, half seed after removal of embryo;
8, supporting plate with hole.
553
water for counterbalance was placed. Continuous addition of water from a burette, fixed above the beaker, allowed a controlled increase of the pressure exerted by the rod on the inner integument in the micropylar region. The stream of water was added very near the surface of the water in the beaker, on the wall of the beaker, to avoid the effect of the kinetic energy due to the fall of water. The pressure was increased at a rate of 0·85 atm S-l. The seed coat was observed continuously through the stereoscopic microscope. When the rod pierced the integument the increase in pressure was discontinued, the level of the water in the burette recorded and the pressure calculated.
Preparation of seed extracts
Extracts were prepared either from whole seeds or separately from the aril, the outer integument, or the endosperm together with inner integument after separation of the parts from imbibed seeds. Usually 12·5 g of whole dry seeds, or the relevant parts separated from them, were dried in a stream of air at 60°C to initial weight, ground to a powder and extracted with 50 ml of water for 30 min at 70°C with continuous stirring. The extract was filtered through two layers of gauze and centrifuged at 7500 g for 15 min. The cleared extracts were either concentrated under vacuum at 60°C or diluted with water as required.
at Frankfurt Univesity L
ibrary on August 22, 2014
http://aob.oxfordjournals.org/D
ownloaded from

554 Blumenthal et al.-Germination of Iris Seeds
After measurement of their osmotic potential (Knauer osmometer), the extracts were sterilized by passing through a 0·45 pm filter and incorporated into the Gamborg solid medium at known concentrations.
Fractionation of the extracts
The aqueous extracts prepared as described above were loaded either onto a CM-Sephadex-50 column (cation exchanger) or a QAE-Sephadex-50 column (anion exchanger) and eluted with 50 ml of water and the eluates were concentrated to 25 mI.
A further fractionation of the CM-Sephadex-50 eluate was achieved by concentrating it (in vacuo, 60°C) to 2 ml, loading onto a Whatman No.3 chromatography paper, and separating by descending chromatography with water as solvent. After drying, each chromatogram was cut in two to separate a high Rr and a low Rf half, and each was extracted twice with 50 ml water for 30 min. The extracts were pooled, filtered through Whatman No. 41 filter paper and brought to the desired volume. These extracts were tested for presence of germination inhibitors as described above.
Light microscopy. Median longitudinal sections and serial cross sections were hand cut from imbibed seeds. The sections were stained by ruthenium red (for hemicelluloses and pectic substances), Sudan IV (for suberin and other lipophilic substances), and phloroglucinol-HCI (for lignin: Jensen, 1962). Permanent sections were prepared from median segments of the seed 2 mm thick, fixed in FAA (a 9:5:5 mixture of 70 per cent ethanol + formaldehyde + acetic acid), dehydrated in a graded series of alcohol and cedar oil and embedded in paraffin. Microtome sections (12 pm) were cut and stained with: alcian green-safranin, Sudan IV, or ruthenium red.
Scanning electron microscopy. Imbibed seeds were cut longitudinally in two halves, dehydrated in a graded series of ethanol, and critical point dried with solvent-substituted liquid carbon dioxide. They were coated with a thin layer of gold and examined with a JEOL JSM 35 scanning electron microscope.
RESULTS
Germination inhibitors
The effect of various concentrations of seed extract of I. lorteti was tested using the growth of isolated embryos as a bioassay.
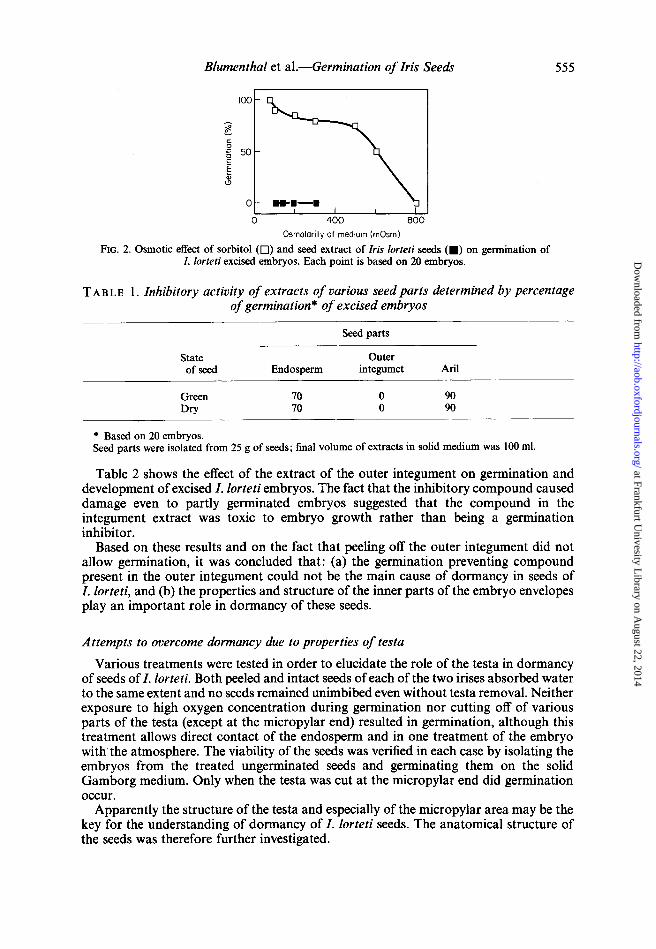
To evaluate the possibility that the inhibitory effect might be osmotic in nature (Lerner, Mayer and Evenari, 1959), the effect of the whole seed extract was compared to the effect of sorbitol at different osmolarities. Figure 2 shows that seed extracts do contain a germination preventing factor and that the effect is not osmotic; a concentration of 600 mOsm of sorbitol was necessary to reduce the germination of the embryos by 50 per cent, while a seed extract of 100 mOsm prevented germination completely.
The seed extract was filtered through ion exchange columns. The eluate from the CM-Sephadex inhibited embryo germination while that from the QAE-Sephadex had no inhibitory activity indicating that the inhibitory compound(s) is anionic. The eluate from the CM-Sephadex column was further purified by paper chromatography. After elution with water the inhibitory compound was found to have an R f of about 0·5.
Extraction of dissected seeds showed that the greater part of the inhibitory activity is located in the outer integument with slight inhibitory activity present also in the endosperm (Table I). Though most of the inhibitory substance was located in the outer integument, peeling this integument away did not allow germination to occur.
at Frankfurt Univesity L
ibrary on August 22, 2014
http://aob.oxfordjournals.org/D
ownloaded from
Blumenthal et al.-Germination of Iris Seeds
o .. -.-. o 400
Osmolarity of medium (mOsm)
FIG. 2. Osmotic effect of sorbitol (0) and seed extract of Iris lorteti seeds (.) on germination of I. lorteti excised embryos. Each point is based on 20 embryos.
555
TAB LE 1. Inhibitory activity of extracts of various seed parts determined by percentage of germination* of excised embryos
Seed parts
State Outer of seed Endosperm integumet Aril
Green 70 0 90 Dry 70 0 90
• Based on 20 embryos. Seed parts were isolated from 25 g of seeds; final volume of extracts in solid medium was 100 ml.
Table 2 shows the effect of the extract of the outer integument on germination and development of excised I. lorteti embryos. The fact that the inhibitory compound caused damage even to partly germinated embryos suggested that the compound in the integument extract was toxic to embryo growth rather than being a germination inhibitor.
Based on these results and on the fact that peeling off the outer integument did not allow germination, it was concluded that: (a) the germination preventing compound present in the outer integument could not be the main cause of dormancy in seeds of I. lorteti, and (b) the properties and structure of the inner parts of the embryo envelopes play an important role in dormancy of these seeds.
Attempts to overcome dormancy due to properties of testa
Various treatments were tested in order to elucidate the role of the testa in dormancy of seeds of I. lorteti. Both peeled and intact seeds of each of the two irises absorbed water to the same extent and no seeds remained unimbibed even without testa removal. Neither exposure to high oxygen concentration during germination nor cutting off of various parts of the testa (except at the micropylar end) resulted in germination, although this treatment allows direct contact of the endosperm and in one treatment of the embryo with the atmosphere. The viability of the seeds was verified in each case by isolating the embryos from the treated ungerminated seeds and germinating them on the solid Gamborg medium. Only when the testa was cut at the micropylar end did germination occur.
Apparently the structure of the testa and especially of the micropylar area may be the key for the understanding of dormancy of I. lorteti seeds. The anatomical structure of the seeds was therefore further investigated.
at Frankfurt Univesity L
ibrary on August 22, 2014
http://aob.oxfordjournals.org/D
ownloaded from

556 Blumenthal et aI.-Germination of Iris Seeds
TAB LE 2. Effect of exposure of excised embryos of Iris lorteti to medium containing a water soluble extract of the outer integument ofl. lorteti seeds for various periods
Periods of incubation (total = 19 d)
1st 2nd
9·5 (SM) 9·5(SM)
4 (EM) 15 (SM)
8 (EM) 11 (SM)
12 (EM) 7 (SM)
9·5 (EM) 9·5 (EM) 4 (SM) 15 (EM) 8 (SM) 11 (EM)
12 (SM) 7 (EM)
State of embryo after 19 d*
Normal seedling consisting of rootlet, additional root and one leaf. (Control)
Delayed germination and development. Complete recovery after release of embryo from effect of inhibitor
Ungerminated, partly brown embryo, tip of cotyledon greened after transfer to medium without extract
Ungerminated, partly brown embryo, tip of cotyledon greened after transfer to medium without extract
Brown embryo Brown germinated embryo Brown germinated embryo plus elongated rootlet Seedling including rootlet and leaf with arrested development
* Assessments based on 20 embryos in each treatment. SM, standard medium. EM, medium containing outer integument extract. Outer integuments from 12·5 g seeds, extracted with
50 ml water and diluted I : 1 with agar medium.
VB Em
Ch
A
En 01 II
FIG. 3. Schematic drawing of median longitudinal section of an iris seed from the Oncocyclus section. M, micropyle; A, aril; Em, embryo; VB, vascular bundle; Ch, chalaza; ST, suberized tissue; II, inner
integument; 01, outer integument; En, endosperm. 01, II and A together form the seed coat.
Structure of the iris seed
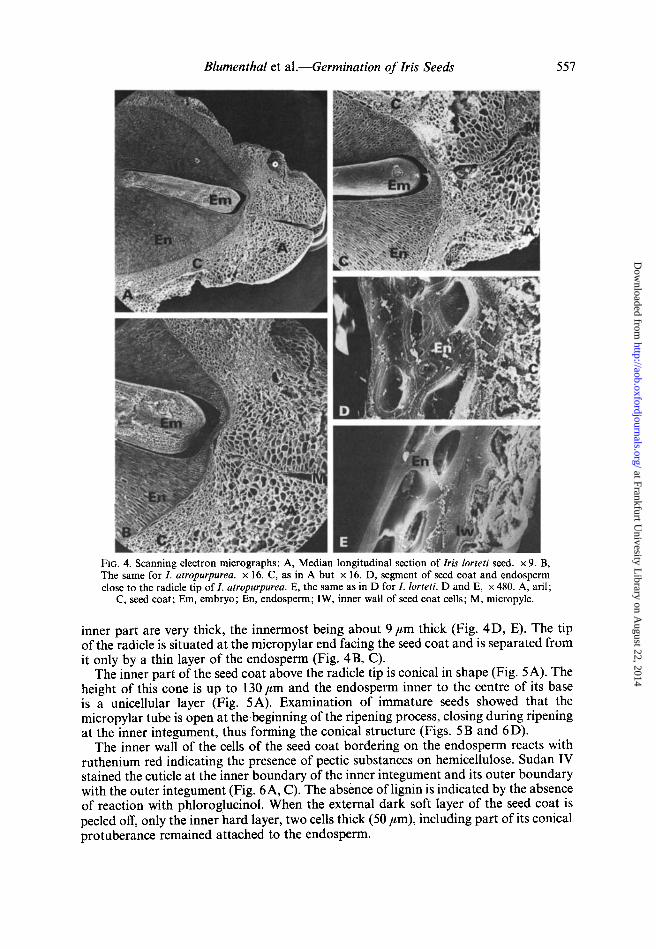
Examination ofthe seeds of the two types of irises showed a great similarity in general structure, comprising a seed coat consisting of two integuments, with an ariI around the micropyle, an endosperm that constitutes most of the volume of the seed, and an embryo embedded in the endosperm (Fig. 3).
The outer part of the seed coat consists of dark, soft, thin-walled cells, which can be easily peeled off (Fig. 4A, B, C). At the micropylar end the seed coat increases in thickness forming the arii. The micropyle passes through the middle of the aril but is closed at the inner part of the seed coat (Fig. 4A, B, C). The walls of the cells of this
at Frankfurt Univesity L
ibrary on August 22, 2014
http://aob.oxfordjournals.org/D
ownloaded from
Blumenthal et al.-Germination of Iris Seeds
FIG. 4. Scanning electron micrographs: A, Median longitudinal section of Iris lorteN seed. x 9. B, The same for l. atropurpurea. x 16. C, as in A but x 16. D, segment of seed coat and endosperm close to the radicle tip of I. atropurpurea. E, the same as in D for I. lorteti. D and E, x 480. A, aril;
C, seed coat; Em, embryo; En, endosperm; IW, inner wall of seed coat cells; M, micropyle.
557
inner part are very thick, the innermost being about 9/lm thick (Fig. 4 D, E). The tip of the radicle is situated at the micropylar end facing the seed coat and is separated from it only by a thin layer of the endosperm (Fig. 4B, C).
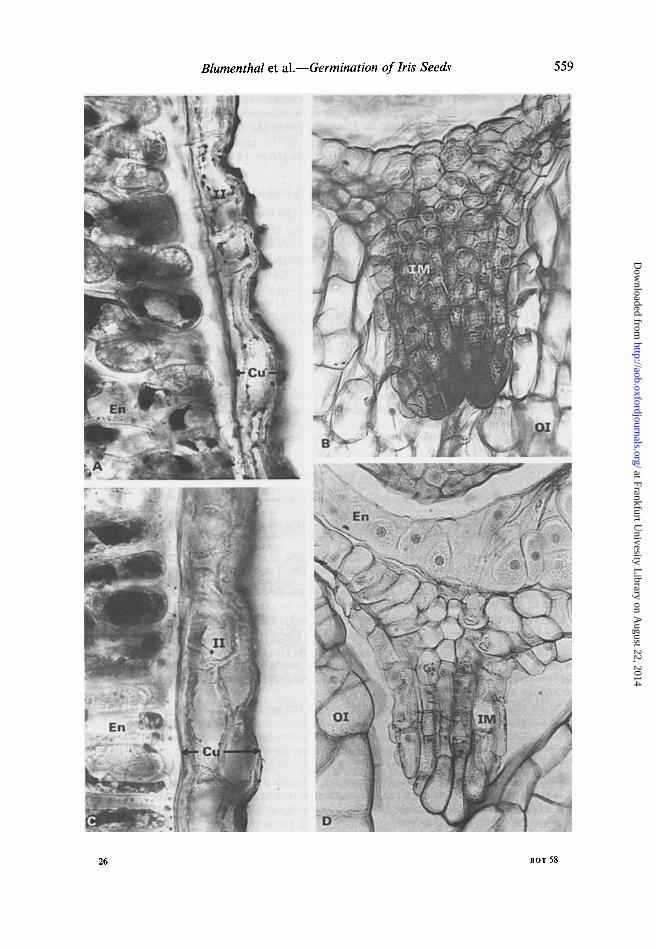
The inner part of the seed coat above the radicle tip is conical in shape (Fig. 5A). The height of this cone is up to 130/lm and the endosperm inner to the centre of its base is a unicellular layer (Fig. 5A). Examination of immature seeds showed that the micropylar tube is open at the beginning of the ripening process, closing during ripening at the inner integument, thus forming the conical structure (Figs. 5B and 6D).
The inner wall of the cells of the seed coat bordering on the endosperm reacts with ruthenium red indicating the presence of pectic substances on hemicellulose. Sudan IV stained the cuticle at the inner boundary of the inner integument and its outer boundary with the outer integument (Fig. 6A, C). The absence oflignin is indicated by the absence of reaction with phloroglucinol. When the external dark soft layer of the seed coat is peeled off, only the inner hard layer, two cells thick (50 /lm), including part of its conical protuberance remained attached to the endosperm.
at Frankfurt Univesity L
ibrary on August 22, 2014
http://aob.oxfordjournals.org/D
ownloaded from

558 Blumenthal et al.-Germination of Iris Seeds
FIG. 5. Photomicrographs of segments of Iris atropurpurea seeds. A, Radicle tip close to inner micropylar end. x 300. B, Inner micropylar end of unripe seed. x 330. C, Seed coat; CS, conical protuberance; En, endosperm; 1M, flared margins of inner integument of micropyle; M, micropyle;
R, radicle.
A vascular bundle passes through the aril inside the external integument, along the seed towards the chalazal region. Above the chalazal region suberized tissue can be identified (Fig. 3).
The only anatomical difference between the seeds of the two Iris species was found in conical protuberance of the inner integument at the micropylar side. In the mature
FIG. 6. Photomicrographs of sections of Iris seeds. l. lorteti: A, Part of the inner integument and endosperm stained with Sudan IV. x 330. B, inner micropylar end of ripe seed. x 330. I. atropurpurea: C, Part of inner integument and endosperm stained with Sudan IV. x 330. D, inner micropylar end of ripe seed. x 330. Note the relatively small cubically shaped cells in the conical protuberance of the inner integument in l. lorteti seed (B) as compared to long shaped cells in the conical protuberance of the inner integument of l. atropurpurea seed (D). Cu, cuticle; En, endosperm; 1M, flared margins of inner integument fused together forming the conical protuberance; II, inner
integument; 01, outer integument.
at Frankfurt Univesity L
ibrary on August 22, 2014
http://aob.oxfordjournals.org/D
ownloaded from
Blumenthal et al.-Germination of Iris Seeds 559
26 BOT 58
at Frankfurt Univesity L
ibrary on August 22, 2014
http://aob.oxfordjournals.org/D
ownloaded from

560 Blumenthal et al.-Germination of Iris Seeds
seeds of I. lorteti this region consists of relatively small cubically shaped cells (Fig. 6 B). In I. atropurpurea in this region the cells are approximately twice as long as those in I. lorteti (Fig. 6D). Therefore, while in I. atropurpurea the tissue facing the edge of the radicle is only four to five cells long (eight to 10 cross walls), in I. lorteti the corresponding tissue consists of eight to nine cells (16 to 18 cross walls) (Fig. 6B, D).
Measurement of the mechanical strength of the micropylar region
The anatomical difference between the seeds of the Iris species suggests the possibility that the protruding radicle in the germinating seed of I. lorteti may encounter a much greater resistance than the radicle of the seed of I. atropurpurea. The hypothesis was tested experimentally by measurement of the force required to rupture the seed coat at the micropylar region. The technique is described in detail in Materials and Methods and the results presented in Table 3. The pressure which must be overcome in Pancratium maritimum in order to pierce the inner integument of the seed (the outer integument was removed) was estimated in the same way for comparison (see Table 3). This shows clearly that the seed coat of l. lorteti requires approximately twice as much pressure as that of I. atropurpurea before it ruptures. In both Iris species much more pressure is required to pierce the seed coat than in Pancratium.
TABLE 3. Pressure required to pierce the inner integument at the micropylar end of Iris lorteti, I. atropurpurea and Pancratium maritimum seeds
Species
I.lorteti I. atropurpurea P. maritimum
Mean pressure (atm)*
135±12 77±9 1O±3
* Based on nine replicate detenninations. Values shown ±SE.
DISCUSSION
The resistance of the seed coat of I.lortetiis amongst the highest recorded in the literature (Table 4). The hardness of the seed coat appears to be the main cause for seed dormancy in irises, and therefore, no germination of Iris seeds can occur unless the structure of the seed coat in the micropylar region is loosened. These results also explain the difference in the degree of dormancy of the two iris species. The results also explain why cutting the testa only at the micropylar end enables the seeds to germinate. This method was very successful and is now being used to germinate Iris seeds in the field for commercial use and as a test assay in the laboratory.
Earlier attempts to overcome the dormancy of Iris seeds by cutting off the integument at the micropylar end have been reported in the literature. Crocker (1906) and Mitchell (1949) succeeded in obtaining 97 per cent germination in an unspecified iris but they did not investigate the structure of the integument or the possible reasons for dormancy. Werckmeister (1962), on the other hand, failed to obtain any germination in Iris seeds by cutting the inner integument; there are not enough data in Werckmeister's paper to explain the discrepancy between his results and the results of Crocker, Mitchell and ours.
Arditti and Pray (1969) claimed that the main cause of dormancy in irises is due to the presence of dormancy inducing compounds in the endosperm. We have also found a slight germination inhibition by extracts prepared from endosperm (plus inner
at Frankfurt Univesity L
ibrary on August 22, 2014
http://aob.oxfordjournals.org/D
ownloaded from

Blumenthal et al.-Germination of Iris Seeds
TABLE 4. Pressure necessary for root protrusion of the germinating seed
Species
Juglans nigra L. Juglans arenaria L. Carya ovata Xanthium pennsylvanicum Capsicum annuum
Mean pressure (atm)
30 50 25
1·7 66
Reference
Crocker, Thornton and Schroeder (1946) Crocker et al. (1946) Crocker et al. (1946) Esahi and Leopold (1968) Watkin and Daniel (1983)
Data compiled from Ib in-2 and transformed into atmospheres.
561
integument) but it is evident that neither this inhibitor nor the more active germinationpreventing substance(s) present in the outer. integument, can be the main cause of dormancy of these seeds. Some interaction between the effect of this inhibitor and the hardness of the testa cannot, at present, be ruled out completely, but the mechanical resistance of the testa at the micropylar region seems to be the main factor preventing germination. On the basis of this mechanism the differences in germination behaviour of the two Iris species can easily be explained.
ACKNOWLEDGEMENTS
Our thanks are due to Mr Michael Deshe from Sdema, Israel and to ' Ampil Nurseries', Ramat Zvi, Israel for supplying the Iris seeds and to Dr Yacov Aharonowicz from the Institute of Microbiology, Hadassah Medical School, Jerusalem for suggesting the method of disinfecting the seeds.
LITERATURE CITED
ARDITTI, J. and PRAY, T. R., 1966. A review of seed germination data from the genus Iris by Rodionenko (1961). Aril Society Yearbook 15-16.
-- -- 1969. Dormancy factors in Iris seeds. American Journal of Botany 56,254--9. AVISHAI, M., 1977. Species relationship and cytogenetic affinities in section Oncocyclus of the genus Iris. Ph.D.
Thesis, Hebrew University, Jerusalem. CROCKER, W., 1906. Role of seed coats in delayed germination. Botanical Gazette 42, 265-91. -- THORNTON, N. C. and SCHROEDER, E. M., 1946. Internal pressure necessary to break shells of nuts and
the role of the shells in delayed germination. Contributions of the Boyce Thompson Institute 14, 173-201. EsAHI, J. and LEOPOLD, A. C., 1968. Physical forces in dormancy and germination of Xanthium seeds. Plant
Physiology 43,871-6. GAMBORG, O. L., MILLER, R. A. and OJIMA, K., 1968. Nutrient requirement of suspension cultures of soybean
root cells. Experimental Cell Research SO, 151-8. JENSEN, W. A., 1962. Botanical Histochemistry. Freeman, San Francisco. JORGENSEN, C. J. C., 1965. Inhibitory effects of Iris seed extracts on germination of indicator plants. Bulletin
of the American Iris Society 179, 27-33. KARTASCHOFF, R., 1958. Die Keimuerzogenung der Samen von Iris pseudacorus L. Berichte der Schweizerischen
Botanischen Gesellschaft 68, 145-82. KIDD, K. K., 1964. A review offactors in Iris seed germination. Bulletin of the American Iris Society 181, 14--22. LERNER, H. R., MAYER, A. M. and EVENARI, M., 1959. The nature of the germination inhibitors present in
dispersal units of Zygophyllum dumosum and Trigonella arabica. Physiologia Plantarum 12, 245-50. MITCHELL, S., 1949. Irisfor Every Garden. M. Barrows, New York. RANDOLPH, C. F., 1945. Embryo seed culture. Bulletin of the American Iris Society 97, 33-45. -- and Cox, L. G., 1943. Factors influencing the germination of Iris seed and the relation of inhibiting
substances to embryo dormancy. Proceedings of the American Society of Horticultural Science 43,284--300. --and RANDOLPH, R., 1955. Embryo culture of Iris seeds. Bulletin of the American Iris Society 139, 7-17. STOLZ, C. P., 1968. Iris seed dormancy. Physiologia Plantarum 21,1328-31. WATKINS, J. T. and DANIEL, J. C., 1983. Mechanical resistance of the seed coat and endosperm during
germination of Capsicum annuum at low temperature. Plant Physiology 72, 146--50. WERCKMEISTER, P., 1962. Dber embryonales Wachstum auserhalb des Samens, Keimung und Keimverzug.
Untersuchungen an Bastardsamen und Embryonen der Gattung Iris L. Biologischer Zentralblatt 6,667-90.
26·2
at Frankfurt Univesity L
ibrary on August 22, 2014
http://aob.oxfordjournals.org/D
ownloaded from