fundamentals of geobiology (knoll/fundamentals of geobiology) || supplemental images
TRANSCRIPT
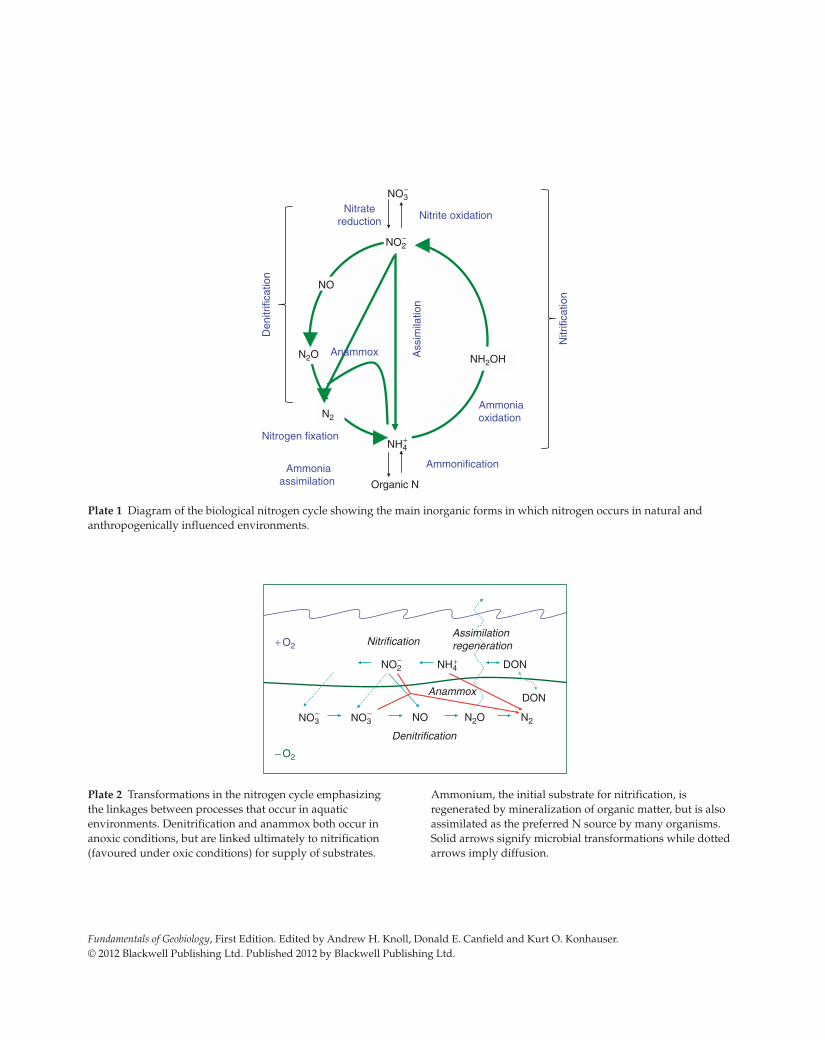
Fundamentals of Geobiology, First Edition. Edited by Andrew H. Knoll, Donald E. Canfield and Kurt O. Konhauser.
© 2012 Blackwell Publishing Ltd. Published 2012 by Blackwell Publishing Ltd.
N2
Nitr
ifica
tion
Den
itrifi
catio
n
Organic N
Ass
imila
tion
N2O
NO
NH2OH
AmmonificationAmmoniaassimilation
Nitrogen fixation
Nitrite oxidationNitrate
reduction
Ammoniaoxidation
Anammox
NO3−
NO2−
NH4+
Plate 1 Diagram of the biological nitrogen cycle showing the main inorganic forms in which nitrogen occurs in natural and
anthropogenically influenced environments.
NO N2O N2
DON
+ O2
− O2
Nitrification
Denitrification
Anammox
Assimilationregeneration
DON
NO2−
NO3− NO3
−
NH4+
Plate 2 Transformations in the nitrogen cycle emphasizing
the linkages between processes that occur in aquatic
environments. Denitrification and anammox both occur in
anoxic conditions, but are linked ultimately to nitrification
(favoured under oxic conditions) for supply of substrates.
Ammonium, the initial substrate for nitrification, is
regenerated by mineralization of organic matter, but is also
assimilated as the preferred N source by many organisms.
Solid arrows signify microbial transformations while dotted
arrows imply diffusion.
Knoll_bins.indd 1Knoll_bins.indd 1 2/16/2012 2:11:57 AM2/16/2012 2:11:57 AM
–40
–20
0
20
40
60
12
4
8
0
0
0.5
1.0
1.5
0.1
1.0
10
0
2
4
6
???
Δ33S
δ33S
f -ratio
Sulfate (mM)
Sulfur reservoir
x 1020 mol
Relative BIFdeposition
Deep water
O2
H2S
Fe2+
???
???
0 500 1000 1500 2000 2500 3000 3500 4000
Time (Ma)
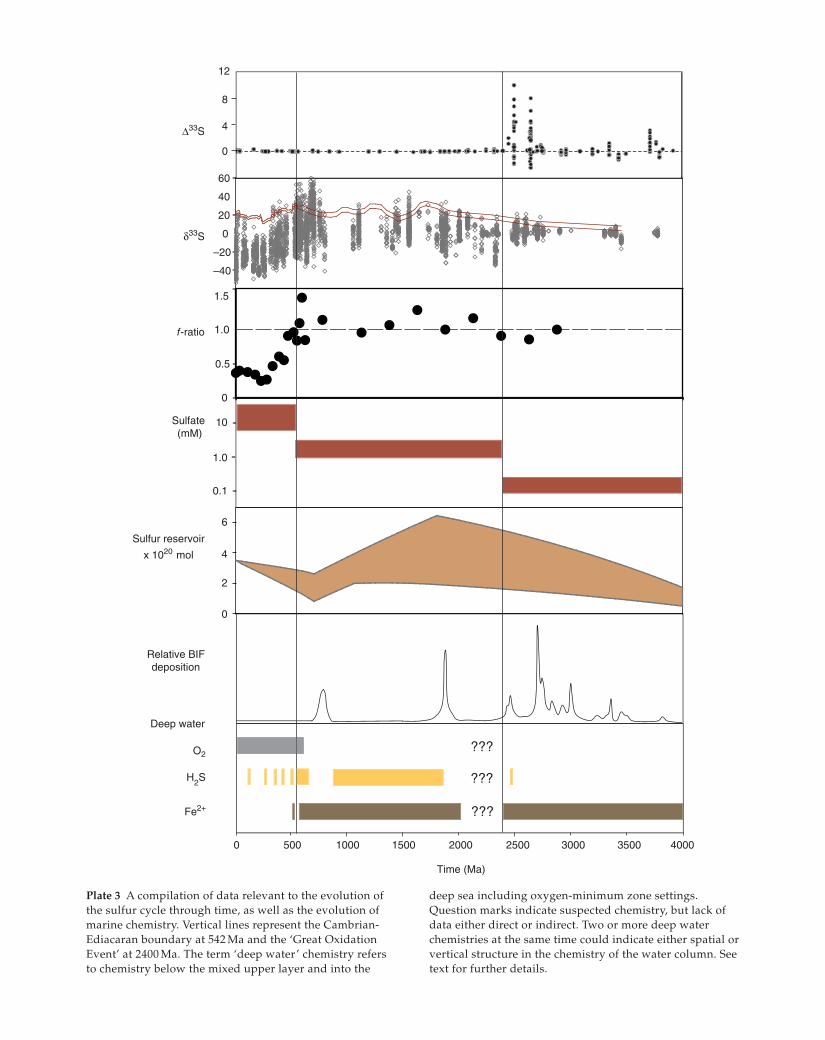
Plate 3 A compilation of data relevant to the evolution of
the sulfur cycle through time, as well as the evolution of
marine chemistry. Vertical lines represent the Cambrian-
Ediacaran boundary at 542 Ma and the ‘Great Oxidation
Event’ at 2400 Ma. The term ‘deep water’ chemistry refers
to chemistry below the mixed upper layer and into the
deep sea including oxygen-minimum zone settings.
Question marks indicate suspected chemistry, but lack of
data either direct or indirect. Two or more deep water
chemistries at the same time could indicate either spatial or
vertical structure in the chemistry of the water column. See
text for further details.
Knoll_bins.indd 2Knoll_bins.indd 2 2/16/2012 2:11:58 AM2/16/2012 2:11:58 AM
(a)HCO3
–
Ca2+ Ca2+
Ca2+
Ca2+Ca2+
S-layer
Ca2+
Anionicligands
Calcitenucleation
OH–
Cyanobacterium
CO32–
(b)+ +Fe2+ 2Fe(OH)3 Fe3O4+ 4H2O2OH–
Fe(lll)-reducing bacterium
Fe2+
2e–
Fe2+
Fe(OH)3
Magnetitenucleation
EPS
CH3COO–
Fe(OH)3
OH–
H2O + CO32– HCO3
– OH–+
HCO3– +H2O CH2O + O2 + OH–
+ +2H+ 2Fe(OH)3--> + 4OH–+ 2H2O2e– 2Fe2+
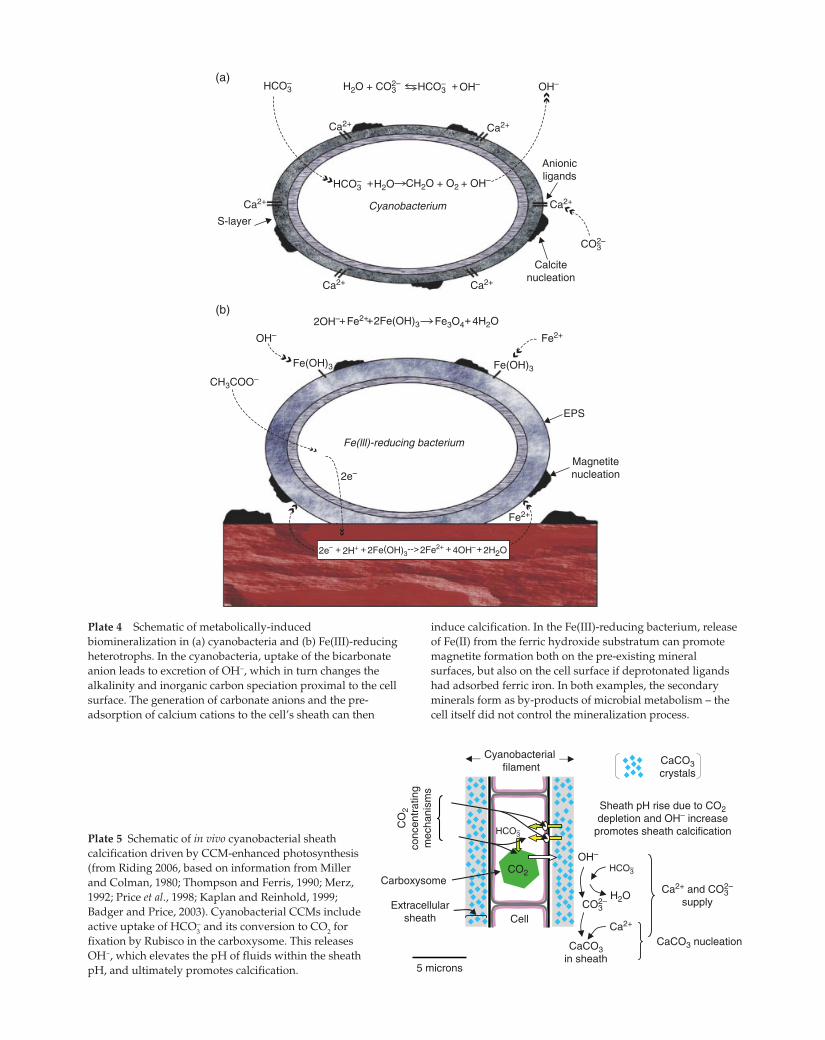
Plate 4 Schematic of metabolically-induced
biomineralization in (a) cyanobacteria and (b) Fe(III)-reducing
heterotrophs. In the cyanobacteria, uptake of the bicarbonate
anion leads to excretion of OH−, which in turn changes the
alkalinity and inorganic carbon speciation proximal to the cell
surface. The generation of carbonate anions and the pre-
adsorption of calcium cations to the cell’s sheath can then
induce calcification. In the Fe(III)-reducing bacterium, release
of Fe(II) from the ferric hydroxide substratum can promote
magnetite formation both on the pre-existing mineral
surfaces, but also on the cell surface if deprotonated ligands
had adsorbed ferric iron. In both examples, the secondary
minerals form as by-products of microbial metabolism – the
cell itself did not control the mineralization process.
HCO3–
HCO3–
Carboxysome
Extracellularsheath
Sheath pH rise due to CO2depletion and OH– increase
promotes sheath calcification
OH–
CO32–
Ca2+
H2O
CaCO3 in sheath
CO
2co
ncen
trat
ing
mec
hani
sms
Ca2+ and CO32–
supply
CaCO3 nucleation
CO2
Cyanobacterial filament
Cell
CaCO3crystals
5 microns
Plate 5 Schematic of in vivo cyanobacterial sheath
calcification driven by CCM-enhanced photosynthesis
(from Riding 2006, based on information from Miller
and Colman, 1980; Thompson and Ferris, 1990; Merz,
1992; Price et al., 1998; Kaplan and Reinhold, 1999;
Badger and Price, 2003). Cyanobacterial CCMs include
active uptake of HCO3− and its conversion to CO
2 for
fixation by Rubisco in the carboxysome. This releases
OH−, which elevates the pH of fluids within the sheath
pH, and ultimately promotes calcification.
Knoll_bins.indd 3Knoll_bins.indd 3 2/16/2012 2:11:58 AM2/16/2012 2:11:58 AM

Infe
rred
C
O2
tren
d (a
) 20
0 40
20 0
40
500
1000
0
Mill
ions
of y
ears
ago
a c
She
ath
calc
ifica
tion
in
duce
d de
spite
el
evat
ed C
O2
550–
150
Ma,
cal
cifie
d sh
eath
abu
ndan
ce b
road
ly
corr
espo
nds
with
mar
ine
carb
onat
e sa
tura
tion
stat
e
~35
0 M
a,
shea
th
calc
ifica
tion
incr
ease
as
CO
2 de
clin
ed
Saturation state Ωcalcite
Sheath calcified cyanobacteria
abundance
Cen
ozoi
c,
mar
ine
shea
th
calc
ifica
tion
scar
ce
to a
bsen
t
A
nom
alou
s el
evat
ed
satu
ratio
n st
ate
Mar
ine
wh
iting
s an
d sh
eath
ca
lcifi
catio
n re
duce
d by
lo
wer
car
bona
te
satu
ratio
n st
ate
Firs
t sh
eath
ca
lcifi
ed
cyan
obac
teria
Car
bona
te
mud
do
min
ated
sh
elve
s
Infe
rred
CC
M
indu
ctio
n as
C
O2
decl
ined
be
low
~10
PA
L
Infe
rred
in
cept
ion
of
biog
enic
‘w
hitin
gs’
She
ath
calc
ified
cy
anob
acte
ria
wid
espr
ead
GE
OC
AR
B II
I m
odel
led
CO
2 tr
end
Cal
cifie
d sh
eath
s sc
arce
b
a
Pos
sibl
y
due
to lo
w
tem
pera
ture
an
d lo
w
satu
ratio
n st
ate
10
PA
L C
O2
Infe
rred
thre
sho
ld
belo
w w
hich
C
CM
s in
duce
d
1500
Gla
ciat
ions
2 5
4 1
3 6
7 8
CO2 ratio to present-day pre-industrial atmospheric level (PAL)
Pla
te 6
C
on
ject
ura
l h
isto
ry o
f cy
an
ob
act
eria
l m
ari
ne
shea
th c
alc
ific
ati
on
an
d p
ico
pla
nk
tic
‘wh
itin
g’
pre
cip
itati
on
. T
he
Pro
tero
zo
ic i
nfe
rred
CO
2 t
ren
d i
s b
ase
d o
n (
a)
Sh
eld
on
(2006),
(b
) K
ah
an
d R
idin
g (
2007),
(c)
Hy
de
et a
l. (2
000)
an
d (
d)
Rid
gw
ell
et a
l. (2
003);
th
e co
nti
nu
ou
s tr
end
lin
e is
fro
m B
ern
er &
Ko
thav
ala
(2001, fi
g. 13);
th
e N
eop
rote
rozo
ic
gla
ciati
on
s is
fro
m W
alt
er e
t al.
(2000);
th
e o
ccu
rren
ces
of
mari
ne
shea
th c
alc
ifie
d c
yan
ob
act
eria
is
fro
m A
rp e
t al.
(2001, fi
g. 3d
); t
he
calc
ula
ted
carb
on
ate
satu
rati
on
sta
tes
are
fro
m R
idin
g a
nd
Lia
ng
(2005b
, fi
g. 5);
an
d t
he
thre
sho
ld b
elo
w w
hic
h C
CM
s are
in
du
ced
is
base
d o
n B
ad
ger
et a
l. (2
002).
Sev
eral
key
dev
elo
pm
ents
can
be
infe
rred
fro
m t
he
fig
ure
. (1
) P
ho
tosy
nth
etic
‘w
hit
ing
s’, as
refl
ecte
d b
y w
ides
pre
ad
carb
on
ate
mu
d s
edim
enta
tio
n, m
ay
hav
e b
een
tri
gg
ered
as
CO
2 r
edu
ced
pH
bu
ffer
ing
(se
e A
rp e
t al.,
2001;
Rid
ing
, 2006).
(2)
A f
urt
her
dec
lin
e b
elo
w ~
10 P
AL
CO
2 i
nd
uce
d C
CM
dev
elo
pm
ent
an
d s
hea
th c
alc
ific
ati
on
at
~1200 M
a (
Kah
an
d R
idin
g, 2007).
(3)
Calc
ifie
d s
hea
ths
wer
e
wid
esp
read
in
th
e ea
rly
Neo
pro
tero
zo
ic (
see
refe
ren
ces
in K
no
ll a
nd
Sem
ikh
ato
v 1
998),
bu
t (4
) b
ecam
e sc
arc
e d
uri
ng
‘S
no
wb
all
’ g
laci
ati
on
s, p
oss
ibly
du
e to
red
uct
ion
in
CC
M
dev
elo
pm
ent
as
low
tem
per
atu
res
fav
ou
red
dif
fusi
ve
entr
y o
f C
O2 i
nto
cel
ls, an
d d
ue
to l
ow
er s
eaw
ate
r sa
tura
tio
n s
tate
ref
lect
ing
red
uct
ion
in
bo
th t
emp
eratu
re a
nd
pC
O2. (5
)
Sh
eath
calc
ific
ati
on
was
com
mo
n i
n m
ari
ne
env
iro
nm
ents
du
rin
g t
he
earl
y-m
id P
ala
eozo
ic d
esp
ite
elev
ate
d C
O2, su
gg
esti
ng
th
at
on
ce C
CM
s h
ad
dev
elo
ped
th
ey w
ere
read
ily
ind
uce
d w
her
e ca
rbo
n l
imit
ati
on
dev
elo
ped
, su
ch a
s m
icro
bia
l m
ats
. T
hro
ug
ho
ut
mu
ch o
f th
e P
ala
eozo
ic a
nd
earl
y M
eso
zo
ic, ca
lcif
ied
sh
eath
ab
un
dan
ce a
pp
ears
to
vary
wit
h
carb
on
ate
satu
rati
on
sta
te. (6
) A
s C
O2 d
ecli
ned
in
th
e L
ate
Dev
on
ian
-Earl
y M
issi
ssip
pia
n, ca
lcif
ied
sh
eath
ab
un
dan
ce t
emp
ora
rily
in
crea
sed
, p
oss
ibly
ref
lect
ing
en
han
ced
CC
M i
nd
uct
ion
, b
ut
then
dec
lin
ed a
s th
e sa
tura
tio
n s
tate
dro
pp
ed i
n t
he
Mis
siss
ipp
ian
-Pen
nsy
lvan
ian
(R
idin
g, 2009).
(7)
Des
pit
e a h
igh
calc
ula
ted
satu
rati
on
sta
te i
n t
he
Late
Cre
tace
ou
s-P
ala
eog
ene,
pla
nk
tic
calc
ifie
rs p
rob
ab
ly r
edu
ced
th
e act
ual
satu
rati
on
sta
te s
uff
icie
ntl
y t
o i
nh
ibit
cy
an
ob
act
eria
l ca
lcif
icati
on
. T
hu
s, c
alc
ifie
d s
hea
ths
wer
e sc
arc
e in
mari
ne
env
iro
nm
ents
. (8
) S
ince
th
e E
oce
ne,
lo
w c
arb
on
ate
satu
rati
on
– d
ue
to l
ow
lev
els
of
bo
th C
a i
on
s an
d p
CO
2– i
s re
flec
ted
in
ex
trem
e sc
arc
ity
in
sh
eath
calc
ific
ati
on
in
mari
ne
env
iro
nm
ents
.
Knoll_bins.indd 4Knoll_bins.indd 4 2/16/2012 2:12:01 AM2/16/2012 2:12:01 AM
(a) (d)
(b) (e)
(c) (f)
<121>
<110>
<121>
<110>
<121>
<110>
<121>
<110>
<121>
<110>
<121>
<110>
Lattice oxygen Fetet1Feoct2
Capping oxygen
OA
Fetet1
Feoct1
OBFetet2Feoct2Fetet1OC
Lattice oxygen
Surface A
1.2 ± 0.1 Å
3.3 ± 0.1 Å
0.4 ± 0.1 Å
Surface A’
Surface BSurface Aα β
Fetet1/Fetet2
Feoct2 Capping oxygen
4.54.03.53.02.52.0
Ver
tical
dis
tanc
e (Å
)
1.51.00.50.0
Feoct1
[111][110]
A
A’
B
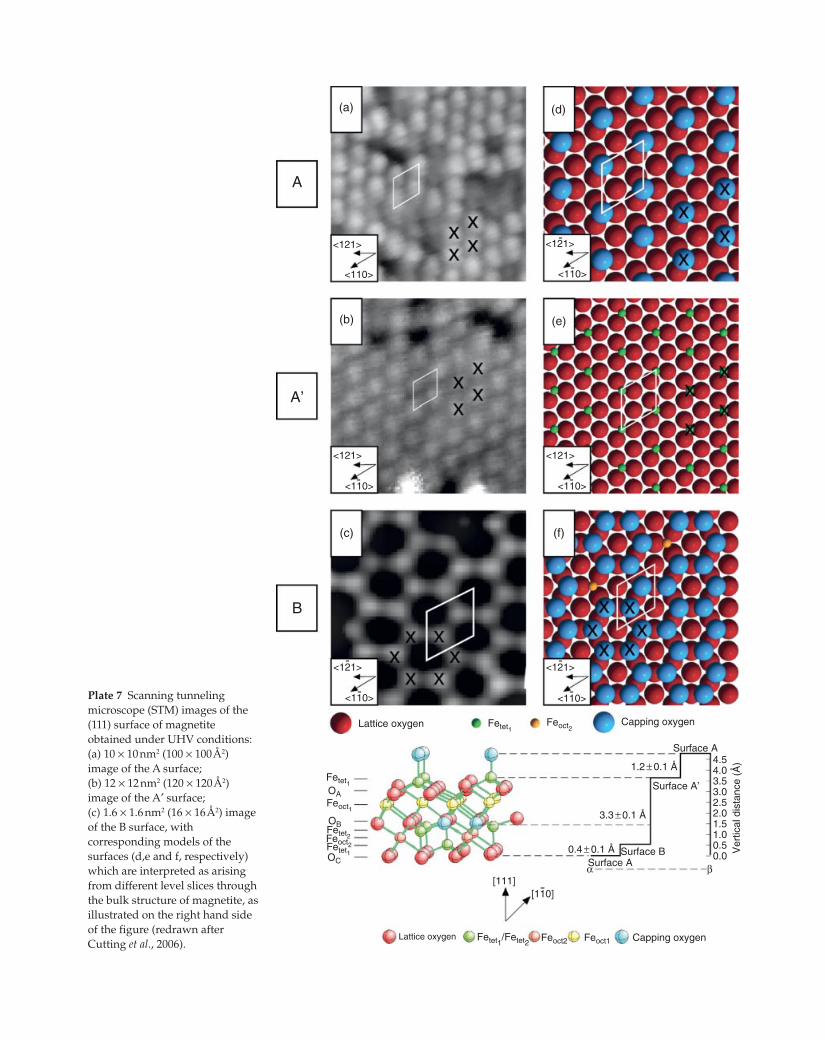
Plate 7 Scanning tunneling
microscope (STM) images of the
(111) surface of magnetite
obtained under UHV conditions:
(a) 10 × 10 nm2 (100 × 100 Å2)
image of the A surface;
(b) 12 × 12 nm2 (120 × 120 Å2)
image of the A′ surface;
(c) 1.6 × 1.6 nm2 (16 × 16 Å2) image
of the B surface, with
corresponding models of the
surfaces (d,e and f, respectively)
which are interpreted as arising
from different level slices through
the bulk structure of magnetite, as
illustrated on the right hand side
of the figure (redrawn after
Cutting et al., 2006).
Knoll_bins.indd 5Knoll_bins.indd 5 2/16/2012 2:12:01 AM2/16/2012 2:12:01 AM
4.1e + 003
12C
3.7e + 003
3.2e + 003
2.8e + 003
2.3e + 003
1.8e + 003
1.4e + 003
9.3e + 002
4.7e + 002
12.
12C14N1.0e + 003
10 um
(c)(a)
(b)
(d)
9.1e + 002
8.0e + 002
6.8e + 002
5.7e + 002
4.5e + 002
3.4e + 002
2.3e + 002
1.1e + 002
0.00
32S
4.4e + 002
3.9e + 002
3.4e + 002
2.9e + 002
2.4e + 002
1.9e + 002
1.5e + 002
97.
49.
0.00
(e)
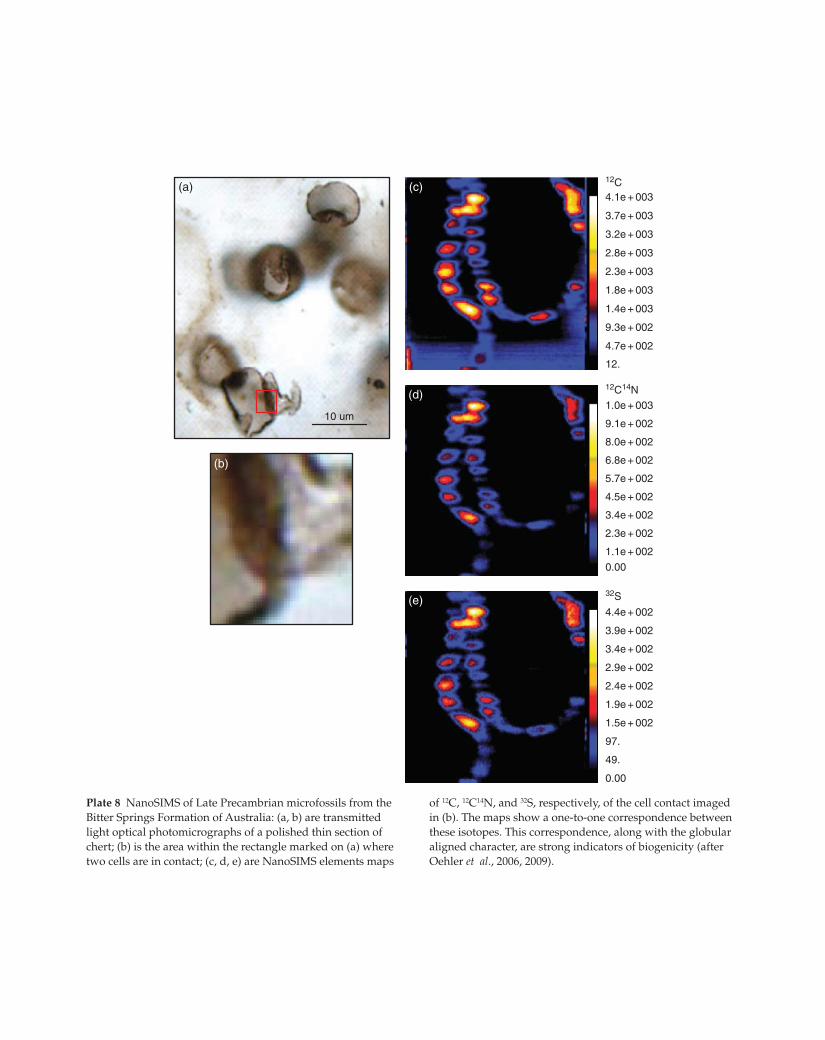
Plate 8 NanoSIMS of Late Precambrian microfossils from the
Bitter Springs Formation of Australia: (a, b) are transmitted
light optical photomicrographs of a polished thin section of
chert; (b) is the area within the rectangle marked on (a) where
two cells are in contact; (c, d, e) are NanoSIMS elements maps
of 12C, 12C14N, and 32S, respectively, of the cell contact imaged
in (b). The maps show a one-to-one correspondence between
these isotopes. This correspondence, along with the globular
aligned character, are strong indicators of biogenicity (after
Oehler et al., 2006, 2009).
Knoll_bins.indd 6Knoll_bins.indd 6 2/16/2012 2:12:06 AM2/16/2012 2:12:06 AM

Bulksolution
Carapace
Carapacesupportstructure
Basalbiofilm
Mineralsurface
Uncolonisedmineral surface
Intra-biofilmfluid flow
321.4μm
25.3μm
31.5 μm
321.4 μm
Primary colonising bacteria
Intra-biofilm bacterial colonies
Carapace bacterial colonies
Carapace polysaccharides/EPS
Basal polysaccharides/EPS
Fluid flow
Plate 9 Confocal scanning laser microscope (CSLM)
image of a biofilm grown between two quartz glass
plates by introducing a nutrient solution and inoculating
with Pseudomonas aeruginosa. Below the image is a
schematic diagram showing the various components of
the biofilm (after Brydie et al., 2004, 2009).
Knoll_bins.indd 7Knoll_bins.indd 7 2/16/2012 2:12:07 AM2/16/2012 2:12:07 AM
Fe-O4 weeks
2 weeks
1 week
Fe 2p3/2
Fe 2p1/2
× 102
5
10
15
20
25
30
35
CP
S
740 735 730 725 720 715 710 705 700Binding energy (eV)
(b)
CP
S
Fe 2p3/2
1 week
2 weeks
4 weeks
Fe 2p1/2
× 102
10
15
20
25
740 735 730 725 720 715 710 705 700Binding energy (eV)
(c) (d)
80×101
CP
S
706050403020
Fe3p
58 56 54
As 3d
2 weeks
1 weeks
4 weeks
As(V)-OAs(lll)-O
52 50 48Binding energy (eV)
46 44 42 40 38
10
(a)
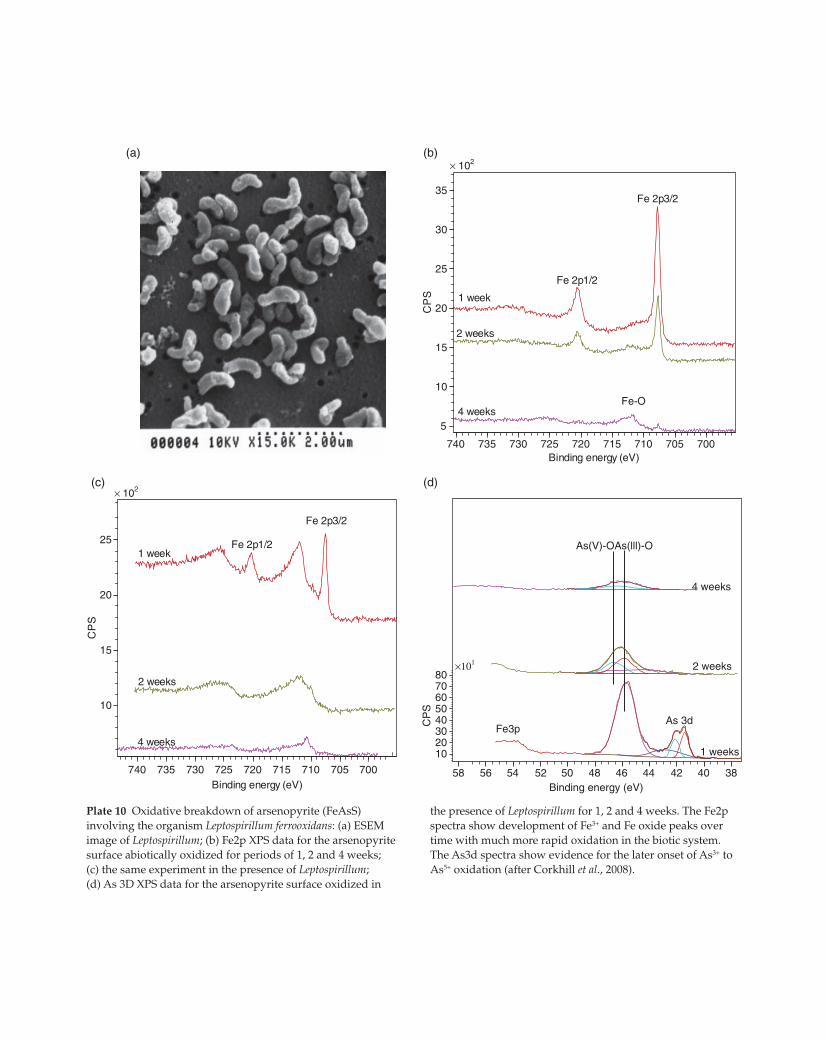
Plate 10 Oxidative breakdown of arsenopyrite (FeAsS)
involving the organism Leptospirillum ferrooxidans: (a) ESEM
image of Leptospirillum; (b) Fe2p XPS data for the arsenopyrite
surface abiotically oxidized for periods of 1, 2 and 4 weeks;
(c) the same experiment in the presence of Leptospirillum;
(d) As 3D XPS data for the arsenopyrite surface oxidized in
the presence of Leptospirillum for 1, 2 and 4 weeks. The Fe2p
spectra show development of Fe3+ and Fe oxide peaks over
time with much more rapid oxidation in the biotic system.
The As3d spectra show evidence for the later onset of As3+ to
As5+ oxidation (after Corkhill et al., 2008).
Knoll_bins.indd 8Knoll_bins.indd 8 2/16/2012 2:12:12 AM2/16/2012 2:12:12 AM
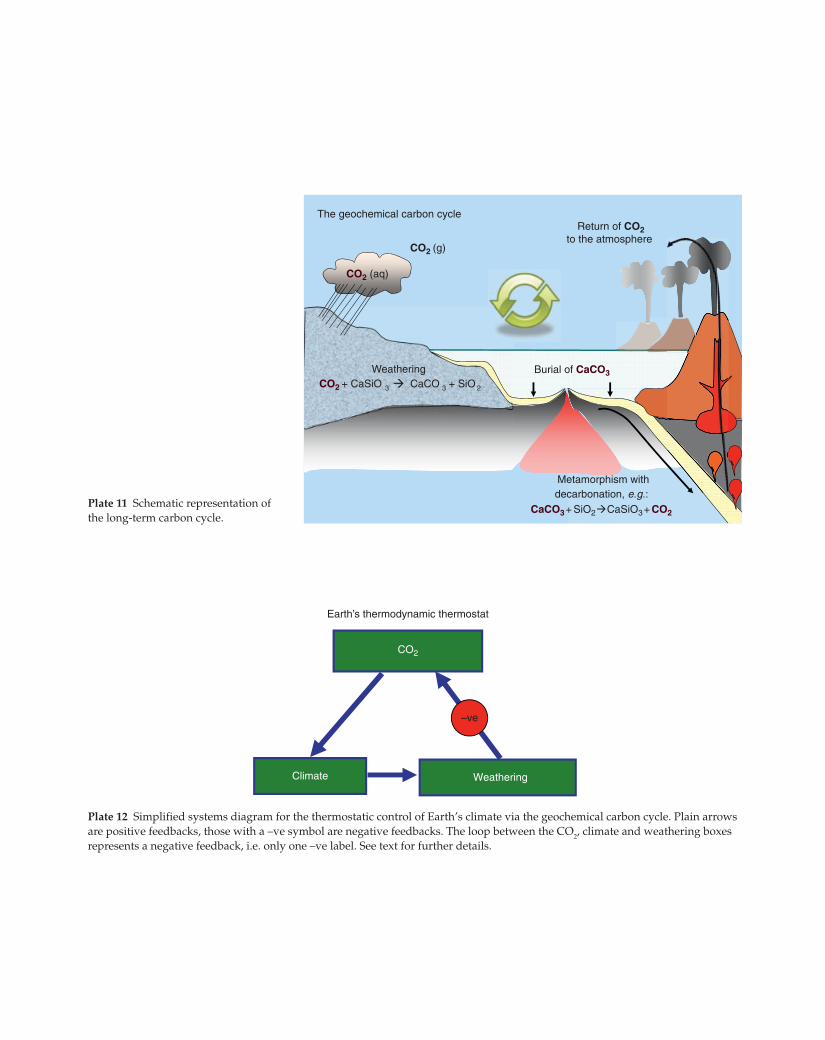
Return of CO2to the atmosphere
Burial of CaCO3WeatheringCO2 + CaSiO 3 CaCO 3 + SiO 2
Metamorphism withdecarbonation, e.g.:
The geochemical carbon cycle
CO2 (aq)
CO2 (g)
CaCO3+ SiO2 CaSiO3+ CO2Plate 11 Schematic representation of
the long-term carbon cycle.
Earth’s thermodynamic thermostat
WeatheringClimate
CO2
–ve
Plate 12 Simplified systems diagram for the thermostatic control of Earth’s climate via the geochemical carbon cycle. Plain arrows
are positive feedbacks, those with a –ve symbol are negative feedbacks. The loop between the CO2, climate and weathering boxes
represents a negative feedback, i.e. only one –ve label. See text for further details.
Knoll_bins.indd 9Knoll_bins.indd 9 2/16/2012 2:12:13 AM2/16/2012 2:12:13 AM
Linking taxonomic diversitywith metabolic activity e.g.
Raman-FISH, SIPs
Meta-genomics,transcriptomics and
proteomics
Niche characterizationEcological lifestyle
Population/community/ecosystemdynamics
Systems biologymetabolic networks
Biodiversity assessments:Who’s there?
Metabolic potential and activityassessments:
What are they doing?
Targeted enrichment
culturing
Patterns of microbial diversity
?
16S rRNA gene assessments
Statistical analyses linkingenvironmental metadatawith community structure
ABC
D GEF
JIH
13C-labeledsubstrate DNA or
RNA
Ecosystem models, testmodel predictions
Classical geneticsand biochemistry
12C13C
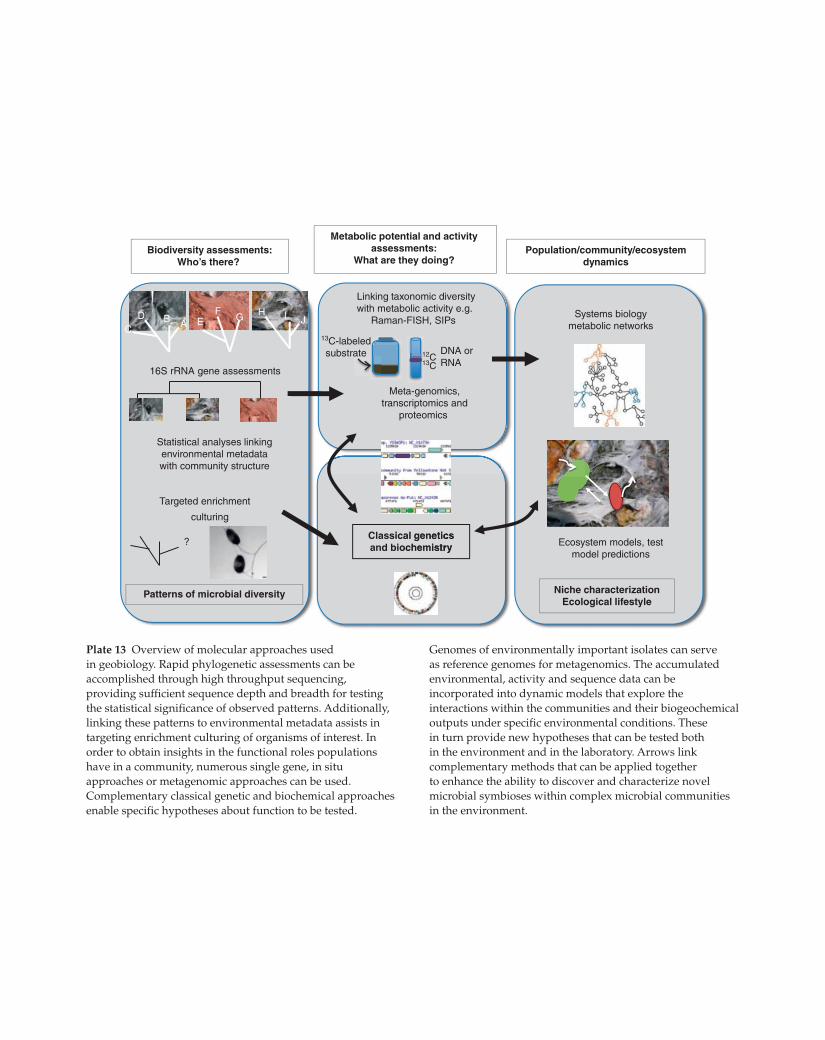
Plate 13 Overview of molecular approaches used
in geobiology. Rapid phylogenetic assessments can be
accomplished through high throughput sequencing,
providing sufficient sequence depth and breadth for testing
the statistical significance of observed patterns. Additionally,
linking these patterns to environmental metadata assists in
targeting enrichment culturing of organisms of interest. In
order to obtain insights in the functional roles populations
have in a community, numerous single gene, in situ
approaches or metagenomic approaches can be used.
Complementary classical genetic and biochemical approaches
enable specific hypotheses about function to be tested.
Genomes of environmentally important isolates can serve
as reference genomes for metagenomics. The accumulated
environmental, activity and sequence data can be
incorporated into dynamic models that explore the
interactions within the communities and their biogeochemical
outputs under specific environmental conditions. These
in turn provide new hypotheses that can be tested both
in the environment and in the laboratory. Arrows link
complementary methods that can be applied together
to enhance the ability to discover and characterize novel
microbial symbioses within complex microbial communities
in the environment.
Knoll_bins.indd 10Knoll_bins.indd 10 2/16/2012 2:12:14 AM2/16/2012 2:12:14 AM
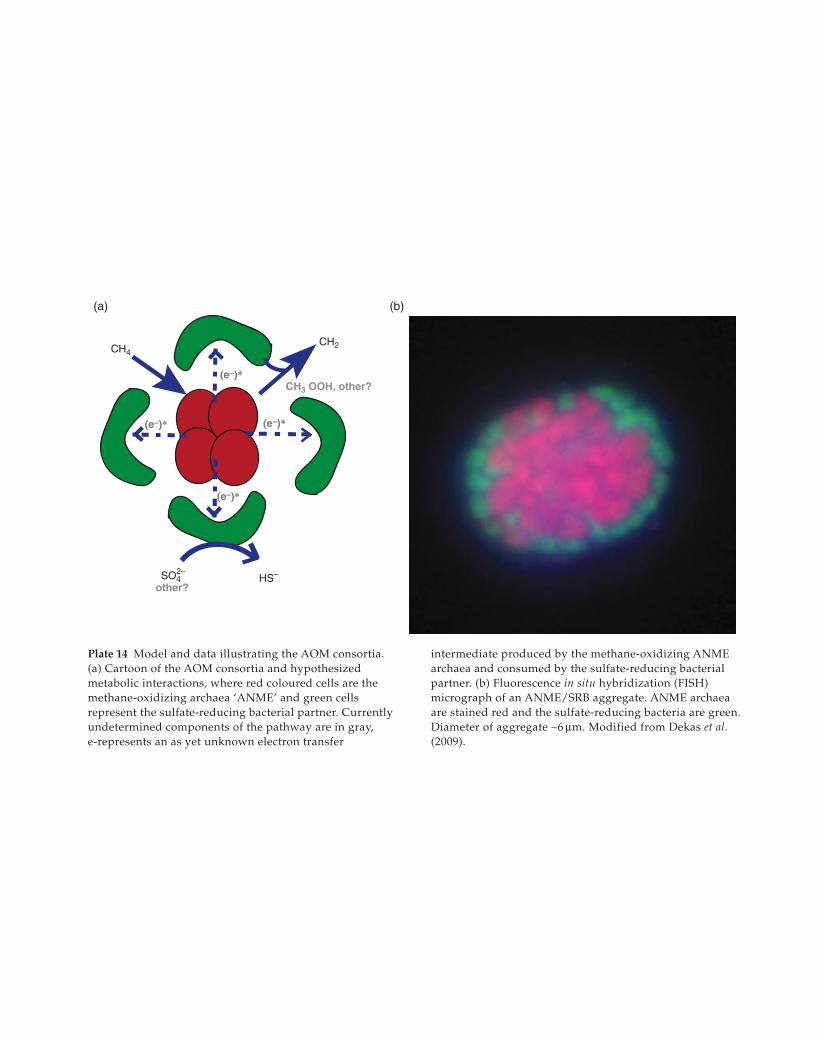
SO42–
HS–
CH4
(a) (b)
CH2
(e–)*
(e–)*
(e–)*
(e–)*
CH3 OOH, other?
other?
Plate 14 Model and data illustrating the AOM consortia.
(a) Cartoon of the AOM consortia and hypothesized
metabolic interactions, where red coloured cells are the
methane-oxidizing archaea ‘ANME’ and green cells
represent the sulfate-reducing bacterial partner. Currently
undetermined components of the pathway are in gray,
e-represents an as yet unknown electron transfer
intermediate produced by the methane-oxidizing ANME
archaea and consumed by the sulfate-reducing bacterial
partner. (b) Fluorescence in situ hybridization (FISH)
micrograph of an ANME/SRB aggregate. ANME archaea
are stained red and the sulfate-reducing bacteria are green.
Diameter of aggregate ∼6 μm. Modified from Dekas et al. (2009).
Knoll_bins.indd 11Knoll_bins.indd 11 2/16/2012 2:12:16 AM2/16/2012 2:12:16 AM
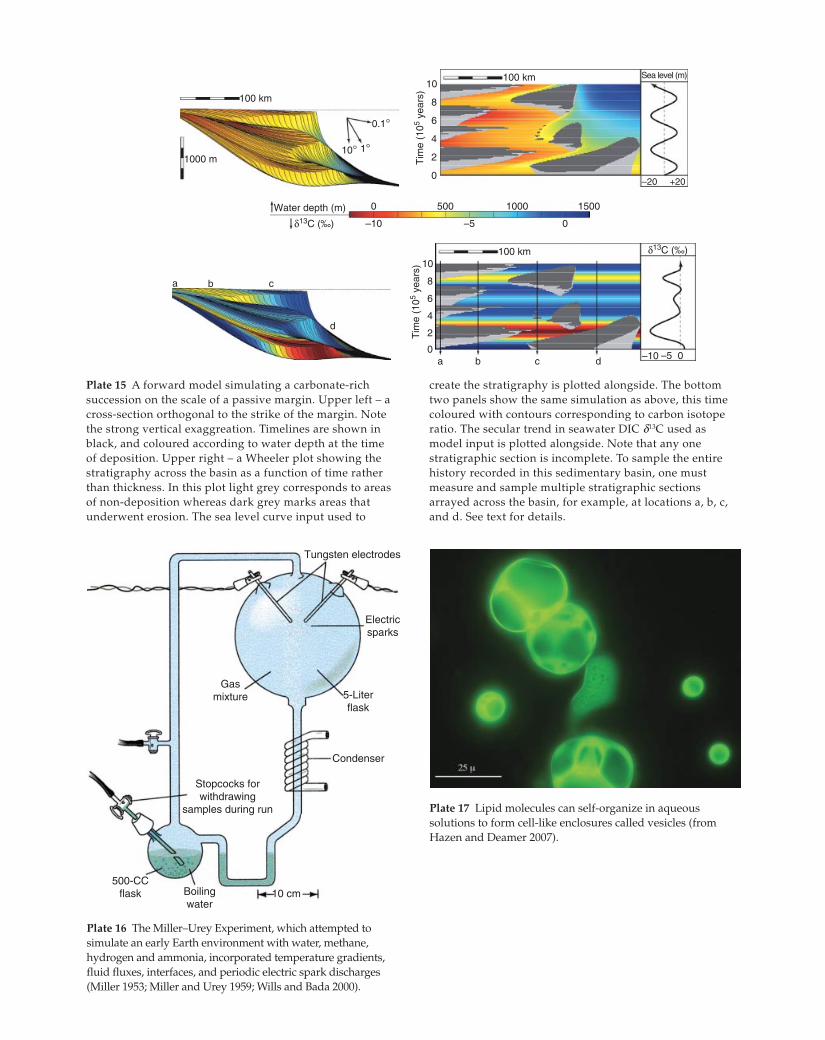
Tungsten electrodes
Electricsparks
5-Literflask
Condenser
10 cmBoilingwater
500-CCflask
Stopcocks forwithdrawing
samples during run
Gasmixture
Plate 16 The Miller–Urey Experiment, which attempted to
simulate an early Earth environment with water, methane,
hydrogen and ammonia, incorporated temperature gradients,
fluid fluxes, interfaces, and periodic electric spark discharges
(Miller 1953; Miller and Urey 1959; Wills and Bada 2000).
Plate 17 Lipid molecules can self-organize in aqueous
solutions to form cell-like enclosures called vesicles (from
Hazen and Deamer 2007).
–20
Sea level (m)
+20
10100 km
8
6
4
2
0
Tim
e (1
05 ye
ars)
1000 m
100 km
10° 1°
0.1°
Water depth (m)
–10
0 500 1000 1500
0–5δ13C (‰)
a b c
d
100 km10
8
6
4
2
0a b c d –10 –5 0
Tim
e (1
05 ye
ars)
δ13C (‰)
Plate 15 A forward model simulating a carbonate-rich
succession on the scale of a passive margin. Upper left – a
cross-section orthogonal to the strike of the margin. Note
the strong vertical exaggreation. Timelines are shown in
black, and coloured according to water depth at the time
of deposition. Upper right – a Wheeler plot showing the
stratigraphy across the basin as a function of time rather
than thickness. In this plot light grey corresponds to areas
of non-deposition whereas dark grey marks areas that
underwent erosion. The sea level curve input used to
create the stratigraphy is plotted alongside. The bottom
two panels show the same simulation as above, this time
coloured with contours corresponding to carbon isotope
ratio. The secular trend in seawater DIC d13C used as
model input is plotted alongside. Note that any one
stratigraphic section is incomplete. To sample the entire
history recorded in this sedimentary basin, one must
measure and sample multiple stratigraphic sections
arrayed across the basin, for example, at locations a, b, c,
and d. See text for details.
Knoll_bins.indd 12Knoll_bins.indd 12 2/16/2012 2:12:17 AM2/16/2012 2:12:17 AM
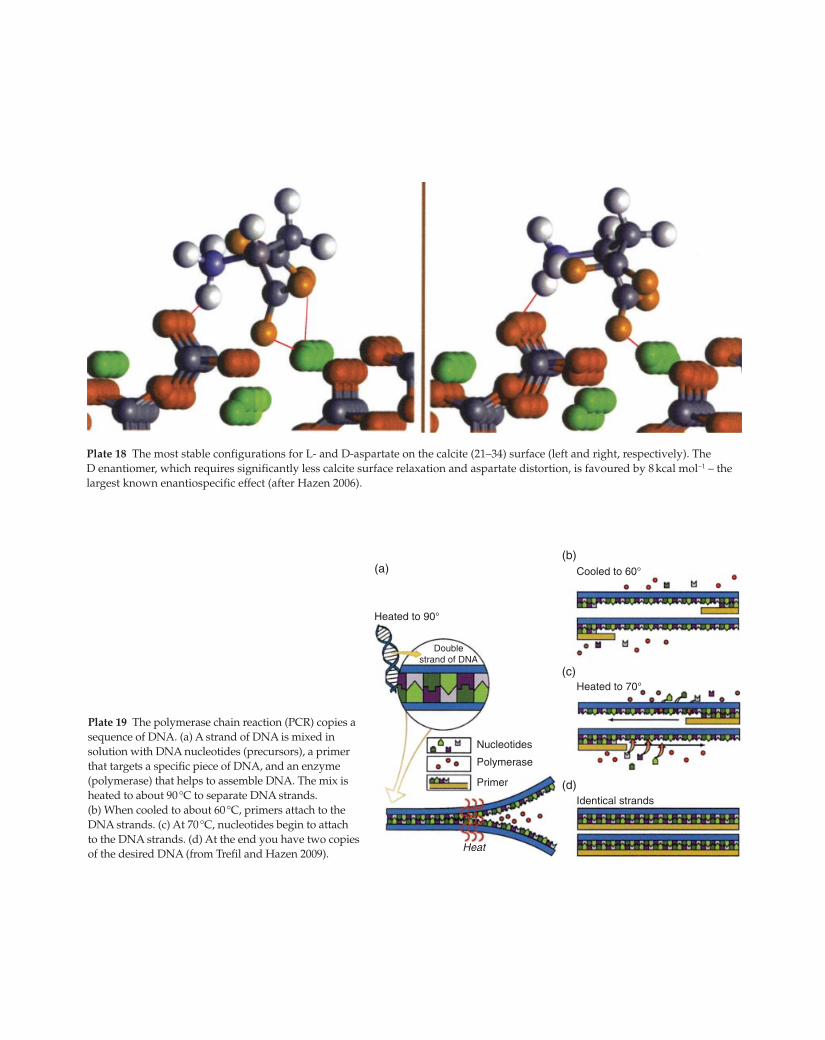
Plate 18 The most stable configurations for L- and D-aspartate on the calcite (21–34) surface (left and right, respectively). The
D enantiomer, which requires significantly less calcite surface relaxation and aspartate distortion, is favoured by 8 kcal mol−1 – the
largest known enantiospecific effect (after Hazen 2006).
(a)
(d)
(c)
(b)Cooled to 60°
Heated to 70°
Heated to 90°
Identical strands
Heat
Primer
Nucleotides
Doublestrand of DNA
Polymerase
Plate 19 The polymerase chain reaction (PCR) copies a
sequence of DNA. (a) A strand of DNA is mixed in
solution with DNA nucleotides (precursors), a primer
that targets a specific piece of DNA, and an enzyme
(polymerase) that helps to assemble DNA. The mix is
heated to about 90 °C to separate DNA strands.
(b) When cooled to about 60 °C, primers attach to the
DNA strands. (c) At 70 °C, nucleotides begin to attach
to the DNA strands. (d) At the end you have two copies
of the desired DNA (from Trefil and Hazen 2009).
Knoll_bins.indd 13Knoll_bins.indd 13 2/16/2012 2:12:20 AM2/16/2012 2:12:20 AM
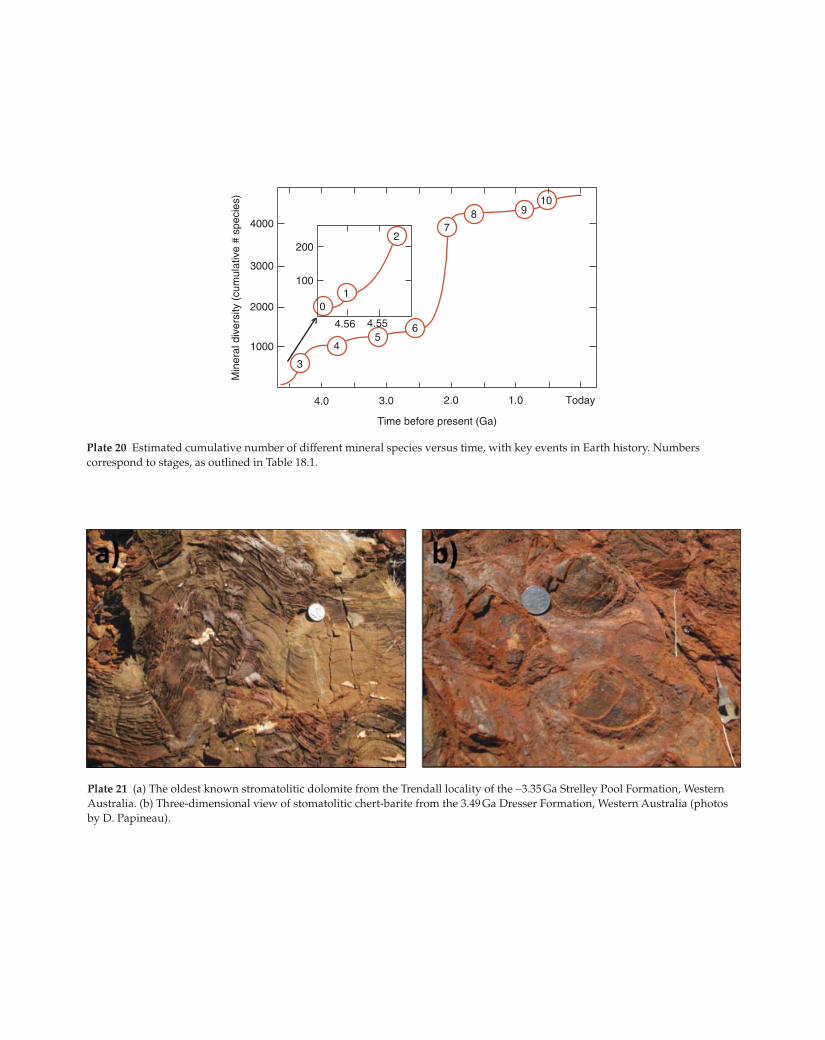
Plate 21 (a) The oldest known stromatolitic dolomite from the Trendall locality of the ∼3.35 Ga Strelley Pool Formation, Western
Australia. (b) Three-dimensional view of stomatolitic chert-barite from the 3.49 Ga Dresser Formation, Western Australia (photos
by D. Papineau).
Time before present (Ga)
4000
3000
2000
1000
Min
eral
div
ersi
ty (
cum
ulat
ive
# sp
ecie
s)
4.0 3.0 2.0
4
3
56
72
10
200
100
4.56 4.55
8 910
1.0 Today
Plate 20 Estimated cumulative number of different mineral species versus time, with key events in Earth history. Numbers
correspond to stages, as outlined in Table 18.1.
Knoll_bins.indd 14Knoll_bins.indd 14 2/16/2012 2:12:23 AM2/16/2012 2:12:23 AM

Plate 22 (a) Slab of a stromatolitic jasper banded iron
formation from the ∼1.88 Ga Biwabik Formation, Minnesota,
USA, (b) stromatolitic chert-dolomite from the ∼1.9 Ga
McLeary Formation, Belcher Islands, Canada, (c) stromatolitic
dolomite from the 2.06–2.09 Ga Rantamaa Formation, Finland,
(d) stromatolitic phosphorite (carbonate-fluorapatite and
dolomite) from the 2.0 Ga Jhamarkotra Formation, Rajasthan,
India (photos by D. Papineau).
Knoll_bins.indd 15Knoll_bins.indd 15 2/16/2012 2:12:28 AM2/16/2012 2:12:28 AM

Plate 23 (a) Chert-magnetite-siderite banded iron formation
from the Neoarchean Hunter Mine Group, Canada, (b)
chert-magnetite banded iron formation from the Neoarchean
Bababudan Supracrustal Belt, India, (c) jasper-chert banded
iron formation from the Paleoproterozoic Negaunee Formation,
Michigan, United States, (d) sulfide facies banded iron
formation from the ∼2.0 Ga Pathavaara Formation, Finland. All
coins are about 2.5 cm in diameter (photos by D. Papineau).
Knoll_bins.indd 16Knoll_bins.indd 16 2/16/2012 2:12:36 AM2/16/2012 2:12:36 AM
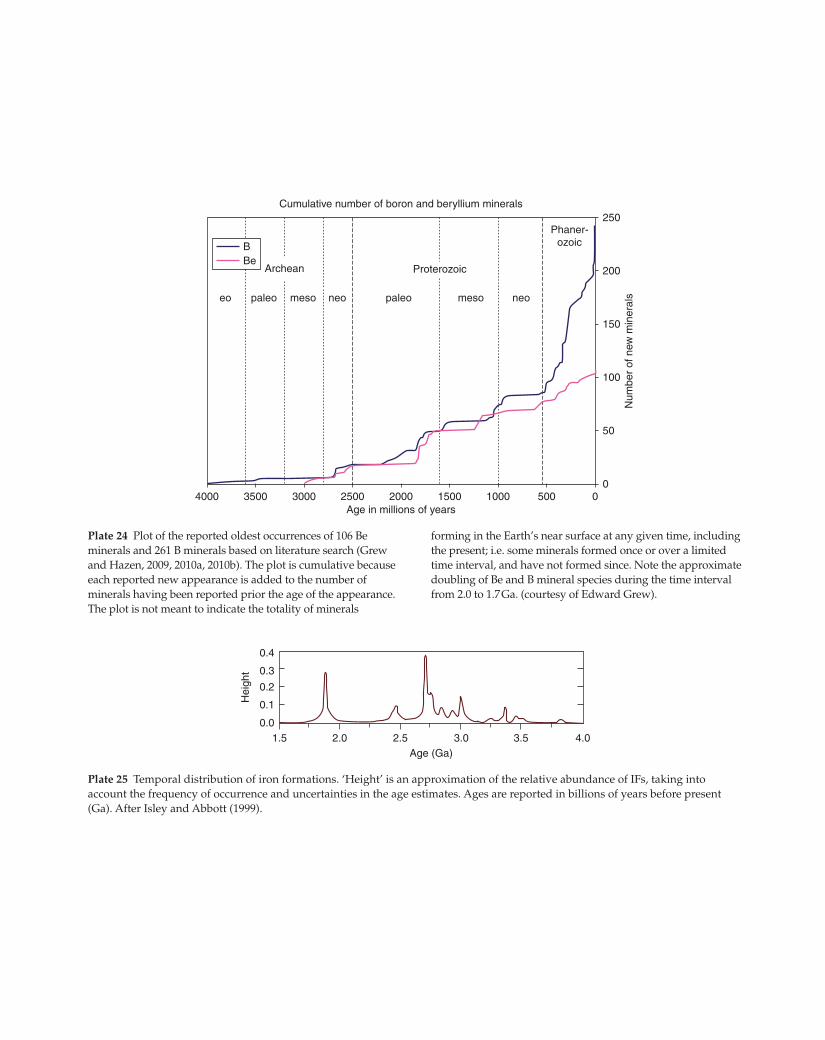
1.50.0
0.1
0.2
0.3
0.4
2.0 2.5Age (Ga)
Hei
ght
3.0 3.5 4.0
Plate 25 Temporal distribution of iron formations. ‘Height’ is an approximation of the relative abundance of IFs, taking into
account the frequency of occurrence and uncertainties in the age estimates. Ages are reported in billions of years before present
(Ga). After Isley and Abbott (1999).
BBe
Cumulative number of boron and beryllium minerals
Archean
eo paleo meso neo paleo
Proterozoic
meso neo
Phaner-ozoic
4000 3500 2500 1500 1000 500 00
50
100
150
200
250
3000 2000Age in millions of years
Num
ber
of n
ew m
iner
als
Plate 24 Plot of the reported oldest occurrences of 106 Be
minerals and 261 B minerals based on literature search (Grew
and Hazen, 2009, 2010a, 2010b). The plot is cumulative because
each reported new appearance is added to the number of
minerals having been reported prior the age of the appearance.
The plot is not meant to indicate the totality of minerals
forming in the Earth’s near surface at any given time, including
the present; i.e. some minerals formed once or over a limited
time interval, and have not formed since. Note the approximate
doubling of Be and B mineral species during the time interval
from 2.0 to 1.7 Ga. (courtesy of Edward Grew).
Knoll_bins.indd 17Knoll_bins.indd 17 2/16/2012 2:12:45 AM2/16/2012 2:12:45 AM
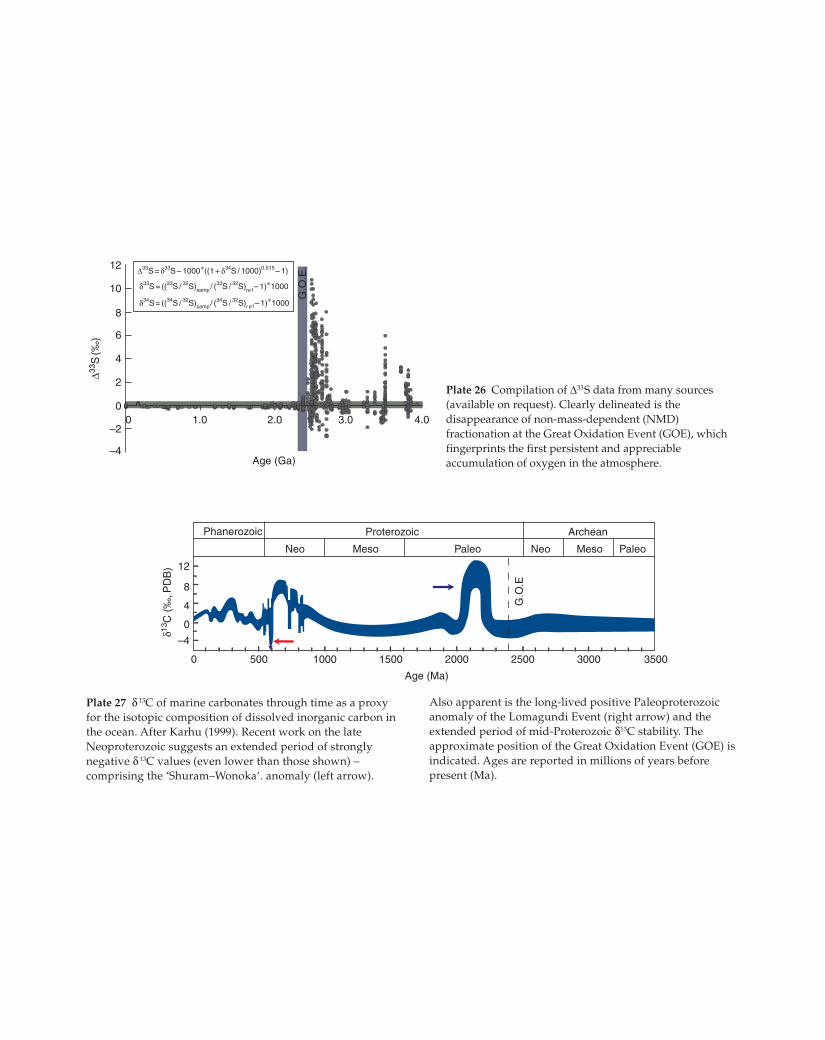
250020001500
Neo NeoMeso Meso
Archean
Paleo Paleo
Age (Ma)
10005000
–4
0
4
8
12
Phanerozoic Proterozoic
3000 3500
δ13C
(‰
, PD
B)
G.O
.E
Plate 27 δ 13C of marine carbonates through time as a proxy
for the isotopic composition of dissolved inorganic carbon in
the ocean. After Karhu (1999). Recent work on the late
Neoproterozoic suggests an extended period of strongly
negative δ 13C values (even lower than those shown) –
comprising the ‘Shuram–Wonoka‘. anomaly (left arrow).
Also apparent is the long-lived positive Paleoproterozoic
anomaly of the Lomagundi Event (right arrow) and the
extended period of mid-Proterozoic δ13C stability. The
approximate position of the Great Oxidation Event (GOE) is
indicated. Ages are reported in millions of years before
present (Ma).
G.O
.E.
4
2
0
Δ33S
(‰)
–2
–4
0 1.0
Δ33S = δ33S – 1000∗((1 + δ34S / 1000)0.515– 1)
δ34S = ((34S / 32S)samp/ (34S / 32S)r e f– 1)∗1000
δ33S = ((33S / 32S)samp/ (33S / 32S)re f– 1)∗1000
2.0
Age (Ga)
3.0 4.0
6
8
10
12
Plate 26 Compilation of Δ33S data from many sources
(available on request). Clearly delineated is the
disappearance of non-mass-dependent (NMD)
fractionation at the Great Oxidation Event (GOE), which
fingerprints the first persistent and appreciable
accumulation of oxygen in the atmosphere.
Knoll_bins.indd 18Knoll_bins.indd 18 2/16/2012 2:12:46 AM2/16/2012 2:12:46 AM
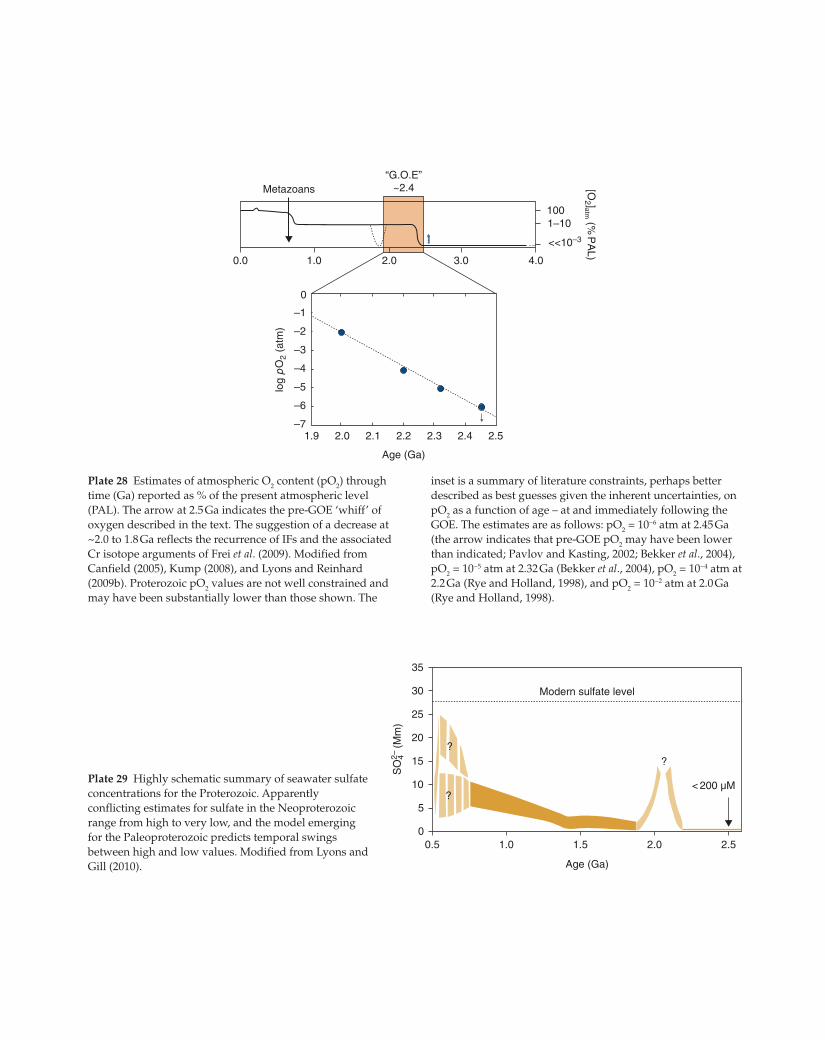
–1
–2
–3
–4
–5
–6
–7
0
1.9 2.0 2.1 2.2 2.3 2.4 2.5
0.0 1.0 2.0 3.0 4.0
Age (Ga)
Metazoans
1001–10
<<10–3
“G.O.E”~2.4
log
pO
2 (a
tm)
[O2 ]atm (%
PA
L)
Plate 28 Estimates of atmospheric O2 content (pO
2) through
time (Ga) reported as % of the present atmospheric level
(PAL). The arrow at 2.5 Ga indicates the pre-GOE ‘whiff’ of
oxygen described in the text. The suggestion of a decrease at
~2.0 to 1.8 Ga reflects the recurrence of IFs and the associated
Cr isotope arguments of Frei et al. (2009). Modified from
Canfield (2005), Kump (2008), and Lyons and Reinhard
(2009b). Proterozoic pO2 values are not well constrained and
may have been substantially lower than those shown. The
inset is a summary of literature constraints, perhaps better
described as best guesses given the inherent uncertainties, on
pO2 as a function of age – at and immediately following the
GOE. The estimates are as follows: pO2 = 10−6 atm at 2.45 Ga
(the arrow indicates that pre-GOE pO2 may have been lower
than indicated; Pavlov and Kasting, 2002; Bekker et al., 2004),
pO2 = 10−5 atm at 2.32 Ga (Bekker et al., 2004), pO
2 = 10−4 atm at
2.2 Ga (Rye and Holland, 1998), and pO2 = 10−2 atm at 2.0 Ga
(Rye and Holland, 1998).
SO
42–(M
m)
35
30
25
20
15
10
5
00.5 1.0 1.5 2.0 2.5
Age (Ga)
< 200 μM
?
Modern sulfate level
?
?
Plate 29 Highly schematic summary of seawater sulfate
concentrations for the Proterozoic. Apparently
conflicting estimates for sulfate in the Neoproterozoic
range from high to very low, and the model emerging
for the Paleoproterozoic predicts temporal swings
between high and low values. Modified from Lyons and
Gill (2010).
Knoll_bins.indd 19Knoll_bins.indd 19 2/16/2012 2:12:46 AM2/16/2012 2:12:46 AM
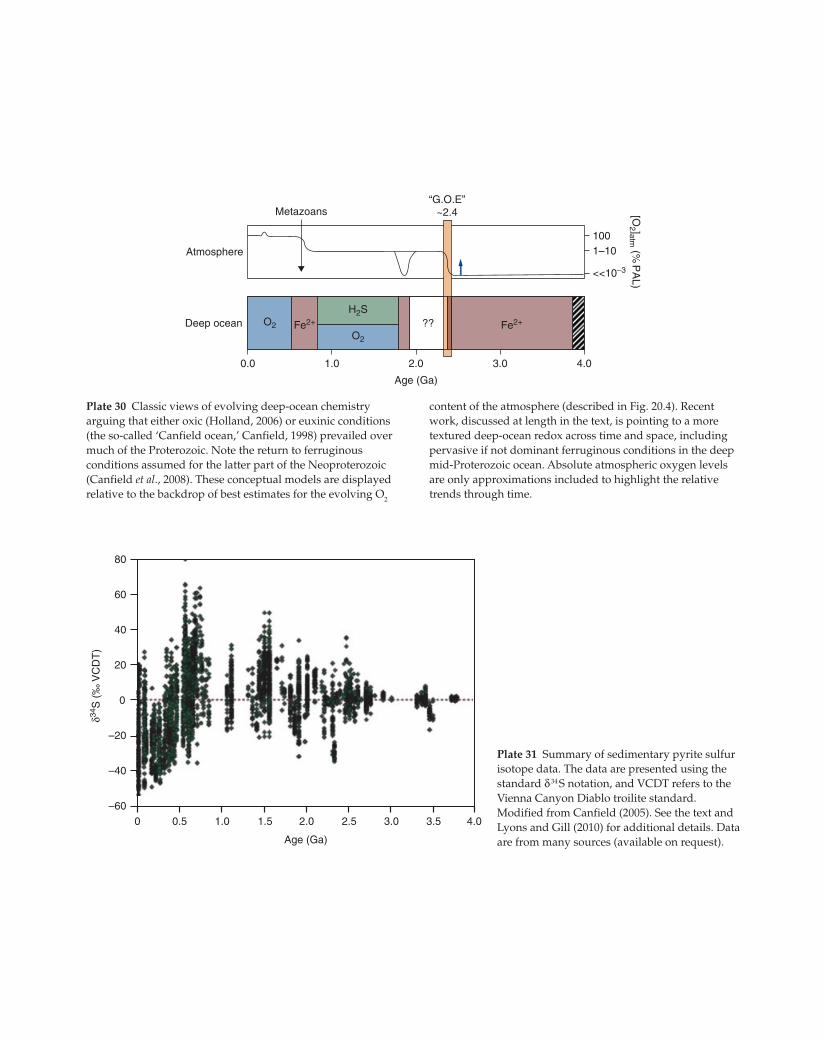
60
40
20
0
–20
–40
–600
δ34S
(‰
VC
DT
)
0.5 1.0 1.5 2.0
Age (Ga)
2.5 3.0 3.5 4.0
80
Plate 31 Summary of sedimentary pyrite sulfur
isotope data. The data are presented using the
standard δ 34 S notation, and VCDT refers to the
Vienna Canyon Diablo troilite standard.
Modified from Canfield (2005). See the text and
Lyons and Gill (2010) for additional details. Data
are from many sources (available on request).
O2O2
Fe2+ Fe2+
H2S
0.0 1.0 2.0 3.0 4.0
Age (Ga)
Atmosphere
Deep ocean
Metazoans“G.O.E”
~2.4
1001–10
<<10–3
[O2 ]atm
(%P
AL)
??
Plate 30 Classic views of evolving deep-ocean chemistry
arguing that either oxic (Holland, 2006) or euxinic conditions
(the so-called ‘Canfield ocean,’ Canfield, 1998) prevailed over
much of the Proterozoic. Note the return to ferruginous
conditions assumed for the latter part of the Neoproterozoic
(Canfield et al., 2008). These conceptual models are displayed
relative to the backdrop of best estimates for the evolving O2
content of the atmosphere (described in Fig. 20.4). Recent
work, discussed at length in the text, is pointing to a more
textured deep-ocean redox across time and space, including
pervasive if not dominant ferruginous conditions in the deep
mid-Proterozoic ocean. Absolute atmospheric oxygen levels
are only approximations included to highlight the relative
trends through time.
Knoll_bins.indd 20Knoll_bins.indd 20 2/16/2012 2:12:47 AM2/16/2012 2:12:47 AM
δ13C (‰)
Pristane & phytane
Pristane & phytane
ca. 550 Myr
n-alkanesn-alkanes
Kerogen
Kerogen–26
–29
–32
“Precambrian” “Phanerozoic”
Plate 33 A fundamental switch in stable carbon isotopic
patterns (δ13C) is found for hydrocarbon constituents of
extractable rock bitumen (free n-alkanes and the acyclic
isoprenoids, pristine and phytane) versus kerogen (from
Logan et al., 1995) for ancient sedimentary rocks. The switch
in isotopic ordering appears to occur around 550 million years
ago (Kelly et al., 2008). Absolute δ13C (‰) values vary from
sample to sample.
(a)
(b)
(c)
[O2] ( )
[ΣH2S] ( )
[CH2O]xH2S
FeS2
SO42–
Fe2+
FeS2
[ΣH2S] ( )
(?)Fe2+
[CH2O]xH2S
FeS2
FeOOH(?)
[O2] ( )
[O2] ( )SO4
2–
(?)
[Fe2+] ( )
[O2] ( ) [O2] ( )SO4
2–
FeOOH(?)
(?)
[Fe ] ( )
Fe2+
[CH2O]xH2S
FeS2
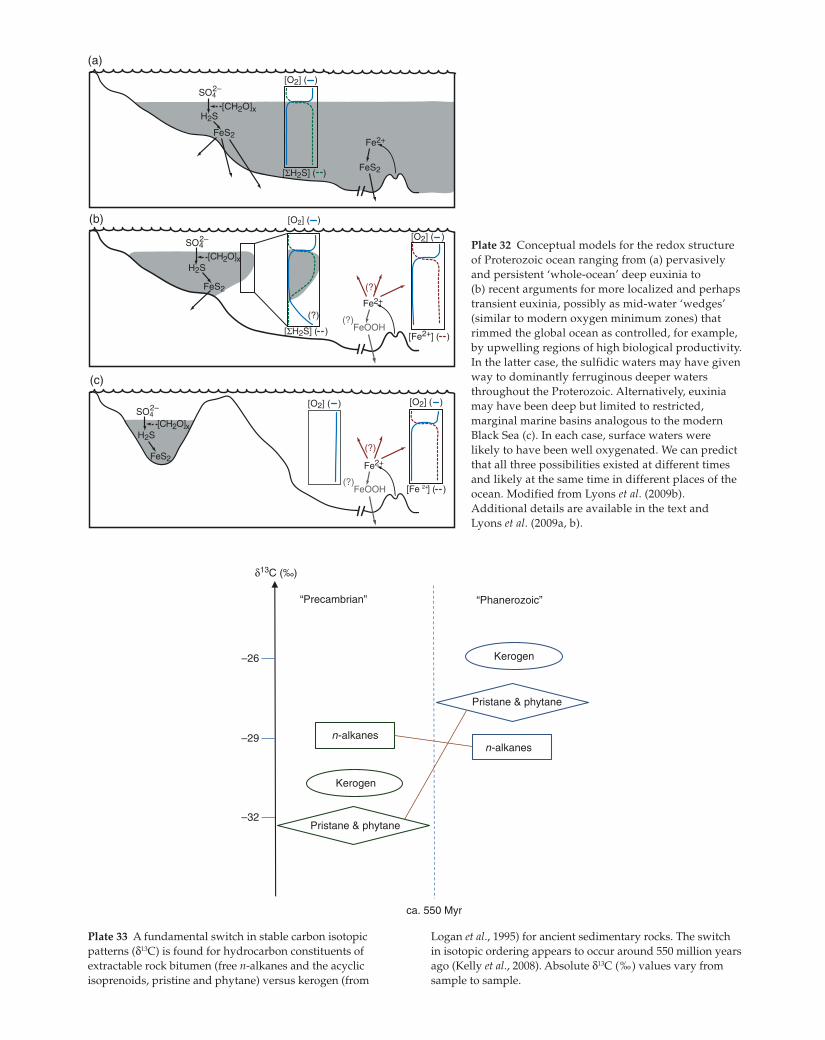
Plate 32 Conceptual models for the redox structure
of Proterozoic ocean ranging from (a) pervasively
and persistent ‘whole-ocean’ deep euxinia to
(b) recent arguments for more localized and perhaps
transient euxinia, possibly as mid-water ‘wedges’
(similar to modern oxygen minimum zones) that
rimmed the global ocean as controlled, for example,
by upwelling regions of high biological productivity.
In the latter case, the sulfidic waters may have given
way to dominantly ferruginous deeper waters
throughout the Proterozoic. Alternatively, euxinia
may have been deep but limited to restricted,
marginal marine basins analogous to the modern
Black Sea (c). In each case, surface waters were
likely to have been well oxygenated. We can predict
that all three possibilities existed at different times
and likely at the same time in different places of the
ocean. Modified from Lyons et al. (2009b).
Additional details are available in the text and
Lyons et al. (2009a, b).
Knoll_bins.indd 21Knoll_bins.indd 21 2/16/2012 2:12:48 AM2/16/2012 2:12:48 AM
Opi
stho
kont
sAm
oebo
zoan
s
Red algae
Green algae
& land plantsH
apto
phyt
esDiat
oms
Brown algae
Chromalveolates
Plants
Rhizarians
Excavates
Choanoflagellates
Animals
Fungi
Lobose amoebae
Slime m
olds
Cili
ates
Api
com
plex
ans
Din
ofla
gella
tes
Oom
ycet
es
Chrysophyte
s
Foraminifers
RadiolariansCercozoansEuglenidsKinetoplastids
Parabasalids
Diplom
onads
Xanthophyte
algae
Uni
kont
s
Bikonts
1200 Ma
750 Ma
750
Ma
750 Ma
1000 Ma
Tappania850 Mafungus?
Melanocyrillium~750 Ma
lobose amoebae
Melicerion~750 Ma
cercozoan
Palaeovaucheria>1000 Ma
xanthophyteBangiomorpha~1200 Mared alga
Protocladus~750 Ma
green alga
Chlorarachniophytes
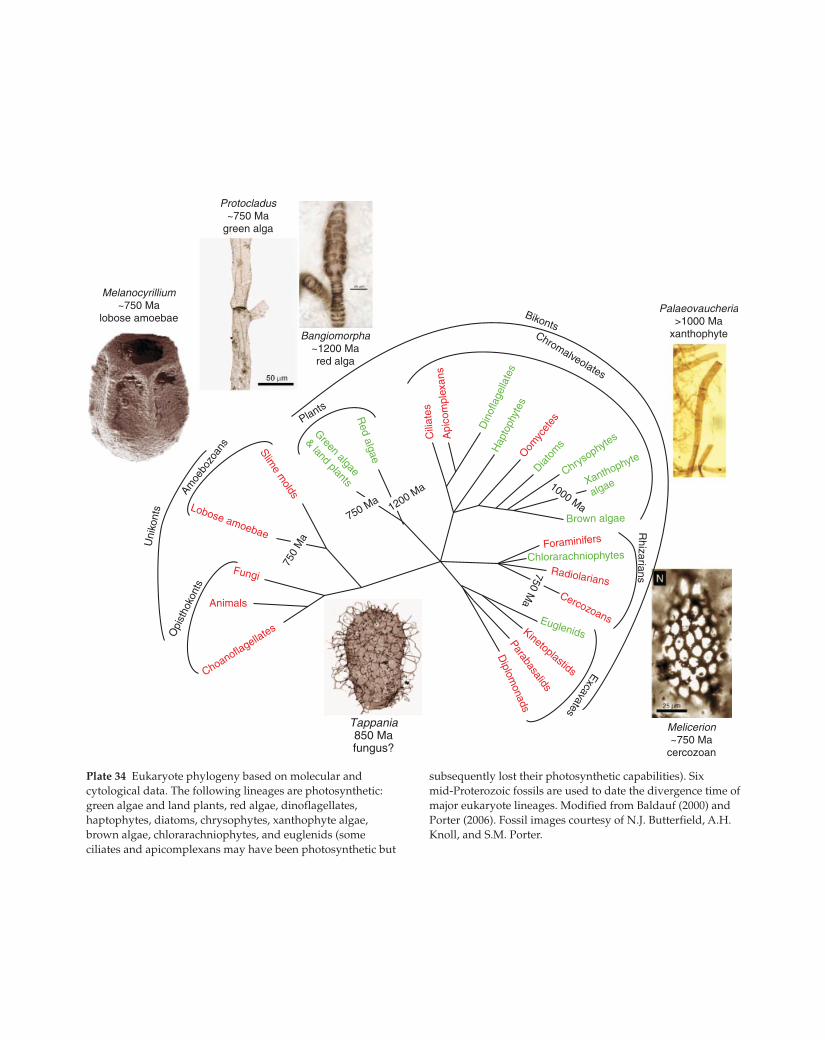
Plate 34 Eukaryote phylogeny based on molecular and
cytological data. The following lineages are photosynthetic:
green algae and land plants, red algae, dinoflagellates,
haptophytes, diatoms, chrysophytes, xanthophyte algae,
brown algae, chlorarachniophytes, and euglenids (some
ciliates and apicomplexans may have been photosynthetic but
subsequently lost their photosynthetic capabilities). Six
mid-Proterozoic fossils are used to date the divergence time of
major eukaryote lineages. Modified from Baldauf (2000) and
Porter (2006). Fossil images courtesy of N.J. Butterfield, A.H.
Knoll, and S.M. Porter.
Knoll_bins.indd 22Knoll_bins.indd 22 2/16/2012 2:12:48 AM2/16/2012 2:12:48 AM
Kimberella
540
545
550
555
560
565
570
575
580
585
590
595
600
605
610
615
620
625
630
635
640
Edi
acar
anC
ambr
ian
Cry
ogen
ian
–8 –4 0 4
δ13C (‰)
Marinoan (Nantuo) Glaciation
Gaskiers Glaciation
EN3 (Shuram)
EN1
EN2
Sou
th C
hina
DP
A
Aus
tral
ian
DP
A
Dou
shan
tuo
anim
al e
mbr
yos,
cni
daria
ns(?
)
Larg
e st
em-g
roup
ani
mal
s(?)
Larg
e m
obile
bila
teria
ns
Ani
mal
bio
min
eral
izat
ion
Dou
shan
tuo
mac
roal
gae
Fractofusus
Kimberella
Cloudina
Konglingiphyton
Megaclonophycus
Tianzhushania
Meghystricho-sphaeridium
Tanarium
Sinocyclocyclicus
?? ??
????
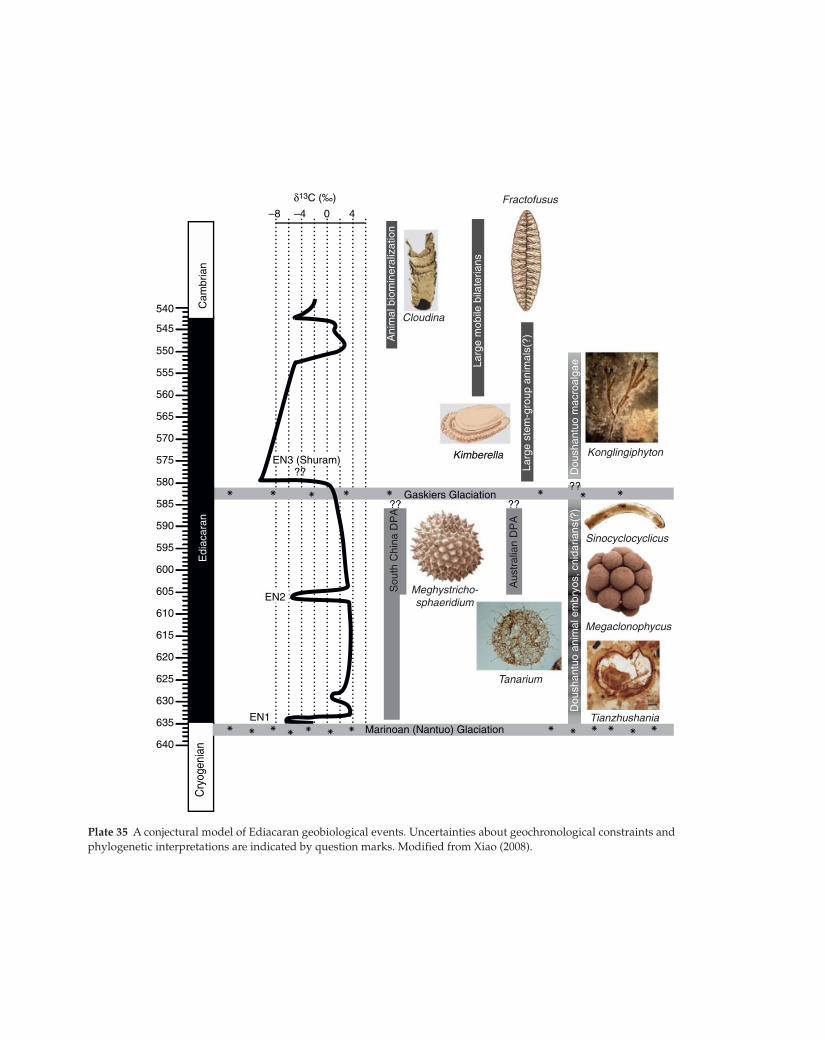
Plate 35 A conjectural model of Ediacaran geobiological events. Uncertainties about geochronological constraints and
phylogenetic interpretations are indicated by question marks. Modified from Xiao (2008).
Knoll_bins.indd 23Knoll_bins.indd 23 2/16/2012 2:12:49 AM2/16/2012 2:12:49 AM

Age (Myr)
Neogene
Penn.
Miss.
Silu
r.P
aleo
c.C
reta
ceou
sJu
rass
icT
riass
icP
erm
ian
Dev
onia
nO
rdov
icia
nC
ambr
ian
Paleoc.
Eoc.
Early
Oligoc.
Late
Early
Late
Early
Middle
Late
Early
Late
Middle
Late
Early
MiddleLate
Early
Middle
Middle
Late
EarlyMiddle
Early
Middle
Late
50
100
150
200
250
300
350
400
450
500
F
J
K
M
R
O
B
A
G
C
H
I
L
P
D
N
Q
(o)?
Ordovician
Silurian
0 1 2 3 64 5 0–2 –1–3–4–5
Sedgwickiizone
Convolutuszone
–27–28–29–30–31
(n)
(m)
Llandov.
Wenl.
0 1 2 3 64 5–1 –2 –1–3–4–5–6
(l)Ludlow
Wenlock
–2–3–4–5–60 1 2 3 64 5 7
(d)
E. Trias.
M. Trias.
0 1 2 3 4–1 7 8
zoneYabeina
Neoschw.zone
EarlyGuad.
5 6 73 4
(f)
(h)
Serp.
Tournais.
Visean
21 222019 230 1 2 3 64 5
(p)
Mohawk
Chazyan
0 1 2 3 4–2 –1–3
Marj.
Delam.
E. Cam.
0 1 2–2 –1–3 –4–5–6–7–8–9–10–11
(r)Marj.
Sunwapt.
Stept.
0 1 2 3 4–2 –1–3 5
(q)δ13C
δ13C
δ13C
δ13C
δ13C
δ13C
δ13C
δ13C δ13C
δ13C
δ13C
δ13C
δ13C
δ13C
δ13C
δ13C
δ13C
δ13C
0 1 2 3 4–2 –1–3 –4–5–6–7–8–9–10
Silur.
Devon.(k)
(j)Givet.
Frasn.
Famenn.
–3–4–5–60 1 2 3
(i)
Mississippian
1 2 3 64 5 7 –2 –1–3–4–5–6–7–8 0
Famennian
E
Guad.
Loping.
0 1 2 3 4 5–2 –1–3 –3–4–5–6–7
(g)
(a)
34Ma
Eoc.
Olig.
0 1 2 0 1 2
33Ma
(b)
Paleoc.
Eoc.
0 1 2–1
57.2Ma
57.4Ma
0–2 –1
Perm.
Trias.
–2 –1–3–4–5–6–70 1 2 3 4–2 –1
(e)
–6–7–8–9–10
Stept.
Marj.
δ18O
δ18O
δ18O
δ18O
δ18O
δ18O
δ18O
δ18O
δ18O
δ18O
δ18O
δ18O δ18O
δ18O
(c)
Trias.
Jur.
0 1 2–2 –11 2 3 4 5
Plate 36 Stable isotope excursions that have been documented
in shallow marine strata in association with mass extinctions.
Eighteen intervals (A–R) contain a total of 26 such δ13 C
excursions. Corresponding to these, and trending in the same
direction, are 19 published δ18 O excursions, which are
displayed in the plots to the right of those depicting δ13 C.
Encircled letters on the left indicate temporal positions of
excursions. Blue indicates association with global cooling and
red, with global warming; black indicates absence of published
evidence of associated climate change. Horizontal scales
represent magnitudes of δ13 C and δ18 O excursions in ‰. Light
δ13 C in N is for organic carbon rather than carbonates, and
heavy δ18 O in H is for conodonts rather than bulk or skeletal
carbonate. Ordinates represent stratigraphic positions of
samples and are neither precisely linear with respect to time
nor scaled the same for all graphs (after Stanley, 2010).
Knoll_bins.indd 24Knoll_bins.indd 24 2/16/2012 2:12:51 AM2/16/2012 2:12:51 AM