free radical theory of aging: beneficial effect of adding antioxidants to the maternal mouse diet on...
TRANSCRIPT

Age, Vol. 2, 109.122, October, 1979
FREE RADICAL THEORY OF AGING: BENEFICIAL EFFECT OF ADDING ANTIOXIDANTS TO THE MATERNAL MOUSE DIET ON LIFE SPAN OF OFFSPRING; POSSIBLE EXPLANATION OF THE SEX DIFFERENCE IN LONGEVITY
Denham Harman, M.D., Ph.D.,* and Dennis E. Eddy, Ph.D. + Departments of Medicine and Biochemistry University of Nebraska College of Medicine Omaha, Nebraska 68105
Abstract
Endogenous free radical reactions may have an adverse effect on the life span. Based on this pos- sibility a number of free radical reaction inhibitors have been shown to increase the average life span of mice when added to the diet throughout life starting shortly after weaning.
The purpose of this study was to determine if exposure to free radical reaction inhibitors, by including them in the maternal diet during the period of about 40 days of high mitotic and meta- bolic activity between conception and weaning, would also increase the life span. This was found to be the case. The largest increase in average life span was obtained by adding 0.5 percent by weight of the water soluble compound, 2-mercap- toethylamine, to the maternal diet; the increase was 14.7 percent for male offspring and 8.6 per- cent for females.
Consideration of the data of this study in the light of early events in embryogenesis, led to the suggestion that the greater longevity of females is due, at least in part, to greater protection of fe- male embryos from free radical reaction damage during the period before random inactivation of one of their two X-chromosomes.
Introduction
Free radical reactions, initiated by both enzymatic and non-enzymatic means, go on continuously throughout the cells and tissues (1). It would be ex- pected because of the high chemical reactivity of the free radicals, intermediates formed in free rad- ical reactions, that all components of the body would be subject to some degree of chemical change in a more-or-less random manner. Based on the possibility (2-4) that these free radical in- duced changes have an adverse effect on the life span, a number of free radical reaction inhibitors have been shown to increase the average life span when added to the diet of mice throughout life starting shortly after weaning (5-10).
*To whom all correspondence should be addressed. + Present address: The Quaker Oats Company,
John Stuart Laboratories, 617 West Main Street, Barrington, III. 60010
Accumulation of random damage from free rad- ical reactions in the early period of life may also be sufficient to shorten the life span, owing to the high mitotic and metabolic rates (11, 12) char- acteristic of the first stages of life coupled with the probability that adverse changes may be passed on to daughter cells. In accord with this are radiation studies which show that the early stages of life are relatively easily damaged (13, 14) by free radical reactions initiated in utero by a brief exposure to ionizing radiation; administra- tion of 2-mercaptoethylamine (2-MEA) to the mother before i[radiation (15) decreases such damage. Thus, exposure to free radical reaction inhibitors during the relatively short time, from conception to weaning, may have a beneficial ef- fect on longevity. To evaluate this possibility, two experiments were conducted in which groups of female mice were maintained on a semi-synthetic diet, with and without an added free radical reac- tion inhibitor, from one month before mating until their offspring were weaned. The male and female offspring were separated, placed on a pelleted commercial diet, and survival curves obtained.
Results
Experiment 1
All the female Swiss mice apparently became pregnant; it cannot be stated with certainty that all became pregnant as there were 5 mice per cage. Whether or not the antioxidants influenced the number of offspring per mouse could not be determined because many were cannibalized. The number of offspring, percent surviving, and aver- age weight is given in Table 1 every fourth month, from age 6 months through 34 months for each group: control, 0.1%w (percent by weight) and 0.2%w 1,2-dihydro-6-ethoxy-2, 2, 4-trimethyl- quinoline (Santoquin), 0.5%w and 1.0%w 2-mer- captoethylamine hydrochloride (2-MEA), 1.0%w sodium hypophosphite (NaH2PO2)(NaHP), 0.1%w and 0.4%w butylated hydroxytoluene (BHT) and 0.1%w and 0.2%w butylated hydroxyanisole (BHA). The average life span __. standard devia- tion, the maximum life span, the difference in months and in percent between the controls and antioxidant groups, as well as between the cor- responding male and female groups, are presented in Table 2. Figure 1 shows the percent of male offspring surviving as a function of age in months for the control and three antioxidant groups, 0.2%w Santoquin, 0.5%w 2-MEA and 1.0%w NaHP; corresponding female data are pre- sented in Figure 2. The average weights of the four male groups are shown in Figure 3 and those for females in Figure 4.
109
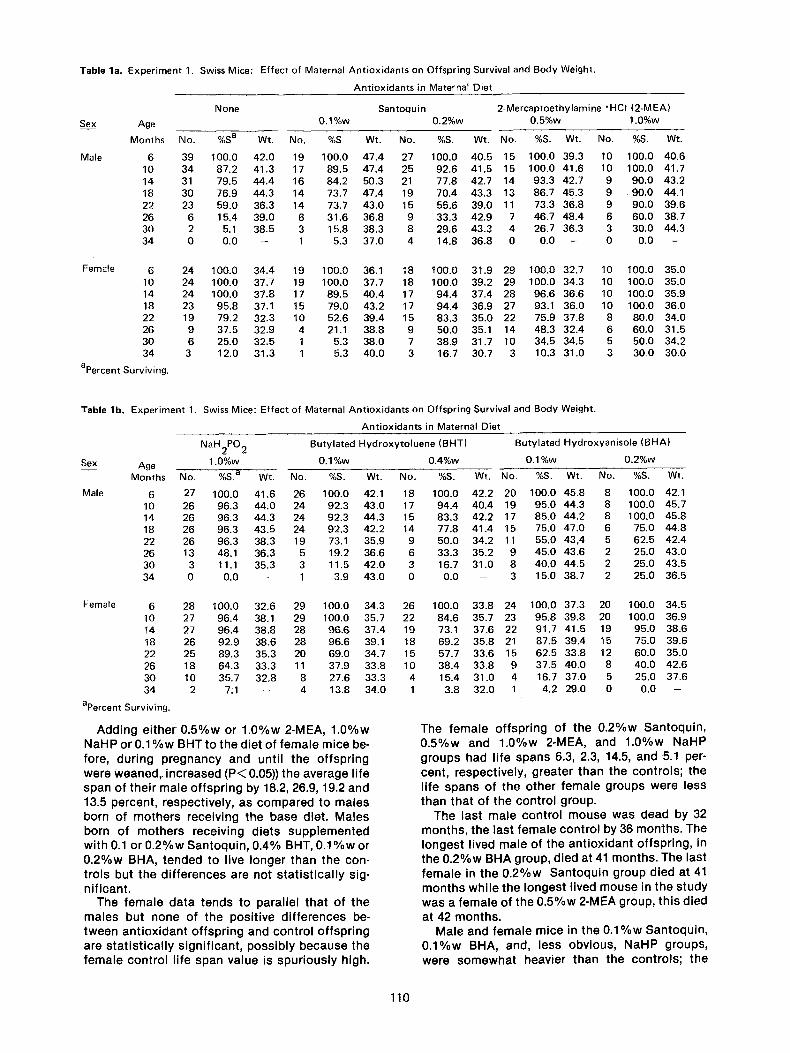
Table la. Experiment 1. Swiss Mice: Effect of Maternal Antioxidants on Offspring Survival and Body Weight.
Antioxidants in Maternal Diet
Santoquin 2-Mercaptoethylamine "HCI (2-MEA) Sex Age 0.1%w 0.2%w 0.5%w 1.0%w
Months No. %S a Wt. No. %S Wt. No. %S. Wt. No. %S. Wt. No. %S. Wt.
Male 6 39 100.0 42.0 19 100.0 47.4 27 100.0 40.5 15 100.0 39.3 10 100.0 40.6 10 34 87.2 41.3 17 89.5 47.4 25 92.6 41.5 15 100.0 41.6 10 100.0 41.7 14 31 79.5 44.4 16 84.2 50.3 21 77.8 42.7 14 93.3 42.7 9 90.0 43.2 18 30 76.9 44.3 14 73.7 47.4 19 70.4 43.3 13 86.7 45.3 9 90.0 44.1 22 23 59.0 36.3 14 73.7 43.0 15 55.6 39.0 11 73.3 36.8 9 90.0 39.6 26 6 15.4 39.0 6 31.6 36.8 9 33.3 42.9 7 46.7 48.4 6 60.0 38.7 30 2 5.1 38.5 3 15.8 38.3 8 29.6 43.3 4 26.7 36.3 3 30.0 44.3 34 0 0.0 - 1 5.3 37.0 4 14.8 36.8 0 0.0 -- 0 0.0 --
None
Female 6 24 100.0 34.4 19 100.0 36.1 18 t00.0 31.9 29 100.0 32.7 10 100.0 35.0 10 24 100.0 37.7 19 100.0 37.7 18 100.0 39.2 29 100.0 34.3 10 100.0 35.0 14 24 100.0 37.8 17 89.5 40.4 17 94.4 37.4 28 96.6 36.6 10 100.0 35.9 18 23 95.8 37.1 15 79.0 43.2 17 94.4 36.9 27 93.1 36.0 10 100.0 36.0 22 19 79.2 32.3 10 52.6 39.4 15 83.3 35.0 22 75.9 37.8 8 80.0 34.0 26 9 37.5 32.9 4 21.1 38.8 9 50.0 35.1 14 48.3 32.4 6 60.0 31.5 30 6 25.0 32.5 1 5.3 38.0 7 38.9 31.7 10 34.5 34.5 5 50.0 34.2 34 3 12.0 31.3 1 5.3 40.0 3 16.7 30.7 3 10.3 31.0 3 30.0 30.0
apercent Surviving.
Table lb. Experiment 1. Swiss Mice: Effect of Maternal Antioxidants on Offspring Survival and Body Weight.
Antioxidants in Maternal Diet
Sex
Male
NaH2PO 2 Butylated Hydroxytoluene (BHT) Butylated Hydroxyanisole (BHA)
Age 1.0%w 0.1%w 0.4%w 0.1%w 0.2%w
Months No. %S. a W~. No. %S. Wt. No. %S. Wt. No. %S. Wt. No. %S. Wt.
6 27 100.0 41.6 26 100.0 42.1 18 100.0 42.2 20 100.0 45.8 8 100.0 42.1 10 26 96.3 44.0 24 92.3 43.0 17 94.4 40.4 19 95.0 44.3 8 100.0 45.7 14 26 96.3 44.3 24 92.3 44.3 15 83.3 42.2 17 85.0 44.2 8 100.0 45.8 18 26 96.3 43.5 24 92.3 42.2 14 77.8 41.4 15 75.0 47.0 6 75.0 44.8 22 26 96.3 38.3 19 73.1 35.9 9 50.0 34.2 11 55.0 43,4 5 62.5 42.4 26 13 48.1 36.3 5 19.2 36.6 6 33.3 35.2 9 45.0 43.6 2 25.0 43.0 30 3 11.1 35.3 3 11.5 42.0 3 16.7 31.0 8 40.0 44.5 2 25.0 43.5 34 0 0.0 - 1 3.9 43.0 0 0.0 - 3 15.0 38.7 2 25.0 36.5
Female 6 28 100.0 32.6 29 100.0 34.3 26 100.0 33.8 24 100.0 37.3 20 100.0 34.5 10 27 96.4 38.1 29 100.0 35.7 22 84.6 35.7 23 95.8 39.8 20 100.0 36.9 14 27 96.4 38.8 28 96.6 37.4 19 73.1 37.6 22 91.7 41.5 19 95.0 38.6 18 26 92.9 38.6 28 96.6 39.1 18 69.2 35.8 21 87.5 39.4 15 75.0 39.6 22 25 89.3 35.3 20 69.0 34.7 15 57.7 33.6 15 62.5 33.8 12 60.0 35.0 26 18 64.3 33.3 11 37.9 33.8 10 38.4 33.8 9 37.5 40.0 8 40.0 42.6 30 10 35.7 32.8 8 27.6 33.3 4 15.4 31.0 4 16.7 37.0 5 25.0 37.6 34 2 7:1 -- 4 13.8 34.0 1 3.8 32.0 1 4.2 29.0 0 0.0 --
apercent Surviving.
A d d i n g e i t h e r 0 . 5 % w or 1 . 0 % w 2-MEA, 1 . 0 % w N a H P or 0 . 1 % w BHT to t he d ie t o f f e m a l e m ice be- fore, du r i ng p r e g n a n c y and unt i l t he o f f sp r i ng we re w e a n e d , i nc reased ( P < 0.05)) the a v e r a g e l i fe span o f t he i r m a l e o f f sp r i ng by 18.2, 26.9, 19.2 and 13.5 percen t , respec t i ve l y , as c o m p a r e d to ma les born o f m o t h e r s rece iv ing t he base d ie t . M a l e s born of m o t h e r s rece iv ing d ie ts s u p p l e m e n t e d w i t h 0.1 or 0 . 2 % w S a n t o q u i n , 0 .4% BHT, 0 . 1 % w or 0 . 2 % w BHA, t e n d e d to l ive l onge r t han the con- t ro l s but t he d i f f e r e n c e s are no t s t a t i s t i c a l l y sig- n i f i can t .
The f e m a l e d a t a t ends to pa ra l l e l t ha t o f t he m a l e s bu t none of the pos i t i ve d i f f e rences be- t w e e n a n t i o x i d a n t o f f sp r i ng and con t ro l o f f sp r i ng are s t a t i s t i c a l l y s i gn i f i can t , p o s s i b l y b e c a u s e the f e m a l e con t ro l l i fe span v a l u e is s p u r i o u s l y h igh.
The f e m a l e o f f sp r i ng of t he 0 . 2 % w S a n t o q u i n , 0 . 5 % w and 1 . 0 % w 2-MEA, and 1 . 0 % w N a H P g roups had l i fe spans 6.3, 2.3, 14.5, and 5.1 per- cent , respec t i ve l y , g rea te r t han the con t ro l s ; t he l i fe s p a n s of the o t h e r f e m a l e g roups we re less t han tha t o f t he con t ro l g roup.
The las t m a l e c o n t r o l m o u s e was dead by 32 m o n t h s , t he las t f e m a l e con t ro l by 36 m o n t h s . The l o n g e s t l ived ma le of t he a n t i o x i d a n t o f f sp r i ng , in the 0 . 2 % w BHA group, d ied at 41 m o n t h s . The last f e m a l e in the 0 . 2 % w S a n t o q u i n g roup d ied at 41 m o n t h s w h i l e the l onges t l ived m o u s e in the s tudy was a f e m a l e of t he 0 . 5 % w 2-MEA group, th i s d ied
at 42 m o n t h s . Ma le and f e m a l e m ice in t he 0 . 1 % w S a n t o q u i n ,
0 . 1 % w BHA, and, less obv ious , NaHP groups , we re s o m e w h a t heav ie r t han the con t ro l s ; t he
110
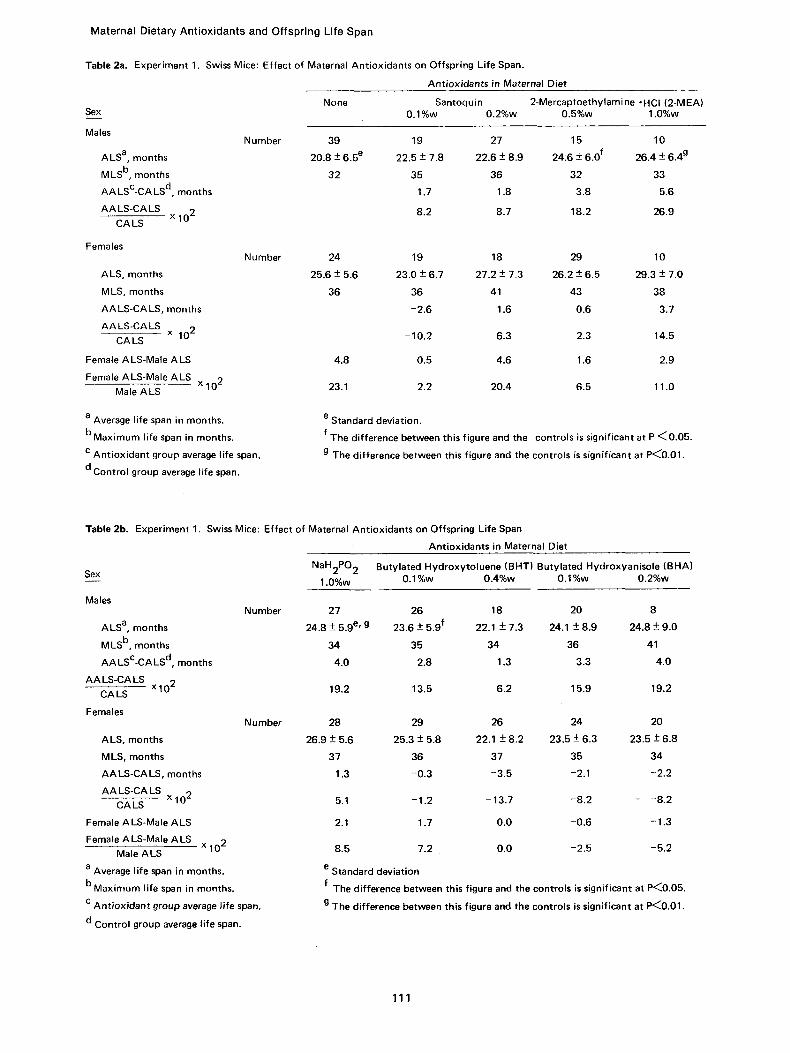
Materna l Dietary An t iox idan ts and Of fspr ing Li fe Span
Table 2a. Experiment 1. Swiss Mice: Effect of Maternal Antioxidants on Offspring Life Span.
Antioxidants in Maternal Diet
Sex
Males
ALS a, months
MLS b, months
AALsc-CALS d, months AALS-CALS
x 102 CA LS
Females
ALS, months
MLS, months
AALS-CALS, months
AALS-CALS x 102 CA LS
Female ALS-Male ALS
Female ALS-Male ALS x 102 Male ALS
a Average life span in months.
b Maximum life span in months.
c Ant ioxidant group average life span.
d Control group average life span.
Number
Number
None Santoquin 2-Mercaptoethylamine ~ (2-MEA) 0.1%w 0.2%w 0.5%w 1.0%w
39 19 27 15 10
20.8 + 6.5 e 22.5 -+ 7.8 22.6 -+ 8.9 24:6 -+ 6.0 f 26.4 -+ 6.4 g
32 35 36 32 33
1.7 1.8 3.8 5.6
8.2 8.7 18.2 26.9
24
25.6 -+ 5.6
36
19 18 29 10
23.0 -+ 6.7 27.2 -+ 7.3 26.2 -+ 6.5 29.3 -+ 7.0
36 41 43 38
-2.6 1.6 0.6 3.7
-10.2 6.3 2.3 14.5
4.8 0.5 4.6 1.6 2.9
23.1 2.2 20.4 6.5 11.0
e Standard deviation.
f The difference between this figure and the controls is significant at P ~0 .05 .
g The difference between this figure and the controls is significant at P~0.01.
Table 2b. Experiment 1. Swiss Mice: Effect of Maternal Antioxidants on Offspring Life Span
Antioxidants in Maternal Diet
Sex
Males
ALS a, months
MLS b, months AALsc-CALS d, months
AALS-CALS x102 CA LS
Females
ALS, months
MLS, months
AA LS-CA LS, months
AA LS-CA LS x 102
CALS
Female ALS-Male ALS
Female ALS-Male ALS x 102 Male ALS
a Average life span in months.
b Maximum life span in months.
c Ant iox idant group average l ife span.
d Control group average life span.
Number
Number
NaH2PO 2
1.0%w
Butylated Hydroxytoluene (BHT) Butylated Hydroxyanisole (BHA) 0.1%w 0.4%w 0.1%w 0.2%w
27 26 18 20 8
24.8 -+ 5.9 e' g 23.6 -+ 5.9 f 22.1 -+ 7.3 24.1 -+ 8.9 24.8 -+ 9.0
34 35 34 36 41
4.0 2.8 1.3 3.3 4.0
19.2 13.5 6.2 15.9 19.2
28 29 26 24 20
26 .9+5 .6 25.3-+5.8 22.1 -+8.2 23.5-+6.3 23.5-+6.8
37 36 37 35 34
1.3 -0 .3 -3 .5 -2.1 -2 .2
5.1 -1.2 -13.7 -8 .2 - -8 .2
2.1 1.7 0.0 -0 .6 -1.3
8.5 7.2 0.0 -2.5 -5 .2
e Standard deviation f The difference between this figure and the controls is significant at P~0.05.
g The difference between this figure and the controls is significant at P<~0.01.
111
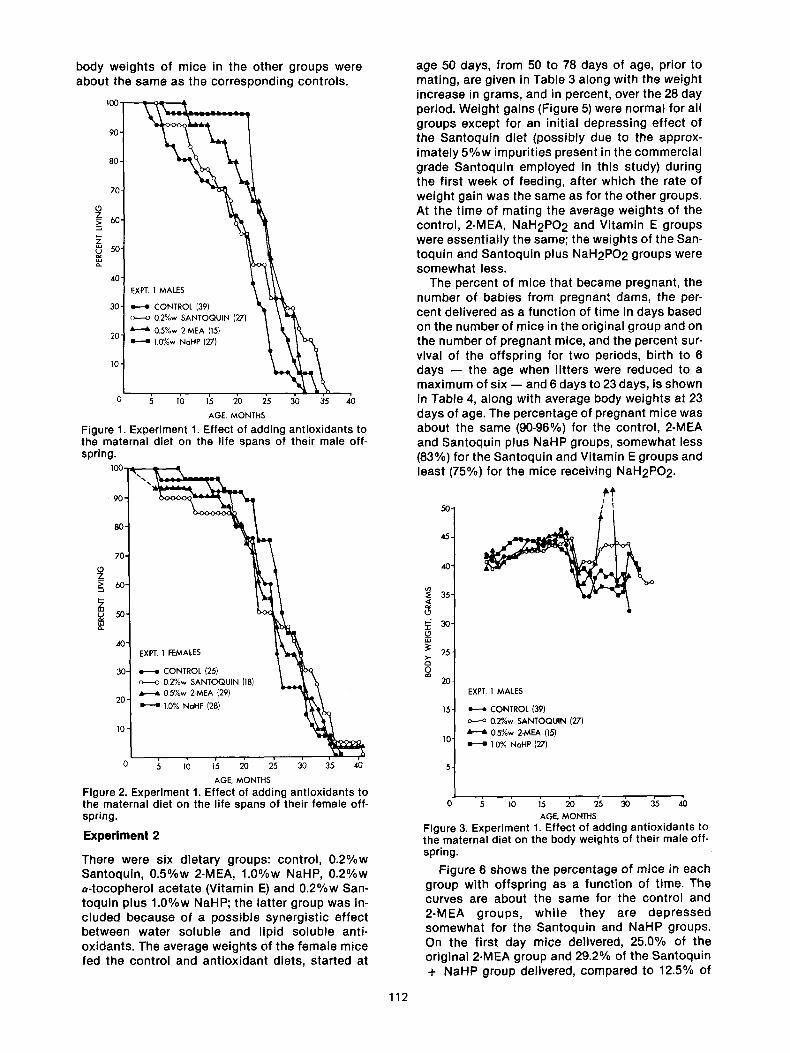
body weights of mice in the other groups were about the same as the corresponding controls.
100, 4
90-
70-
60-
50-
40-
30-
20-
10-
EXPT. 1 MALES
H CONTROL (39) o---o 0.2%w SANTOQUIN (27)
0.5%w 2-MEA (15) H 1.0%w NoHP (27)
0 5 10 15 20 25 30 35 40
AGE, MONTHS
Figure 1. Experiment 1. Effect of adding antioxidants to the maternal diet on the life spans of their male off- spring.
1 0 0 . - v - ~ - - e . - ~ l
90'
80-
70-
O z_ > 6o-
~_ ~0-
40-
20"
EXPT. 1 FEMALES
; : CONTROL (25) o ~ o 0.2%w SANTOQUIN (18) "- • 0.5%w 2-MEA (29)
: : 1.0% NaHP (28)
10"
0 5 10 15 20 25 30 35 40
AGE. MONTH S
Figure 2. Experiment 1. Effect of adding antioxidants to the maternal diet on the life spans of their female off- spring.
E x p e r i m e n t 2
There were six dietary groups: control, 0.2%w Santoquin, 0.5%w 2-MEA, 1.0%w NaHP, 0.2%w e-tocopherol acetate (Vitamin E) and 0.2%w San- toquin plus 1.0%w NaHP; the latter group was in- cluded because of a possible synergistic effect between water soluble and lipid soluble anti- oxidants. The average weights of the female mice fed the control and antioxidant diets, started at
age 50 days, from 50 to 78 days of age, prior to mating, are given in Table 3 along with the weight increase in grams, and in percent, over the 28 day period. Weight gains (Figure 5) were normal for all groups except for an initial depressing effect of the Santoquin diet (possibly due to the approx- imately 5%w impurities present in the commercial grade Santoquin employed in this study) during the first week of feeding, after which the rate of weight gain was the same as for the other groups. At the time of mating the average weights of the control, 2-MEA, NaH2PO 2 and Vitamin E groups were essentially the same; the weights of the San- toquin and Santoquin plus NaH2PO2 groups were somewhat less.
The percent of mice that became pregnant, the number of babies from pregnant dams, the per- cent delivered as a function of time in days based on the number of mice in the original group and on the number of pregnant mice, and the percent sur- vival of the offspring for two periods, birth to 6 days - - the age when litters were reduced to a maximum of six - - and 6 days to 23 days, is shown in Table 4, along with average body weights at 23 days of age. The percentage of pregnant mice was about the same (90-96%) for the control, 2-MEA and Santoquin plus NaHP groups, somewhat less (83%) for the Santoquin and Vitamin E groups and least (75%) for the mice receiving NaH2PO2.
5o. /~
40
35
30 �84 =o
25
o 20
15
10
EXPT. 1 MALES
e--e CONTROL (39) o ~ o 0.2%w SANTOQUIN (27)
0.5%w 2-MEA (15) H 10% NoHP (27)
.~ ib 1's io i5 sb is 4b AGE, MONTHS
Figure 3. Experiment 1. Effect of adding antioxidants to the maternal diet on the body weights of their male off- spring.
Figure 6 shows the percentage of mice in each group with offspring as a function of time. The curves are about the same for the control and 2-MEA groups, while they are depressed somewhat for the Santoquin and NaHP groups. On the first day mice delivered, 25.0% of the original 2-MEA group and 29.2% of the Santoquin + NaHP group delivered, compared to 12.5% of
112
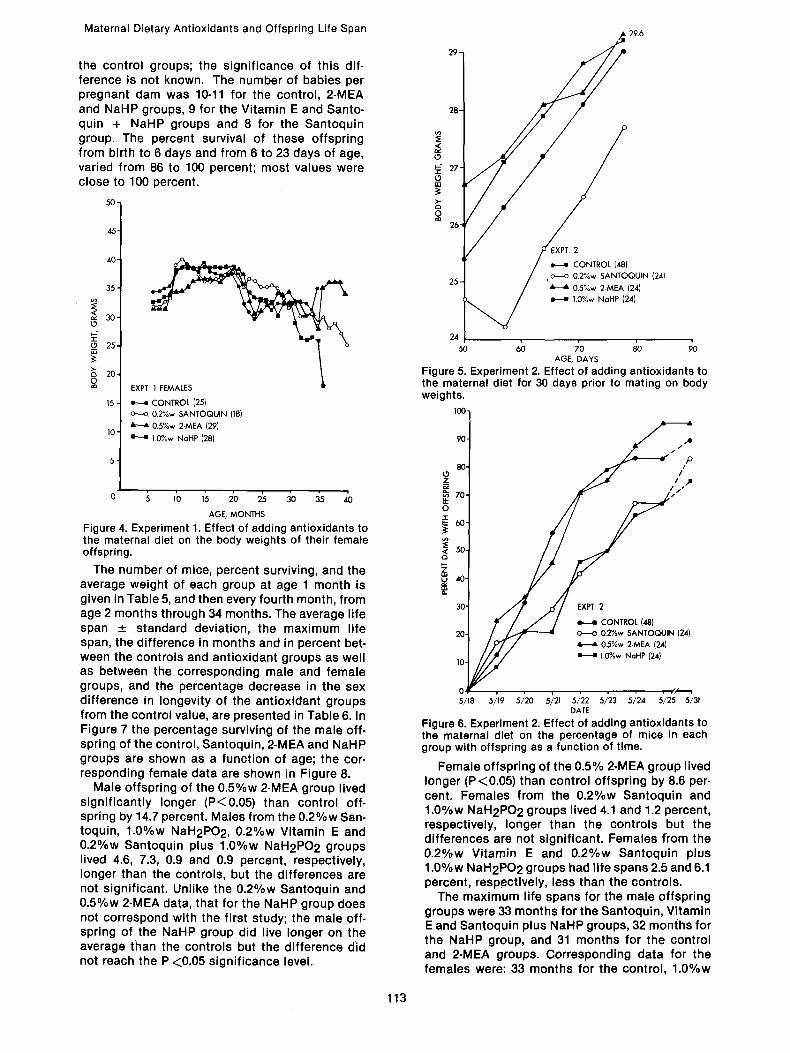
the control groups; the significance of this dif- ference is not known. The number of babies per pregnant dam was 10-11 for the control, 2-MEA and NaHP groups, 9 for the Vitamin E and Santo- q u i n + NaHP groups and 8 for the Santoquin group. The percent survival of these offspring from birth to 6 days and from 6 to 23 days of age, varied from 86 to 100 percent; most values were close to 100 percent.
50-
29-
35-
< 30~
25,
20 o
15,
10 �84
EXPT. 1 FEMALES
H CONTROL (25) 0.2%w SANTOQUlN (18) 0.5%w 2-MEA (29) 1.0%w NaHP (28)
45-
4oJ
0 ~ lb 1~ 2'o 2'5 i0 i5 4b AGE, MONTHS
Figure 4. Experiment 1. Effect of adding antioxidants to the maternal diet on the body weights of their female offspring.
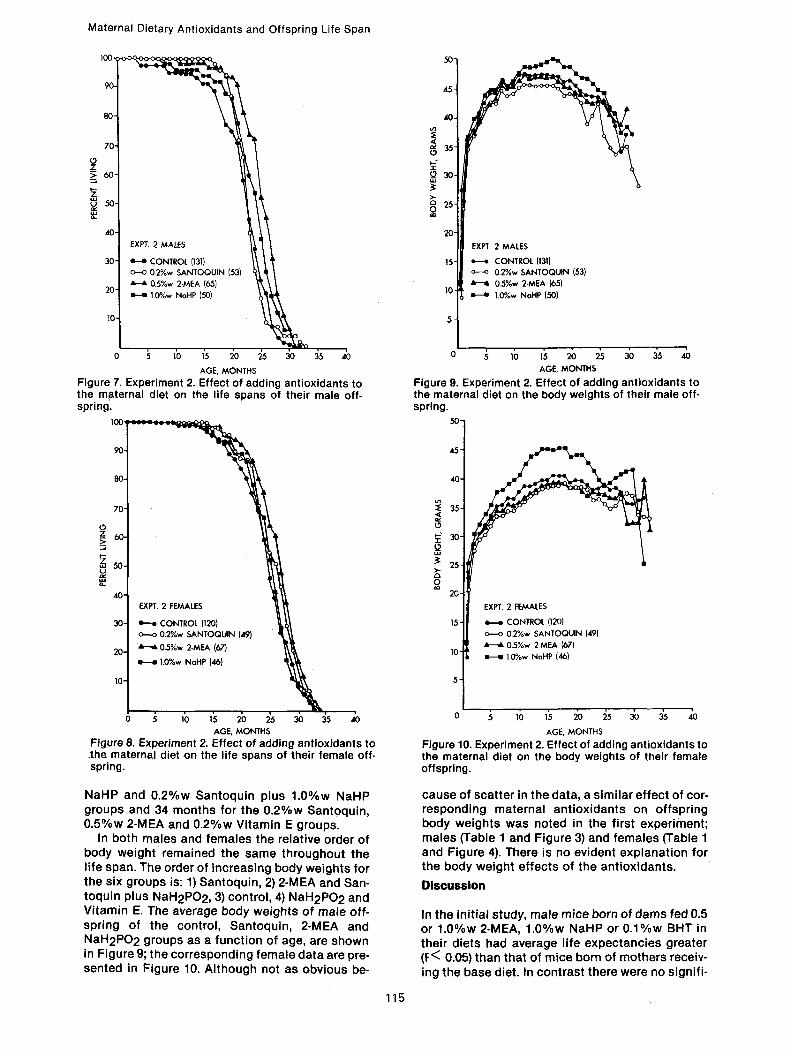
The number of mice, percent surviving, and the average weight of each group at age 1 month is given in Table 5, and then every fourth month, from age 2 months through 34 months. The average life span _+ standard deviation, the maximum life span, the difference in months and in percent bet- ween the controls and antioxidant groups as well as between the corresponding male and female groups, and the percentage decrease in the sex difference in longevity of the antioxidant groups from the control value, are presented in Table 6. In Figure 7 the percentage surviving of the male off- spring of the control, Santoquin, 2-MEA and NaHP groups are shown as a function of age; the cor- responding female data are shown in Figure 8.
Male offspring of the 0.5%w 2-MEA group lived significantly longer (P<0.05) than control off- spring by 14.7 percent. Males from the 0.2%w San- toquin, 1.0%w NaH2PO2, 0.2%w Vitamin E and 0.2%w Santoquin plus 1.0%w NaH2PO 2 groups lived 4.6, 7.3, 0.9 and 0.9 percent, respectively, longer than the controls, but the differences are not significant. Unlike the 0.2%w Santoquin and 0.5%w 2-MEA data, that for the NaHP group does not correspond with the first study; the male off- spring of the NaHP group did live longer on the average than the controls but the difference did not reach the P <0.05 significance level.
28-
25
O 27-
o=
0 26
90 �84
80. ('1 Z
70.
0
~6o.
< 50. Q
30"
20-
10-
Ol 5/18
EXPT. 2
CONTROL (48) o.--o 0.2%w SANTOQUIN (24)
0.5%w 2-MEA (24) 1.0%w NaHP (24)
24 I 5o ~ ~o ~;o ~;o
AGE, DAYS
Figure 5. Experiment 2. Effect of adding antioxidants to the maternal diet for 30 days prior to mating on body weights.
100.
Maternal Dietary Antioxidants and Offspring Life Span �9 29.6
/
EXPT. 2
H CONTROL (48) 02%w SANTOQUIN (24) 0.5%w 2-MEA (24)
H 1.0%w NaHP (24}
. /,z..~l 5719 5/'20 5~21 5722 5/'23 5/;24 5/25 5/31
DATE
Figure 6. Experiment 2. Effect of adding antioxidants to the maternal diet on the percentage of mice in each group with offspring as a function of time.
Female offspring of the 0.5% 2-MEA group lived longer (P<0.05) than control offspring by 8.6 per- cent. Females from the 0.2%w Santoquin and 1.0%w NaH2PO2 groups lived 4.1 and 1.2 percent, respectively, longer than the controls but the differences are not significant. Females from the 0.2%w Vitamin E and 0.2%w Santoquin plus 1.0%w NaH2PO2 groups had life spans 2.5 and 6.1 percent, respectively, less than the controls.
The maximum life spans for the male offspring groups were 33 months for the Santoquin, Vitamin E and Santoquin plus NaHP groups, 32 months for the NaHP group, and 31 months for the control and 2-MEA groups. Corresponding data for the females were: 33 months for the control, 1.0%w
113
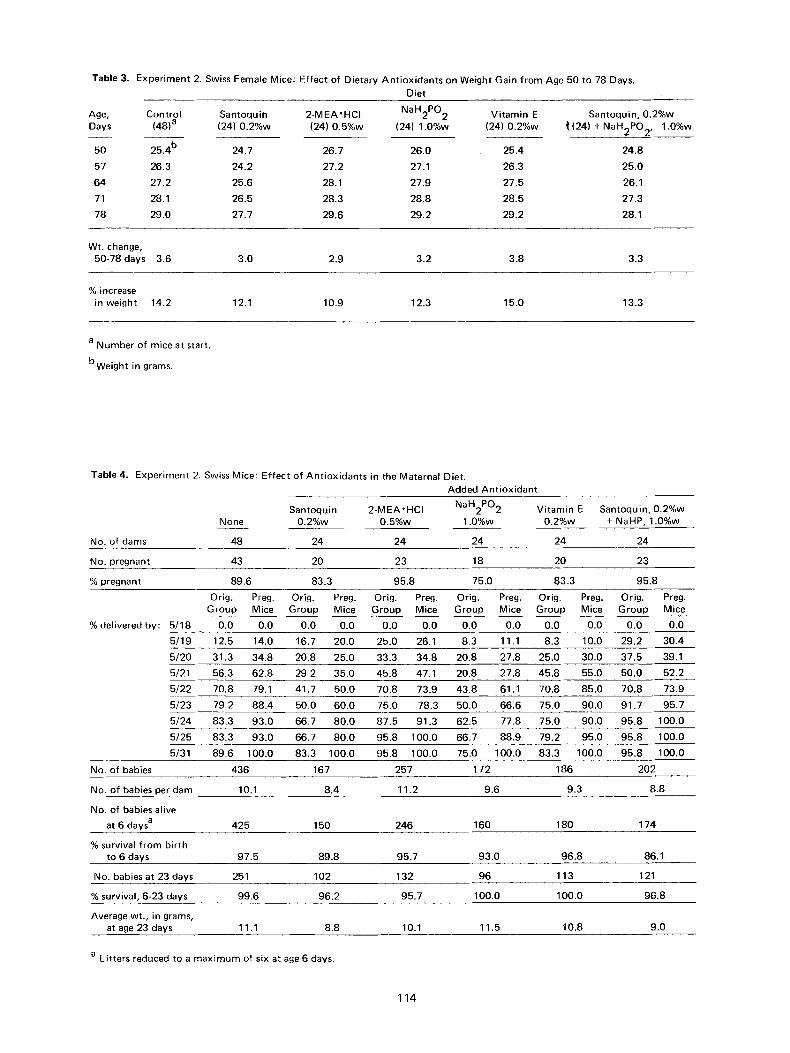
Table 3. Experiment 2. Swiss Female Mice: Effect of Dietary Ant ioxidants on Weight Gain from Age 50 to 78 Days. Diet
Age, Control Santoquin 2-MEA'HCI NaH2PO 2 Vitamin E Santoquin, 0.2%w Days (48) a (24) 0.2%w (24) 0.5%w (24) 1.0%w (24) 0.2%w /((24) + NaH2PO2, 1.0%w
50 25.4 b 24.7 26.7 26.0 25.4 24.8
57 26.3 24.2 27.2 27.1 26.3 25.0
64 27.2 25.6 28.1 27.9 27.5 26.1
71 28.1 26.5 28.3 28.8 28.5 27.3
78 29.0 27.7 29.6 29.2 29.2 28.1
Wt. change, 50-78 days 3.6 3.0 2.9 3.2 3.8 3.3
% increase in weight 14.2 12.1 10.9 12.3 15.0 13.3
a Number of mice at start.
bweight in grams.
Table 4. Experiment 2. Swiss Mice: Effect of Antioxidants in the Maternal Diet. Added Ant ioxidant
Santoquin 2-MEA'HCI NaH2PO2 Vitamin E Santoquin, 0.2%w None 0.2%w 0.5%w 1.0%w 0.2%w + NaHP, 1.0%w
No. of dams 48 24 24 24 24 24
No. pregnant 43 20 23 18 20 23
% pregnant 89.6 83.3 95.8 75.0 83.3 95.8
Orig. Preg. Orig. Preg. Orig. Preg. Orig. Preg. Orig. Preg. Orig. Preg. Group Mice Group Mice Group Mice Group Mice Group Mice Group Mice
% delivered by: 5/18 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
5/19 12.5 14.0 16.7 20.0 25.0 26.1 8.3 11.1 8.3 10.0 29.2 30.4
5/20 31.3 34.8 20.8 25.0 33.3 34.8 20.8 27.8 25.0 30.0 37.5 39.1
5/21 56.3 62.8 29 2 35.0 45.8 47.1 20.8 27.8 45.8 55.0 50.0 52.2
5/22 70.8 79.1 41.7 50.0 70.8 73.9 43.8 61.1 70.8 85.0 70.8 73.9
5/23 79.2 88.4 50.0 60.0 75.0 78.3 50.0 66.6 75.0 90.0 91.7 95.7
5/24 83.3 93.0 66.7 80.0 87.5 91.3 62.5 77.8 75.0 90.0 95.8 100.0
5/25 83.3 93.0 66.7 80.0 95.8 100.0 66.7 88.9 79.2 95.0 95.8 100.0
5/31 89.6 100.0 83.3 100.0 95.8 100.0 75.0 100.0 83.3 100.0 95.8 100.0
No. of babies 436 167 257 172 186 202
No. of babies per dam 10.1 8.4 11.2 9.6 9.3 8.8
No. of babies alive
at 6 days a 425 150 246 160 180 174
% survival from birth to 6 days 97.5 89.8 95.7 93.0 96.8 86.1
No. babies at 23 days 251 102 132 96 113 121
% survival, 6-23 days 99.6 96.2 95.7 100.0 100.0 96.8
Average wt., in grams, at age 23 days 11.1 8.8 10.1 11.5 10.8 9.0
a Litters reduced to a maximum of six at age 6 days.
114
Maternal Dietary Antioxidants and Offspring Life Span
100 , ~
80,
O Z
60-
~ 5o-
40-
t EXPT. 2 MALES
30- H CONTROL (131) o--o 0.2%w SANTOQUIN (53)
0.5%w 2-MEA (65) 20- H 1.0%w NaHP (50)
o ~ lb 1'5 ~o ~ :~o is 4b AGE, MONTHS
Figure 7. Experiment 2. Effect of adding antioxidants to the maternal cliet on the life spans of their male off- spring.
O z
~5o. u
30-
EXPT. 2 FEMALES
e~e CONTROL (120) o ~ o 0.2%w SANTOQUIN (49)
0.5%w 2-MEA (67)
H 1.0%w NaHP (46)
50'
45-
40
~35 �84 t-- _~ 30:
~25- 2
20- EXPT. 2 MALES
15- ~ CONTROL (131) o--..o 0.2%w SANTOQUIN (53)
05%w 2-MEA (65) 10 1.0%w NoHP (50}
5
0 ~ 15 1~ 2b ~ 3b ~ 4b AGE, MONTHS
Figure 9. Experiment 2. Effect of adding antioxidants to the maternal diet on the body weights of their male off- spring.
.50"1
45-
40-
35"
-y- 30- =o_
25- o
20-
15"
10-
5-
EXPT. 2 FEMALES
H CONTROL (120) 0.2%w SANTOQUIN (49) 0.5%w 2-MEA (67)
H 1.0%w NaHP (46)
0 5 10 15 20 25 30 35 40 AGE, MONTHS
Figure 8. Experiment 2. Effect of adding antioxidants to 1he maternal diet on the life spans of their female off- spring.
NaHP and 0 .2%w Santoquin plus 1.0%w NaHP groups and 34 months for the 0 .2%w Santoquin, 0 .5%w 2-MEA and 0 .2%w Vitamin E groups.
In both males and females the relative order of body weight remained the same throughout the life span. The order of increasing body weights for the six groups is: 1) Santoquin, 2) 2-MEA and San- toquin plus NaH2PO2, 3) control, 4) NaH2PO 2 and Vitamin E. The average body weights of male off- spring of the control, Santoquin, 2-MEA and NaH2PO2 groups as a function of age, are shown in Figure 9; the corresponding female data are pre- sented in Figure 10. Although not as obvious be-
115
0 5 10 15 20 25 30 35 40
AGE, MONTHS
Figure 10. Experiment 2. Effect of adding antioxidants to the maternal diet on the body weights of their female offspring.
cause of scatter in the data, a similar effect of cor- responding maternal antioxidants on offspring body weights was noted in the first experiment; males (Table 1 and Figure 3) and females (Table 1 and Figure 4). There is no evident explanation for the body weight effects of the antioxidants.
Discussion
In the initial study, male mice born of dams fed 0.5 or 1.0%w 2-MEA, 1.0%w NaHP or 0 .1%w BHT in their diets had average life expectancies greater (F< 0.05) than that of mice born of mothers receiv- ing the base diet. In contrast there were no signifi-
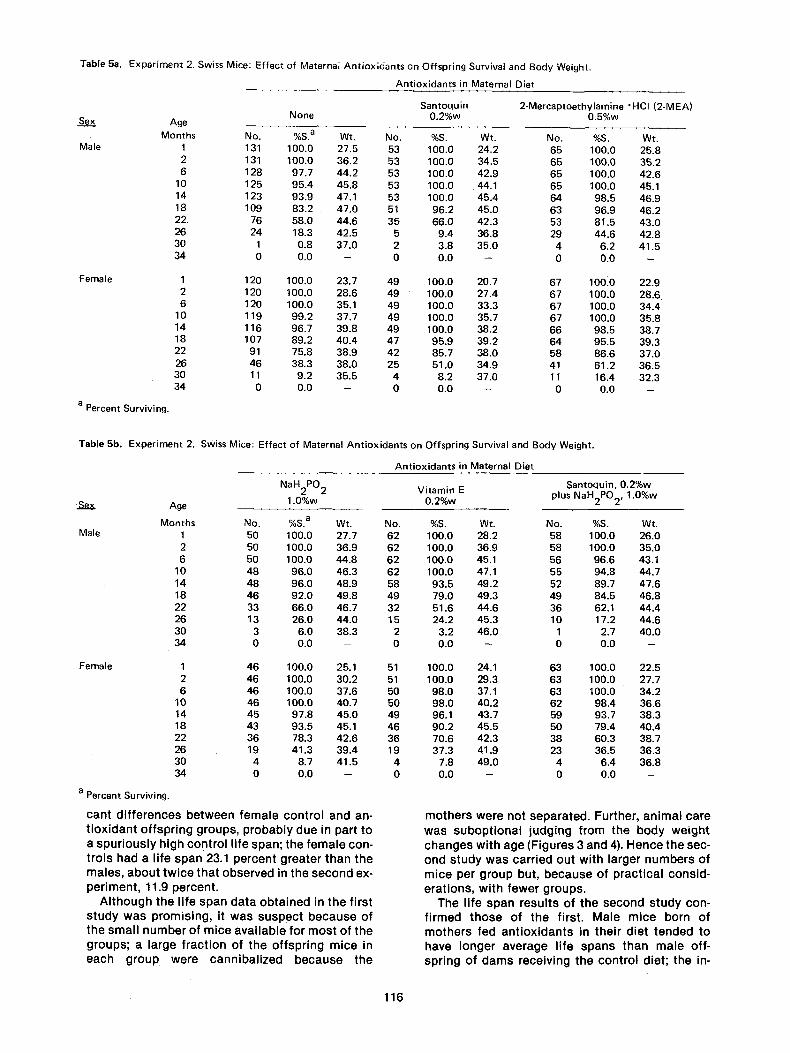
Table 5a. Experiment 2. Swiss Mice: Effect of Maternal Antioxidants on Offspring Survival and Body Weight.
Se__._~x
Male
Female
a Percent Surviving.
Antioxidants in Maternal Diet
Santoquin 2-Mercaptoethylamine ~ (2-MEA) None 0.2%w 0.5%w Age
Months No. %S. a Wt. No. %S. Wt. No. %S. Wt. 1 131 100.0 27.5 53 100.0 24.2 65 100.0 25.8 2 t31 100.0 36.2 53 100.0 34.5 65 100.0 35.2 6 128 97.7 44.2 53 100.0 42.9 65 100.0 42.6
10 125 95.4 45.8 53 100.0 44.1 65 100.0 45.1 14 123 93.9 47.1 53 100.0 45.4 64 98.5 46.9 18 109 83.2 47.0 51 96.2 45.0 63 96.9 46.2 22. 76 58.0 44.6 35 66.0 42.3 53 81.5 43.0 26 24 18.3 42.5 5 9.4 36.8 29 44.6 42.8 30 1 0.8 37.0 2 3.8 35.0 4 6.2 41.5 34 0 0.0 - 0 0.0 - 0 0.0 -
1 120 100.0 23.7 49 100.0 20,7 67 100.0 22.9 2 120 100.0 28.6 49 100.0 27.4 67 100.0 28.6 6 120 100.0 35.1 49 100.0 33.3 67 100.0 34.4
10 119 99.2 37.7 49 100.0 35.7 67 100.0 35.8 14 116 96.7 39.8 49 100.0 38,2 66 98.5 38.7 18 107 89.2 40.4 47 95.9 39.2 64 95.5 39.3 22 91 75.8 38.9 42 85.7 38.0 58 86.6 37.0 26 46 38.3 38.0 25 51.0 34.9 41 61.2 36.5 30 11 9.2 35.5 4 8.2 37,0 11 16.4 32.3 34 0 0.0 -- 0 0.0 -- 0 0.0 --
Table 5b. Experiment 2. Swiss Mice: Effect of Maternal Antioxidants on Offspring Survival and Body Weight.
Antioxidants in Maternal Diet
NaH2P02 Vitamin E Age 1.0%w 0.2%w
Months No. %S. a Wt. No. %S. Wt. Male 1 50 100.0 27.7 62 100.0 28.2
2 50 100.0 36.9 62 100.0 36.9 6 50 100.0 44.8 62 100.0 45.1
10 48 96.0 46.3 62 100.0 47.1 14 48 96.0 48.9 58 93.5 49.2 18 46 92.0 49,8 49 79.0 49.3 22 33 66.0 46.7 32 51.6 44.6 26 13 26.0 44.0 15 24.2 45.3 30 3 6.0 38.3 2 3.2 46.0 34 0 0.0 -- 0 0.0 --
Female 1 46 100.0 25.1 51 100.0 24.1 2 46 100.0 30.2 51 100.0 29.3 6 46 100.0 37.6 50 98.0 37.1
10 46 100.0 40.7 50 98.0 40.2 14 45 97.8 45.0 49 96.1 43.7 18 43 93.5 45.1 46 90.2 45.5 22 36 78.3 42.6 36 70.6 42.3 26 19 41.3 39.4 19 37.3 41.9 30 4 8.7 41.5 4 7.8 49.0 34 0 0.0 -- 0 0.0 --
a Percent Surviving.
can t d i f f e r e n c e s b e t w e e n f e m a l e con t ro l and an- t i o x i d a n t o f f sp r i ng g roups , p r o b a b l y due in par t to a s p u r i o u s l y h igh con t ro l l i fe span; the f e m a l e con- t ro i s had a l i fe span 23.1 pe rcen t g rea te r t han the ma les , a b o u t t w i c e t ha t obse rved in the s e c o n d ex- pe r imen t , 11.9 percen t .
A l t h o u g h the l i fe span d a t a o b t a i n e d in the f i rs t s t u d y w a s p rom is i ng , it was suspec t b e c a u s e of the sma l l n u m b e r o f m i ce a v a i l a b l e for m o s t o f t he g roups ; a la rge f r ac t i on of t he o f f sp r i ng m i ce in each g r o u p we re c a n n i b a l i z e d b e c a u s e t he
Santoquin, 0.2%w plus NaH2PO 2, 1.0%w
No. %8. Wt. 58 100.0 26.0 58 100.0 35.0 56 96.6 43.1 55 94.8 44.7 52 89.7 47.6 49 84.5 46.8 36 62.1 44.4 10 17.2 44.6
1 2.7 40.0 0 0.0 -
63 100.0 22.5 63 100.0 27.7 63 100.0 34.2 62 98.4 36.6 59 93.7 38.3 50 79.4 40.4 38 60.3 38.7 23 36.5 36.3
4 6.4 36.8 0 0.0 --
m o t h e r s we re no t sepa ra ted . Fur ther , a n i m a l ca re was s u b o p t i o n a l j u d g i n g f rom the b o d y w e i g h t c h a n g e s w i t h age (F igures 3 and 4). H e n c e the sec- ond s t u d y w a s car r ied ou t w i th la rger n u m b e r s o f m i ce per g roup but , b e c a u s e of p rac t i ca l cons id - e ra t i ons , w i t h f ewe r g roups .
The l i fe span resu l t s o f t he second s t u d y con- f i rmed t h o s e of t he f i rst . Ma le m ice born of m o t h e r s fed a n t i o x i d a n t s in the i r d ie t t ended to have l onge r a v e r a g e l i fe spans t han m a l e of f - sp r ing o f c lams rece iv ing the con t ro l d ie t ; t he in-
116

Maternal Dietary Antioxidants and Offspring Life Span
crease was significant, P<0.05, for the 2-MEA group. The female data were similar but the dif- ferences between treated and control offspring were less; as with the males, only females of the 2-MEA group lived significantly longer than the control offspring (P<0.05). Although the San- toquin group offspring did not live significantly longer than the controls, they had the lowest early mortality rate of any of the offspring groups; the first male Santoquin offspring died at 17 months and the first female at 15 months. There is no ob- vious explanation for this effect of Santoquin. The results of adding both a lipid soluble antioxidant, Santoquin, and a water soluble one, NaH2PO2, to the maternal diet were disappointing. Instead of a synergistic effect, the combination reduced off- spring life span, females more than males. This re- sult, like that of a similar antioxidant combination on the humoral response of mouse splenic cells to sheep red blood cells (16), may be due to the tox- icity of relatively high antioxidant concentrations.
Life span data for the four groups in common in the two studies - - control, Santoquin (0.2%w), 2-MEA (0.5%w) and NaH2PO2 (1.0%w) - - are presented in Table 7; the data are in good agree- ment considering the relatively small numbers in the groups of the first experiment. The relative effectiveness of the antioxidants on life span, as well as their other effects (Table 4), reflect the dif- ferences between the antioxidants, including water and lipid solubility, absorption, metabolism, excretion, concentration in oviducal (17) and uter- ine fluids, ease of crossing the placental barrier (18, 19), concentration in the mothers milk, con- centration time course in the cells and t issues, and intervention in normal metabolic processes, both involving free radical reactions and other- wise.
These two studies demonstrate that addition of suitable free radical reaction inhibitors to the ma- ternal diet, from one month before mating until weaning, can significantly increase the average life span of their offspring; there were no certain effects on the maximum life span. The most effec- tive compound, 2-MEA, at a dietary concentration of 0.5%w, increased the average life span of male offspring by about 15 percent and of females by around 8 percent.
The beneficial effect of maternal antioxidants on offspring life span is attributed to suppression of damage normally produced by endogenous free radical reactions during the approximately 40 day period between conception and weaning. Other explanations can be advanced. For example, ma- ternal malnutrition can have a significant deleter- ious effect (20) on the complex processes (21) in- volved in ovulation, fertilization, embryogenesis and fetal development. Hence in this study, the differences between the dietary groups (Table 4) in fertility, gestation period, percentage of offspring surviving from birth to 6 days and 6 days to 23 days of age, body weight at 23 days as well as off-
spring life span, may have been the result of variable degrees of protection from oxidative damage by the antioxidants of a dietary com- ponent(s) present in adequate amounts for nutri- tion under normal circumstances but inadequate under the stress of pregnancy. Further, anti- oxidants have a multiplicity of effects on bio- logical systems (22, 23), not all necessarily related to their free radical reaction activity. Thus, 2-MEA depletes liver glycogen in mice (24) while Vitamin E inhibits membrane Na+ K+-ATPase (25). Con- ceivably, non-free radical related effects of antiox- idants in the mother and/or offspring could be responsible for the differences noted in the dietary groups. However, the most likely expla- nation of the beneficial effects of maternal anti- oxidants on their offspring is a decrease in free radical reaction mediated damage leading to long- term adverse consequences, including alterations in DNA, both nuclear and mitochondrial, and changes in cell surface glycoproteins involved in morphogenesis (26).
Consideration of early events in embryogenesis in the light of the present work ~rovides a possible explanation for the greater longevity of females. In the second study female control offspring lived 11.9 percent longer than the male control off- spring; this sex difference in longevity is essen- tially the same as for humans, 11.4 percent (27). The two water soluble compounds, 2-MEA and NaH2PO2, reduced the sex diference in longevity to 6.0 and 5.6 percent, respectively, while the two lipid soluble compounds, Santoquin and Vitamin E, had essentially no effect in the first case and a modest one in the second, the sex differences being 11.0 and 8.6 percent, respectively; the lon- gevity sex difference for the Santoquin plus NaH2PO 2 groups, 4.1 percent, is similar to that for 2-MEA and NaH2PO 2. Similar results were ob- tained in the first study but were ignored at first because of uncertainty in the data due to the small numbers of mice in the various groups and probable suboptional animal care.
The developing mouse embryo (11, 12, 28, 29) reaches the 8 cell, 16-32 cell (morula), and late blastocyst stage at about 60, 72-84, and 108 hours, respectively, after ovulation. Around 72 hours after ovulation, the 16-32 cell embryo emerges from the fallopian tube (30) into the uterus where implantation occurs approximately 48 hours later during the late blastocyst state of development. Between the 8 cell and morula stages, about con- incident with the change from tubal to uterine en- vironment, there is a marked increase in oxygen consumption (11) by the embryo which persists, at least until implantation. Intrauterine oxygen serves as the source of oxygen for the embryo. Studies in the rat (31) show that at ovulation the oxygen tension in the uterine cavity rises from about 22 mm Hg, 7 mm below the venous pO 2 of 29 mm, to around 38 mm on the first day after ovulation, and remains elevated until implantation
117
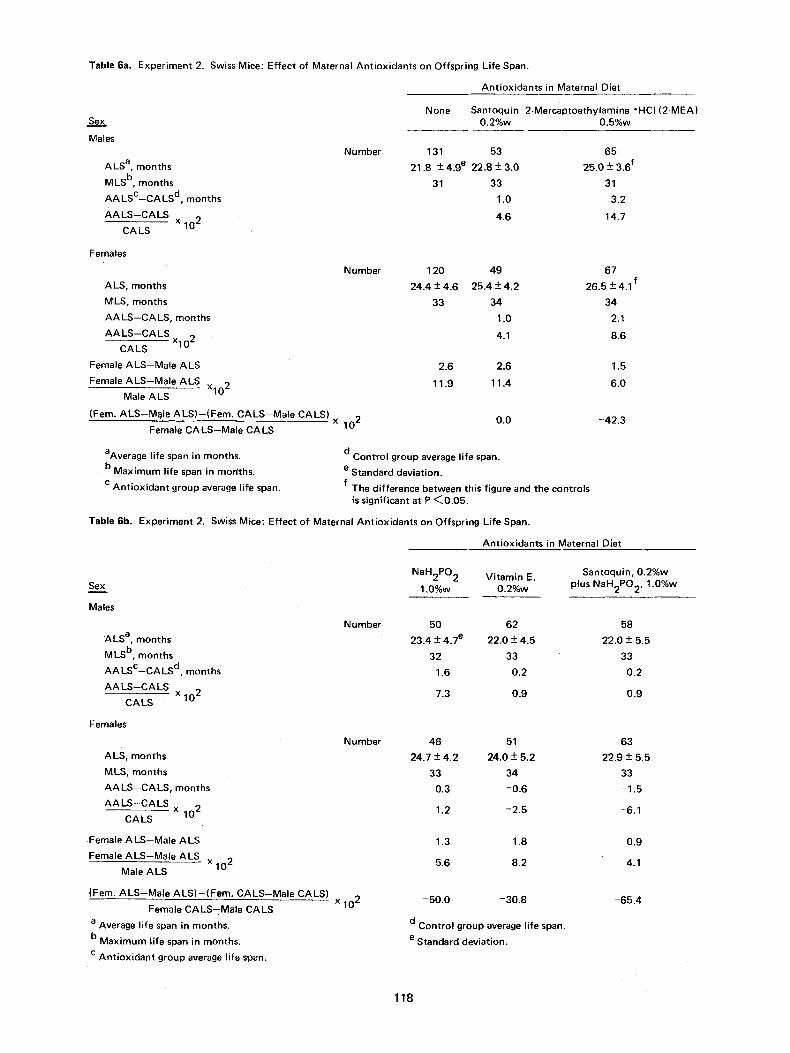
Table 6a. Experiment 2, Swiss Mice: Effect of Maternal Ant• on Offspring Life Span.
Seix,
Males
ALS a, months
MLS b, months
AALsc--CALS d, months
AALS--CALS x C A LS 102
Ant• in Maternal Diet
None Santoquin 2-Mercaptoethylamine "HCI (2-MEA 0.2%w 0.5%w
Number 131 53 65
21.8 +- 4.9 e 22.8 +- 3.0 25.0 +- 3.6 f
31 33 31
1.0 3.2
4.6 14.7
Females
ALS, months
MLS, months
AA LS--CALS, months
AALS--CALS x 102
CA LS
Female ALS--Male ALS
Female ALS,Male ALS x102 Male ALS
(Fern. ALS--M~Ie ALS)--(Fem. CALS--Male CALS)
Female CALS--Male CALS
aAverage life span in months.
b Maximum life span in morlths.
c Antioxidant group average life span.
Number 120 49 67
24.4 + 4.6 25.4 • 4.2 26.5 • 4.1 f
33 34 34
1.0 2.1
4.1 8.6
2.6 2.6 1.5
11.9 11.4 6.0
x 102 0.0 -42.3
d Control group average life span.
e Standard deviation. f The difference between this figure and the controls
is significant at P <0.05,
Table 6b. Experiment 2. Swiss Mice: Effect of Maternal Ant• on Offspring Life Span.
Ant• in Maternal Diet
Sex
Males
ALS a, months
MLS b, months
A A L s c ' C A L S d, months
AALS--CALS x
CALS 102
Females
ALS, months
MLS, months
AALS--CALS, months
AA LS --C A LS X
CALS 102
Female ALS--Male ALS
Female ALS--Male ALS x 102
Male ALS
(Fern. ALS-Male ALS)--(Fem. CALS-Male CALS)
Female CALS--Male CALS
a Average life span in months.
b Maximum life span in months.
c Antioxidant group average life span.
Number
Number
x 102
NaH2PO2 Vitamin E, 1.0%w 0.2%w
50 62
23.4 • 4.7 e 22.0 + 4.5
32 33
1.6 0.2
7.3 0.9
46 51
24.7 • 4.2 24.0 + 5.2
33 34
0.3 -0.6
1.2 -2.5
1.3 1.8
5.6 8.2
-50.0 -30.8
d Control group average life span.
e Standard deviation.
Santoquin, 0.2%w plus NaH2PO2, 1.0%w
58
22.0 • 5.5
33
0.2
0,9
63
22.9•
33
-1.5
-6.1
0.9
4.1
-65.4
118

Maternal Dietary Antioxidants and Offspring Life Span
Table 7. Swiss Mice: Effect of Adding Antioxidants to the Maternal Diets on the Life Spans a of Their Offspring.
Antioxidants in Maternal Diet
None Santoquin 2-MEA NaH2P02 0.2%w 0.5%w 1.0%w
Males
Experiment 1 20.8 -+ 6.5 c (39) b 22.6 + 8.9 (27) 24.6 -+ 6.0 (15) 24.8 + 5.9 (27)
Experiment 2 22.2 + 4.0 (128) 22.8 -+ 3.0 (53) 25.0 -+ 3.6 (65) 23.4 -+ 4.7 (50)
Females
Experiment 1 25.6 -+ 5.6 (24) 27.2 + 7.3 (18) 26.2 -+ 6.5 (29) 26.9 -+ 5.6 (28)
Experiment 2 24.4 -+ 4.6 (120) 25.4 + 4.2 (49) 26.5 -+4.1 (67) 24.7 -+4.2 (46)
a Survival at 6 months set at 100 percent. b
Number alive at 6 months of age. c Standard deviation.
occurs when the pO 2 again increases. The rat intrauterine oxygen tension, presumably mouse intrauterine oxygen tensions are similar, is rela- tively high compared to the average values in tissues (32).
Prior to some time between the 40-50 cell blastocyst stage, fourth day of gestation, and well after implantation, ninth day of gestation, female embryos have two functioning X-chromosomes while male embryos have one (33-35). The X-chromosome codes for glucose-6-phosphate de- hydrogenase (G6PD) (36). G6PD is a key enzyme in maintaining cell levels of NADPH which in turn regulate the level of glutathione (37). Glutathione, the major cellular sulphydryl compound, serves to minimize free radical damage to the organism by acting as a free radical reaction inhibitor (38), in the same manner as other mercaptans, and as a hydrogen donor for glutathione peroxidase in the reduction of hydrogen peroxide and hydroperox- ides (39). One of the two functional X-chromo- somes is inactivated (34, 35) in female embryos at random between the fourth and ninth day of gesta- tion. Hence, during the period of time, probably at least 48 hours, between the 8 cell stage and X-chromosome inactivation, a period of high ox- idative metabolism, high mitotic rate and ex- posure to relatively high environmental oxygen tension, female embryos may be better protected from free radical reaction than males. In short, during the period that the female embryo possesses two functioning X-chromosomes per cell, it should be better protected from damaging free radical reactions than the male embryo with its one X-chromosome per cell because the X-chromosome codes for G6PD, an enzyme which indirectly enhances cellular protection from free radical reactions by raising the cellular levels of glutathione and glutathlone peroxidase.
Assuming that at least part of the "damage", for example, alterations in nuclear and mitochon- drial DNA, is passed on to the daughter cells, male offspring would be "older" at birth than females and have a shorter life span. Addition of free radical reaction inhibitors to the maternal diet would be expected to result in about equal con-
centrations of the inhibitors in male and female embyros, but the relative increase in the ability to limit free radical reaction damage should be greater in the males prior to the random inactiva- tion of one of the two functioning X-chromosomes present in the female cells. This differential pro- tective effect of maternal antioxidants before X-chromosome inactivation may be measured by the percentage decrease in the sex differences in longevity of the antioxidant groups from the con- trol value. An antioxidant that reduces the sex differential in longevity may or may not increase offspring life span. Most of the time spent by the embryo prior to X-chromosome inactivation is passed before implantation. Hence the free radical reaction protect ive effect before X-chromosome inactivation depends largely on passage of the antioxidant into the oviducal and uterine fluids whereas later in development it is in- fluenced by the placental barrier and excretion in- to the mother's milk.
In the second study the percentage decreases in the sex longevity difference in the 2-MEA, NaH2PO 2, Santoquin, Vitamin E, and Santoquin plus NaH2PO 2 groups were -42.3, -50.0, 0.0, -30.8 and -65.4 percent, respectively. The water soluble antioxidants were more effective than the lipid soluble ones in reducing the sex difference in longevity as well as, generally, in increasing off- spring longevity. These observations probably reflect both antioxidant differences and the higher molar concentration of the water soluble antiox- idants in the diets; the molar concentrations in the diets, relative to Vitamin E, of 2-MEA, NaH2PO2, Santoquin, Vitamin E, and Santoquin plus NaH2PO 2 were 10.2, 22.4, 2.2, 1.0, and 2.2 plus 22.4, respectively.
In view of the present work, human maternal diets formulated to minimize more-or-less random endogenous free radical reactions, for example, by decreasing the intake of substances such as polyunsaturated lipids and copper that might reasonably be expected to participate readily in free radical reactions (40), might increase the life span of both sexes while decreasing the normal difference in life span between the sexes.
119

Experimental Procedures
Experiment 1
Charles River CD strain female mice (Charles River Breeding Laboratories, Wilmington, Massa- chusetts) were obtained at weaning and caged 5 per cage (stainless steel, 12x8x5 inches). The mice were maintained in an air-conditioned room at 75 ~ - 78 ~ at a humidity of 50-60 percent. Cages were changed 2-3 times per week; the bedding was sterilized shredded corn cobs (San-i-Cel, Paxton Processing Company, Paxton, II1.). The mice were placed on a semi-synthetic diet containing 5%w lard (41, 42) as a sole source of lipid, plus 2.5 mg. of ,-tocopherol acetate per 100 grams of finished diet, until 50 days of age when they were divided into dietary groups by adding nothing, control, or one of several antioxidants to the diet: 0.1%w and 0.2%w Santoquin (MW -- 216), 0.5%w and 1.0%w 2-MEA (MW of the hydrochloride = 114), 1.0%w NaHP (MW of the monohydrate = 106), 0.1%w and 0.4%w BHT (MW = 220), and 0.1%w and 0.2%w BHA (MW = 180); BHA has not previously been used in mouse aging studies. There were 20 mice in the control group and 10 in each of the ex- perimental groups.
Four weeks after being placed on the special diets one CD male, same age as the females, was placed in each cage for a period of one week; all the females apparently became pregnant. Mice conceived during this period were born 17-22 days after the males were removed. Many of the off- spring were cannibalized, probably largely because the original five females per cage were kept in the same cage with their offspring until weaning. Although it would have been desirable to have had 40-60 mice for each group in the life span study, it was elected to proceed with the mice available for it was not feasible at the time to repeat the breeding phase.
When the remaining offspring were 4 weeks old the males and females were placed in separate cages (10 per cage in most cases) and kept on a commercial pelleted mouse chow (Rockland, Teklad, Inc., Monmouth, II1.) for the rest of their lives; cages were not consolidated as mice died. Mice were counted and weighed monthly starting at age 6 months; data were inadvertently not tabulated for the period prior to 6 months of age.
Experiment 2
This experiment was conducted in the same man- ner as the first except that: 1) initially four mice were caged per cage, 2) 48 females were mated in the control group and 24 in each of the treated groups; 0.2%w Santoquin, 0.5%w 2-MEA, 1.0%w NaHP, 0.2%w ,-tocopherol acetate, and 0.2%w Santoquin plus 1.0%w NaHP; the molar amounts in the diet of the first four groups, relative to Vitamin E, were 2.2, 10.2, 22.4 and 1.0, respec- tively, 3) after the male was removed, each of the
four females was placed in a separate cage, 4) at age six days the litters were reduced to 6, and 5) mice were weighed and counted monthly starting at age one month.
Acknowledgement
The authors gratefully acknowledge the financial assistance of the Paul F. Glenn Foundation, the Monsanto Company, the late Mrs. Adah Millard of Omaha, Nebraska, and subsequently, of the Millard Trust. The conscientious assistance of Larry W. Lass helped make the study possible.
Abbreviations
S a n t o q u i n is 1 ,2-d ihyd ro-6-et hoxy-2, 2,4-trimethylquinoline. 2-MEA is 2-mercaptoethyl- amine oHCI. NaHP is NaH2PO2.H20, sodium hypophosphite. BHT is butylated hydroxytoluene. BHA is butylated hydroxyanisole. Vitamin E is ,-tocopherol acetate. NADPH is nicotinamide adenine dinucleotide phosphate. G6PD is glucose- 6-phosphate dehydrogenase.
References
1. Isenberg, I.: Free radicals in tissue. Physiol. Rev., 44: 487-517, 1964.
2. Harman, D.: Aging: a theory based on free radical and radiation chemistry. J. Gerontol., 11: 298-300, 1956.
3. Harman, D.: Role of free radicals in muta- tion, cancer, aging, and the maintenance of life. Radiation Res., 16: 753-763, 1962.
4. Harman, D.: Prolongation of life: role of free radical reactions in aging. J. Amer. Geriatrics Soc., 17: 721-735, 1969.
5. Harman, D.: Prolongation of the normal life span by radiation protection chemicals. J. Gerontol., 12: 257-263, 1957.
6. Hun-Hoi, N. P., and Ratsimamanger, A. R.: Age retardation in the rat by nordihydro- guaiaretic acid. C. r. Seanc. Soc. Biol., 153: 1180-1182, 1959.
7. Harman, D.: Prolongation of the normal life span and inhibition of spontaneous cancer by antioxidants. J. Gerontol., 16: 247-254, 1961.
8. Harman, D.: Free radical theory of aging: effect of free radical reaction inhibitors on the mortality rate of male LAF 1 mice. J. Gerontol., 23: 476-482, 1968.
120

Maternal Dietary Antioxidants and Offspring Life Span
9.
10.
11.
12.
13.
14.
15.
16.
17.
18.
19.
20.
21.
Comfort, A.: Effect of ethoxyquin on the longevity of C3H mice. Nature, 229: 254-255, 1971.
Emanuel, N. M.: Free radicals and the action of inhibitors of radical processes under pathological states and aging in living organisms and in man. Quart. Rev. Bio- physics, 9: 283-308, 1976.
Mills, Jr., R. M., and Brinster, R. L.: Oxygen consumption of preimplantation mouse em- bryos. Exper. Cell Res., 47: 337-344, 1967.
McLaren, A.: The embryo, in Reproduction in Mammals, Book 2, Embryonic and Fetal Development, edited by Austin, C. R., and Short, R. V., Cambridge, University Press, 1972, pp. 1-42.
Thompson, J. F.: Protection by thiols, in Radiation Protection in Mammals, New York, Reinhold Publishing Corp., 1962, pp. 64-65.
Bacq, Z. M., and Alexander, P.: Delayed effects, in Fundamentals of Radiobiology, New York, Pergamon Press, 1961, pp. 447-448.
Rugh, R., and Clugston, H.: Protection of mouse fetus against X-irradiation death. Science, 123: 28-29, 1956.
Harman, D., Heidrick, M. D., and Eddy, D. E.: Free radical theory of aging: effect of free radical reaction inhibitors on the immune response. J. Amer. Geriatrics Soc., 25: 400-407, 1977.
Beier, H. M.: Oviducal and uterine fluids. J. Reprod. Fert., 37: 221-237, 1974.
Hagerman, D. D., and Villee, C. A.: Transport function of the placenta. Physiol. Rev., 40: 313-330, 1960.
Moya, F., and Thorndike, V.: Passage of drugs across the placenta. Amer. J. Obst. & Gyn., 84: 1778-1798, 1962.
Jacobson, H. N.: Nutrition, in Scientific F o u n d a t i o n s of O b s t e t r i c s and Gynaecology, 2nd Edition, edited by Philipp, E. E., Barnes, J., and Newton, M., Chicago, Year Book Medical Publishers, 1977, pp. 511-520.
Scientific Foundations of Obstetrics and Gynaecology, 2nd Edition, edited by Philipp, E. E., Barnes, J., and Newton, M., Chicago Year Book Medical Publishers, 1977.
22.
23.
24.
25.
26.
31.
27.
28.
29.
30.
32.
Comfort, A.: The Biology of Senescence, 3rd Edition. New York, Elsevier North Holland, 1979, pp. 272-273.
. l . , t Leltevte, P., and Liebecq, C.: Metabolic effects of sulfur-containing radioprotective agents, in International Encyclopedia of Pharmacology and Therapeutics, Section 79, Sulfur-Containing Radioprotective Agents, edited by Bacq, Z. M., New York, Pergamon Press, 1975, pp. 205-281.
Bacq, Z. M., and Fischer, P.: The action of cysteamine on liver glycogen. Arch. Internat. Physiol., 61: 417-418, 1953.
Kawaic, K., Nakao, M., Nakao, T., and Katsui, G.: Inhibition of membrane adenosine tri- phosphatase by alpha-tocopherol and its derivatives. Amer. J. Clin. Nutr., 27: 978-994, 1974.
Jenkinson, E. J., and Billington, W. D.: Cell surface properties of early mammalian em- bryos, in Concepts in Mammalian Embryo- genesis, edi ted by Sherman, M. I., Cambridge, The MIT Press, 1977, pp. 235-266.
Yochim, J. M., and Mitchell, J. A.: Intra- uterine oxygen tension in the rat during pro- gestation: its possible relation to car- bohydrate metabolism and the regulation of nidation. Endicrinol., 83: 706-713, 1968.
Vital Statistics of the United States, 1973. Vol. II, Section 5. U. S. Depart. of Health, Education and Welfare. Public Health Ser- vice. Health Resources Administration. National Center for Health Statist ics, Rockville, Maryland, 1975.
Brinster, tivity in Biochim.
R. L: Lactate dehydrogenase ac- the preimplanted mouse embryo. Biophys. Acta, 110: 439-441, 1965.
Snell, G. D., and Stevens, L. C.: Early em- bryology, in Biology of the Laboratory Mouse, edited by Green, E. L., New York, McGraw-Hall, 1966, pp. 205-245.
Lewis, W. H., and Wright, E. S.: On the early development of the mouse egg. Contrib. Em- bryol. Carnegie Inst. Wash., 25: No. 148, 113-144, 1935.
Lubbers, D. W.: Quantitative measurement and description of oxygen supply to the tissue, in Oxygen and Physiological Func- tion, edited by Jobis, F. F., Dallas, Profes- sional Information Library, 1977, p. 254-275.
121

33.
34.
35.
36.
37.
Chapman, V. M., West, J. D., and Adler, D. A.: Gene t i c s of ear ly m a m m a l i a n em- bryogenesis, in Concepts in Mammalian Embryogenesis, edited by Sherman, M. I., Cambridge, the MIT Press, 1977, pp. 109-135.
Gartler, S. M., and Andrino, R. J.: Mammalian X-chromosome inactivation, in Adv. in Human Genetics, Vol. 7, edited by Harris, H., and Hirschhorn, K., New York, Plenum Press, 1976, pp. 99-140.
Epstein, C. J.: Expression of the mammalian X-chromosome before and after fertilization. Science, 175: 1467-1468, 1972.
Kirkman, H. N.: Glucose-6-phosphate dehy- drogenase, in Adv. in Human Genetics, Vol. 2, edited by Harris, H., and Hirschhorn, K., New York, Plenum Press, 1971, pp. 1-60.
Beutler, E.: Glutathione reductase, in Glutathione, edited by Flohe, L., Benohr, M. Ch., Sies, H., Waller, H. D., and Wendel, A., Stuttgart, Georg Theime, 1974, pp. 109-114.
38. Chapman, W. H., and Cronkite, E. P.: Further s tud ies on the benef ic ia l ef fect of glutathione on X-irradiated mice. Proc. Soc. Exptl. Biol. Med., 75: 318-322, 1950.
39. Flohe, L. Gunzler, W. A., and Ladenstein, R,: Glutathione peroxidase, in Glutathione: Metabolism and Function, edited by Arias, I. M., and Jakoby, W. B., New York, Raven Press, 1976, pp. 115-138.
40. Harman, D.: Free radical theory of aging: nutrit ional implications. Age, 1: 145-152, 1978.
41. Harman, D.: Free radical theory of aging: effect of free radical reaction inhibitors on the mortality rate of male LAF 1 mice. J. Gerontol., 23: 476-482, 1968.
42. Harman, D.: Free radical theory of aging: effect of the amount and degree of unsatura- tion of dietary fat on mortality rate. J. Geron- tol., 26: 451-457, 1971.
122