functional properties of strychnine-sensitive glycine receptors expressed in xenopus oocytes...
TRANSCRIPT

28 Neuroscience Research, 11 (1991 ) 28 -40 ' 1991 Elsevier Scientific Publishers Ireland, Ltd. 0168-0102/91/$03.50
NEURES 00440
Functional properties of strychnine-sensitive glycine receptors expressed in Xenopus oocytes injected
with a single mRNA
H. Akagi 1.., K. Hirai 2 and F. Hishinuma 2 /
Department of Neurosctence and 2 Department of Molecular Biology, Mitsubishi Kasei Institute of Life Sciences, Tokyo (Japan)
(Received 27 November 1990; Accepted 17 January 1991)
Key words." cDNA cloning; Chloride ion channel; Glycine receptor; Polymerase chain reaction; Site-directed mutagenesis; Xenopus oocytes
SUMMARY
Mature rat spinal cord cDNA libraries constructed in Xgtl0 and XZAPII were screened with an oligonuc- leotide probe (39 mer), and 4 clones that possess DNA-inserts encoding a glyeine receptor subunit were obtained. The cloned cDNAs were used to reconstruct the nucleotide sequence of the full-length open reading frame consisting of 1350 base pairs (bp) as well as the 5'-(184 bp) and 3'-(591 bp) non-coding regions. Synthetic RNA transcribed in vitro from the glycine receptor cDNA induced Xenopus oocytes to synthesize functional glycine receptor that generated large CI- currents. The electrophysiological properties of the wild-type receptor and some mutant receptors produced by site-directed mutagenesis were analyzed.
INTRODUCTION
Glycine is one of the leading neurotransmitters in the vertebrate central nervous system, especially in the spinal cord. In spinal motoneurons, glycine causes neuronal inhibition which is due to an increase in C1- permeability through a specific receptor 6~4°. The glycine receptor, like GABA A and nicotinic acetylcholine receptors, is a complex of ligand-binding sites and ion channel, namely multiple subunit proteins assembled in the neuronal membrane like a cylinder form an ion channel 7'37.
Because the glycinergic system plays an important central role in a variety of physio- logical functions, it is crucial to investigate the properties of glycine receptors at the molecular level, including identification of regions which concern ligand (agonist/ antagonist) binding and ion channel pores. A powerful approach for this type of analysis is to modify the receptor protein using techniques of recombinant D N A and site-directed mutagenesis 9. As a first step in this approach, it is essential to isolate the complementary (c) D N A that encodes the receptor to determine its primary structure. Using a rat spinal cord cDNA library, an attempt was made by Grenningloh et al. t° who reported the
* Present address and correspondence: Hiroyuki Akagi, Department of Pharmacology, Gunma University, School of Medicine, Showa-machi, Maebashi-shi, Gunma 371, Japan.

29
nucleotide sequence of a glycine receptor cDNA and deduced the amino acid sequence of the glycine receptor subunit (referred to as the '48-kDa subunit'). However, the con- structed cDNA lacked the 5'-region including the initiation codon, ATG, and following in-frame sequence encoding a part of the prepro-sequence (signal sequence) of the protein. We have also tried to isolate a clone encoding the full size of the glycine receptor subunit and obtained several cDNAs which overlap and cover the entire open reading frame (ORF) of the receptor protein. Sequential analysis revealed that the ORF of the cDNA is identical to that of the 48-kDa subunit clone except in one base, but the sequence of the 3'-non-coding region differs considerably. A single species of RNA transcribed in vitro from the cloned cDNA induced formation of functional glycine receptors in Xenopus oocytes. Studies on modifying the receptor subunit by point mutation techniques have been carried out to define important regions for glycine receptor function.
MATERIALS AND METHODS
Isolation of cDNAs Total RNA was isolated from spinal cords of mature (8-12 weeks or 25 days old)
Wistar rats by the phenol /ch loroform method and poly(A)-rich fractions were isolated using oligo-dT cellulose 35. A portion of the poly(A) + RNAs from 25-day-old rats was further size-fractionated by sucrose-density-gradient ultracentrifugation as described pre- viously, and each fraction was assessed for its expressional potency of functional glycine receptor in Xenopus oocytes 2"3. Both non- and fractionated RNAs were used for prepara- tion of cDNA libraries. The random as well as oligo-dT primed libraries were constructed with two species of bacteriophage, Xgtl0 and XZAPII as vectors (titer of 3 x 105 to 3 x 10 7 pfu/ml) . The libraries were screened with a 32p-labeled oligodeoxynucleotide, AsLBS (5 ' -GTAGTGCq 'TGGTGCAGTATCTCAGATCCTTCTCTTCCTT-3 ' ) prepared by an automated DNA sYnthesizer (Applied Biosystems). For sequencing, positive cDNAs derived from Xgtl0 libraries were subcloned into plasmid vectors, pUC18 or pUC19, and those in XZAPII were excised directly into pBluescript S K ( - ) in vivo. Nucleotide sequencing was carried out by the chain termination method 28. The sequences of both sense and antisense strands were determined. A cDNA fragment, PCR8, was obtained by the use of polymerase chain reaction (PCR) with two synthetic oligonucleo- tides, LBS ( 5 ' - A A G G A A G A G A A G G A T C T G A G A T A C T G C A C C A A G C A C T A C - 3 ' ) and AsTGA ( 5 ' - G A G C C T T C A C T T G T T G T G G A C G T C C T C T C T C C G G A C G A T C - 3 ' ) as primers.
Reconstruction of glycine receptor cDNAs encodes full-length ORF and functional expression The glycine receptor cDNA that encodes full-length ORF (pGRA1) was constructed
from two clones, p1201 and PCR8. The SacI-PstI fragment of p1201 was replaced by that from PCR8 (the PstI site is in the pUC vectors). The cDNA pGRA1 was subcloned into plasmid pSPT18 or pSPT19, expression vectors bearing T3 and SP6 promoters. After purification of the plasmid DNA by CsC1 centrifugation, they were transcribed in vitro to obtain RNAs encoding the receptor subunit. The RNA was suspended in water at a concentration of 1 /~g//~l (calculated from the OD260 value) after capping at the 5'-end, and stored at - 8 0 ° C. Fifty nl of the RNA solution (50-100 ng//~l) were injected into oocytes of Xenopus laevis. After incubation of the oocytes in modified Barth's solution (for salt compositions, see Ref. 35) for 2-5 days at 20 o C, they were assayed electrophysi- ologically. The oocytes were perfused continuously with frog Ringer solution at room

30
temperature (24-27 o C) and voltage-clamped at a holding potential of - 6 0 to - 7 0 mV with two glass capillary microelectrodes. Drugs were applied by including them in perfusion medium. In some experiments, the C1- concentration of the perfusate was reduced from 128 to 30 mM by replacing NaC1 with Na isethionate.
Preparation of cDNAs of the glycine receptors by site-directed mutagenesis Mutant cDNAs that were modified for a part of the clone, pGRA1, were prepared by
the PCR methods described by Ho et al. ~-~ Two primers, E1201H (5 ' -GGGAATTCG- CACCCCCATAACTCGTGGACTTTACAGCA-3 ' ) and AsTGA (for sequence, see above) were used in addition to various mutant primers as described. The mutant cDNAs were subcloned into pSPT19 and the sequences surrounding the mutated regions were confirmed. Mutant RNAs were synthesized as described above.
RESULTS
Isolation and sequence determination of cDNAs encoding a glycine receptor subunit In previous reports, we showed that functional glycine receptors could be synthesized
from two different types of mRNAs: one is a large molecule ( - 10 kb) and the other is relatively small (2-3 kb) 2'3. The adult rat spinal cord is richer in the large glycine receptor mRNA whereas the neonatal (3 days old) cord preferentially expresses the smaller RNA. When total poly-(A) + RNA from 25-day-old rat spinal cord was ultracentrifuged in a sucrose gradient solution, most of the mRNA expressing functional glycine receptor was distributed in heavy-density fractions (Frs. 4 -6 in total Frs. of 22. The 28S ribosomal RNA was found in Frs. 8-9.) This sedimentation pattern accords with that of adult glycine receptor mRNA, and differs from that of the neonatal one. Therefore, it seemed that the major type of glycine receptor mRNA present in the spinal cord of 25-day-old rats is the large transcript (the adult type). For constructing cDNA libraries, the fractionated mRNAs (Fr. 5), as well as total poly-(A) + RNAs from 25-day-old and 10-12-week-old rat spinal cords, were used as templates. The libraries constructed in bacteriophages Xgtl0 and ~,ZAPII were screened with a synthetic oligonucleotide, AsLBS (Fig. 1; for sequence, see Methods), and 4 positive clones were obtained. The clones were analyzed by DNA sequencing. One of the positive clones, p903, had a DNA insert consisting of 861 base pairs (bp) in which a sequence of 612 bp was identical to that of the 48-kDa subunit clone already reported 1°. It had an additional sequence of 248 bp at the 5'-directed region where an in-frame A TG codon was present. We presumed that this ATG is a putative in-frame initiation codon, because there is a nonsense codon, TAA ( - 1 1 1 to -109) , in the upstream region of the ATG (see Fig. 2). Another clone, p1005, overlapped with the sequence in p903 at the 5'-region and it also had an extra sequence of 1264 bp nucleotides in the 3'-region in which a putative stop codon, TGA, existed. Thus, the glycine receptor cDNA which included the full-length ORF was constructed (Fig. 1). Figure 2 shows the nucleotide and deduced amino acid sequences of the glycine receptor subunit; this consists of 184 bp of 5'-non-coding, 1350 bp of ORF and 591 bp of 3'-non-coding region. As judged from the deduced amino acid sequence and N-terminal amino acid (Ala 1 in Fig. 2) determined by Grenningloh et al. 1°, the s i s a l peptide seems to consist of 28 amino acids. Almost all of the amino acids in the signal peptide region are hydrophobic ones; however, there are 6 charged amino acids (Fig. 2). Interestingly, a sequence of G l u - 4 - A l a - A s p - A l a - a precedes the N-terminus of mature glycine receptor, which is a consensus sequence cleavable by dipeptidylaminopeptidase 16.
The nucleotide sequence of the ORF region is identical to that of the 48-kDa clone m except for one nucleotide at 1110. This region in the 48-kDa clone has an ApaI-cleaving
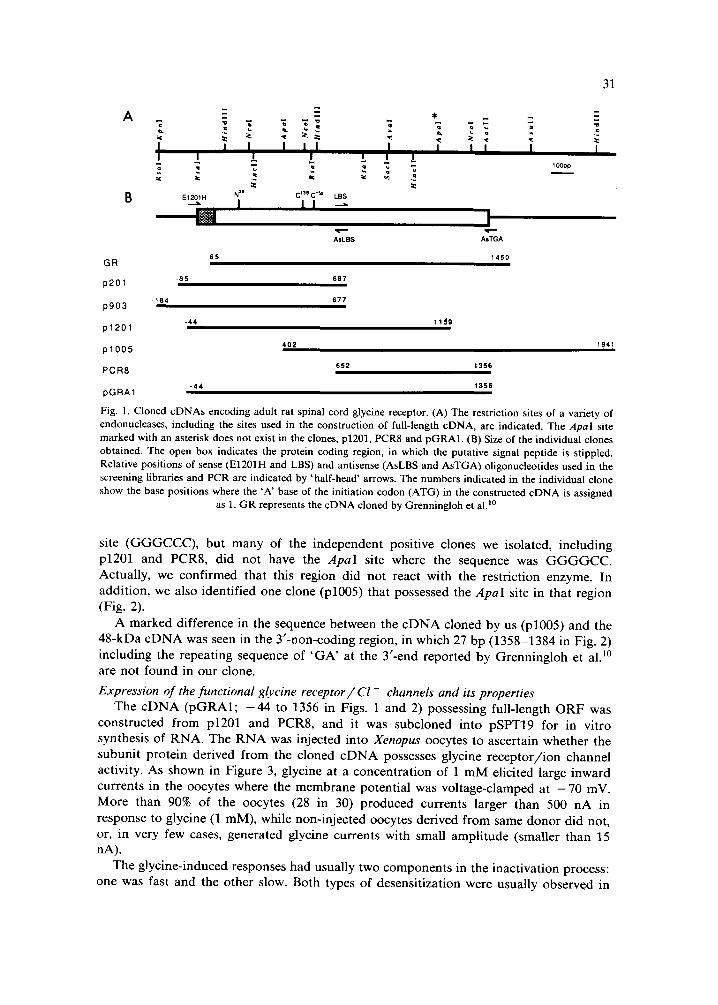
31
A
GR
p201
p903
p 1 2 0 1
p 1 0 0 5
P C R 8
= = ,
I , i , , l r i , I I I I I
E1201H N~J C1~e C~ LBS
AsLBS AsTGA
65 1450
I 1 oo~
-85 687
-184 677
-44 11~9
402 1941
652 1356
-44 1356 p G R A 1
Fig. l. Cloned cDNAs encoding adult rat spinal cord glycine receptor. (A) The restriction sites of a variety of endonucleases, including the sites used in the construction of full-length cDNA, are indicated. The Apal site marked with an asterisk does not exist in the clones, p1201, PCR8 and pGRA1. (B) Size of the individual clones obtained. The open box indicates the protein coding region, in which the putative signal peptide is stippled. Relative positions of sense (E1201H and LBS) and antisense (AsLBS and AsTGA) oligonucleotides used in the screening libraries and PCR are indicated by 'half-head' arrows. The numbers indicated in the individual clone show the base positions where the 'A' base of the initiation codon (ATG) in the constructed cDNA is assigned
as 1. GR represents the eDNA cloned by Grenningloh et al? °
site ( G G G C C C ) , bu t many of the i n d e p e n d e n t posi t ive clones we isolated, inc lud ing p1201 and PCR8, d id not have the ApaI site where the sequence was G G G G C C . Actua l ly , we conf i rmed that this region d id not react with the res t r ic t ion enzyme. In add i t ion , we also ident i f ied one c lone (p1005) that possessed the ApaI site in that region (Fig. 2).
A marked di f ference in the sequence be tween the e D N A cloned by us (p1005) and the 48 -kDa e D N A was seen in the 3 ' -non-cod ing region, in which 27 bp (1358-1384 in Fig. 2) inc lud ing the repea t ing sequence of ' G A ' at the 3 ' - end repor ted b y G r e n n i n g l o h et al. a° are no t found in our clone.
Expression of the functional glycine receptor/Cl - channels and its properties The c D N A ( p G R A 1 ; - 4 4 to 1356 in Figs. 1 and 2) possess ing ful l - length O R F was
cons t ruc ted f rom p1201 and PCR8, and it was subc loned into pSPT19 for in v i t ro synthesis of R N A . The R N A was injected into Xenopus oocytes to ascer ta in whether the subuni t p ro te in der ived f rom the c loned e D N A possesses glycine r e c e p t o r / i o n channel act ivi ty. As shown in F igure 3, glycine at a concen t ra t ion of 1 m M elici ted large i nward cur ren ts in the oocytes where the m e m b r a n e po ten t ia l was vo l t age-c lamped at - 7 0 mV. M o r e than 90% of the oocytes (28 in 30) p r o d u c e d currents larger than 500 n A in response to glycine (1 mM), while non- in jec ted oocytes der ived f rom same d o n o r d id not, or, in very few cases, genera ted glycine currents with small ampl i t ude (smal ler than 15 nA).
The g lyc ine- induced responses had usual ly two c ompone n t s in the inac t iva t ion process : one was fast and the o ther slow. Both types of desens i t iza t ion were usual ly obse rved in

32
- 1 8 4
- 1 0 5
- 2 5
+ 4 0
+ i 0 0
+ 1 8 0
+ 2 2 0
+ 2 8 0
+ 3 4 0
+ 4 0 0
+ 4 8 0
+ 5 2 0
+ 5 8 0
+ 6 4 0
+700
+760
+820
+880
+ 9 4 0
+1000
+1060
+1120
+1180
+1240
+1300
+1352
+1441
+ 1 5 2 0
+ 1 5 8 9
+1678
+1757
+1838
+1815
GCGGTACCTTAGCTGCCTGGCTGCAATTCTCCCCCCCAGCTCTGCCACCTCGTCCCTCTACTCCCTGCCCCCTTAAACA
TCTGGATTATTACCCCCAGTGGCGTTCTCCGGCTTCGTAAGTGCCCATCGGAAACATCTTTGCACCCCCATAACTCGTG
GACTTTACAGCACAAGAATCTGGAAG ATG TAC AGC TTC AAC ACT CTG CGA TTC TAC CTT TGG GAG
ACC ATT GTA TTC Thr Ile Val Phe
AAG CCT ATG TCA Lys Pro Met Set
GCC AGG ATC AGA Ala Arg Ile ArE
AAC AGC TTT GGT Asn Ser Phe Gly
CAG CAG TGG AAC Gln Gln Trp Asn
GAC CCA TCC ATG Asp Pro Set Met
GCC CAC TTC CAC Ala His Phe His
GTC CTC TAC AGC Val Leu Tyr Ser
CCG ATG GAC GTA Pro Met Asp Val
CTC ATC TTT GAG Leu Ile Phe Glu
CAG TTT ATC CTG Gln Phe Ile Leu
AAA TTC ACC TGC Lys Phe Thr Cys
CAG ATG TAC ATC Gln Met Tyr Ile
ATG GAT GCT GCA Met Asp Ala Ala
CAG AGC TCT GGC Gln Ser Ser Gly
TGG ATG GCT GTT Trp Met Ala Val
TTT GTG TCC CGG Phe Val Ser Arg
GAT GAT GAG GGT Asp Asp Glu Gly
CTG CAA GCC AAG Leu Gln Ala Lys
GCT CCT GCA CCG Ala Pro Ala Pro
AAG ATC GAC AAA Lys Ile Asp Lys
TAC TGG ATC ATC Tyr Trp Ile Ile
Met Tyr -28
TTC AGC CTT GCT GCT Phe Set Leu Ala Ala
-i0
CCC TCG GAC TTC CTG Pro Ser Asp Phe Leu i0
CCC AAC TTT AAA GGT Pro Ash Phe Lys GIy 3O
TCT ATC GCC GAG ACA Ser Ile Ala Glu Thr 5O
GAC CCC CGT CTC GCC A~ Pro Arg Leu Ala
TTG GAT TCC ATC TGG Leu Asp Ser Ile Trp 90
GAG ATC ACC ACG GAC Glu Ile Thr Thr Asp ii0
ATC AGA ATC ACC CTG lle Arg Ile Thr Leu 130
CAG ACA TGT ATC ATG Gln Thr Cys Ile Met 150
TGG CAA GAG CAA GGA Tr Gln Glu Gln GIy 17~ AAG GAA GAG AAG GAT LI~0S Olu GIu Lys Asp
ATT GAG GCC CGA TTC lle Glu Ala ArE Phe 210
CCC AGC CTG CTT ATC Pro Set Leu Leu Ile 23O
CCA GCT CGT GTG GGA Pro Ala ArE Val Gly 25O
TCC CGA GCC TCC CTA Ser Arg Ala Ser Leu 270
TGC CTG CTC TTC GTG C s Leu Leu Phe Val 2~o CAN CAC AAG GAA CTC Gin His Lys Glu Leu 310
GGA GAA GGC CGC TTC G133~ GIu GIy Arg Phe
GAT GGC ATC TCT GTC As Gly Ile Set Val as~ TCC AAG TCC CCG GAG Set Lys Ser Pro Glu 370
ATA TCT CGC ATC GGT Ile Set ArE I1e Gly 390
TAC AAG ATC GTC CGG T r Lys Ile Val ArE 4~o
Ser Phe
TCC AAA Set Lys
GAT AAG Asp Lys
CCT CCT Pro Pro
ACC ATG Thr Met
TAC AAT Tyr Asn
AAG CCT Lys Pro
AAC AAG Asn Lys
ACT CTG Thr Leu
CAA CTG Gln Leu
GCT GTG Ala Val
CTG AGA Leu Arg
CAC CTG His Leu
GTC ATC Val Ile
CTG GGC Leu G1y
CCC AAG Pro Lys
TTC TCG Phe Ser
CTT CGA Leu ArE
AAC TTC ASh Phe
HAG GGT Lys Gly
GAG ATG Glu Met
TTC CCC Phe Pro
AGA GAG Arg Olu
Asn Thr Leu
GAG GCT GAC GIu Ala Asp
CTT ATG GGA Leu Met GIy
GTG AAC GTG Val Asn Val
GAC TAC AGG Asp Tyr Arg
GAA TAT CCT Glu Tyr Pro
GAC TTG TTC Asp Leu Phe
CTG CTG AGA Leu Leu Arg
GCC TGC CCC Ala Cys Pro
GAA AGC TTT Glu Ser Phe
CAG GTG GCA Gln Val Ala
TAC TGC ACC Tyr Cys Thr
GAA CGG CAG Glu Arg Gln
CTG TCC TGG Leu Ser Trp
ATC ACC ACA Ile Thr Thr
GTG TCC TAC Val Ser Tyr
GCC CTG CTG Ala Leu Leu
TTT AGG AGG Phe Arg Arg
TCC GCC TAT Ser Ala Tyr
GCC AAC AAC Ala ASh ASh
CGG AAA CTC Arg Lys Leu
ATG GCC TTC Met Ala Phe
GAC GTC CAC Asp Val His
A r g P h e T y r L e u T r p G l u 20
GCT GCC CGC TCT GCA CCC A l a A l a A r g S e t A l a P r o
1 ~1
AGG ACT TCT GGG TAT GAT A ~ T h r S e t G l y T y r A s p
AGT TGC AAC ATC TTC ATC S e r C y s A s n I l e P h e I l e
40
GTC AAC ATC TTC CTG AGG V a l A s n I 1 e P h e L e u A r g
60
GAC GAC TCT CTG GAC CTT A ~ A s p S e r L o u A s p L o u
TTT GCC AAT GAG AAG GGG P h e A l a A s n G l u L y s G l y 100
ATC TCC CGG AAC GGC AAC lie Set Arg ASh Oly Asn 120
ATG GAC CTG AAG AAT TTC Met Asp Leu Lys Asn Phe 140
GGT TAT ACC ATG AAC GAC GII6~ Tyr Thr Met ASh Asp
GAT GGA CTG ACC CTG CCT As Gly Leu Thr Leu Pro
AAG CAC TAG AAC ACA GGT L s H i s T y r A s n T h r G l y 2~o ATG GGC TAC TAC CTG ATC Met Gly Tyr Tyr Leu Ile 220
ATC TCC TTC TGG ATC AAC Ile Ser Phe Trp Ile Asn 240
GTG CTC ACC ATG ACC ACA Val Leu Thr Met Thr Thr 250
GTG AAA GCT ATT GAC ATC Val Lys Ala Ile Asp Ile 280
GAA TAT GCC GCT GTC AAC Glu Tyr Ala Ala Val Asn 3OO
AAG CGG CGA CAT CAC HAG L3~0S Arg Arg His His Lys
GGG ATG GGG CCA GCC TGC G134~ Met Gly Pro Ala Cys
AAC AAC ACC ACG AAC CCC Asn ASh Thr Thr ASh Pro 350
TTC ATC CAG AGA OCC AAG Phe Ile Gln ArE A1a Lys 380
CTC ATC TTC AAC ATG TTC Leu Ile Phe Asn Met Phe 4O0
AAC AAG TGA AGGCTCTGGGA Asn Lys End 420
CGGGGT GGGGGGGAGCTG6GAGAGGGTGC~TGGGAT GAGCACAGAGGC T ({GGAGCCAAGGGAGGAGGAAGAGA~AAGAG
C A ~ A ~ A ~ A T ~ T C C C T ~ C A ~ C A A G T ~ A A T A ~ A A T C C A C A A ~ G ~ T ~ A T A C A T C ~ A ~ G T G T T A C A A T ~ T ~ A T C T A G
CCTCCAGGGGCGTGAGGTGGGAGGGCCTTTTCGAATACAGCAACTGGGGACCATGAGGGGTGGGACAGGGGTCAGGGAG
GGGATTGGGAA~%TCACTGCTTTCAAAATTC~AGTCCATTGGGTTGAGGGGGGC~GGGGGGcTCCTGTTTGGAGGTCAA TGCTGCCTGCCCCAAAAACAGGAAGGCAGTGT~LATATATATATAACATTTAACCATG~TTAACATTAATCGAAAACCCA
ccAGAAcAAAAGATACGTCTCTT~EACCAGCCTAGTAACTGCTTAAACTTTTAAAGTCCCCAA~GTGATGQACAATACT TCCCAGAGTAGAAATGGACACGAATAAGCTTGCTGTGGTCTATGCTCCCATCACGCTC TTACAA~CGCCATGT C~GTCT
TTAAACTGACCCCCGTGGACACAT GCC
Fig. 2. Nucleotide and deduced amino acid sequences of rat glycine receptor subumt. Numbers on the left and beneath the am/no acid denote nucleotide and an-nno acid positions respectively. Assignment of amino acids are made from Ala, the putative N-terminal amino acid of matured ~ycine receptor TM. An upstream nonsense codon is underlined. The nucleotide, G, marked with an asterisk is found in p1201, PCR8 and pGRA], but not in
p1005 clone, in which the base is replaced by C.

33
1 ~ glycine A i - -
B
1 iN olycine
7 f
Strychnine
Gly a-Ala Gly Gly
Fig. 3. Glycine- and fl-alanine-induced currents recorded from Xenopus oocytes injected with a single RNA (25 ng/50 nl) transcribed from pGRA1. Oocytes were voltage-clamped at -70 mV, and drugs were applied by perfusion during periods indicated by horizontal lines. Downward deflection denotes inward current. (A) Two characteristic examples of glycine currents in oocytes derived from a single donor. (B) The expressed glycine receptor responded not only to glycine but to fl-alanine (1 mM) to generate the inward currents, and the
glycine response was markedly reduced by strychnine (200 nM).
the same oocyte, but the extent contributed by each type differed from oocyte to oocyte; in some oocytes the slow phase was preferentially seen (Fig. 3A-i) and in others the fast phase greatly contributed to the inactivation (Fig. 3A-ii). Both types of the response were markedly reduced in the presence of 200 nM strychnine, a potent glycine receptor antagonist, and were immediately recovered after washing out the antagonist (Fig. 3B). By contrast, bicuculline, a GABA A receptor antagonist, did not affect the glycine-induced response even at a concentration of 50/~M (data not shown).
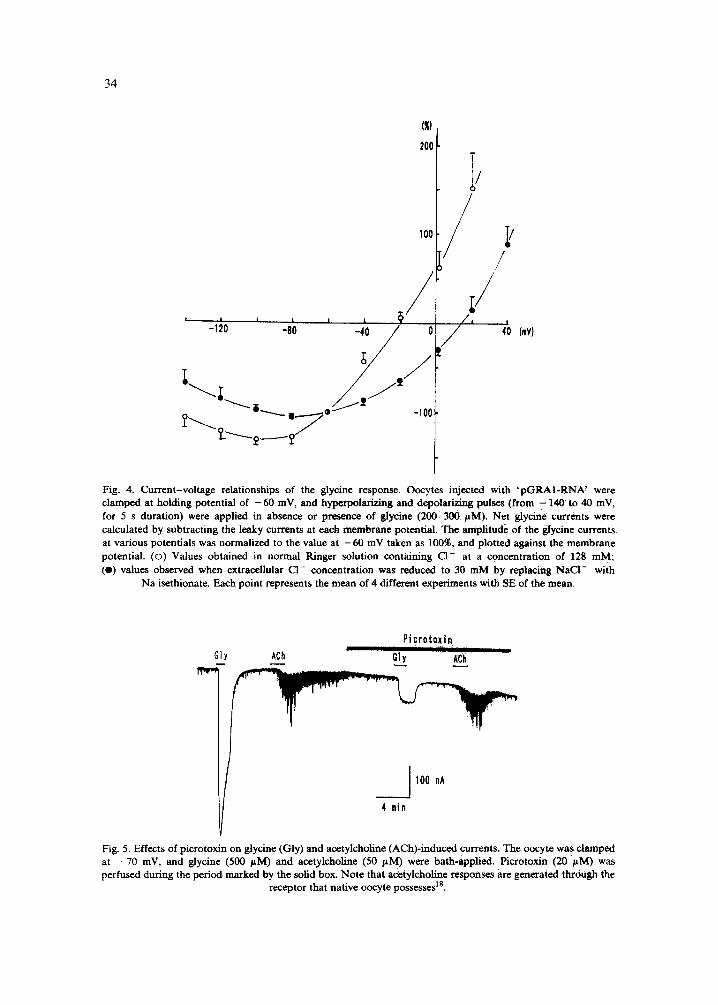
The glycine-induced currents inverted direction from inward to outward at about - 2 0 mV, which was close to equilibrium potential of C1- in oocytes 21 (Fig. 4). Furthermore, the reversal potential shifted in the positive direction when the extracellular C1- con- centration was reduced to 30 mM by substituting Na isethionate for NaC1 (Fig. 4). Therefore, the main carrier of the glycine currents would be chloride ions.
The C1- currents generated by glycine were depressed by picrotoxin, a blocker of certain types of C1- channels (Fig. 5). It has been demonstrated that picrotoxin can block not only GABA currents but also glycine currents in dissociated mammalian neurons 4 and in oocytes injected with rat spinal cord mRNAs 2, indicating that this effect is not the result of a 'cloning artifact'. It is known that in Xenopus oocyte, acetylcholine, serotonin, glutamate and some peptide transmitters/hormones produce C1- currents accompanied by membrane current oscillations tl't2"a4.18.2°,22.34.39. These responses are thought to be generated through intrinsic C1- channels which native oocytes possess 18'21'23"24"36. The oscillatory response induced by acetylcholine was not blocked by picrotoxin (Fig. 5),
34
(~),
200 l
100
/
/J /J j
/ 4o (mY)
Fig. 4. Current-voltage relationships of the glycine response. Oocytes injected with 'pGRA1-RNA' were clamped at holding potential of - 60 mV, and hyperpolarizing and depolarizing pulses (from - 140 to 40 mV, for 5 s duration) were applied in absence or presence of glyeine (200-300 /~M). Net glyeine currents were calculated by subtracting the leaky currents at each membrane potential. The amplitude of the glycine currents at various potentials was normalized to the value at - 6 0 mV taken as 100%, and plotted against the membrane potential. (o) Values obtained in normal Ringer solution containing C1- at a concentration of 128 mM; (e) values observed when extracellular CI- concentration was reduced to 30 mM by replacing NaCt- with
Na isethionate. Each point represents the mean of 4 different experiments with SE of the mean.
Gly
Picrotoxin I I IIII h i ( [ I I _ _
ACh Gly kCh
~ J 100 n^
4 rain
Fig. 5. Effects of picrotoxan on glycine (Gly) and acetylcholine (ACh)-induced currents. The oocyte was clamped at - 7 0 mV, and glycine (500 /zM) and acetylcholine (50 /~M) were bath-applied~ Picrotoxin (20 /~M) was perfused during the period marked by the solid box. Note that acetylcholine responses are generated through the
receptor that native oocyte possesses TM.

Relative amplitude of response 0 50 100 (~} f i I
Glycine ~ ~ ~ #-A]anine ~ a-Alanine
Taurine
Serine
GABA
35
Fig. 6. Comparison of potencies of glycine and other related compounds for generating currents. Oocytes injected with pGRAI RNA were clamped at -70 mV and test compounds at concentration of 1 mM were sequentially applied to a single oocyte at an interval of 5 rain. To avoid cross-desensitization effects, the order of application of chemicals was usually fixed from bottom (GABA) to top (glycine), but applications in reverse order did not change the magnitude of the responses. Each column represents the relative amplitudes
(mean + SEM, n = 6) where the peak amplitude of glycine currents was taken as 100%.
suggesting that C1- channels gated by glycine are newly synthesized and result from the injection of the RNA.
Agonist specificity in activating the cloned glycine receptor was examined in oocytes injected with synthetic RNA. fl-Alanine, a closely related amino acid to glycine, also produced C1- currents (Figs. 3 and 6) and the response was blocked by strychnine and picrotoxin (data not shown). The potency of fl-alanine as a glycine receptor agonist was usually about one-third that of glycine (29.2 _ 0.9%, n = 6) when compared at the same concentration (1 raM), but in some oocytes the peak amplitude of the ~-alanine response was greater than half of the glycine response (e.g., see Fig. 3). Other related amino acids, taurine, a-alanine and serine act as very weak agonists, while GABA did not elicit any currents (Fig. 6). Other compounds such as cysteamine, malonic acid, glycineamide, aminomethane sulfonate and a dipeptide, glycyl-glycine, were essentially ineffective.
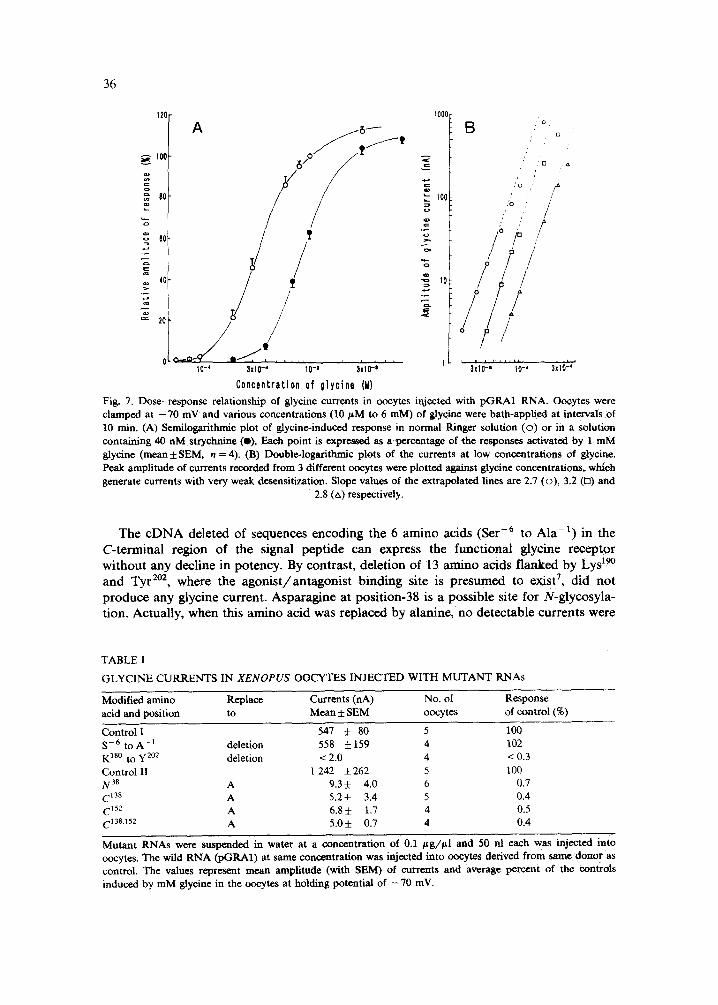
Figure 7 shows the concentration-response relationships on glycine currents. The minimum concentration for detecting glycine currents (1 nA) was 20-40 /~M and the responses reached their maximum at concentrations of 1-3 raM. Double logarithmic plots at low glycine concentrations gave slopes of 2.7-3.2 (Fig. 7B). These values were similar to those in the case observed in oocytes injected with poly(A) ÷ mRNAs isolated from various animal sources 2"13'22 and in cultured mouse spinal neurons 5, suggesting that 3 glycine molecules are required for opening of the CI - channels. Strychnine at a low concentration (40 nM, the estimated IC5o value) blocked the glycine response in an apparently competitive manner (Fig. 7A). In the presence of a higher concentration of strychnine (200 nM), glycine (up to 100 mM)-induced responses did not reach maximal values.
Modification of glycine receptor subunit by site-directed mutagenesis and properties of the modified receptors
From the deduced amino acid sequence of the glycine receptor subunit, its membrane topology was inferred (for model, see Ref. 7). The next important step is to determine which part of the protein is associated with which functional property. To solve this question, modification of the cDNA by site-directed mutagenesis was attempted. Some of the results are summarized in Table I.
36
120
10C
80
eO
o~ 40 >
~: 20
1000
A ~ s f
lo0
10 -4 3X10 -4 10-a 3X10 -z
Concent ra t ion of 91ycine (M)
/ • D
/ ,'D 4
/O " ," r' /
,'O ,"
," ,/ /
3x10 ~G I0 ~4 3x10"
Fig. 7. Dose-response relationship of glycine currents in oocytes injected with pGKA1 RNA. Oocytes were clamped at - 7 0 mV and various concentrations (10/xM to 6 raM) of glycine were bath-applied at intervals of 10 rain. (A) Sernilogarithmie plot of glycine-induced response in normal Ringer solution (o ) or in a solution containing 40 nM strychnine (O). Each point is expressed as a percentage of the responses activated by 1 mM glycine (mean ± SEM, n = 4). (B) Double-logarithmic plots of the currents at low concentrations of glycine. Peak amplitude of currents recorded from 3 different ooeytes were plotted against glyeine concentrations, which generate currents with very weak desensitization. Slope values of the extrapolated lines are 2.7 (o) , 3.2 (n) and
2.8 (zx) respectively.
The cDNA deleted of sequences encoding the 6 amino acids (Ser -6 to Ala -1) in the C-terminal region of the signal peptide can express the functional glycine receptor without any decline in potency. By contrast, deletion of 13 amino acids flanked by Lys 19° and Tyr 2°2, where the agonist/antagonist binding site is presumed to exist 7, did not produce any glycine current. Asparagine at position-38 is a possible site for N-glycosyla- tion. Actually, when this amino acid was replaced by alanine, no detectable currents were
TABLE I
GLYCINE CURRENTS IN XENOPUS OOCYTES INJECTED WITH MUTANT RNAs
Modified amino Replace Currents (hA) No. of Response acid and position to Mean ± SEM oocytes of control (%)
Control I 547 ± 80 5 100 S -6 to A -1 deletion 558 +159 4 102 K 18° to y202 deletion < 2.0 4 < 0.3 Control II 1 242 +262 5 100 N 38 A 9.3+ 4.0 6 0.7 C 138 A 5.2+ 3.4 5 0.4 C 152 A 6.8± 1.7 4 0.5 C 13sa~2 A 5.0+ 0.7 4 0.4
Mutant RNAs were suspended in water at a concentration of 0.1 # g / # l and 50 nl each was injected into oocytes. The wild RNA (pGRA1) at same concentration was injected into oocytes derived from same donor as control. The values represent mean amplitude (with SEM) of currents and average percent of the controls induced by mM glycine in the oocytes at holding potential of - 7 0 mV.

37
observed. The mutant receptors with either or both cysteines at positions 138 and 152 substituted by alanine also resulted in very low expression.
DISCUSSION
In the present study, the sequence of glycine receptor cDNAs including the full-length protein coding region of a receptor subunit protein was determined. When compared with the nucleotide sequence of the 48-kDa subunit cDNA reported by Grenningloh et al. 1°, the sequence of the coding region in many clones that we isolated fully match with each other, except that at one base, at 1110, C is substituted for G. However, we obtained an additional clone in which the ApaI-cleaving site does exist, supposing that the single-base substitution occurred naturally on this point. Another difference was found in the 3'-trailing sequence where 27 bases (1358-1384) were substituted for those in the 48-kDa subunit cDNA. We neither isolate any cDNA identical to that of 48-kDa subunit from our cDNA libraries nor succeed in amplification of the 48-kDa cDNA by PCR using primers corresponding to these sequences, suggesting that the mRNA of the glycine receptor does not possess this sequence, or, if present, is in very small proportions. This mismatch may result from sequence polymorphisms between different strains of rats (Sprague-Dawley vs. Wistar), or from alternative splicing events.
The size of the ORF was determined to be 1.35 kb. This value is much smaller than the mRNA which is - 10 kb in size as determined by both Northern blot hybridization 3'1° and combined with analysis using sucrose density gradient and functional assays 2'3. This suggests that the mRNA possesses an extremely long 5'- a n d / o r 3'-non-coding region. Indeed, there is no consensus sequence for polyadenylation signal (AATAAA) in the 3'-non-coding region within the 591 bp that has been determined.
The sequence of the 5'-upstream region including the signal peptide domain was determined in the present study. It is reported that the N-terminal amino acid of the mature subunit protein is Ala 1 (Fig. 2, see Ref. 10) and therefore, we assumed that the signal peptide consists of 28 amino acids; the size of the signal peptide is in good agreement with that of other membrane proteins. In the signal sequence, however, there are 6 charged amino acids present; in particular, a sequence of G l u - a - A l a - A s p - A l a -1 (a consensus sequence cleavable by dipeptidylaminopeptidase 16) precedes the N-terminal amino acid, Ala 1, identified by the protein sequencing 1°. This is very rare because the signal peptide usually consists of hydrophobic amino acids 38. It is possible that the receptor subunit is processed by two (e.g. a signal peptidase and a dipeptidyl amino- peptidase 16) or more enzymes.
A single RNA transcribed from the cloned cDNA synthesized functional glycine receptor in Xenopus oocytes. This result agrees with that observed using a chimeric clone constructed with the incomplete 48-kDa subunit cDNA and with a sequence encoding the signal peptide of a GABA A receptor subunit 29'33. It has been reported that injection of single RNA encoding the a- or fl-subunit of GABA A receptors induces functional GABA-gated C1- channels 8'32. However, its relative amplitude is very small, whereas co-injection of a- and fl-subunit RNAs induces larger GABA currents 27"3°. A similar case has been reported for the nicotinic acetylcholine receptor; it requires hetero-oligomeric assembly consisting of a-, fl-, and 7- or 8-subunits for functional receptors ~7. These contrast with what we and others observed for the glycine receptors. A single subunit RNA alone induces the formation of glycine receptors in oocytes which produce C1- currents of very large amplitude. Moreover, the expressed receptors possess properties essentially indistinguishable from those of glycine receptors observed in mammalian

38
neurons 4 and in oocytes injected with total poly-(A) ~ RNA 1'2, with respect to agonist and antagonist activities and to Cl- channel function. Presumably, single receptor subunits assemble as a tetra- or pentameric oligomer to form Cl channels 19"3~'3v.
During sustained application of glycine, the evoked response cannot hold at peak current and begins to decay. This desensitization phase of glycine receptors expressed by pGRA1-RNA consists of two components- - fas t and slow in time course--al though the extent of the two phases varies in individual oocytes (see Fig. 3). Similar desensitization patterns in glycine currents have been reported for mammalian neurons recorded by the whole cell clamp technique 4. In this case, it was proposed that functionally different glycine receptors give rise to different C1- currents. In the present experiments, however. the expressed glycine receptors, which appear to be homomeric because they are synthe- sized from a single subunit mRNA, possess two different desensitization kinetics. Many possibilities can account for this mechanism; for instance, (1) multiple post-translational mechanisms take part in the receptor subunit processing, (2) different extent of phos- phorylation on individual receptor subunits, and (3) altered assembly in terms of tertiary a n d / o r quaternary structures. Further experiments are required to solve this question.
As mentioned above, the glycine receptor expressed in oocytes seems to be a homo- oligomeric one, which possesses a fully functional Cl- channel. This result contrasts with reports that functional glycine receptors in neurons are composed of heterogeneous subunits 7. So far, we do not know whether the homo-oligomeric receptors are functional in the mammalian central nervous system. Apart from this, however, it is without doubt that the Xenopus oocytes system combined with a single subunit mRNA encoding the glycine receptor provides a great advantage in defining the essential sites of the receptor necessary for function as ion channels. Preliminary experiments to determine this was performed using site-specific mutagenesis. Modification of Asn 38 to Ala, which is a putative N-glycosylation site, resulted in failure of receptor expression. It is reported that in the nicotinic acetylcholine receptor, the disulfide bond of two cysteine residues is important for function. In the case of the glycine receptor, it is also important, because the response was markedly reduced when the Cys at position-138 or -152 was substituted by Ala. Further experiments are required to clarify whether the lack of glycine receptor function in oocytes injected with many mutant RNAs is due to instability of the synthesized subunits, to failure in membrane inversion, or to synthesis of stable protein without function.
ACKNOWLEDGEMENTS
We would like to thank Ms. K. Akita and F. Ozawa for their technical assistance in the preparation of mRNAs and synthesis of oligonucleotides, and Dr. Y. Kikuchi for his kind advice on the PCR method. We are also grateful to Drs. K. Nagahari and K. Takahashi of Mitsubishi Kasei Co. Ltd. for helpful discussions during the cloning work, and to Dr. Y. Kudo for his encouragement throughout this work. Part of this work was supported by research funds from the Ichiro Kanehara Foundation and the Naito Foundation.
NOTE ADDED IN PROOF
The nucleotide sequence data for an adult rat glycine receptor ~xl reported in this paper will appear in the DDBJ, EMBL and GenBank Nucleotide Sequence Databases under the accession number D00833.

39
REFERENCES
1 Akagi, H. and Miledi, R., Expression of glycine and other amino acid receptors by rat spinal cord mRNA in Xenopus oocytes, Neurosci. Lett., 95 (1988), 262-268.
2 Akagi, H. and Miledi, R., Heterogeneity of glycine receptors and their messenger RNAs in rat brain and spinal cord, Science, 242 (1988) 270-273.
3 Akagi, H., Patton, D.E. and Miledi, R., Discrimination of heterogenous mRNAs encoding strychnine-sensi- tive glycine receptors in Xenopus oocytes by antisense oligonucleotides, Proc. Natl. Acad. Sci. USA, 86 (1989) 8103-8107.
4 Akaike, N. and Kaneda, M., Glycine-gated chloride current in acutely isolated rat hypothalamic neurons, J. Neurophysiol., 62 (1989) 1400-1409.
5 Barker, J.L., Macdonald, R.L. and Smith, T.G., Voltage-clamp analysis of amino acid currents in cultured mammalian neurons, J. Gen. Physiol., 70 (1977) la.
6 Barker, J.L., GABA and glycine: ion channel mechanism. In M.A. Rogawski and J.L. Barker (Eds.), Neurotransmitter Actions in the Vertebrate Nervous System, Plenum, New York, 1985, pp. 71-100.
7 Betz, H., Schmitt, B., Becker, C.-M., Grenningloh, G. and Rienitz, A., The glycine receptor. In A.D. Strosberg, (Ed.), The Molecular Biology of Receptors, Eliss, Chichester, 1987, pp. 128-138.
8 Blair, L.A., Levitan, E., Marshall, J., Dionne, V.E. and Barnard, E.A., Single subunits of the GABA A receptor form ion channels with properties of the native receptor, Science, 242 (1988) 577-579.
9 Dani, J.A., Site-directed mutagenesis and single-channel currents define the ionic channel of the nicotinic acetylcholine receptor, Trends Neurosci., 12 (1989) 125-128.
10 Grenningloh, G., Rienitz, A., Schmitt, B., Methfessel, C., Zensen, M., Beyreutther, K., Gundelfinger, E.D. and Betz, H., The strychnine-binding subunit of the glycine receptor shows homology with nicotinic acetylcholine receptors, Nature, 328 (1987) 215-220.
11 Gundersen, C.B., Miledi, R. and Parker, I., Serotonin receptors induced by exogenous messenger RNA in Xenopus oocytes, Proc. R. Soc. Lond., B219 (1983) 103-109.
12 Gundersen, C.B., Miledi, R. and Parker, I., Glutamate and kainate receptors induced by rat brain messenger RNA in Xenopus oocytes, Proc. R. Soc. Lond., B221 (1984) 127-143.
13 Gundersen, C.B., Miledi, R. and Parker, I., Properties of human glycine receptors expressed in Xenopus oocytes, Proc. R. Soc. Lond., B221 (1984) 235-244.
14 Hirono, C., Ito, I. and Sugiyama, H., Neurotensin and acetylcholine evoke common responses in frog oocytes injected with rat brain messenger ribonucleic acid. J. Physiol. (Lond.), 382 (1987) 523-535.
15 Ho, S.N., Hunt, H.D., Horton, R.M., Pullen, J.K., and Pease, L.R., Site-directed mutagenesis by overlap extension using the polymerase chain reaction, Gene, 77 (1989) 51-59.
16 Kreil, G., Processing of precursors by dipeptidylaminopeptidases: a case of molecular ticketing. Trends Biochem. Sci., 15 (1990) 23-26.
17 Kurosaki, T., Fukuda, K., Konno, T., Mori, Y., Tanaka, K., Mishina, M. and Numa, S., Functional properties of nicotinic acetylcholine receptor subunits expressed in various combinations, FEBS Lett., 214 (1987) 253-258.
18 Kusano, K., Miledi, R. and Stinnakre, J., Cholinergic and catecholaminergic receptors in the Xenopus oocyte membrane, J. Physiol. (Lond), 328 (1982) 143-170.
19 Langosch, D., Thomas, L. and Betz, H., Conserved quaternary structure of ligand-gated ion channels: the post synaptic glycine receptor is a pentamer, Proc. Natl. Acad. ScL USA, 85 (1988) 7394-7398.
20 Meyerhof, W., Morley, S., Schwarz, J. and Richter, D., Receptors for neuropeptides are induced by exogenous poly-(A) + RNA in oocytes from Xenopus laevis, Proc. Natl. Acad Sci. USA, 85 (1988) 714-717.
21 Miledi, R., Parker, I. and Sumikawa, K., Transplanting receptors from brains into oocytes. In Fidia Research Foundation Neuroscience Award Lectures, 1Iol. 3., Raven, New York, 1989, pp. 57-90.
22 Nomura, Y., Kaneko, S., Kato, K., Yamagishi, S. and Sugiyama, H., lnositol phosphate formation and chloride current responses induced by acetylcholine and serotonin through GTP-binding proteins in Xenopus oocytes after injection of rat brain messenger RNA, Mol. Brain Res., 2 (1987) 113-123.
23 Oron, Y., Dascal, N., Nadler, E. and Lupu, M., Inositol 1,4,5-triphosphate mimics muscarinic response in Xenopus oocytes. Nature, 313 (1985) 141-143.
24 Parker, I. and Miledi, R., Changes in intracellular calcium and in membrane currents evoked by injection of inositol triphosphate into Xenopus oocytes, Proc. R. Soc. Lond., B228 (1986) 307-315.
25 Parker, I, Sumikawa, K. and Miledi, R., Neurotensin and substance P receptors expressed in Xenopus oocytes by messenger RNA from rat brain. Proc. R. Soc. Lond., B229 (1986) 151-159.
26 Parker, I., Sumikawa, K. and Miledi, R., Responses to GABA, glycine and fl-alanine induced in Xenopus oocytes by messenger RNA from chick and rat brain, Proc. R. Soc. Lond, B233 (1988) 201-216.

40
27 Pritchett, D.B., Sontheimer, H., Shivers, B.D., Ymer, S., Kettenman, H., Schofield, P. and Seeburg, P.H., Importance of a novel GABA A receptor subunit for benzodiazepine pharmacology. Nature. 338 (1989) 582-585.
28 Sanger, F., Nicklen, S. and Coulson, A.R., DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sei. USA, 74 (1977) 5463-5467.
29 Schmieden, V., Grenningloh, G., Schofield, P.R. and Betz, H., Functional expression in Xenopus oocytes of the strychnine binding 48kd subunit of the glycine receptor, EMBO J., 8 (1989) 695-700.
30 Schofield, P.R., Darlison, M.G., Fujita, N., Burr, D.R., Stephenson, F.A., Rodriguez, H., Rhee, L.M., Ramachandran, J., Reale, V., Glencorse, T.A., Seeburg, P.H. and Barnard, E.A., Sequence and functional expression of the GABA A receptor shows a ligand-gated receptor super-family, Nature 328 (1987) 221-227.
31 Schofield, P.R. and Abbott, A., Molecular pharmacology and drug action: structural information casts light on ligand binding, Trends Pharmacol. Sci., 10 (1989) 207-212.
32 Shivers, B.D., Killisch, I., Sprengel, R., Sontheimer, H., Kohler, M., Schofield, P. and Seeburg, P.H., Two novel GABA A receptor subunits exist in distinct neuronal subpopulations, Neuron, 3 (1989) 327-337.
33 Sontheimer, H., Becket, C.-M., Pritchett, D.B., Schofield, P.R., Grenningloh, G., Kettenmann, H., Betz, H. and Seeburg, P.H., Functional chloride channels by mammalian cell expression of rat glycine receptor subunit, Neuron, 2 (1989) 1491-1497.
34 Sugiyama, H., Ito, I and Hirono, C., A new type of glutamate receptor linked to inositol phospholipid metabolism, Nature, 325 (1987) 531-533.
35 Sumikawa, K., Parker, I. and Miledi, R., Expression of neurotransmitter receptors and voltage-activated channels from brain mRNA in Xenopus oocytes, Meth. Neurosci., ] (1989) 30-45.
36 Takahashi, T., Neher, E. and Sakmann, B., Rat brain serotonin receptors in Xenopus oocytes are coupled by intracellular calcium to endogenous channels, Proc. Natl. Acad. Sci. USA, 84 (1987) 5063-5067.
37 Unwin, N., The structure of ion channels in membrane of excitable cells, Neuron, 3 (1989) 665-676. 38 Verner, K. and Schatz, G.. Protein translocation across membrane, Science, 241 (1988) 1307-1313. 39 Woodward, R.M. and Miledi, R., Membrane currents elicited by porcine vasoactive intestinal peptide (VIP)
in follicle-enclosed Xenopus oocytes, Proc. R. Soc. Lond., B231, (1987) 489-497. 40 Young, A.B. and Macdonald, R.L., Glycine as a spinal cord neurotransmitter. In R.A. Davidoff (Ed.),
Handbook of The Spinal Cord, Vol. 1, Marcel Dekker, New York, 1983, pp. 1-43.