formation of bone tissue in culture from isolated bone...
TRANSCRIPT

FORMATION OF BONE TISSUE IN CULTURE
FROM ISOLATED BONE CELLS
ITZHAK BINDERMAN, DAN DUKSIN, ARIEH HARELL,EPHRAIM KATZIR (KATCHALSKI), and LEO SACHS
From the Hard Tissue Unit, Department of Endocrinology, Ichilov Medical Centre, Tel-Aviv, andthe Departments of Biophysics and Genetics, The Weizmann Institute of Science, Rehovot, Israel
INTRODUCTION
Various theories of mammalian bone calcificationhave been proposed, each of them based on bio-logical and biophysical phenomena's taking placein living bone (1) . Calcification in bone tissue ispreceded by the formation of an extracellularorganic matrix, primarily secreted by bone cells(2) . The bone-forming cells, active osteoblastsand osteocytes, control the synthesis of this extra-cellular matrix and regulate the exchange be-tween ions present in the bulk extracellular fluidsand those present in mineralizing bone collagen(3) .
Recent evidence established the role of bonecells in forming functional units that are vitallyconcerned with calcium homeostasis (4) . It istherefore of importance for the study of the bio-
ABSTRACTA system is described for the formation of bone tissue in culture from isolated rat bonecells . The isolated bone cells were obtained from embryonic rat calvarium and perios-teum or from traumatized, lifted periosteum of young rats . The cells were cultured for aperiod of up to 8 wk, during which time the morphological, biochemical, and functionalproperties of the cultures were studied . Formation of bone tissue by these isolated bonecells was shown, in that the cells demonstrated osteoblastic morphology in light andelectron microscopy, the collagen formed was similar to bone collagen, there was minerali-zation specific for bone, and the cells reacted to the hormone calcitonin by increased cal-cium ion uptake . Calcification of the fine structure of the cells and the matrix is described .Three stages in the calcification process were observed by electron microscopy . It is con-cluded that these bone cells growing in vitro are able to function in a way similar to suchcells in vivo . This tissue culture system starting from isolated bone cells is therefore suitablefor studies on the structure and function of bone .
chemical and biophysical aspects of living bone todevelop a model system of living osteoblasts inwhich differentiation, secretion of bone collagen,and calcification can proceed .
After the pioneering work of Fell (5), fewattempts have been reported in the literature togrow bone cells capable of forming bone collagenin tissue culture . Most of these studies were donein organ cultures in which whole bone or frag-ments of bones were cultured and were analyzedhistologically and biochemically (6, 7) . A methodfor culturing isolated bone cells was reported byRose (8), using a special multipurpose culturechamber. The osteogenic capacity of embryonicchicken fibroblasts in tissue culture was demon-strated by Fitton Jackson (9). Bard and his
THE JOURNAL OF CELL BIOLOGY . VOLUME 61, 1974 . pages 4 2 7 -439
427
on May 20, 2018jcb.rupress.org Downloaded from http://doi.org/10.1083/jcb.61.2.427Published Online: 1 May, 1974 | Supp Info:

coworkers (10) recently reported the isolationof bone cells from various sources . The cells,however, were kept for a short time only and didnot undergo calcification . At the same time, mor-phological and histochemical evidence suggeststhat the functional activity of bone cells is cor-related to their ultrastructure (11, 12) . In thesestudies both mineralized (13) and decalcified (14)bone specimens taken from living tissue havebeen used . Nevertheless, the various steps involvedin bone formation from the deposition of theextracellular matrix up to its calcification couldnot be investigated in detail due to many techni-cal obstacles (15, 16) .
In this paper, a new in vitro tissue culture sys-tem is introduced for the maintenance, growth,differentiation, and calcification of cells isolatedfrom bone and periosteum . The cultures areshown to contain functioning bone cells . Histo-chemical and biochemical data are presentedconfirming the formation of calcified bone tissuein the cultures . Morphological features of youngand old bone tissue in culture are described andcompared to the fine structure of bone in vivo . Apreliminary note on this work was published else-where (17) .
MATERIALS AND METHODS
Cells and Cell Cultures
Bone cells were obtained from three sources . (a)Calvaria, comprising the frontal, parietal, andoccipital bones, were removed from CR rat embryos(Charles River Breeding Laboratories, Inc ., Wil-mington, Mass .), 16-20 days after gestation, washedwell in phosphate-buffered saline (PBS) and separatedinto periosteum and bone proper. Each tissue wastreated separately . The separated tissues were mincedwell individually to give very small pieces . Thesewere incubated for 2 h at 37 °C in a 1 :1 mixture oftrypsin solution 0 .25% (1 :300, ICN Nutritional Bio-chemicals Div., International Chemical & NuclearCorp., Cleveland, Ohio) . 0 .137 M NaCl, 5.4 X 10 -3M KCI, 4.17 X 10-3 M NaHCO3, 5 .5 X 10-3 Mglucose, and a solution of disodium versenate (EDTA)(0 .02%), 0 .137 M NaCl, 2.7 X 10-3 M MC1, 8 .1 X10-3 M NaH2PO4, and 1.47 X 10-3 M KH2PO4.This solution was replaced every 30 min, and cellswere harvested by slow centrifugation (100 g) in aserological centrifuge (model 2070, Falcon Plastics,Div. Becton, Dickinson and Co ., Oxnard, Calif.) .The cells were counted by means of a hemocytom-eter; there were approximately 2 X 10 5 cells percalvarium and 10 6 cells per periosteum . Secondaryrat embryo cell cultures (18) served as a fibroblast
428
THE JOURNAL OF CELL BIOLOGY • VOLUME 61, 1974
control for the calvaria and periosteum cell cultures .(b) The periosteum was circumferentially peeledfrom 3-w old CR male rats, according to a methodpreviously described (19) . 24 h after the operationaltrauma, at a time when a veritable explosion of celldivision takes place, the traumatized tissue was re-moved, minced well, and incubated in the trypsin-EDTA mixture as described under (a) . Sham-operated animals were used as controls .
The cells obtained by both techniques were sus-pended in BGJ medium (Gibco formula, [GrandIsland Biological Co ., Grant Island, N . Y .] "peppedup" by Fitton Jackson's modification), supplementedwith 10%o fetal calf serum (FCS) (Gibco), counted,and inoculated in 60 X 15-mm plastic petri dishes(Falcon Plastics, Division of B-D Laboratories, LosAngeles, Calif .) . Seeding was at 2 X 10 4 cells perplate . The plates were incubated up to 8 wk at 37 °Cin moist 90% air and 10% CO2. The medium wasrenewed routinely every 3-4 days .
Histochemical MethodsSTAINING : Routine staining procedure was ac-
cording to Giemsa after methanol fixation .ALKALINE PHOSPHATASE : The Gomori Co-
balt method was used, which produced a black stain .The p-nitrophenyl phosphate-agar method (20) forthe detection of alkaline phosphatase-containing cellsin tissue culture was also employed .CALCIUM PHOSPHATE : Von Kossa's silver
method (21), which produced a black stain, wasused .ELECTRON MICROSCOPY : Pieces of tissue were
removed from the petri dishes with a scalpel andfixed with 3% glutaraldehyde (Schuchardt, Munich)for 60 min. The fixative was washed off with caco-dylate buffer (0.15 M, pH 7 .4), and the samples werestained with a 1 % solution of osmium tetroxide for60 min, dehydrated in alcohol and embedded inEpon. The sections were counterstained with satu-rated uranyl acetate solution for 30 min, and exam-ined in a JEOL Jem-T7 electron microscope .PHOTOGRAPHS : Phase-contrast and bright-field
micrographs were taken with a Zeiss phase-contrastinverted microscope, (Carl Zeiss, Inc., New York) onKodak Pan-X film (Eastman Kodak Co., Rochester,N. Y .) .
Calcitonin EffectThe plates used for the calcitonin (CT) experiments
(5 X 105 cells per plate) were washed three times withEarle's buffer solution (15 ml total) . 2 ml of Earle's so-lution containing 5 mg/% Ca++ and 10 5 cpm 45CaC12(Radiochemical Centre, Amersham, Great Britain),120 uCi/µg Ca++) were added, and the cultures wereincubated for 60 min at room temperature to reach aplateau in 45Ca++ uptake . Sterile lyophilized porcinecalcitonin, 10.25 metabolic clearance rate units/mg
(Armour Pharmaceutical Co ., Phoenix, Ariz .) wasadded to the culture, 0 .1 M metabolic clearance rateunits per plate. Samples were taken from the mediumfor counting at 5-min intervals for up to 60 min afterCT addition. Then the medium was removed and theculture was washed three times with Earle's buffersolution (15 ml total) . The culture was subsequentlydissolved in 1 ml 0.1 N NaOH and counted in aliquid scintillation spectrometer (Packard Tricarb,Packard Instrument Co., Inc ., Downers Grove, Ill .),in a dioxane-based scintillation liquid. The proteinconcentration was determined by the method ofLowry et al. (22) .
RESULTS
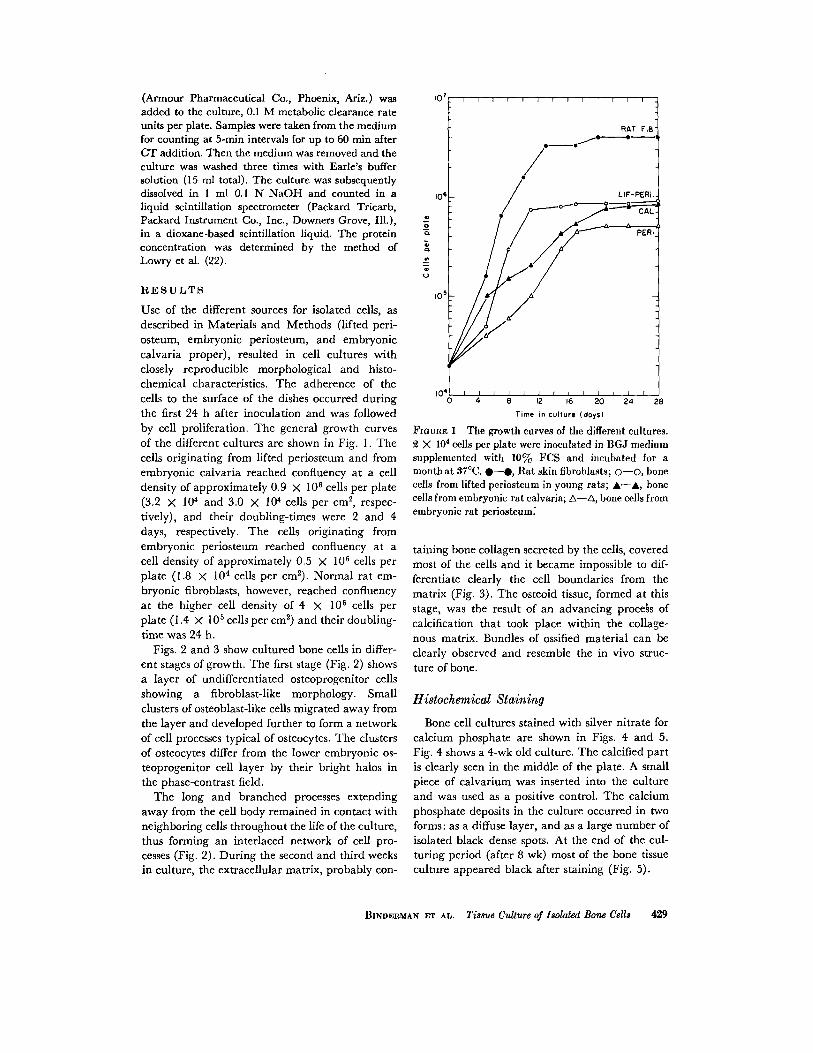
Use of the different sources for isolated cells, asdescribed in Materials and Methods (lifted peri-osteum, embryonic periosteum, and embryoniccalvaria proper), resulted in cell cultures withclosely reproducible morphological and histo-chemical characteristics . The adherence of thecells to the surface of the dishes occurred duringthe first 24 h after inoculation and was followedby cell proliferation . The general growth curvesof the different cultures are shown in Fig . I . Thecells originating from lifted periosteum and fromembryonic calvaria reached confluency at a celldensity of approximately 0 .9 X 10 6 cells per plate(3.2 X 10 4 and 3.0 X 104 cells per cm 2 , respec-tively), and their doubling-times were 2 and 4days, respectively . The cells originating fromembryonic periosteum reached confluency at acell density of approximately 0 .5 X 10 6 cells perplate (1 .8 X 104 cells per cm2) . Normal rat em-bryonic fibroblasts, however, reached confluencyat the higher cell density of 4 X 106 cells perplate (1 .4 X 105 cells per cm2 ) and their doubling-time was 24 h .
Figs. 2 and 3 show cultured bone cells in differ-ent stages of growth . The first stage (Fig . 2) showsa layer of undifferentiated osteoprogenitor cellsshowing a fibroblast-like morphology . Smallclusters of osteoblast-like cells migrated away fromthe layer and developed further to form a networkof cell processes typical of osteocytes . The clustersof osteocytes differ from the lower embryonic os-teoprogenitor cell layer by their bright halos inthe phase-contrast field .
The long and branched processes extendingaway from the cell body remained in contact withneighboring cells throughout the life of the culture,thus forming an interlaced network of cell pro-cesses (Fig . 2) . During the second and third weeksin culture, the extracellular matrix, probably con-
lo'_
BINDERMAN ET AL . Tissue Culture of Isolated Bone Cells
RAT Fi8 -
0
4
8
12
16
20
24
28Time in culture (doys)
FIOUUE 1 The growth curves of the different cultures.2 X 104 cells per plate were inoculated in BGJ mediumsupplemented with 10% FCS and incubated for amonth at 37 °C. •- 0, Rat skin fibroblasts ; O-O, bonecells from lifted periosteum in young rats ; A-A, bonecells from embryonic rat calvaria; A-A, bone cells fromembryonic rat periosteum :
taining bone collagen secreted by the cells, coveredmost of the cells and it became impossible to dif-ferentiate clearly the cell boundaries from thematrix (Fig. 3) . The osteoid tissue, formed at thisstage, was the result of an advancing process ofcalcification that took place within the collage-nous matrix . Bundles of ossified material can beclearly observed and resemble the in vivo struc-ture of bone .
Histochemical Staining
Bone cell cultures stained with silver nitrate forcalcium phosphate are shown in Figs . 4 and 5 .Fig. 4 shows a 4-wk old culture . The calcified partis clearly seen in the middle of the plate . A smallpiece of calvarium was inserted into the cultureand was used as a positive control. The calciumphosphate deposits in the culture occurred in twoforms : as a diffuse layer, and as a large number ofisolated black dense spots . At the end of the cul-turing period (after 8 wk) most of the bone tissueculture appeared black after staining (Fig. 5) .
429
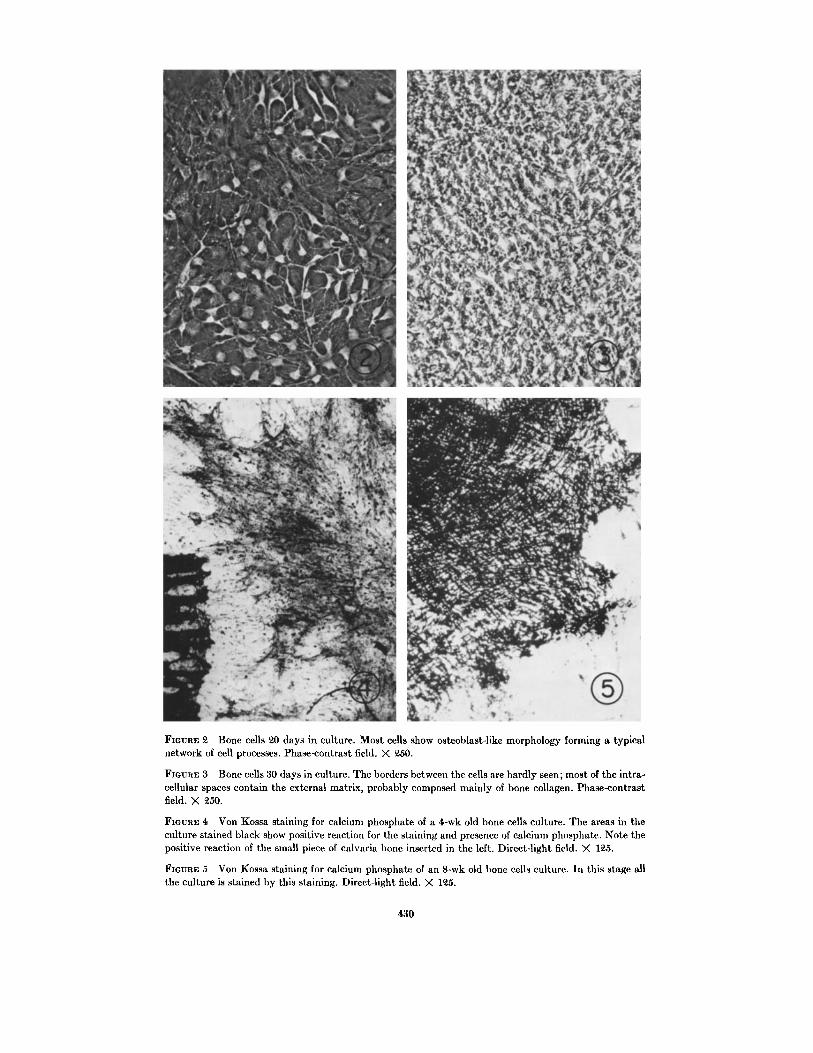
FIGURE 2 Bone cells 20 days in culture . Most cells show osteoblast-like morphology forming a typicalnetwork of cell processes . Phase-contrast field . X 250 .
FIGURE 3 Bone cells 30 days in culture . The borders between the cells are hardly seen ; most of the intra-cellular spaces contain the external matrix, probably composed mainly of bone collagen . Phase-contrastfield. X 250.
FIGURE 4 Von Kossa staining for calcium phosphate of a 4-wk old bone cells culture . The areas in theculture stained black show positive reaction for the staining and presence of calcium phosphate . Note thepositive reaction of the small piece of calvaria bone inserted in the left . Direct-light field . X 125 .
FIGURE 5 Von Kossa staining for calcium phosphate of an 8-wk old bone cells culture . In this stage allthe culture is stained by this staining . Direct-light field . X 125 .
430

Osteoblasts are known to contain an elevatedamount of alkaline phosphatase (23) . Most of theenzymatic activity is associated with the cell mem-branes (23, 24). For this reason histochemicalstaining for alkaline phosphatase was also per-formed. Alkaline phosphatase activity was foundto be located at the surface of the bone cells, asshown by their black envelope . A different stain-ing procedure for alkaline phosphatase, based onthe agar method of Maio and coworkers (20), wasalso used . In the bone cell cultures the agar wasstained by the yellow derivative of p-nitrophenol,establishing the presence of alkaline phosphataseactivity in the bone cell culture . Only negligiblestaining occurred in rat fibroblast culture .
The Effect of Calcitonin
Calcium uptake by cultured bone cells, asmeasured by 45Ca incorporation, was found to befour times as high as by skin fibroblasts under thesame conditions . CT added to bone cell culturescaused a significant rise in 45Ca uptake by thesecells. The hormone, however, had no effect on thecultured rat fibroblasts (Table I) .
Electron Microscopy
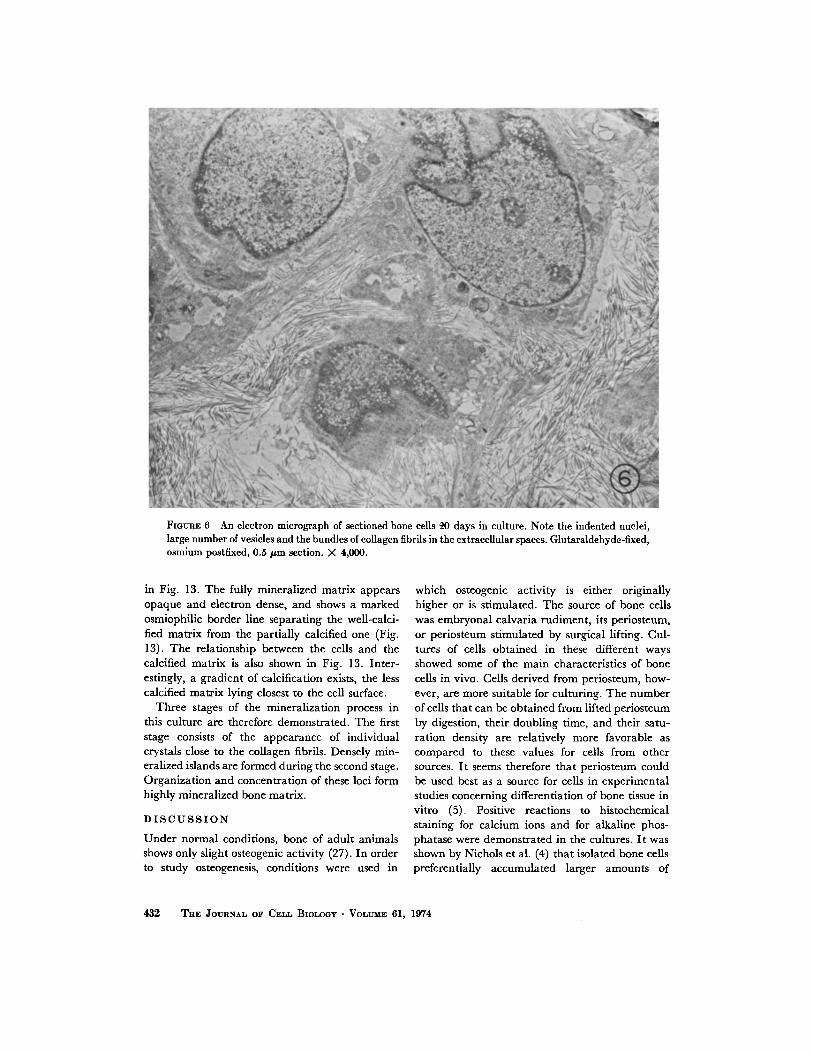
The fine structure studies of the culture weredirected in particular to the process of mineraliza-tion . Results are presented concerning the ultra-structure of the cultured bone cells before andafter the occurrence of calcification .UNCALCIFIED YOUNG CULTURE : During
the differentiation period, osteoblasts surroundedby fine collagen fibers are characteristic of bonetissue formation in culture . A striking feature of ayoung culture (1-3 wk) is the occurrence of os-teoblasts in clusters . In the 3-wk culture (Figs. 6and 7) numerous active osteoblasts are visible . Inthe cells large indented nuclei with well-definedmembranes are seen . Fig . 7 shows a typical cultureof such young bone cells. The cells within a clustershow various structural features at their sites ofcontact. The cell membrane surfaces are eitherclosely apposed or separated from each other byextracellular matrix spaces of variable widthwhich contain mainly short and longer collagenfibrils . The cytoplasm is rich in ribosomes bothfree, and bound to a well-developed rough endo-plasmic reticulum in large mitochondria and inmembranous vesicular structures of the Golgi com-plex (Fig. 7). The latter structures indicate anactive secretory function . The cytoplasm some-
TABLE IThe Effect of Calcitonin on 45Ca Uptake by Bone
Cells and Fibroblasts in Culture
IbCa uptake cpm/mg protein X 10-3CT
mU MRC/plate
Bone cello
Skin fibroblasts
-
4.8 f 0.3
1 .2 f 0 .30 .1
7 .4 f 0.3
1 .3 f 0 .2
times contained numerous myelin-like figures alsotermed lamellar body membranes (25, 26) . Theselaminated membranous cytoplasmic bodies occurin the near vicinity of the mitochondria (Figs . 6-8and 10) . The lamellae composed of electron-dense, layered membranes are arranged eitherconcentrically, as shown in cross sections (Figs . 7and 8), or in parallel, as shown in longitudinalsections (Figs. 8 and 10). These lamellar structuresare known to exist in a variety of cells such as lunggreat alveolar cells (25) or neuronal cells (26) .Their appearance in bone cells, however, hasnot been previously reported .
The extracellular matrix of the young culturecontains unmineralized thin collagen fibrils vary-ing in size and diameter (Figs. 6, 7, 9, and 10) .Some adjacent fibrils are registered to accuratelythat their banding pattern appears continuous(Fig . 9) .Noncalcified matrix contains vesicular struc-
tures and amorphous-looking materials which maybe composed of mucoproteins or glycoproteins(Fig. 10), and which may play an important rolein the calcification process .CALCIFYING CULTURE : Mineralized ma-
trix appears at later stages (8 wk) . The depositionof crystals of calcium phosphate in the extracel-lular matrix of isolated cultured bone cells stronglysupports our contention that it is possible to ob-tain bone formation in this system, Although cal-cification of the matrix usually occurs in the nearvicinity of the collagen fibrils (Figs . 11 and 12), norelationship between the position of the needle-shaped apatite crystals and the collagen bandingpattern was visible . Careful inspection of Fig . 12reveals the presence of vesicle-like structuresrandomly dispersed in the collagen matrix con-taining some apatite crystals . Further calcificationof the matrix may occur by continued mineraldeposition in these structures . Eventually, massivecalcification occurs also between these structures,giving rise to large mineralized areas within thematrix. These subsequent stages can be observed
BINDERMAN ET AL . Tissue Culture of Isolated Bone Cells 431
in Fig. 13 . The fully mineralized matrix appearsopaque and electron dense, and shows a markedosmiophilic border line separating the well-calci-fied matrix from the partially calcified one (Fig .13) . The relationship between the cells and thecalcified matrix is also shown in Fig . 13. Inter-estingly, a gradient of calcification exists, the lesscalcified matrix lying closest to the cell surface .
Three stages of the mineralization process inthis culture are therefore demonstrated . The firststage consists of the appearance of individualcrystals close to the collagen fibrils . Densely min-eralized islands are formed during the second stage .Organization and concentration of these loci formhighly mineralized bone matrix .
DISCUSSION
Under normal conditions, bone of adult animalsshows only slight osteogenic activity (27) . In orderto study osteogenesis, conditions were used in
432
FIGURE 6 An electron micrograph of sectioned bone cells 4,0 days in culture . Note the indented nuclei,large number of vesicles and the bundles of collagen fibrils in the extracellular spaces . Glutaraldehyde-fixed,osmium postfixed, 0 .5 µm section . X 4,000.
THE JOURNAL OF CELL BxoLoGY • VOLUME 61, 1974
which osteogenic activity is either originallyhigher or is stimulated . The source of bone cellswas embryonal calvaria rudiment, its periosteum,or periosteum stimulated by surgical lifting. Cul-tures of cells obtained in these different waysshowed some of the main characteristics of bonecells in vivo . Cells derived from periosteum, how-ever, are more suitable for culturing. The numberof cells that can be obtained from lifted periosteumby digestion, their doubling time, and their satu-ration density are relatively more favorable ascompared to these values for cells from othersources . It seems therefore that periosteum couldbe used best as a source for cells in experimentalstudies concerning differentiation of bone tissue invitro (5) . Positive reactions to histochemicalstaining for calcium ions and for alkaline phos-phatase were demonstrated in the cultures . It wasshown by Nichols et al . (4) that isolated bone cellspreferentially accumulated larger amounts of

FIGURE 7 Young bone cells 3 wk in culture. Cells show indented nuclei N, Golgi apparatus GA, abundantendoplasmic reticulum ER, vacuoles Ve, and myelin-like figures MP. The matrix contains collagen fibrilsCo. Glutaraldehyde-fixed, osmium-postfixed, 0 .5 µm section . X 30,000 .
BINDERMAN ET AL. Ti33ue Culture of Isolated Bone Cells
433
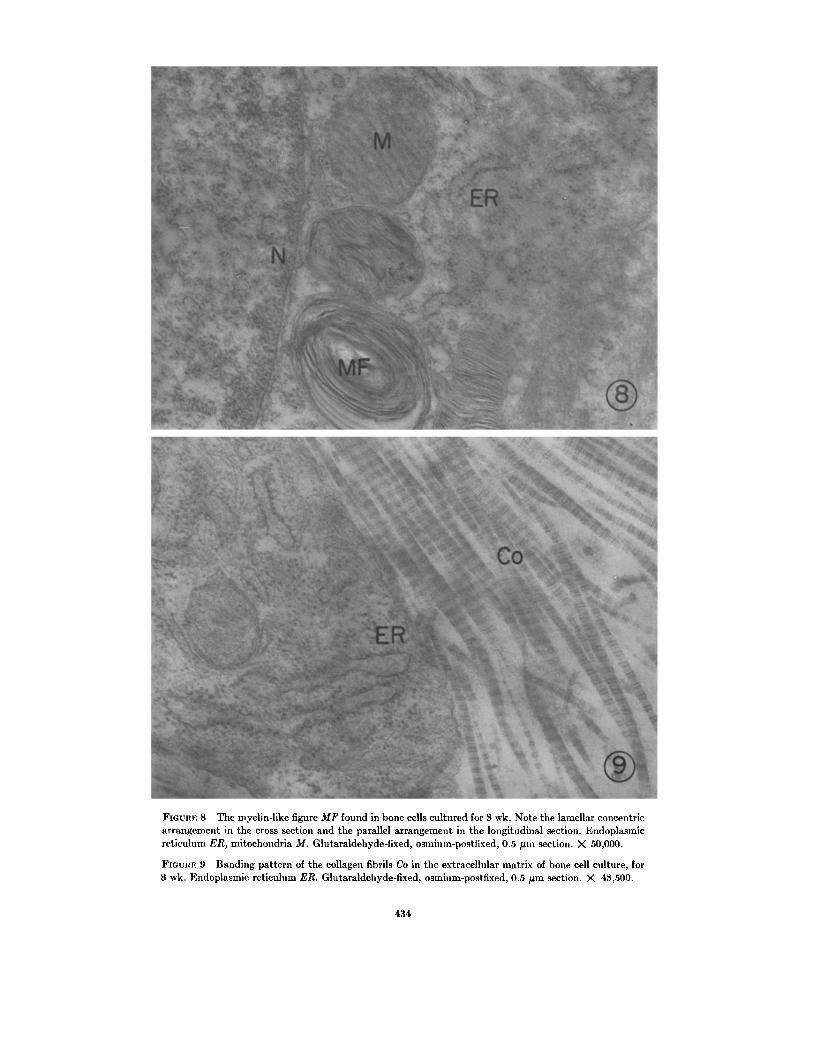
FIGURE 8 The myelin-like figure MF found in bone cells cultured for 3 wk. Note the lamellar concentricarrangement in the cross section and the parallel arrangement in the longitudinal section . Endoplasmicreticulum ER, mitochondria M. Glutaraldehyde-fixed, osmium-postfixed, 0 .5 µm section . X 50,000 .
FIGURE 9 Banding pattern of the collagen fibrils Co in the extracellular matrix of bone cell culture, for3 wk. Endoplasmic reticulum ER . Glutaraldehyde-fixed, osmium-postfixed, 0 .5 µm section . X 43,500.
4 34

FIGURE 10 Young bone cell 3 wk in culture . Note the indented nucleus N, myelin-like figures MF, andlong mitochondria M. The extracellular matrix contains collagen fibrils Co and vesicles V. Glutaraldehyde-fixed, osmium-postfixed, 0 .5 µm section . X 33,600.
calcium than do other cells in the organism .Moreover, bone cells have an increased calciumuptake when incubated in the presence of calci-tonin (28) . Our technique, by which isolated (28)and cultured bone cells react similarly and exhibit
an increase in calcium uptake in response to calci-tonin, may serve as a significant method for theidentification of metabolically active bone cells .
This method permits observation in culturedishes of the different stages of calcification of theextracellular matrix, and makes it possible to com-pare this process to bone formation in vivo .
The study of bone tissue formation in cell cul-tures has several advantages. a Bone specimensprepared for electron microscopy are often sub-jected to prolonged decalcification (4), which isnot necessary for the cultures . b The vital proper-ties of the cells are more readily conserved in tissueculture than in whole bone tissue, since the fixa-tives are more accessible to the thin culture speci-mens. c Bone tissue formation can easily be fol-lowed step by step .
The overall picture of cultured bone cells andthe matrix is similar to that of the bone tissue de-scribed in in vivo studies (14-16) . In the earlystages, short collagen fibrils are observed adjacentto, or merging with, the cell surface of the osteo-
blasts . Similarly, such structures were found alsoin embryonic fowl osteoblast cultures (29) . Well-developed rough endoplasmic reticulum, numer-ous mitochondria, a vesicle-rich Golgi apparatus(30), and both microtubules and microfilamentsin the inner parts of the cell surface indicate thatthese cells actively synthesize matrix components .Recent biochemical findings (31-33) suggest
that transformation of procollagen into collagenoccurs at the cell membrane, and that the secretedcollagen aggregates into fibrils immediately aftersecretion through the cell membrane . The mor-phological appearance of cultured bone cells andits matrix support this suggestion .
In the early stages of bone tissue formation inour culture, fine mineral crystallites are presentembedded in an electron-dense, granular material
BINDERMAN ET AL . Tissue Culture of Isolated Bone Cells 435

FIGURE 11 Mineralized collagen fibrils from a six-wk old bone cell culture . Glutaraldehyde-fixed, os-mium-postfixed 0.5 µm section . X 27,800 .
FIGURE 12 Extracellular matrix of bone cells cultured for 8 wk. Partially mineralized collagen fibrils,Co, and vesicles, v. Glutaraldehyde-fixed, osmium-postfixed, 0 .5 µm section . X80,600 .
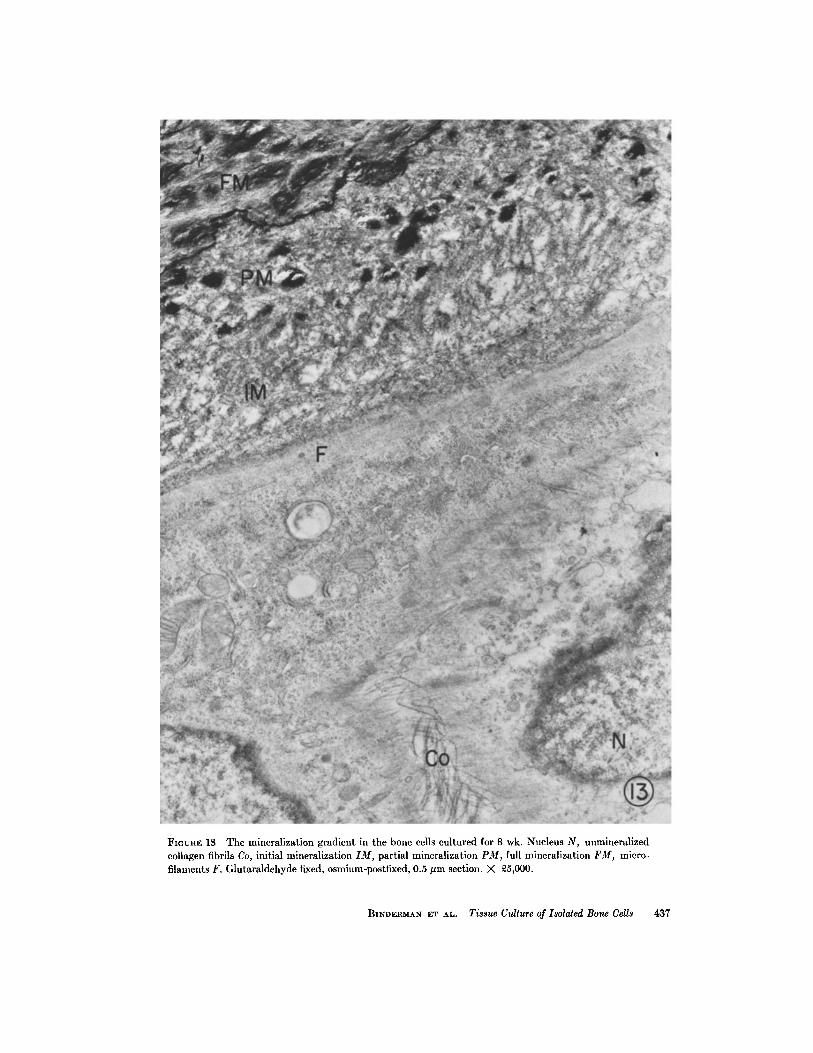
FIGURE 13 The mineralization gradient in the bone cells cultured for 8 wk . Nucleus N, unmineralizedcollagen fibrils Co, initial mineralization IM, partial mineralization PM, full mineralization FM, micro-filaments F. Glutaraldehyde fixed, osmium-postfixed, 0 .5 µm section . X 25,000 .
BINDERMAN ET AL . Tissue Culture of Isolated Bone Cells
437

coating small groups of collagen fibrils . This ob-servation is in agreement with the results of pre-vious studies showing that the collagenous matrixis the site for deposition of calcium phosphatecrystals . This has been demonstrated in culturedembryonic fowl bone (34), in rachitic osteoid tis-sue healing in vivo (35), in calcifying turkey legtendons (36), in reconstituted collagen fibrils cal-cifying in vitro (37, 38), and in demineralized bonematrix calcifying in a metastable buffer (39) .
While apatite crystals appear to be arrangedalong the collagen fibers, a close morphologicalrelationship between the collagen banding-patternand the position and orientation of the crystals
was not apparent. Such findings were also re-ported elsewhere (16) .
Further calcification leads to patches of mineraldeposition with no definite relation to collagenfibers . These patches seem to be connected withvesicle-like structures, which may be similar to thenoncollagenous bone nodules described elsewhere(40,41) .During the last stage of bone development in
culture, fully calcified areas appear . These areasare usually bordered by a distinct osmiophilic line .A gradient of calcification exists, the fully calcifiedmatrix being furthest removed from the bone cells .There is no doubt, however, that the bone cells
exert some control over the mineralization of thematrix surrounding them . This has been sug-gested to be the case during the initiation of the
mineralization process (42) . At this stage it is notknown whether the bone cells also actively par-ticipate in the massive calcification, or this processoccurs by a physicochemical mechanism of crystalnucleation and growth .
Chemical analysis' of the collagen producedby bone cells in culture showed that the chemicalcomposition of this collagen with respect to alde-hydes and cross-links is similar to that of collagenproduced by bone tissues in vivo (43) .
Our system for culturing bone cells appears tobe a successful attempt to grow bone tissue in vitrostarting from isolated bone cells . Formation of bonetissue by these cells is indicated by the followingfindings : (a) the cells show osteoblastic morphologyin light and electron microscopy ; (b) the collagenformed is similar to bone collagen ; (c) Specificmineralization occurs in the culture, and themineral stains by the von Kossa method; (d) The
I G. Mechanic, personal communication .
438 THE JOURNAL OF CELL BIOLOGY • VOLUME 61, 1974
cells react to calcitonin hormone and take upmore calcium ions in the presence of the hormone .
The introduction of this tissue culture systemfor investigation of the differentiation, growth, andfunction of bone cells enables us to study the roleof the cells and their constituents in the differentstages of bone formation, and the influence ofhormones and other factors in this process
We thank Dr. B. N. Bachra, University of Leiden,Netherlands for stimulating discussions and for hiscritical reading of the manuscript. We are gratefulto Mrs. Brita Hardy, Department of BiologicalUltrastructure, The Weizmann Institute of Science,for the preparation of the electron micrographs,and to Mrs. Aliza Treves for her excellent andcapable technical assistance.
Received for publication 25 May 1973, and in revisedform 11 December 1973 .
REFERENCES
1 . BACHRA, B. N. 1970 Int. Rev. Connect. Tissue Res.5 :165 .
BERNARD, G. W. 1971 . In Cellular and Mo-lecular Renewal in the Mammalian Body . I . L .Cameron, and J . D. Thrasher, editors . Aca-demic Press, Inc., New York . 373.
3. RASMUSSEN, H., J . FEINBLATT, N. NAGATA, andM. PECHET. 1970. Fed. Proc. 29 :1190.
4. NICHOLS, G . JR ., P. HIRSCHMANN, and P . ROGERS.1971 . In Cellular Mechanisms for CalciumTransfer and Homeostasis . G. Nichols, Jr .,and R. H. Wasserman, editors. AcademicPress, Inc., New York . 211 .
5 . FELL, H . B . 1932 . J. Anat. 66:157 .6 . RAISZ, L . G., and I. NIEMANN . 1969 . Endocri-
nology . 85:446.7 . GOLDHABER, P. 1958. Arch. Pathol. 66:635 .8. ROSE, G. G., and T. O. SHINDLER . 1964. Tex.
Rep. Biol. Med. 22 :174 .9 . FITTON-JACKSON, S . 1965 . In Structure and
Function of Connective and Skeletal Tissue.Butterworth & Co. (Publishers) Ltd ., London277 .
10. BARD D. R., M. T. DICKENS, A. V. SMITH, andJ. M. ZAREK, 1972. Nature (Lond.) . 236:314.
11 . DOTY, S . B., and B. H. SCHOFIELD. 1972 .Histochem . J . 4:245 .
12. HOLTROP, M. E. 1972 . Calcif. Tissue Res . 9:140.13 . SCOTT, B. L., and D. C. PEASE. 1956 . Anta . Rec .
126:465.14. WASSERMANN, F., and J. A. YAEGER . 1965. Z.
Zellforsch . Mikrosk . Anat. 67 :636 .15. DUDLEY, H. R., and D. SPIRO. 1961 . J. Biophys.
Biochem. Cytol . 11 :627 .
2 .

16 . CAMERON, D. A. 1972 . In The Biochemistry andPhysiology of Bone. G. H. Bourne, editor .Academic, Press Inc ., New York . 1 :191 .
17 . BINDERMAN, I . D ., DUKSIN, L . SACHS, and A .HARELL . 1972 . Abstr. IVth InternationalCongress Endocrinology 256 :181 .
18 . YOUNGNER, J. S. 1954 . Proc . Soc . Exp. Biol. Med.85:202 .
19 . PIATIER-PIKETTY, D., and J . ZUCMAN . 1971 .Isr. J. Med. Sci. 7 :447.
20 . MAIO, J. J ., and L. L. DE CARLI . 1962 . Nature,(Lond. ) . 196:600 .
21 . KossA, J. V. 1901 . Beitr . Pathol. Anat . Allg .Pathol . 29 :163 .
22 . LOWRY, O . H., N. J . RosEBROUGH, A. L. FARR,and R . J . RANDALL . 1951 . J. Biol. Chem .193 :265 .
23 . HEKKELMAN, J. W ., and A. MOSKALEWSKI . 1969 .Exp . Cell Res. 58 :283 .
24 . YOSHIKI, S., T. UMEDA, and Y . KURAHASHI .1972 . Histochemie . 29 :296 .
25 . SMITH, D. S ., U . SMITH, and J . W . RYAN . 1972 .Tissue Cell. 4 :457 .
26. BRADY, R . O ., and E. H. KOLODNY . 1972 . Prog .Med. Genet. 8 :225 .
27 . PRITCHARD, J . J . 1972. In The Biochemistry andPhysiology of Bone. G. H. Bourne, editor .Academic Press, Inc., New York . 1 .
28 . HARELL, A ., I . BINDERMAN, and G . A. RODAN .1973 . Endocrinology . 92 :550.
29 . FITTON JACKSON, S. 1960 . In Bone as a Tissue .K. Rodahl, J . T. Nicholson, and E. M . Brown,editors. McGraw-Hill Book Company, NewYork . 165 .
30. ROSE, G. G. 1961 . J. Biophys. Biochem . Cytol .9 :463 .
31 . DIEGELMANN, R . F ., and B. PETERKOFSKY . 1972 .Proc . Natl . Acad. Sci . U. S. A . 69 :892 .
32 . EHRLICH, H . P ., and P . BORNSTEIN . 1972 . Nat .New Biol . 238 :257 .
33 . PIEZ, K. A . 1972 . In Current Topics in Bio-chemistry, C. B. Anfinsen, R. F. Goldberger,and A. N. Schechter, editors, Academic Press,Inc., New York . 101 .
34. FITTON-JACKSON, S ., and J. T . RANDALL . 1956 .In Bone Structure and Metabolism . G. E. W.Wolstenholme and C . M. O'Connor, editors.Churchill Ltd. London . Ciba Found . Symp . 47 .
35. ROBINSON, R. A., and H . SHELDON . 1960 . Cal-cification in Biological Systems . Am. Assoc .Ado. Sci . Pub[. 64 :261 .
36 . NYLEN, M . U., SCOTT, D . B., and U . M. MOSLEY .1960 . Calcification in Biological Systems . Am .Assoc. Adv . Sci. Publi. 64:129 .
37 . GLIMCHER, M. J . 1959. Rev . Mod . Phys . 31 :359 .38 . GLIMCHER, M. J . 1960. Calcification in Bio-
logical Systems . Am. Assoc. Adv . Sci . Publ .64 :421 .
39 . BACHAA, B. N. 1972 Calcif. Tissue Res . 8 :287 .40 . BERNARD, G . W., and D. C . PEASE . 1969. Am . J.
Anat . 125 :271 .41 . BERNARD, G . W. 1971 . In Cellular and Mo-
lecular Renewal in the Mammalian Body .I . L . Cameron, and J. D . Thrasher, editors .Academic Press, Inc ., New York . 373 .
42 . STEENDIJK, R ., and A. BOYDE . 1973. Calcif.Tissue Res . 11 :242 .
43 . MECHANIC, G ., P . M. GALLOP, and M. L. TAN-ZER . Biochem . Biophys. Res. Commun . 45 :644 .
BINDERMAN ET AL . Tissue Culture of Isolated Bone Cells
439