food derived from glyphosate tolerant … a416... · 5 cp4 epsps is present in low concentrations...
TRANSCRIPT

FOOD DERIVED FROM GLYPHOSATE TOLERANT
CORN LINE NK603
A Safety Assessment
TECHNICAL REPORT SERIES NO. 25
FOOD STANDARDS AUSTRALIA NEW ZEALAND
May 2003 © Food Standards Australia New Zealand 2003 ISBN 0 642 34550 3 ISSN 1448-3017 Published June 2003 This work is copyright. Apart from any use as permitted under the Copyright Act 1968, no part may be reproduced by any process without prior written permission from Food Standards Australia New Zealand Food (FSANZ). Requests and inquiries concerning reproduction and rights should be addressed to the Information Officer, Food Standards Australia New Zealand, PO Box 7168, Canberra BC, ACT 2610. An electronic version of this work is available on the Food Standards Australia New Zealand (FSANZ) website at http://www.foodstandards.gov.au . This electronic version may be downloaded, displayed, printed and reproduced in unaltered form only for your personal, non-commercial use or use within your organisation. Food Standards Australia New Zealand Australia New Zealand PO Box 7186 PO Box 10599 Canberra BC ACT 2610 Wellington Australia New Zealand Tel +61 2 6271 2241 Tel +64 4 473 9942 Fax +61 2 6271 2278 Fax +64 4 473 9855 Email [email protected] Email [email protected]

2

3
TABLE OF CONTENTS
SUMMARY ..............................................................................................................................4
BACKGROUND ......................................................................................................................6
HISTORY OF USE..................................................................................................................6
DESCRIPTION OF THE GENETIC MODIFICATION....................................................7 Methods used in the genetic modification .............................................................................7 Function and regulation of novel genes .................................................................................7 Characterisation of the genes in the plant ..............................................................................9 Stability of genetic changes .................................................................................................11 Impact on human health from the potential transfer of novel genetic material to cells of the human digestive tract ...........................................................................................................11 Conclusion ...........................................................................................................................12
CHARACTERISATION OF NOVEL PROTEIN..............................................................13 Biochemical function and phenotypic effects......................................................................13 Protein expression analyses .................................................................................................14 Potential toxicity of novel protein........................................................................................16 Potential allergenicity of novel protein................................................................................18 Conclusion ...........................................................................................................................20
COMPARATIVE ANALYSES.............................................................................................20 Nutrient analyis....................................................................................................................21 Levels of anti-nutrients ........................................................................................................27 Ability to support typical growth and well-being ................................................................27 Conclusion ...........................................................................................................................31
ACKNOWLEDGEMENTS ..................................................................................................31
REFERENCES.......................................................................................................................32

4
SUMMARY Glyphosate-tolerant corn line NK603 has been developed primarily for agricultural purposes to provide growers with an additional variety of corn that has been engineered for tolerance to the broad spectrum herbicide, glyphosate. Food derived from this GM corn line has been evaluated to determine its suitability for human consumption. The evaluation criteria included characterisation of the transferred genes, analysis of changes at the DNA, protein and whole food levels, and assessment of the potential allergenicity and toxicity of any newly expressed proteins. Examination of these criteria has enabled both the intended and unintended changes to be identified, characterised and evaluated for safety. Nature of the genetic modification In corn line NK603, the glyphosate-tolerance trait has been introduced by the addition of a bacterial gene encoding the EPSPS protein, a key enzyme in the biosynthesis of aromatic amino acids in plants and microbes. The mode of action of glyphosate is to bind to the plant EPSPS protein, thereby impairing its normal enzyme activity, subsequently resulting in plant cell death. The bacterial form of the enzyme (denoted as CP4 EPSPS) has a lower affinity for glyphosate, so that when present in plant cells, the activity of the introduced enzyme replaces the sensitive plant EPSPS enzyme. The result is that the engineered plant is able to function in the presence of the herbicide. Line NK603 contains two linked copies of the CP4 EPSPS gene, each with separate regulatory sequences. One copy is expressed from the rice actin promoter and intron while the second is expressed from the enhanced cauliflower mosaic virus promoter, which have both been shown to direct constitutive protein expression in corn. Additional regulatory sequences in common include an optimised chloroplast transit peptide sequence, to direct translocation of the CP4 EPSPS protein to chloroplasts where the protein is functionally active, and a NOS 3’ untranslated region providing the appropriate eukaryotic polyadenylation signal. Because a purified segment of DNA was used in the transformation, no extraneous bacterial genes, including laboratory marker genes, were transferred. History of use Corn has been cultivated for centuries and has been safely consumed as food and feed for thousands of years. It is a staple food for a significant proportion of the world’s population. Sweet corn varieties are grown for human consumption and corn-based products are routinely used in a wide range of foods. Grain and by-products from processing of corn are also used as animal feedstuffs. Characterisation of novel protein The novel protein expressed in corn line NK603 is CP4 EPSPS and the close sequence variant CP4 EPSPS L214P, which differs from CP4 EPSPS by one amino acid. The CP4 EPSPS is a bacterial form of an enzyme already naturally occurring in all plants including corn. This enzyme is involved in amino acid biosynthesis and the bacterial enzyme, unlike the plant enzyme, is able to function in the presence of the herbicide glyphosate. The CP4 EPSPS has been completely sequenced and encodes a 47.6 kDa protein consisting of a single polypeptide of 455 amino acids (Padgette et al., 1996). The deduced amino acid sequence of the CP4 EPSPS with the CTP2 transit peptide (amino acids 1-76) was also assessed. The characterisation of the CP4 EPSPS and variant CP4 EPSPS L214P included an examination of the three dimensional folding patterns of the enzyme and amino acid sequence homology at the active site enabling comparision with the structure and function of the native corn EPSPS. The bacterial enzyme exhibits approximately 50% amino acid sequence similarity with plant EPSPS enzymes (eg. soybean, corn and petunia).

5
CP4 EPSPS is present in low concentrations in the grain and at higher concentrations in the forage of corn line NK603. A higher level of novel protein expression in the green tissues of the plant (corresponding to the forage) is consistent with the functional rice actin and viral promoters used in the gene constructs. Although the level of expression is low, it is sufficient to confer tolerance to glyphosate at the level of the whole plant. The mean CP4 EPSPS protein levels in NK603 grain were comparable at the non-replicated sites (11.0 µg/g fw) and the replicated sites (10.6 µg/g fw) indicating that the novel protein is expressed at approximately the same levels either within a site or across geographically dispersed sites. Comparative analyses The results of extensive compositional analyses on glyphosate-treated plants grown at multiple locations demonstrate that the levels of the important components in NK603 corn grain (protein, total fat, carbohydrate, ash, fibre, fatty acids, amino acids, minerals and moisture) are not different from the non-transformed parental line. In addition, analyses for Vitamin E, phytic acid and trypsin inhibitor confirmed that the modification has not resulted in any variation to these minor components. Statistical analysis of the results for fatty acids and amino acids showed that some minor differences between the transformed line and non-transformed control line occurred at one or two of the trial sites. However, the nature of the differences was not consistent across all sites in the two major studies and therefore the differences were considered to reflect random variation that is characteristic of large-scale plant analyses. Moreover, all compositional results from the transformed line were well within the ranges observed for commercial non-transformed lines for each of the parameters investigated. Corn line NK603 was also shown to be equivalent to its non-transformed counterpart in the ability to support typical growth and well-being in animal feeding studies where grain from test and control lines was included in the diet of rapidly growing broiler chickens and grower-finisher swine, two commercially produced species. In addition, a 13 week feeding study in laboratory rats comparing diets containing NK603 corn with several non-transformed corn varieties did not identify any treatment –related changes in clinical parameters or tissue pathology. These studies support the conclusion that the genetic modification has not resulted in changes in the nutritional qualities of corn line NK603 when compared to other commercial varieties. Conclusion No potential public health and safety concerns have been identified in the assessment of glyphosate tolerant corn line NK603. On the basis of the data assessed, herbicide-tolerant corn line NK603 is equivalent to other commercially available corn in terms of its safety and nutritional adequacy.

6
FOOD DERIVED FROM GLYPHOSATE TOLERANT CORN LINE NK603:
A SAFETY ASSESSMENT
BACKGROUND A safety assessment has been conducted on food derived from corn that has been genetically modified to be tolerant to the herbicide glyphosate. The modified corn is referred to as glyphosate tolerant corn line NK603. Herbicide tolerance is achieved through expression in the plant of the bacterial gene encoding the EPSPS protein, a key enzyme in the biosynthesis of aromatic amino acids in plants and microbes. Glyphosate directly affects the shikimate biosynthetic pathway in plants. The mode of action of glyphosate is to specifically bind to and block the activity of 5-enolpyruvylshikimate-3-phosphate synthase (EPSPS), an essential enzyme involved in the biosynthesis of aromatic amino acids in all plants, bacteria and fungi. Blocking the enzyme results in the breakdown of the synthesis of essential aromatic amino acids in cells, ultimately leading to the death of the plant. However, the enzyme produced from the introduced epsps gene has a reduced affinity for the herbicide compared with the corn EPSPS enzyme, and thus imparts glyphosate tolerance to the whole plant. The bacterial CP4 EPSPS is used also in glyphosate tolerant varieties of soybean, canola, sugar beet and cotton. Foods derived from these modified crop lines have previously been assessed for safety by FSANZ and have been approved for food use in Australia and New Zealand under Standard 1.5.2 – Food Produced Using Gene Technology of the Australian New Zealand Food Standards Code (the Code). Corn is used predominantly as an ingredient in the manufacture of breakfast cereals, baking products, extruded confectionery and corn chips. Maize starch is used extensively by the food industry for the manufacture of many processed foods including dessert mixes and canned foods. Despite the diverse uses of corn products in many foods, corn is a relatively minor crop in both Australia and New Zealand, with a declining area planted over the last decade. Consequently, there is a requirement to import products such as high-fructose corn syrup and maize starch to meet manufacturing demand. The glyphosate-tolerance trait has not been introduced into sweet corn or popcorn varieties and therefore the whole kernel from corn line NK603 is not consumed directly as food, but rather is processed into various corn fractions. HISTORY OF USE Corn (Zea mays L., also called maize) has a long history of safe use as a food for both humans and other animals. Being the only important cereal crop indigenous to North America, it has been utilised for thousands of years and was the foundation of the extensive North and South American ancient civilisations. Corn seed was carried to Europe centuries ago, where it became established as an important crop in southern latitudes, moving rapidly to Africa, Asia and other parts of the world. In countries where corn is an important crop, it is the principal component of livestock feeds, and most of it is fed to farm animals, particularly to ruminants. In only a few countries is corn a major constituent of human diets. In developed countries, corn is consumed mainly as popcorn, sweet corn, corn snack foods and occasionally as corn bread. Corn is also an important source of the sweeteners, starches, oil and alcohol used in many foods, beverages and numerous other products. In the United States, corn is the largest crop in terms of planted acreage, total production and crop value. While corn is generally used as a high energy animal feed, it is also a very suitable raw

7
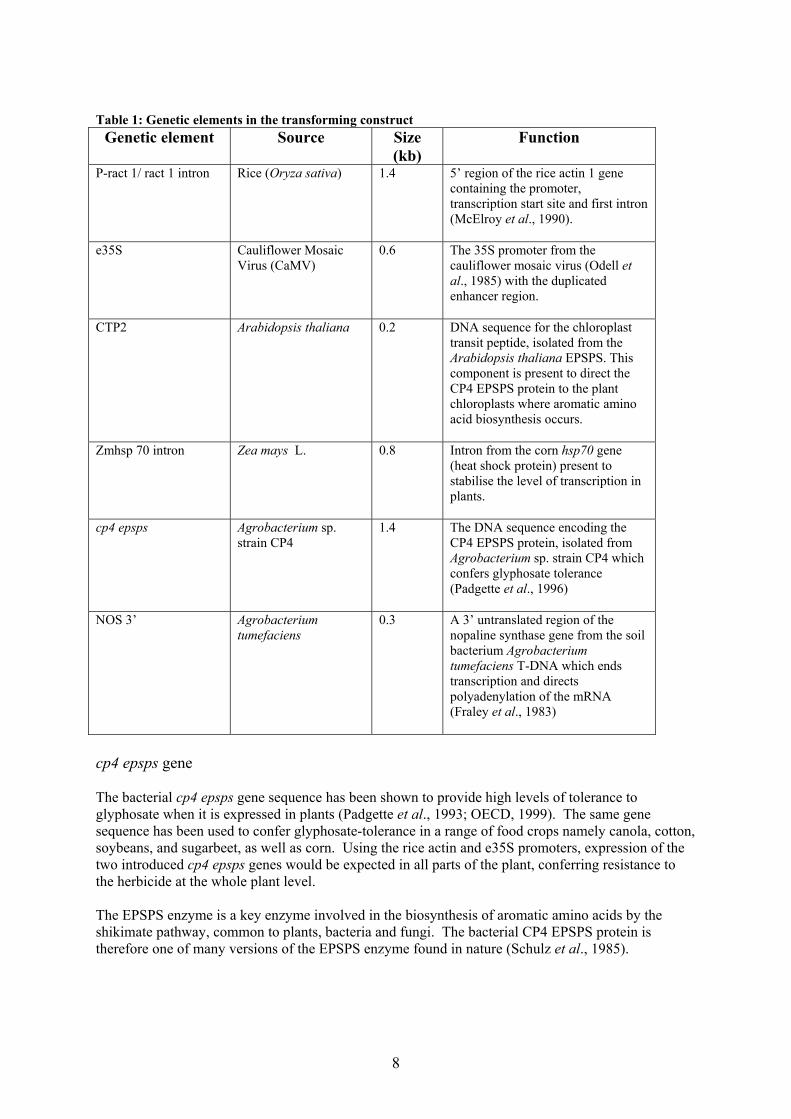
material for the manufacture of starch which is largely converted to a variety of products for human consumption, such as sweetener and fermentation products including high fructose corn syrup and ethanol. Corn oil is commercially processed from the germ and accounts for approximately nine percent of domestic vegetable oil production. Little whole kernel or processed corn is consumed by humans worldwide when compared to these corn-based food ingredients that are used in the manufacture of many foods including bakery and dairy goods, beverages, confections and meat products. DESCRIPTION OF THE GENETIC MODIFICATION Methods used in the genetic modification Corn line NK603 was generated by transformation of embryogenic corn (Zea mays) cells using a particle acceleration method. This method of transformation allowed for a specific segment of plasmid DNA, purified by gel electrophoresis and incorporating only the genes of interest together with essential controlling elements, to be transferred to the plant genome. Since the introduced DNA contained a gene coding for herbicide tolerance (in this case, the cp4 epsps gene), the plant cells were grown in the presence of glyphosate and only those cells that carry the DNA modification are able to grow successfully. The independent plant line, NK603, was subsequently developed from cultivation of the transformed corn cells. Function and regulation of novel genes A specific DNA segment of 6706 base pairs (bp) was purified from plasmid PV-ZMGT32 by agarose gel electrophoresis and subsequently used in the transformation of embryogenic corn cells. The purified fragment consisted of two adjacent gene expression cassettes, each comprising a single copy of the cp4 epsps gene fused to an optimised chloroplast transit peptide sequence and separate controlling DNA elements essential for expression in plant cells (see below and Table 1). The segment does not contain an antibiotic resistance selectable marker gene or bacterial origin of replication sequences. In the first (5’ end) expression cassette, the cp4 epsps gene is under the regulation of the rice actin promoter and rice actin intron. The second cassette, which is fused to the 3’ end of the first, consists of the cp4 epsps gene regulated by the enhanced cauliflower mosaic virus 35S promoter (e35S) and intron from the corn heat shock protein 70 (HSP70). Both expression cassettes incorporate the 3’untranslated region of the nopaline synthase gene (NOS 3’) for signal polyadenylation. Diagrammatically, the introduced DNA segment can be represented as follows:
5’ 6706 BP 3’
P-
ract 1
ract 1 intron
CP4 EPSPS
E35S
ZmHSP70 intron
CP4 EPSPS
CTP2 NOS 3’ CTP2 NOS 3’ Although plasmid PV-ZMGT32 contained other bacterial genes and controlling sequences for selection and replication in the laboratory, these sequences were not contained within the gel purified segment used in the transformation and therefore are not present in the plant.
8
Table 1: Genetic elements in the transforming construct
Genetic element Source Size (kb)
Function
P-ract 1/ ract 1 intron Rice (Oryza sativa) 1.4 5’ region of the rice actin 1 gene containing the promoter, transcription start site and first intron (McElroy et al., 1990).
e35S Cauliflower Mosaic Virus (CaMV)
0.6 The 35S promoter from the cauliflower mosaic virus (Odell et al., 1985) with the duplicated enhancer region.
CTP2 Arabidopsis thaliana 0.2 DNA sequence for the chloroplast transit peptide, isolated from the Arabidopsis thaliana EPSPS. This component is present to direct the CP4 EPSPS protein to the plant chloroplasts where aromatic amino acid biosynthesis occurs.
Zmhsp 70 intron Zea mays L. 0.8 Intron from the corn hsp70 gene (heat shock protein) present to stabilise the level of transcription in plants.
cp4 epsps Agrobacterium sp. strain CP4
1.4 The DNA sequence encoding the CP4 EPSPS protein, isolated from Agrobacterium sp. strain CP4 which confers glyphosate tolerance (Padgette et al., 1996)
NOS 3’ Agrobacterium tumefaciens
0.3 A 3’ untranslated region of the nopaline synthase gene from the soil bacterium Agrobacterium tumefaciens T-DNA which ends transcription and directs polyadenylation of the mRNA (Fraley et al., 1983)
cp4 epsps gene The bacterial cp4 epsps gene sequence has been shown to provide high levels of tolerance to glyphosate when it is expressed in plants (Padgette et al., 1993; OECD, 1999). The same gene sequence has been used to confer glyphosate-tolerance in a range of food crops namely canola, cotton, soybeans, and sugarbeet, as well as corn. Using the rice actin and e35S promoters, expression of the two introduced cp4 epsps genes would be expected in all parts of the plant, conferring resistance to the herbicide at the whole plant level. The EPSPS enzyme is a key enzyme involved in the biosynthesis of aromatic amino acids by the shikimate pathway, common to plants, bacteria and fungi. The bacterial CP4 EPSPS protein is therefore one of many versions of the EPSPS enzyme found in nature (Schulz et al., 1985).

9
However, the CP4 EPSPS protein has a high catalytic efficiency compared to most other EPSPS enzymes (Barry et al., 1992; Padgette et al. 1993 & 1996) and, in addition, is highly tolerant to glyphosate due to a lower binding affinity with that herbicide. The mechanism of action of glyphosate is to bind specifically to the EPSPS protein, blocking the enzyme activity, and thereby interfering with normal protein synthesis in plant cells, leading to plant death. Plants that express the CP4 EPSPS gene are tolerant to glyphosate due to the continued activity of the enzyme in the presence of the herbicide, allowing normal cellular functions to continue. The CP4 and native corn EPSPS enzymes are therefore functionally equivalent, except for their affinity for glyphosate. Chloroplast transit peptide In both plant gene expression cassettes, the CP4 EPSPS coding sequence is fused to a chloroplast transit peptide (CTP2) whose sequence is based on the CTP isolated from Arabidopsis thaliana EPSPS. The purpose of the CTP is to direct the new protein to the chloroplast, where the enzymes of the shikimate pathway operate in plant cells, and therefore where the endogenous corn EPSPS enzyme is naturally transported. Transit peptides are commonly occurring molecular mechanisms to facilitate intracellular transport of proteins between compartments within a cell. The CTP is typically cleaved from the mature protein on uptake into the chloroplast, and subsequently rapidly degraded. Characterisation of the genes in the plant Studies evaluated: Cavato, T.A., Deng, M.Y. and Lirette, R.P.. Confirmation of the Genomic DNA Sequences Flanking the 5’ and 3’ Ends of the Insert in Roundup Ready Corn Event NK603. Monsanto Laboratory Project 00-01-46-30, MSL 16857, completed October 2000. Deng, M.Y., Lirette, R.P., Cavato, T.A. and Sidhu, R.S.. Molecular characterisation of Roundup Ready (CP4 EPSPS) Corn Line NK603. Monsanto Laboratory Project 99-01-46-26, MSL 16214, completed October 1999. Kesterson, N.K., Reiser, S.E., Cavato, T.A. and Lirette, R.P.. PCR and DNA Sequence Analysis of the Insert in Roundup Ready Maize Event NK603. Monsanto Laboratory Project 01-01-46-25, MSL 17588, completed January 2002. Silanovich, A., Hileman, R.E., and Astwood, J.D.. Amended Report for Bioinformatic Evaluation of DNA Sequences Flanking the 3’ End of the NK603 Insertion Event: Assessment of Putative Polypeptides. Monsanto Laboratory Project 00-01-46-41, MSL 17005, completed October 2000 (Amendment 1 completed December 2000). Multiple molecular analyses were undertaken in order to characterise the inserted DNA in corn line NK603. Genomic plant DNA was analysed using the standard methodology of Southern blot analysis to determine the insert number and the copy number as well as to provide information about the integrity of the inserted regulatory sequences and to confirm the absence of any of the plasmid backbone sequences. Polymerase chain reaction (PCR) and nucleotide sequencing was used to verify the entire DNA segment, including the junction regions at the site of integration into the plant DNA. The test material used was leaf tissue taken from corn line NK603 grown under greenhouse conditions and treated with Roundup Ultra (64 ounces/acre) at the V2-V3 stage (2-3 leaf collars). Leaf tissue from the untransformed parental corn line LH82 x B73 grown under similar conditions was used as the control material. Data from the analyses support the conclusion that the genome of corn line NK603 contains a single DNA insertion, as determined by multiple Southern blots using different known molecular cleavage sites within the region of the introduced segment. The results from further analysis using different

10
DNA probes are consistent with and support the conclusion that one complete copy of the DNA segment used in the transformation is present. As well as the single complete copy of the DNA segment used in the transformation, the insert also includes a 217 bp fragment of the enhancer region of the rice actin promoter inversely linked at the 3’end of the introduced DNA. The evidence for this was provided by Southern blot analysis and confirmed by DNA sequence analysis of the regions at the ends of the inserted segment. Nucleotide sequence analysis of corn line NK603 The inserted DNA in corn line NK603 has been completely sequenced, providing unequivocal information about the nature and organization of the elements in this modification. These data reveal several molecular changes in the introduced segment in the plant compared with the segment used in the transformation. The results show that the additional 217 bp at the 3’ end includes polylinker sequence (50 bp) and the first 167 bp of the enhancer region of the rice actin promoter. Previously published studies on the rice actin promoter and intron indicate that sequences essential for promoter function are not present in these 167 nucleotides (McElroy et al. 1990). In particular, neither the TATA box nor transcription initiation site is present in the fragment, indicating that the sequence should not function as a promoter. The expected lack of promoter activity was also confirmed experimentally where transcripts initiated by this small fragment were not detected using the extremely sensitive technique of reverse transcriptase-PCR (RT-PCR). The sequencing data also revealed that the first (5’) cp4 epsps gene corresponds exactly to the original plasmid sequence, whereas the second (3’) cp4 epsps gene differs by two nucleotides. One of the nucleotide changes is silent, and does not alter the encoded amino acid. The second nucleotide change results in a single amino acid change to the CP4 EPSPS protein – a proline at position 214 of the protein instead of leucine. These changes were confirmed in DNA obtained from grain representing the F1 generation prior to the start of this study. The F1 generation is the progenitor for all NK603 generations, including the material used in safety assessment studies, as well as the material used to generate all commercial corn varieties developed from this line. The sequence variation therefore has been present since the earliest stages of development of this line. The nucleotide changes occurring in the coding region of the second cp4 epsps gene are minor and, of themselves, do not have implications for food safety. The variant protein, CP4 EPSPS L214P, that is produced as a result of the expression of this gene is discussed in more detail in later sections of this report. Analysis of the 5’ and 3’flanking sequences Further DNA sequence information was obtained with a focus specifically on the site of insertion and the flanking regions adjacent to the inserted DNA. These data supplement the data obtained by multiple Southern hybridisation experiments. Genomic DNA extracted from leaf tissue from corn line NK603 was sequenced to determine the nucleotide sequence at the 5’ and 3’ ends of the newly inserted segment. DNA extracted from the leaves of a non-transformed line B73 was used as a control in the PCR experiments. The sequence data reveal that a segment of chloroplast DNA (305bp) is immediately adjacent to the 3’ end of the introduced DNA segment. It is apparent that this additional DNA has co-integrated with the transformation cassette at the same time. Bioinformatic analysis identified the chloroplast sequence as corresponding to the coding sequence for the α-subunit of chloroplast DNA-directed RNA polymerase and ribosomal protein S11 in maize. This extraneous DNA was not present in the

11
gel-isolated segment used in the transformation process and therefore the origin of the chloroplast DNA was the transformed, embryonic maize cell itself. Given the method of transformation used to generate this line, some DNA rearrangements would reasonably be expected as these have been commonly observed in plant transformations and are often reported in the scientific literature. The rearrangements do not necessarily raise any public health or safety concerns provided that they are fully characterised using detailed molecular and bioinformatic tools. In this case, both the sequence analysis and information from previously published studies (McElroy et al., 1990) indicate that the additional sequence corresponding to a portion of the rice actin promoter at the 3’end of the introduced DNA is non-functional. There are several published examples where host genetic material has been observed to co-integrate with transgenes at the site of integration. It is most likely that this results from the normal DNA repair mechanisms naturally found in living cells and which are utilised deliberately and effectively in the plant transformation process. Moreover, movement of plastid DNA into the genome has been reported to occur naturally in many different plant species including tomato, spinach and rice (see for example Pichersky et al., 1991). The plastid DNA is merely present in the nucleus. Investigation of the chloroplast sequence and surrounding regions, do not reveal characteristics or properties that are likely to raise food safety concerns. Bioinformatic analysis of theoretical peptides encoded by the chloroplast sequence revealed no relevant sequence similarity to known toxins or allergens. The DNA corresponds to the native genetic material of the corn plant and, despite its non-native location in this plant line, has always been a natural part of this food. Stability of genetic changes The stability of the transferred genes was investigated to ascertain plant characteristics over multiple generations. Segregation data for nine generations were statistically analysed, based on the frequency of observed versus expected numbers of progeny with tolerance to glyphosate. The stability of the insert was demonstrated through six generations of crossing and three generations of self pollination. These data show that the herbicide tolerance trait in corn line NK603 is inherited according to predicted patterns, consistent with a single active site of insertion of the CP4 EPSPS into the genomic DNA, segregating according to Mendelian genetics. Southern blot analysis was also conducted to assess the genetic stability of the inserted DNA in this line including, as controls, non-transformed B73 corn DNA and the same B73 DNA spiked with the original plasmid DNA. Genomic DNA extracted from leaf tissues of the F1 generation (the progeny from a R0 back cross) and the fifth generation of back-crossing (BC5F1) of line NK603 and both control samples were appropriately cleaved, and probed with the full-length CTP2-CP4 EPSPS fragment. There were no detectable differences in the observed hybridisation pattern between the DNA extracted from the F1 generation and from the BC5F1 generation. These results demonstrate that the integrated segment in corn line NK603 is stable spanning at least five generations. Impact on human health from the potential transfer of novel genetic material to cells of the human digestive tract The human health considerations in relation to the potential for horizontal gene transfer depend on the nature of the novel genes and must be assessed on a case-by case basis.

12
In 1991, the World Health Organization (WHO) issued a report of a Joint FAO1/WHO Expert Consultation which looked at strategies for assessing the safety of foods produced by biotechnology (WHO 1991). It was concluded by that as DNA from all living organisms is structurally similar, the presence of transferred DNA in food products, in itself poses no health risk to consumers. The major concern in relation to the potential transfer of novel genetic material to cells in the human digestive tract is with antibiotic resistance genes. Antibiotic resistance genes can be present in some transgenic plants as a result of their use as marker genes in the laboratory or in the field. It is generally accepted that there are no safety concerns with regard to the presence in the food of antibiotic resistance gene DNA per se (WHO 1993). There have been concerns expressed, however, that there could be horizontal gene transfer of antibiotic resistance genes from ingested food to microorganisms present in the human digestive tract and that this could compromise the therapeutic use of antibiotics. However, in this case no antibiotic resistance gene is present in corn line NK603 so this is not an issue. In relation to the transfer of other novel genetic material from GM food to bacteria in the human digestive system, this is extremely unlikely to occur because of the number of complex and unlikely steps that would need to take place consecutively. It is equally unlikely that novel genetic material from genetically modified food would transfer to human cells via the digestive tract. In considering the potential impact on human health, it is important to note that humans have always consumed large amounts of DNA as a normal component of food and there is no evidence that this consumption has had any adverse effect on human health. Furthermore, current scientific knowledge has not revealed any DNA sequences from ingested foods that have been incorporated into human DNA. Novel DNA sequences in genetically modified foods comprise only a minute fraction of the total DNA in the food (generally less than 0.01%) and are therefore unlikely to pose any special additional risks compared with the large amount of DNA naturally present in all foods. Conclusion At the molecular level, the analyses indicate that the transformation process has resulted in a single insertion event, comprising one complete copy of the transformation cassette together with an additional small portion of the enhancer region of the rice actin promoter, linked at the 3’ end of the inserted DNA in an inverse orientation. The additional nucleotides are completely identified and are not expected to be functional on the basis of previously published studies delineating the minimum sequence requirements for functionality. Two nucleotide changes are present in the second copy of the cp4 epsps gene, resulting in the expression of a protein, CP4 EPSPS L214P, which varies by one amino acid from the expected CP4 EPSPS sequence. This change was detected in the F1 generation and has therefore been present from the earliest stages of development of corn line NK603, including throughout all of the downstream scientific analyses of this line. Approximately 300 bp of chloroplast DNA has co-integrated during transformation and is adjacent to the insert. The sequence has been identified as corn chloroplast coding sequence, however on the basis of detailed investigation, there is no evidence that its presence has any impact on corn line NK603. The inserted DNA is physically stable and is inherited in a predictable manner over multiple generations.
1 Food and Agriculture Organization.

13
CHARACTERISATION OF NOVEL PROTEIN Biochemical function and phenotypic effects Studies evaluated: Padgette, S.R., Barry, G.F., Re, D.B., Weldon, M., Eichholtz, D.A., Kolacz, K.H. and Kishore, G.M., 1993. Purification, Cloning and Characterisation of a Highly Glyphosate Tolerant EPSP Synthase from Agrobacterium sp. strain CP4. Monsanto Technical Report MSL-12738, St. Louis, Missouri. As part of the safety assessment of glyphosate-tolerant corn line NK603, the assessment examines the expressed products of the introduced genes and considers the levels of new protein in the grain. In this line, the only expressed protein product from the inserted gene cassettes is the CTP2-CP4 EPSPS protein, and the sequence variant CTP2-CP4 EPSPS L214P, which differs from CP4 EPSPS by one amino acid. The EPSPS enzyme catalyses a non-rate limiting step in the shikimate pathway involved in aromatic amino acid biosynthesis in plants and microorganisms. Since EPSPS is naturally present in plants, bacteria and fungi as part of the basic biochemical makeup of the organism, several scientific studies have compared the amino acid sequences and catalytic properties of the enzyme from a wide variety of different sources (see Schultz et al., 1985 and Barry et al., 1992). Data from these studies show that differences in amino acid sequence of the enzyme from different species, including bacteria and fungi, result in varying degrees of sensitivity to glyphosate. The bacterial CP4 version of the EPSPS enzyme introduced into corn line NK603 exhibits a lower binding affinity for glyphosate and thus exhibits high catalytic efficiency in the presence of glyphosate when compared to the native corn EPSPS. The catalytic function of the introduced CP4 EPSPS enzyme is well characterised in plants. It has been established that CP4 EPSPS is highly specific for its natural substrates, shikimate-3-phosphate and phosphoenolpyruvate, similar to the corn enzyme (Padgette et al., 1993). The characterisation included an examination of three dimensional folding patterns of the protein and sequence homology at the active site enabling comparison with the structure and function of the native corn EPSPS. The shikimate pathway does not occur in mammals, where aromatic amino acids are provided from other sources, a fact contributing to the selective toxicity of glyphosate to plants. The CP4 EPSPS has been completely sequenced and encodes a 47.6 kDa protein consisting of a single polypeptide of 455 amino acids (Padgette et al., 1996). The bacterial enzyme exhibits approximately 50% amino acid sequence similarity with plant EPSPS enzymes (eg. soybean, corn and petunia). The degree of similarity of the CP4 EPSPS protein to other EPSPS enzymes naturally present in all food crops (eg. soybean and corn) and in fungal and microbial food sources such as Baker’s yeast (Saccharomyces cerevisiae) and Bacillus subtilis which have been safely consumed by humans for centuries, is evidence that this family of proteins has been an integral part of the food supply throughout history. Variant protein CP4 EPSPS L214P One of two nucleotide changes in the coding region of the second cp4 epsps gene has resulted in a single amino acid substitution in the gene product. The variant protein contains a proline instead of a leucine at position 214 and is therefore referred to as CP4 EPSPS L214P. Bioinformatic and biochemical analyses of the variant protein were assessed in order to compare its properties with CP4 EPSPS protein which is also expressed in corn line NK603. When compared with other EPSPS enzymes, structural modelling diagrams show that the overall predicted secondary and tertiary structure of the variant L214P protein is maintained. Moreover, the

14
amino acid substitution has occurred away from the active site and has not altered the functional activity of the enzyme, which is still equivalent to that of the non-variant CP4 EPSPS. This observation is consistent with previously published reports that compare specific activities and protein sequences of this enzyme from a range of species and conclude that the three-dimensional structure is the key element to enzymatic function for the EPSPS family of proteins, not the precise amino acid sequence (Padgette et al., 1996). Protein expression analyses Under the regulation of the rice actin promoter (expression cassette 1) and the 35S promoter (expression cassette 2), the new protein is expected to occur throughout the whole plant, including the grain, since these promoters have been shown to drive constitutive gene expression in genetically modified corn. Characterisation of the expressed protein and determination of the level of novel gene expression in corn line NK603 was performed by various methods including Western blot analysis (immunoblotting) and an enzyme-linked immunosorbent assay (ELISA). Studies evaluated: Bisop, B.F., 1993. Production of CP4 EPSP synthase in a 100 litre recombinant Escherichia coli fermentation. Monsanto Technical Report MSL-12389, St Louis, Missouri. Harrison, L.A., Leimgruber, M.R., Smith, C.E., Nida, D.L., Taylor, M.L., Gustafson, M., Heeren, B. and Padgette, S.R., 1993. Characterisation of Microbially-Expressed Protein: CP4 EPSPS. Monsanto Technical Report MSL-12901, St Louis, Missouri. Ledesma, B.E. and Sidhu, R.S., 1999. Development and validation of a direct ELISA for quantitation of CP4 5-enolpyruvylshikimate-3-phosphate synthase (CP4 EPSPS) protein in corn tissues from Roundup Ready® plants. Monsanto Technical Report, MSL-16259, St. Louis, Missouri. Lee, T.C. and Astwood, J.D., 1999. Assessment of the Equivalence of CP4 EPSPS Protein Expressed in Escherichia coli and in Roundup Ready® corn lines NK600 and NK603. Monsanto Technical Report MSL-16392, St Louis, Missouri. Western blot analysis The expression of the full-length CP4 EPSPS protein in the grain from corn line NK603 was confirmed by Western blot analysis. Two control materials were used for this study. The first control material was obtained from a non-transformed parental corn line (LH82xB73) that does not contain the genetic material to encode CP4 EPSPS. Grain for the transformed and non-transformed corn lines was collected from field grown plants. The second control material was a non-transformed soybean line, A5403, which likewise does not express CP4 EPSPS. The presence or absence of the CP4 EPSPS gene in the tested lines was established by a PCR detection method. Two reference materials were also used for this study. The primary reference material was in vitro produced CP4 EPSPS derived from recombinant E. coli, transformed with a plasmid encoding the enzyme. A second reference material was used, which was CP4 EPSPS endogenously expressed by a similarly transformed soybean line, AG3701, obtained from Asgrow (Stonington, Illinois). This soybean line is glyphosate-tolerant due to the presence and expression of the bacterial CP4 EPSPS gene, also present in corn line NK603. Immunoblotting involved the use of polyclonal antisera raised in goats against the E. coli produced CP4 EPSPS protein. The results of the Western blot analysis demonstrated that the E. coli produced CP4 EPSPS protein used for the safety studies, the CP4 EPSPS expressed in the glyphosate-tolerant soybeans and the CP4 EPSPS expressed by glyphosate-tolerant corn line NK603 were identical, based on electrophoretic
15
mobility and detection using specific antibodies. Immunoreactive bands at the expected apparent molecular weight (approx. 47 kDa) were observed for all CP4 EPSPS-containing samples, whether E. coli produced protein or extracted from the transformed corn or soybean plants. No immunoreactive bands were detected in the control (untransformed) corn or soybean extracts, confirming the specificity of the antibodies in detecting the expressed protein. ELISA detection Levels of the CP4 EPSPS protein were estimated in both forage and grain samples collected from six non-replicated and two replicated field sites, representative of the major U.S. corn production region during the 1998 growing season. Samples collected from line NK603 and the non-transformed parental control line (LH82xB73) were analysed using ELISA. The CP4 EPSPS protein levels in forage and grain extracts were estimated using a double antibody sandwich ELISA consisting of a monoclonal anti-CP4 EPSPS antibody as the capture antibody and a polyclonal anti-CP4 EPSPS conjugated to horseradish peroxidase (HRP) as the detection antibody. The CP4 EPSPS protein levels in plant tissue extracts were quantified by comparison of the sample absorbance (OD) to the absorbance produced by a range of concentrations of purified CP4 EPSPS reference standard. This protein standard was purified in the laboratory from an E. coli strain expressing the Agrobacterium sp. strain CP4 EPSPS, and fully characterised. The CP4 EPSPS protein levels estimated in corn forage and grain samples are summarised in Table 2. The levels of CP4 EPSPS protein in all non-transformed control samples were below the limit of quantitation (LOQ) of the assay (data not presented). Table 2. Summary of CP4 EPSPS protein levels measured by ELISA in tissues of NK603 corn plants (µg/g fresh weight). The LOQ for forage equals 0.05 µg/g fw, and for grain equals 0.09 µg/g fw. Sites Parameter Forage
µg/g fw Grain
µg/g fw Non-replicated Mean
Range SD
25.5 18.0-31.2
4.5
11.0 6.9-15.6
3.2 Replicated Mean
Range SD
25.9 25.7-26.1
0.3
10.6 9.8-11.3
1.0 All sites Mean
Range SD
25.6 18.0-31.2
3.8
10.9 6.9-15.6
2.6 SD = Standard Deviation As expected, the results of the ELISA show that mature CP4 EPSPS is present in low concentrations in the grain and at higher concentrations in the forage of corn line NK603. A higher level of novel protein expression in the green tissues of the plant (corresponding to the forage) is consistent with the functional rice actin and viral promoters used in the gene constructs. Although the level of expression is low, it is sufficient to confer tolerance to glyphosate at the level of the whole plant. The mean CP4 EPSPS protein levels in NK603 grain were comparable at the non-replicated sites (11.0 µg/g fw) and the replicated sites (10.6 µg/g fw) indicating that the novel protein is expressed at approximately the same levels either within a site or across geographically dispersed sites. The novel protein expressed in corn line NK603 is CP4 EPSPS and the close sequence variant CP4 EPSPS L214P. The CP4 EPSPS is a bacterial form of an enzyme already naturally occurring in all plants including corn. Analysis of the expression of novel protein in the modified line indicates that the new protein is present at low levels in the grain, the part of the plant used as food, and at higher levels in the forage, which is not used for human consumption. No laboratory marker genes, in particular antibiotic resistance genes, were transferred during the plant transformation process.

16
Potential toxicity of novel protein The safety of other foods derived from genetically modified crops containing the CP4 EPSPS protein used in corn line NK603 has been previously addressed in assessments of glyphosate-tolerant soybeans, insect-protected corn, glyphosate-tolerant cotton and glyphosate-tolerant canola. Studies that are of some relevance to an assessment of the potential toxicity and allergenicity of this protein in the context of other GM foods have been published in the scientific literature (for example, Harrison et al., 1996; Hammond et al., 1996). Detailed protein expression analyses have demonstrated that the only new proteins arising from the modification in corn line NK603 are the bacterial CP4 EPSPS enzyme and the close sequence variant CP4 EPSPS L214P, which differs from CP4 EPSPS by one amino acid. The CP4 EPSPS gene has been completely sequenced and encodes a 47.6 kDa protein consisting of a single polypeptide of 455 amino acids. At the amino acid level, this enzyme is similar to other EPSPS enzymes in this family of proteins with a function common to plants and microorganisms. The family of EPSPS proteins are naturally present in foods derived from plants (for example, soybean and tomato) and microbes (for example, Baker’s yeast and Bacillus subtilis) and have no known history of toxicity or allergenicity. The similarity of the CP4 EPSPS protein to these other EPSPS proteins provides supporting evidence for the safety of this protein. Studies evaluated: Harrison, L.A., Bailey, M.R., Leimgruber, R.M., Smith, C.E., Nida, D.L., Taylor, M.L., Gustafson, M.E., Heeren, B. and Padgette, S.R. (1993) Characterisation of Microbially-Expressed Protein: CP4 EPSPS. Monsanto Technical Report MSL-12901, St Louis, Missouri. Hileman, R.E. and Astwood, J.D. (1999b). Bioinformatics Analysis of CP4 EPSPS Protein Sequence Utilising Toxin and Public Domain Genetic Databases. Monsanto Technical Report MSL-16268, St. Louis, Missouri. Naylor, M.W. (1993). Acute Oral Toxicity Study of CP4 EPSPS Protein in Albino Mice. Monsanto Technical Report MSL-13077, St Louis, Missouri. Sequence comparison to known toxins EPSPS proteins from plants and other biological sources have a long history of consumption by humans and have not been associated with toxicity in relation to human health. The bacterial CP4 EPSPS has been specifically tested for potential toxicity using a range of scientific approaches. A database of 4,677 protein sequences (not all unique) associated with toxicity was assembled from publicly available genetic databases such as PIR, SwissProt, EMBL and GenBank. The amino acid sequence of the CP4 EPSPS protein was compared to protein sequences in the toxin database using the FASTA sequence alignment tool. In addition, the amino acid sequence of the CP4 EPSPS protein was compared to all protein sequences in the publicly available sequence databases to screen for structural similarity to other known proteins, including pharmacologically active proteins. As expected from prior examinations and comparisons, the CP4 EPSPS protein shares sequence similarity only with other EPSPS proteins from different biological sources. These computer searches did not reveal other significant structural homology, confirming the lack of similarity of the CP4 EPSPS protein to known protein toxins. Bioinformatic analyses were performed for the variant protein CP4 EPSPS L214P using four types of sequence searches: ALLERGENSEARCH analysis using the variant sequence as a query to search the ALLERGEN3 database and FASTA analyses using the variant sequence to search the ALLERGEN3, TOXIN5 and ALLPEPTIDES databases. As for the non-variant protein, the results of these searches do not indicate any significant structural similarity of the variant protein to known protein toxins or

17
allergens in these databases. As would reasonably be expected for two proteins that differ in only one amino acid in 445, the results obtained for searches of the ALLPEPTIDES and ALLERGEN3 databases using CP4 EPSPS L214P parallel the bioinformatic assessment that was obtained for CP4 EPSPS. Acute oral toxicity study in mice As a further test for potential toxicity, an acute oral toxicity study of CP4 EPSPS in young laboratory mice using purified (>90%) protein produced in E. coli in the laboratory was performed. A separate protein characterisation study was completed in order to confirm the equivalence of the bacterially produced enzyme used in the toxicity study to the protein expressed in the modified plants. The results of the study showed that the purified CP4 EPSPS exhibits the appropriate chemical identity and integrity as determined by gel electrophoresis, Western blot (immunoblotting), N-terminal amino acid sequencing and ELISA. The purified protein also demonstrated functional identity as determined by enzymatic activity. The study was conducted in general compliance with the EPA FIFRA (40 CFR Part 160). A total of 100 animals (50 males and 50 females) were used in this study, ranging from 5.5 weeks to 7 weeks of age. Test groups were randomised for weight and comprised 10 CD-1 mice of each sex per group. The protein preparation containing the CP4 EPSPS was administered as a single dose by gavage to three groups of the mice at dosages of 49, 154 and 572 mg/kg body weight respectively. These doses correspond to 40, 100 and 400 mg/kg of CP4 EPSPS protein based on the level of purity of the protein and ELISA analyses of the dosing solutions. A control group received bovine serum albumin (BSA) at a dosage of 363 mg/kg in the same solution and delivery volume as the test substance. The second control group was administered the carrier solution only, 50 mM sodium bicarbonate. At defined stages throughout the duration of the study, clinical observations were performed for mortality and signs of toxicity, and body weights and food consumption measured. Signs of toxicity include such occurrences as changes in the skin and fur, eyes and mucous membranes, respiratory, autonomic and central nervous systems as well as behavioural changes. At the termination of the study (day 8-9), animals were sacrificed, examined for gross pathology and numerous tissues were collected. Tissues retained from the animals included aorta, adrenals, brain, colon, oesophagus, eyes, gall bladder, heart, kidneys, lung, liver, lymph nodes, muscle, ovaries, pancreas, pituitary, prostate, rectum, salivary gland, seminal vesicles, skin, spinal cord, spleen, stomach, testes, thymus, uterus and bladder. Hollow organs were opened and examined. The results of the study showed no statistically significant differences in group mean body weights, cumulative weight gains or food consumption in any of the groups treated with either BSA or the CP4 protein, when compared with the carrier control group. The data were evaluated according to a decision-tree analysis procedure which, depending on the results of early statistical tests, determined further statistical analysis applied to detect group differences and analyse for trends. All animals survived to the scheduled termination of the study, and there were no clinical signs observed that could be related to the test material. As recorded in Table 3, a small number of general pathological observations were detected in the female mice but these occurred throughout all groups in the study, including both control groups that did not receive the test material, and therefore these findings cannot be related to the treatment. There were no such findings in the male animals in any of the test or control groups in the study.
18
Table 3. Pathology - Incidence of individual gross necropsy alterations, females.
Carrier Control
N=10
BSA Control
N=10
CP4 EPSPS 40 mg/kg
N=10
CP4 EPSPS 100 mg/kg
N=10
CP4 EPSPS 400 mg/kg
N=10 Eye (corneal opacity)
0 0 0 1 0
Kidney (cyst)
0 0 1 0 0
Pituitary (focus)
1 0 0 0 0
Uterus (hydrometra)
2 1 1 1 2
In conclusion, there was no evidence of acute toxicity in mice following a single oral dose of up to 400 mg/kg of CP4 EPSPS protein. This dose level is far in excess of the level of exposure expected from the consumption of modified corn. Potential allergenicity of novel protein Although many foods have been reported to cause allergies in some people, the prevalence of food allergy using prospective, population-based studies has been shown to be less than 2% of adults and 2-7% of infants and children, excluding cases of food intolerances such as enzyme deficiencies. Food allergies are primarily due to an immune reaction to a particular protein or glycoprotein component of the food. Studies evaluated: Astwood, J.D., George, C., Alibhai, M., McCoy, R., Lahman, L., Hammond, B.G., Leach, J.N. and Silvanovich, A. (2001). Safety Assessment of Roundup Ready Corn Event NK603 Containing Genes Encoding CP4 EPSPS and CP4 EPSPS L214P. MSL Number 17600. Hileman, R.E. and Astwood, J.D., 1999a. Bioinformatics Analysis of CP4 EPSPS Protein Sequence Utilising an Allergen Database, Monsanto Technical Report MSL No. 16267, St. Louis, MO. Ream, J.E., Bailey, M.R., Leach, J.N. and Padgette, S.R., 1993. Assessment of the in vitro digestive fate of CP4 EPSP synthase. Monsanto Technical Report MSL-12949. The potential allergenicity of the new protein introduced into corn line NK603 has been assessed by comparing certain molecular and biochemical properties of the new protein to those of known allergens. These include amino acid sequence similarity with known protein allergens, poor digestibility and resistance to processing. Other factors that are taken into account and that may increase the likelihood of allergic oral sensitisation to proteins include the level of food consumption, and the relative quantity of the protein in the food. Digestibility of CP4 EPSPS Typically, most food allergens tend to be stable to the peptic and acidic conditions of the digestive system if they are to reach and pass through the intestinal mucosa to elicit an allergic response (Kimber et al., 1999; Metcalfe et al., 1996). To address the question of potential allergenicity, the physicochemical properties of the CP4 EPSPS protein, which is expressed in the corn grain at low levels were investigated, and its susceptibility to proteolytic degradation was tested. Simulated mammalian gastric and intestinal digestive mixtures (described in the United States

19
Pharmacopoeia, 19902) were established to assess the susceptibility of the CP4 EPSPS protein to in vitro proteolytic digestion. The protein was incubated at approximately 37°C in simulated mammalian gastric fluid (SGF) and simulated intestinal fluid (SIF). At defined periods, the digestions were terminated and the levels of remaining CP4 EPSPS protein were determined by Western blot analysis and enzymatic activity assays. The results show that CP4 EPSPS protein degraded readily in both simulated gastric and intestinal fluids, indicating that it would similarly break down during the processes involved in human digestion. Western blot analyses demonstrated that the half–life of the protein was less than 15 seconds in the gastric system. The results of the activity assay confirmed that the activity of the enzyme had decreased by greater than 84% at the first timepoint (that is, after 2 minutes incubation). There was a strong correlation between the results of the Western blot analysis and the enzymatic activity assay in the SGF experiments, providing evidence that the protein degrades rapidly in the stomach when ingested by mammals as a component of food. In simulated intestinal fluid, the half-life of the CP4 EPSPS protein was less than 10 minutes as determined by Western blot analysis. In addition, the enzyme activity had decreased by greater than 94% after approximately 4.5 hours incubation. Overall, these digestibility results show that the introduced protein in corn line NK603 is readily degraded in a simulated digestive system and similarly readily degraded in the conditions of the mammalian digestive tract. Biochemical studies on CP4 EPSPS L214P Variant CP4 EPSPS L214P was produced in the laboratory by microbial fermentation and purified to 98% homogeneity by standard EPSPS purification techniques. The specific enzymatic activity of the CP4 EPSPS L214P was determined to be equivalent to similarly purified CP4 EPSPS. In the transformed corn, the variant protein confers the same enzymatic function as the non-variant CP4 EPSPS. Based on the structure/function data and the high degree of similarity of the two proteins, the in vitro digestibility of CP4 EPSPS L214P in simulated gastric fluid (SGF) was predicted to parallel that of CP4 EPSPS. This was confirmed by digestibility experiments performed over a range of time points between 15 seconds and 60 minutes. As with previous digestibility experiments, the degree of protein digestion was assessed using SDS-PAGE and gel staining techniques. The results showed that greater than 98% of the CP4 EPSPS L214P protein was rapidly digested within 15 seconds in SGF. Sequence comparison to known allergens A comparison of the amino acid sequence of an introduced protein with the amino acid sequence of known allergens is a further useful indicator of the potential for allergenicity, based on the identification of contiguous identical sequence matches which may be immunologically significant. A database of 567 protein sequences associated with allergy and coeliac disease was assembled from publicly available genetic databases (GenBank, EMBL, PIR and SwissProt) and from current literature. The amino acid sequence of the introduced CP4 EPSPS protein was compared to these assembled sequences using the sequence alignment tool FASTA (see earlier discussion on potential toxicity). The results of the alignment showed that CP4 EPSPS shared no structurally significant sequence similarity to sequences within the assembled allergen database. In addition, the amino acid sequence of the CP4 EPSPS protein was compared to the allergen database using an algorithm that scans for a window of eight identical linearly contiguous amino acids. This
2 The United States Pharmacopaeia, 1990, Volume XXII, NF XVII. United States Pharmacopeial Convention, Inc., Rockville, MD, page 1788.

20
comparison did not find any sequence identities between the introduced protein and the database sequences. Similarly, as was predicted from the structure/function data and the results of the toxin database searches, the variant CP4 EPSPS L214P protein showed no sequence identity to known allergens in the databases using the standardised parameters. Abundance of CP4 EPSPS Most allergens are present as major protein components in a specific food, typically ranging between 1% and 80% of total protein (Astwood and Fuchs, 1996). In contrast, CP4 EPSPS protein is present at approximately 0.01% of the total protein found in the grain of corn line NK603, noting that the grain is composed predominantly of carbohydrate and that protein normally comprises approximately 20%-25% of the grain. Corn flour is therefore the major food product likely to include corn proteins while corn oil and corn syrup are not expected to contain plant proteins including the introduced CP4 EPSPS protein. Conclusion The CP4 EPSPS protein is structurally and biochemically similar to other EPSPS enzymes from various plant and microbial food sources that are currently part of the human diet and have been consumed over a long period without any health concerns. The protein does not exhibit sequence similarity with known toxins and allergens, and does not exhibit the biochemical characteristics of known protein allergens. When fed as a single dose to laboratory mice at levels greatly exceeding the likely human level of exposure through consumption of whole corn grain or flour, there was no evidence of acute toxicity. Furthermore, the novel protein is present in relatively low abundance in the grain and demonstrates digestive lability in conditions that mimic human digestion. The additional biochemical and bioinformatic analyses applied to the variant protein CP4 EPSPS L214P expressed in NK603 corn confirm that it is indistinguishable from the non-variant CP4 EPSPS protein in terms of structural characteristics, enzymatic function, biochemistry and the potential for allergenicity or toxicity. The combined analyses investigating the potential toxicity and allergenicity properties of the novel proteins therefore strongly support the conclusions that corn line NK603, expressing both versions of CP4 EPSPS, does not pose a food safety risk. COMPARATIVE ANALYSES The safety assessment of foods produced using gene technology entails, in this case, evaluating compositional data from the transgenic corn plant in comparison with equivalent data from the parental (or non-transformed) plant line or literature values for the particular crop species. This process involves identifying the key components, including nutrients and any toxicants, characteristic of corn grain and also takes into account the variation in composition due to genetic variability, environmental factors, and post-harvest handling and processing. Studies evaluated: Ridley, W.P., George, C., Nemeth, M.A., Astwood, J.D., Breeze, M.L.* and Sorbet R.**, 2000. Compositional Analyses of Forage and Grain Collected From Roundup Ready Maize Event NK603 Grown in 1999 E.U. Field Trials. Monsanto Technical Report MSL-16897, St Louis, MO. Sidhu, R.S. and Ledesma, B.E., 1999. Introduced Protein Levels and Compositional Analyses of Roundup Ready Corn Line NK603 Tissues Produced in 1998 U.S. Field Trials. Monsanto Technical Report MSL-16278, St. Louis, MO. * Covance Laboratories, Madison, Wisconsin.

21
** Statistical Analysis Facility, Certus International Inc., Chesterfield, MO. Nutrient analyis The key nutrients in corn have been evaluated in order to compare equivalent data from the transformed line NK603, the non-transformed counterpart and published literature ranges obtained for conventional varieties of corn. This evaluation includes a study of the major constituents that are characteristic of whole corn grain, taking account of the natural variation in composition that is known to occur due to genetic variability and multiple environmental factors. Two major studies were conducted to determine the compositional profile of key corn tissues collected from corn line NK603, the non-transformed parental control line and a series of commercial corn hybrids grown under field conditions. Trial sites were selected across the United States corn-growing belt and in multiple sites across Europe. The U.S. sites included two replicated sites in Illinois and Ohio and six non-replicated sites in Iowa, Illinois, Indiana and Kansas. The European sites included four replicated sites located in Germignonville, Janville and L’isle Jourdain in France and Bagnarola, Italy. These sites provided a breadth of environmental conditions representative of regions where corn varieties are grown as commercial products. Sample preparation and collection Grain and forage samples of line NK603, treated with glyphosate herbicide (application rates supplied), and the non-genetically modified parental control line together with other commercial hybrids were collected from the range of sites. In the U.S. trials, several glyphosate-tolerant corn lines, including NK603, as well as the control line were planted at each site. Five different non-transformed commercial reference hybrids were planted at each of the European sites. The test and control substances were characterised at the molecular level by extracting DNA from grain tissue and analysing the DNA by event specific PCR techniques. In general, forage was collected at the late dough/early dent stage by dividing approximately 12 randomly selected plants into three roughly equal segments and placing them on dry ice within 10 mins of collection. Ears were harvested from approximately 12 self-pollinated plants at normal kernel maturity (<32% moisture), dried to a moisture level between 10-20%, shelled and the kernels pooled to provide the grain sample. Forage (stored on dry ice) and grain (stored at ambient temperature) samples were then transferred to the laboratory for compositional analyses. To conduct the analyses, forage and grain samples were collected from glyphosate-tolerant corn line NK603, the non-transformed parental control line (LH82 x B73) and reference hybrids. The control line has the same genetic background as that of the test line but lacks the gene encoding CP4 EPSPS protein. Compositional analyses included measurement of proximates (protein, fat, ash, moisture), acid detergent fibre (ADF), neutral detergent fibre (NDF), amino acids, fatty acids, vitamin E, minerals (calcium, copper, iron, magnesium, manganese, phosphorus, potassium, sodium and zinc), phytic acid and trypsin inhibitor content of the grain. Forage samples were analysed for proximate, ADF and NDF content. Carbohydrate values in both forage and grain were estimated by calculation. Detailed results from the compositional analyses of both the grain and forage on a site-by-site basis were evaluated in the safety assessment, however, not all of the results are presented here. Moreover, as the forage is not consumed by humans, the focus of this assessment was primarily on the results obtained from analyses of the grain samples. The forage data provide supporting evidence of the lack of significant differences in composition throughout the transformed and non-transformed plants, including tissues of the plants that are not part of the human diet.

22
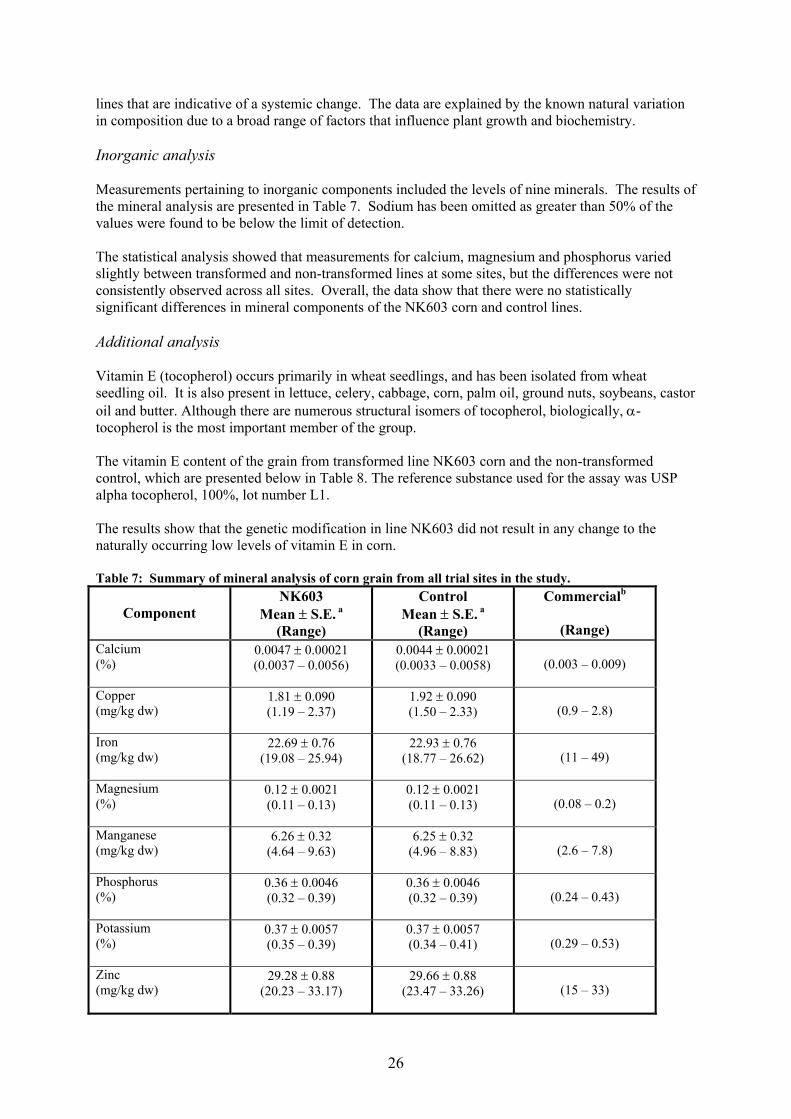
Statistical analysis The compositional data from the U.S. field trials were developed and statistically analysed as three sets of comparisons: analyses for each of the two replicated trials and for a combination of trials at different field sites. Similar multiple statistical analyses were applied to the European trial data. The test event, NK603, was compared to the non-transformed control line to determine statistically significant differences at p<0.05. In addition, the comparison of NK603 to the 95% tolerance interval for the commercial reference varieties was conducted to determine if the range of values for NK603 fell within the population of commercial corn. The data presented here are summary data pertaining to all trials. Analytical methods Detailed information relating to the methods used in the generation of the compositional data was evaluated. For each parameter tested, as well as appropriate technical references, the limit of detection or quantitation was also stated. Methodology was predominantly derived from established references such as American Association of Cereal Chemists (AACC), Association of Official Analytical Chemists (AOAC), American Oil Chemists Society (AOCS), the United States Department of Agriculture (USDA) Agricultural Handbook, or from a range of published literature methods. Proximate analysis – U.S. study The results obtained for the proximate analysis (including fibre) of the grain taken from all trial sites are presented in Table 4. The difference in the mean value for the moisture content between line NK603 and the control line was statistically significant (p-value 0.037), although values were within the commercial range for corn. This difference is not considered to be of biological significance and does not adversely affect the overall nutritional qualities or safety of food derived from the transformed corn line. There was no statistically significant difference between the transformed corn line and the non-transformed control for the remaining proximate analyses. Furthermore, all values were within the measured range for conventionally produced commercial lines of corn and, where comparable data were available, were also within the range reported in the general literature (Watson, 1987). Proximate analysis – European study The statistical evaluation of the combined data from all sites showed that there were no statistically significant differences between line NK603 and the control line for the content of moisture, fat, protein, ash, carbohydrate, ADF and NDF in forage, and for the content of ash, moisture, ADF, NDF, carbohydrate and protein in grain. There was a statistically significant (p<0.001) difference between the two lines in the percentage of total fat in the grain, as shown in Table 5. As this difference was not observed in the forage from the same sample set, nor was it a consistent observation on a site-by-site basis, the result is considered to reflect random fluctuations generally observed when multiple compositional studies are undertaken at a range of different agricultural sites. Moreover, the range of values obtained for both the NK603 line and the control line were within the published literature range (Watson, 1982) for this component.
23
Table 4: Summary of Proximate and Fibre Analysis from all U.S. trial sites.
Component NK 603 corn Mean ± S.E.a
(Range)
Control Mean ± S.E. a
(Range)
Commercialb
(Range) Ash (% dw)
1.44 ± 0.032 (1.28 – 1.75)
1.49 ± 0.032 (1.32 – 1.75)
(0.8 – 1.8)
Carbohydrates (% dw)
82.59 ± 0.39 (80.71 – 84.33)
82.26 ± 0.39 (80.23 – 83.70)
(83.1 – 89.6)
ADF (% dw)
3.79 ± 0.16 (3.14 – 5.17)
3.70 ± 0.16 (2.79 – 4.28)
(2.3 – 5.7)
NDF (% dw)
10.38 ± 0.67 (7.89 – 12.53)
10.32 ± 0.67 (8.25 – 15.42)
(8.2 – 16.1)
Moisture (% fw)
11.08 ± 0.45 (9.01 – 13.30)
11.76 ± 0.45 (8.56 – 14.8)
(6.1 – 15.6)
Total fat (%)
3.54 ± 0.09 (2.92 – 3.94)
3.59 ± 0.09 (2.88 – 4.13)
(1.7 – 4.3)
Protein (% dw)
12.43 ± 0.44 (10.30 – 14.77)
12.66 ± 0.44 (11.02 – 14.84)
(6.7 – 13.4)
dw=dry weight; fw=fresh weight. a The mean of all values ± standard error of the mean. b The range of sample values for commercial lines grown in 1998. (Sidhu 1999) Amino acid analysis Analysis of corn grain and forage included measurement of 18 essential amino acids, excluding glutamine and asparagine. In the U.S. study, with the exception of phenylalanine, none of the amino acid measurements for the grain showed a statistically significant difference between the transformed line NK603 and the non-transformed control. For the amino acid phenylalanine, in grain from NK603 assayed from all sites, the mean value was 5.34 ± 0.032 (% of total), whereas the mean value for phenylalanine in the non-transformed control over all sites was 5.26 ± 0.032 (% of total). These values compare favourably to the commercial range for phenylalanine of 4.7 to 5.5 as represented by lines grown commercially in 1998, and the literature range of 2.9 to 5.7 (Watson, 1982). The magnitude of the difference in the values for phenylalanine between the transformed line and its comparator is small, and is not of concern with respect to food safety. Moreover, both the lines are within the previously reported numerical ranges for this amino acid in commercialised corn grain. Amino acids – European trials Of the eighteen amino acids measured, small statistically significant differences were observed in six, including alanine, arginine, glutamic acid, histidine, lysine and methionine. The mean levels of some amino acids showed small increases (for example, alanine 1%; glutamic acid 2%) while for others the levels showed small decreases (for example, histidine 4%; arginine 6%). The results of the analysis of these combined data are presented in Table 5. As previously described for the proximate analysis results, the differences were not observed consistently across all sites in the study and therefore are not indicative of a general trend but rather reflect normal fluctuations in physiological parameters associated with plants grown at a variety of locations. Furthermore, when data from both the U.S. and
24
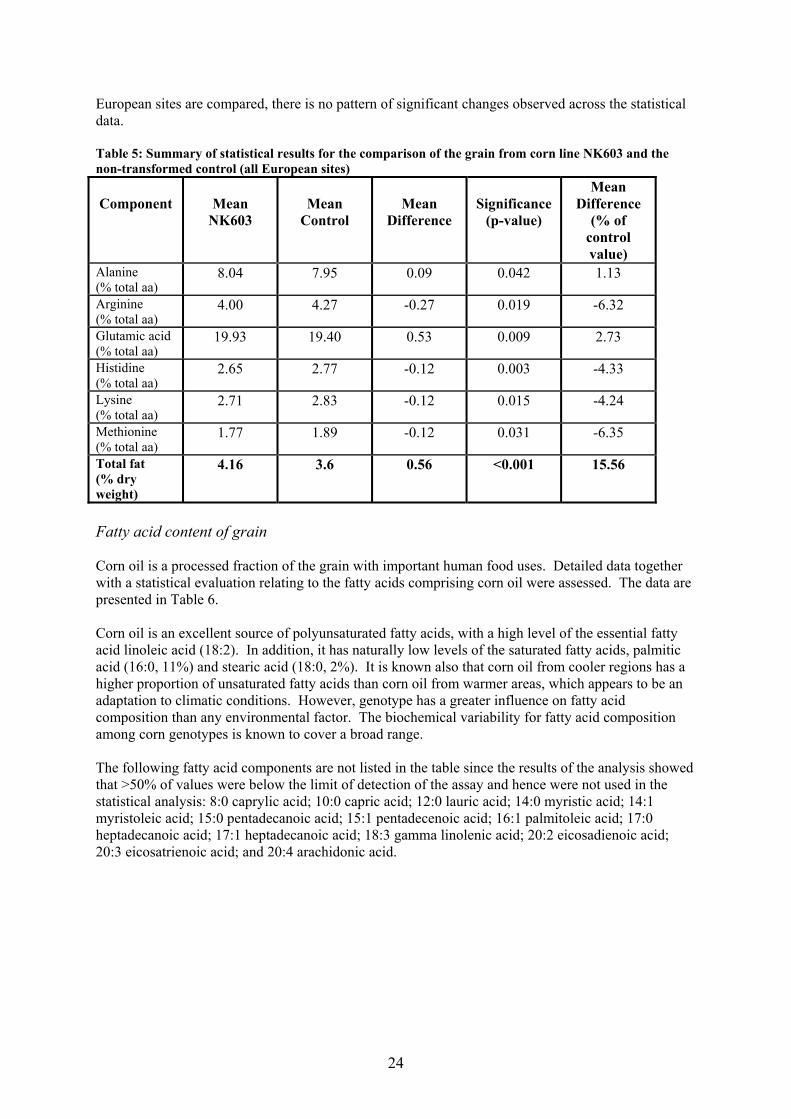
European sites are compared, there is no pattern of significant changes observed across the statistical data. Table 5: Summary of statistical results for the comparison of the grain from corn line NK603 and the non-transformed control (all European sites)
Component
Mean
NK603
Mean
Control
Mean
Difference
Significance
(p-value)
Mean Difference
(% of control value)
Alanine (% total aa)
8.04 7.95 0.09 0.042 1.13
Arginine (% total aa)
4.00 4.27 -0.27 0.019 -6.32
Glutamic acid (% total aa)
19.93 19.40 0.53 0.009 2.73
Histidine (% total aa)
2.65 2.77 -0.12 0.003 -4.33
Lysine (% total aa)
2.71 2.83 -0.12 0.015 -4.24
Methionine (% total aa)
1.77 1.89 -0.12 0.031 -6.35
Total fat (% dry weight)
4.16 3.6 0.56 <0.001 15.56
Fatty acid content of grain Corn oil is a processed fraction of the grain with important human food uses. Detailed data together with a statistical evaluation relating to the fatty acids comprising corn oil were assessed. The data are presented in Table 6. Corn oil is an excellent source of polyunsaturated fatty acids, with a high level of the essential fatty acid linoleic acid (18:2). In addition, it has naturally low levels of the saturated fatty acids, palmitic acid (16:0, 11%) and stearic acid (18:0, 2%). It is known also that corn oil from cooler regions has a higher proportion of unsaturated fatty acids than corn oil from warmer areas, which appears to be an adaptation to climatic conditions. However, genotype has a greater influence on fatty acid composition than any environmental factor. The biochemical variability for fatty acid composition among corn genotypes is known to cover a broad range. The following fatty acid components are not listed in the table since the results of the analysis showed that >50% of values were below the limit of detection of the assay and hence were not used in the statistical analysis: 8:0 caprylic acid; 10:0 capric acid; 12:0 lauric acid; 14:0 myristic acid; 14:1 myristoleic acid; 15:0 pentadecanoic acid; 15:1 pentadecenoic acid; 16:1 palmitoleic acid; 17:0 heptadecanoic acid; 17:1 heptadecanoic acid; 18:3 gamma linolenic acid; 20:2 eicosadienoic acid; 20:3 eicosatrienoic acid; and 20:4 arachidonic acid.
25
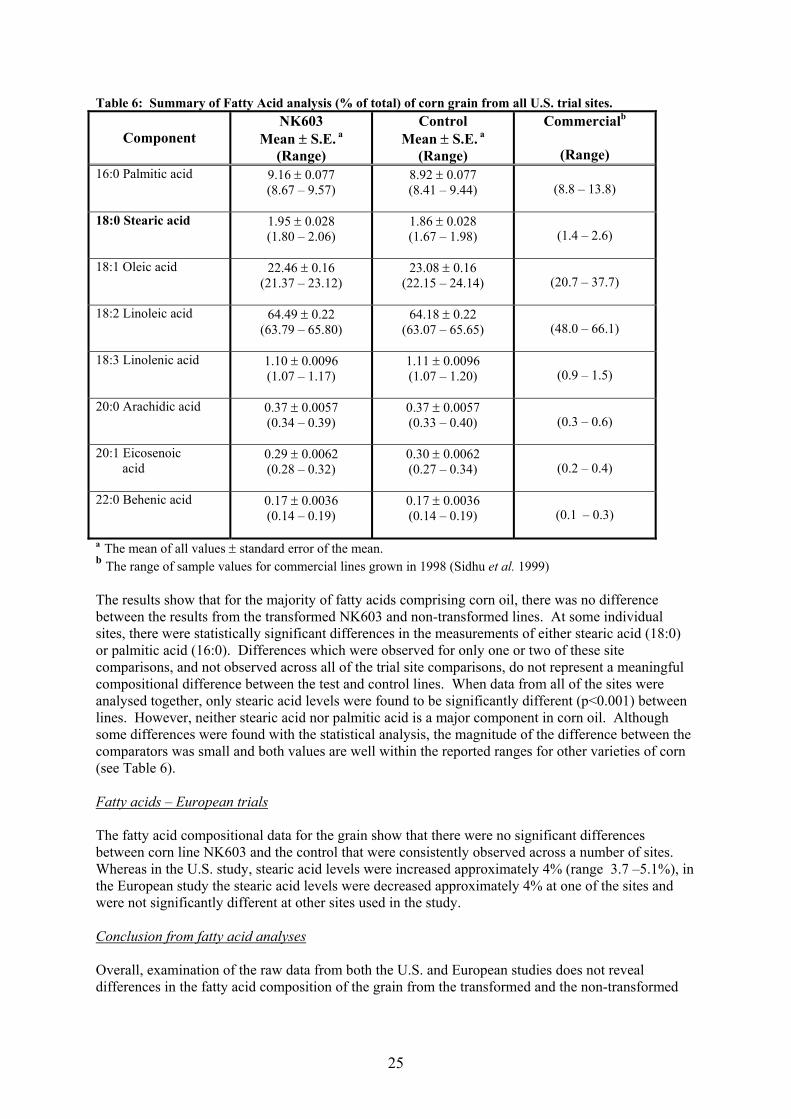
Table 6: Summary of Fatty Acid analysis (% of total) of corn grain from all U.S. trial sites.
Component
NK603 Mean ± S.E. a
(Range)
Control Mean ± S.E. a
(Range)
Commercialb
(Range) 16:0 Palmitic acid
9.16 ± 0.077 (8.67 – 9.57)
8.92 ± 0.077 (8.41 – 9.44)
(8.8 – 13.8)
18:0 Stearic acid
1.95 ± 0.028 (1.80 – 2.06)
1.86 ± 0.028 (1.67 – 1.98)
(1.4 – 2.6)
18:1 Oleic acid
22.46 ± 0.16 (21.37 – 23.12)
23.08 ± 0.16 (22.15 – 24.14)
(20.7 – 37.7)
18:2 Linoleic acid
64.49 ± 0.22 (63.79 – 65.80)
64.18 ± 0.22 (63.07 – 65.65)
(48.0 – 66.1)
18:3 Linolenic acid
1.10 ± 0.0096 (1.07 – 1.17)
1.11 ± 0.0096 (1.07 – 1.20)
(0.9 – 1.5)
20:0 Arachidic acid
0.37 ± 0.0057 (0.34 – 0.39)
0.37 ± 0.0057 (0.33 – 0.40)
(0.3 – 0.6)
20:1 Eicosenoic acid
0.29 ± 0.0062 (0.28 – 0.32)
0.30 ± 0.0062 (0.27 – 0.34)
(0.2 – 0.4)
22:0 Behenic acid
0.17 ± 0.0036 (0.14 – 0.19)
0.17 ± 0.0036 (0.14 – 0.19)
(0.1 – 0.3)
a The mean of all values ± standard error of the mean. b The range of sample values for commercial lines grown in 1998 (Sidhu et al. 1999) The results show that for the majority of fatty acids comprising corn oil, there was no difference between the results from the transformed NK603 and non-transformed lines. At some individual sites, there were statistically significant differences in the measurements of either stearic acid (18:0) or palmitic acid (16:0). Differences which were observed for only one or two of these site comparisons, and not observed across all of the trial site comparisons, do not represent a meaningful compositional difference between the test and control lines. When data from all of the sites were analysed together, only stearic acid levels were found to be significantly different (p<0.001) between lines. However, neither stearic acid nor palmitic acid is a major component in corn oil. Although some differences were found with the statistical analysis, the magnitude of the difference between the comparators was small and both values are well within the reported ranges for other varieties of corn (see Table 6). Fatty acids – European trials The fatty acid compositional data for the grain show that there were no significant differences between corn line NK603 and the control that were consistently observed across a number of sites. Whereas in the U.S. study, stearic acid levels were increased approximately 4% (range 3.7 –5.1%), in the European study the stearic acid levels were decreased approximately 4% at one of the sites and were not significantly different at other sites used in the study. Conclusion from fatty acid analyses Overall, examination of the raw data from both the U.S. and European studies does not reveal differences in the fatty acid composition of the grain from the transformed and the non-transformed
26
lines that are indicative of a systemic change. The data are explained by the known natural variation in composition due to a broad range of factors that influence plant growth and biochemistry. Inorganic analysis Measurements pertaining to inorganic components included the levels of nine minerals. The results of the mineral analysis are presented in Table 7. Sodium has been omitted as greater than 50% of the values were found to be below the limit of detection. The statistical analysis showed that measurements for calcium, magnesium and phosphorus varied slightly between transformed and non-transformed lines at some sites, but the differences were not consistently observed across all sites. Overall, the data show that there were no statistically significant differences in mineral components of the NK603 corn and control lines. Additional analysis Vitamin E (tocopherol) occurs primarily in wheat seedlings, and has been isolated from wheat seedling oil. It is also present in lettuce, celery, cabbage, corn, palm oil, ground nuts, soybeans, castor oil and butter. Although there are numerous structural isomers of tocopherol, biologically, α- tocopherol is the most important member of the group. The vitamin E content of the grain from transformed line NK603 corn and the non-transformed control, which are presented below in Table 8. The reference substance used for the assay was USP alpha tocopherol, 100%, lot number L1. The results show that the genetic modification in line NK603 did not result in any change to the naturally occurring low levels of vitamin E in corn. Table 7: Summary of mineral analysis of corn grain from all trial sites in the study.
Component
NK603 Mean ± S.E. a
(Range)
Control Mean ± S.E. a
(Range)
Commercialb
(Range) Calcium (%)
0.0047 ± 0.00021 (0.0037 – 0.0056)
0.0044 ± 0.00021 (0.0033 – 0.0058)
(0.003 – 0.009)
Copper (mg/kg dw)
1.81 ± 0.090 (1.19 – 2.37)
1.92 ± 0.090 (1.50 – 2.33)
(0.9 – 2.8)
Iron (mg/kg dw)
22.69 ± 0.76 (19.08 – 25.94)
22.93 ± 0.76 (18.77 – 26.62)
(11 – 49)
Magnesium (%)
0.12 ± 0.0021 (0.11 – 0.13)
0.12 ± 0.0021 (0.11 – 0.13)
(0.08 – 0.2)
Manganese (mg/kg dw)
6.26 ± 0.32 (4.64 – 9.63)
6.25 ± 0.32 (4.96 – 8.83)
(2.6 – 7.8)
Phosphorus (%)
0.36 ± 0.0046 (0.32 – 0.39)
0.36 ± 0.0046 (0.32 – 0.39)
(0.24 – 0.43)
Potassium (%)
0.37 ± 0.0057 (0.35 – 0.39)
0.37 ± 0.0057 (0.34 – 0.41)
(0.29 – 0.53)
Zinc (mg/kg dw)
29.28 ± 0.88 (20.23 – 33.17)
29.66 ± 0.88 (23.47 – 33.26)
(15 – 33)
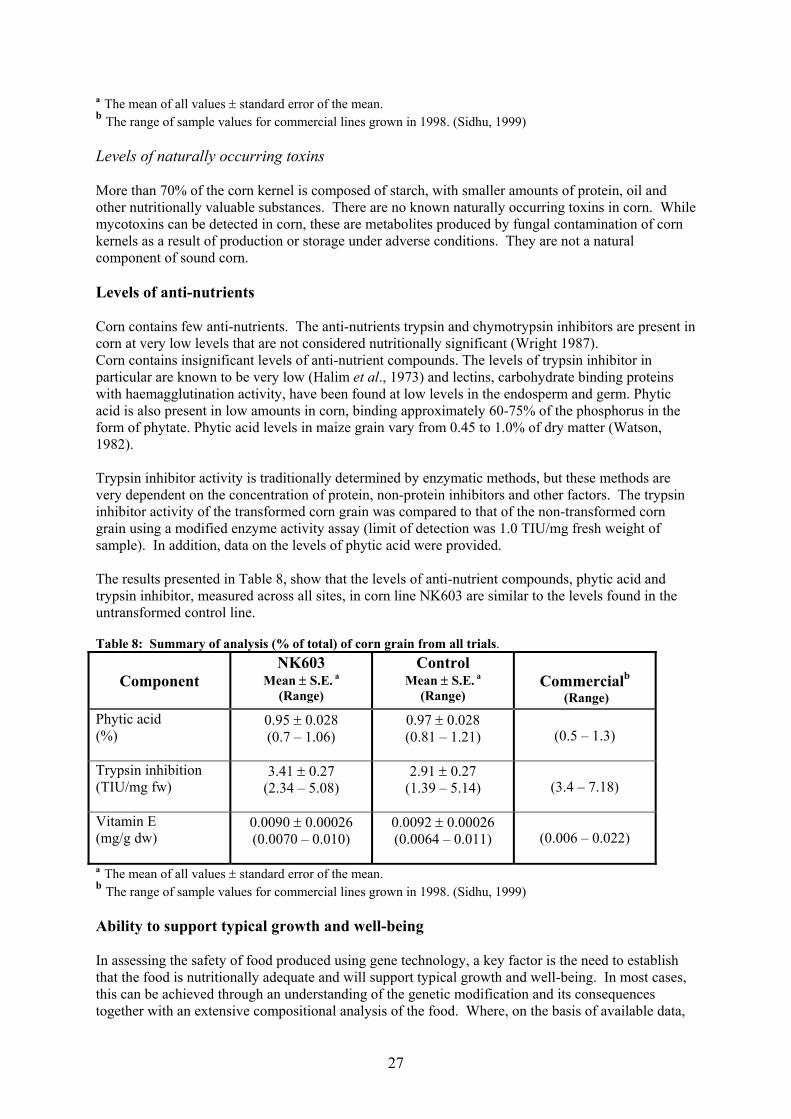
27
a The mean of all values ± standard error of the mean. b The range of sample values for commercial lines grown in 1998. (Sidhu, 1999) Levels of naturally occurring toxins More than 70% of the corn kernel is composed of starch, with smaller amounts of protein, oil and other nutritionally valuable substances. There are no known naturally occurring toxins in corn. While mycotoxins can be detected in corn, these are metabolites produced by fungal contamination of corn kernels as a result of production or storage under adverse conditions. They are not a natural component of sound corn. Levels of anti-nutrients Corn contains few anti-nutrients. The anti-nutrients trypsin and chymotrypsin inhibitors are present in corn at very low levels that are not considered nutritionally significant (Wright 1987). Corn contains insignificant levels of anti-nutrient compounds. The levels of trypsin inhibitor in particular are known to be very low (Halim et al., 1973) and lectins, carbohydrate binding proteins with haemagglutination activity, have been found at low levels in the endosperm and germ. Phytic acid is also present in low amounts in corn, binding approximately 60-75% of the phosphorus in the form of phytate. Phytic acid levels in maize grain vary from 0.45 to 1.0% of dry matter (Watson, 1982). Trypsin inhibitor activity is traditionally determined by enzymatic methods, but these methods are very dependent on the concentration of protein, non-protein inhibitors and other factors. The trypsin inhibitor activity of the transformed corn grain was compared to that of the non-transformed corn grain using a modified enzyme activity assay (limit of detection was 1.0 TIU/mg fresh weight of sample). In addition, data on the levels of phytic acid were provided. The results presented in Table 8, show that the levels of anti-nutrient compounds, phytic acid and trypsin inhibitor, measured across all sites, in corn line NK603 are similar to the levels found in the untransformed control line. Table 8: Summary of analysis (% of total) of corn grain from all trials.
Component
NK603 Mean ± S.E. a
(Range)
Control Mean ± S.E. a
(Range)
Commercialb
(Range) Phytic acid (%)
0.95 ± 0.028 (0.7 – 1.06)
0.97 ± 0.028 (0.81 – 1.21)
(0.5 – 1.3)
Trypsin inhibition (TIU/mg fw)
3.41 ± 0.27 (2.34 – 5.08)
2.91 ± 0.27 (1.39 – 5.14)
(3.4 – 7.18)
Vitamin E (mg/g dw)
0.0090 ± 0.00026 (0.0070 – 0.010)
0.0092 ± 0.00026 (0.0064 – 0.011)
(0.006 – 0.022)
a The mean of all values ± standard error of the mean. b The range of sample values for commercial lines grown in 1998. (Sidhu, 1999) Ability to support typical growth and well-being In assessing the safety of food produced using gene technology, a key factor is the need to establish that the food is nutritionally adequate and will support typical growth and well-being. In most cases, this can be achieved through an understanding of the genetic modification and its consequences together with an extensive compositional analysis of the food. Where, on the basis of available data,

28
there is still concern or doubt in this regard, carefully designed feeding studies in animals may provide further re-assurance that the food is nutritionally adequate. Such studies may be considered necessary where the compositional analysis indicates significant differences in a number of important components or nutrients or where there is concern that the bioavailability of key nutrients may be compromised by the nature of the genetic changes to the food. In view of the compositional data available for corn line NK603 and the technical features of the genetic modification, animal studies were not considered essential to demonstrate the wholesomeness and nutritional adequacy of this food. Nevertheless, several feeding studies using transformed and non-transformed corn grain were provided as additional supporting information and have been included in the safety assessment. Studies evaluated: Dudek, B.R., 2001. 13 Week Feeding Study in Rats with Grain from Roundup Ready Corn (NK603) Preceded by a 1-Week Baseline Food Consumption Determination with PMI Certified Rodent Diet #5002. Project Number: MSE-N 99091, ML-99-253, MSL 17423, MSL 17555. George, B. et al., 2001. Comparison of Broiler Performance When Fed Diets Containing Event NK603, Parental Line or Commercial Corn. Monsanto Study No. 2000-01-39-02. Stanisiewski, E.P., Hartnell, G.F. Fischer, R.L. and Lewis, A.J., 2002. Performance of Pigs Fed Diets Containing Roundup Ready (NK603), Non-Transgenic Control or Conventional Corn Grown During 2000 In Nebraska. MSL 17500 . Feeding study in Broiler Chickens Rapidly growing broiler chickens are sensitive to changes in nutrient quality in diets, and therefore serve as a useful model species to evaluate the wholesomeness of protein/amino acid sources. This study compares the broiler performance and processing parameters of rapidly growing broiler chickens (Gallus domesticus) raised on a diet containing either corn line NK603, the non-transformed parental corn line (B73HTxLH82), or one of five commercially available reference corn lines, over approximately 43 days. Grain from the NK603 and parental lines was produced in field sites in Hawaii, while grain from the five reference lines was produced either in Hawaii or in other locations during the 1999/2000 growing season. All diets were formulated to meet nutritional recommendations, based on individual nutrient analyses for the grain from each test line and control, and to align them with traditional broiler industry uses. From days 1-20, chickens were fed a starter diet containing approximately 55% w/w corn (crude protein ranging from 20.7% – 21.9%). From days 20-42, chickens were fed a grower/finisher diet containing approximately 60% w/w corn. These dietary corn concentrations are within the range used by commercial poultry growers in the United States. No growth promotants or other medications were added to the test diets which were provided ad libitum. The birds, a high-yielding commercial strain (Ross x Ross 508), were one day of age at the beginning of the study, and were separated by gender and randomly assigned to treatments. For each treatment group, there were 100 birds (50 males and 50 females) in 10 pens (10 birds/pen), giving a total of 700 birds. During the course of the study, the birds were examined twice daily for general health, and any abnormal health symptoms were recorded. Any birds sacrificed were weighed, and any deaths were necropsied to determine the possible cause of death. As much as possible, environmental conditions simulated commercial conditions for raising broilers to market weight (around 2 kg) in approximately 42 days.

29
At study termination (day 43 for males, day 44 for females), carcass measurements were taken including those for fat pads which were collected from each bird and weighed. Meat quality assays on breast and thigh meat samples were subsequently conducted. Statistical analyses were performed on starting and final live weights, feed consumption, feed efficiency, chill weight, percent chill weight (for breast, wing, thigh and drum), as well as moisture, protein and fat for breast and thigh meat. Results The rate of chick mortality was at expected levels (average of 1.14% across groups) for commercial feeding trials and was randomly distributed across all treatments. All performance parameters measured were similar for all of the diets, including the NK603 corn, the non-transformed parental corn and commercial reference lines, as well as being comparable to published literature values for Ross x Ross broiler strains. In particular, live weight at day 0 and day 42, total feed intake and feed efficiency were similar across all treatments. Furthermore, no differences were observed in the percentage of moisture, protein and fat in breast meat or in the percentage of protein and fat in thigh meat across treatment diets. Finally, no differences were observed between the treatment groups in terms of wing weight measurements. The results of the broiler feeding study show that there were no differences in parameters tested between birds fed a diet containing corn line NK603 and the non-transformed parental line (B73HT x LH82). In addition, when individual treatment comparisons were made, broiler chickens in general performed and had similar carcass yield and meat composition with diets containing NK603, the parental control, or five commercially available reference lines. The results support the conclusion that there are no differences between the non-transformed control and transformed corn line NK603 in terms of the ability to provide adequate nutrition to rapidly growing broiler chickens. Feeding study in grower and finisher swine This study compares growth performance and carcass quality measurements in growing-finishing pigs provided diets containing either corn line NK603, the non-transformed control corn, or two commercial reference sources of non-genetically modified corn. While the intent of the study was to confirm the nutritional value of NK603 corn for supporting pig performance in a commercial sense, pigs have been widely used in pharmaceutical research and are considered valuable models for assessing the health effects of dietary micronutrients given the similarity of their digestive and cardiovascular systems to that of humans. The experiment used 72 animals of each gender with an initial body weight of 22.6 ± 0.03 kg. Animals were allotted to treatments randomly such that both genders received all four corn hybrids. The animals were sacrificed when the average body weight had reached 116 kg. The nutrient composition of the corn was similar across all lines used in the study in terms of crude protein and total digestible nutrients. Corn was incorporated into the diets at 68.1% (grower1), 74.2% (grower2), 78.1% (finisher1) and 81.8% (finisher2), along with de-hulled soybean meal. Ultrasound measurements of back fat and loin area were taken on the final day of the experiment. Carcass quality measurements were made 24 hours post-mortem. Most parameters measured, including average daily gain (ADG), average daily feed intake (ADFI) and feed efficiency (ADG/ADFI), were not affected by diet but showed an expected difference between the males and females. Loin muscle quality and composition (protein, fat and water percentages) were similar among diets and between genders. In summary, there were no differences between the test and control/reference corn lines used in this study in terms of the growth performance and carcass measurements in growing-finishing pigs. The

30
conclusion from this study is that corn line NK603 is equivalent to the non-transformed corn varieties in terms of its ability to support adequate nutrition in these animals. Feeding study in rats This study was undertaken to compare the responses of laboratory rats (Sprague Dawley) when fed either a diet containing grain from glyphosate-tolerant corn line NK603, or one of several control diets containing either grain from the non-transformed parental variety or from one of a series of non-transformed commercial corn hybrids (designated as reference controls). Young animals (6 weeks of age, 20 rats/gender/diet group) were assigned to one of the following diets for a period of 13 weeks:
11% or 33% (wt/wt) NK603 corn; 11% or 33% (wt/wt) parental control corn; or 33% (wt/wt) reference control corn grain (six commercial hybrids tested).
There were a total number of 10 diet groups involving 400 rats in the study. In the diets composed of 11% test grain (NK603 or parental line), the formulated diet was supplemented with 22% corn grain from a non-transformed commercial hybrid to bring the total corn content in these groups to the standard 33% used in this experiment. The grain samples and diets were analysed for nutrient composition and residues (for example, pesticide residues and mycotoxins). All diets were balanced for similar fat and protein content. Certified rodent diet was administered during week 1 of the study to establish baseline food consumption data for each animal and was followed by administration of the test and control diets from weeks 2 to 14. Food consumption was determined daily for days 1, 2, 3 and during days 4-7 for each of the first two weeks of the study. Following week 2, food consumption was measured weekly. All animals were observed twice daily for morbidity and moribundity. Body weight was recorded at weekly intervals. After 5 and 14 weeks, blood and urine were collected from 10 animals/gender/group for blood chemistry, haematology and qualitative and quantitative urine analyses. Coagulation parameters were determined at the terminal blood collection only. After 14 weeks, all animals were sacrificed and necropsied. Specified tissues were collected according to the protocol and organs were weighed. Selected tissues were examined microscopically. Observations and results There were two mortalities during the study, one from the high dose NK603 male group (at day 82) and the second from a reference control male group (at day 86). Neither death was considered to be diet or treatment related. There were no other adverse clinical reactions observed during the course of the study. The results of the herbicide analysis show that the glyphosate residue in the test grain was 0.09 ppm, slightly above the analytical detection limit of 0.05 ppm. The parent and reference lines were not assayed for glyphosate. The growth of male and female rats fed NK603 corn grain was comparable to that of rats fed grain from the parental control and reference control groups. Body weight gain and food consumption were comparable across all groups. Organ weights were similar across test and control groups and gross pathology findings were unremarkable in test groups and comparable to control groups. The majority of clinical pathology parameters (chemistry, haematology, coagulation, urinalysis) were similar across all groups with only a few exceptions. A closer examination of the few statistically significant differences in clinical parameters demonstrated that these were artefacts of various statistical calculations and were not considered biologically meaningful as they were either not dose related, or the values were within the range of the reference controls. Microscopic examination of tissues

31
showed no differences between rats fed diets containing 33% NK603 corn grain compared to those fed diets with 33% non-transformed grain. In summary, the rats fed grain composed of corn line NK603 responded similarly to those animals fed with parental control and reference control grain diets. Conclusion Comprehensive data from a range of compositional analyses conducted on grain from glyphosate treated corn line NK603 and the non-transformed control were presented for assessment. The compositional components measured included proximates (protein, fat, ash, carbohydrates, moisture, acid detergent fibre and neutral detergent fibre), amino acid composition, fatty acids, and inorganic mineral analysis. The comparison of results and data from the test and control lines demonstrates that there are no compositional differences of biological significance between corn line NK603 and the non-transformed control in any of the components tested. Minor differences observed were not considered to be of concern with respect to food safety as the levels were well within published ranges that are normally expected of commercial corn varieties. Any variations between lines that were found following statistical evaluation of the data were small in magnitude, occurred at random across the trial sites, and were not indicative of a trend that might point to the existence of an unintended effect as a result of the genetic modification. Overall, as is expected with analyses of crop plants including corn, the compositional variation in a line across localities may be greater than the variation between different lines grown at the same site. Finally, three separate feeding studies using broiler chickens, pigs and laboratory rats to test the nutritional adequacy of corn line NK603 compared to its parental control and other commercial control lines showed that there were no differences between the transformed and non-transformed control corn in terms of the ability to support typical growth and nutritional well-being when incorporated into animal diets. ACKNOWLEDGEMENTS FSANZ gratefully acknowledges Dr Michael Ayliffe, Senior Research Scientist at CSIRO Plant Industry, for his time in reviewing this safety assessment report and for providing his expert comments.

32
REFERENCES Astwood, J.D. and Fuchs, R.L. (1996). Allergenicity of foods derived from transgenic plants. In Ortolani, C. and Wuthrich, B. (eds.) Highlights in food allergy. Monographs in Allergy, 32: 105-120. Barry, G., Kishore, G., Padgette, S., Taylor, M., Kolacz, K., Weldon, M., Re, D., Eichholtz, D., Fincher, K. and Hallas, L. (1992). Inhibitors of amino acid biosynthesis: strategies for imparting glyphosate tolerance to crop plants, pp 139-145. In Biosynthesis and Molecular Regulation of Amino Acids in Plants, Singh et al. (eds), American Society of Plant Physiologists. Fraley, R.T., Rogers, S.G., Horsch, R. B., Sanders, P.R., Flick, J.S., Adams, S.P., Bittner, M.L., Brand, L.A., Fink, C.L., Fry, J.S., Galluppi, G.R., Goldberg, S.B., Hoffman, N.L. and Woo S.C. (1983). Expression of bacterial genes in plant cells. Proc. Natl. Acad. Sci. USA, 80: 4803-4807. Hammond, B.G., Vicini, J.L., Hartnell, G.F., Naylor, M.W., Knight, C.D., Robinson, E.H., Fuchs, R.L. and Padgette, S.R. (1996) The Feeding Value of Soybeans Fed to Rats, Chickens, Catfish and Dairy Cattle Is Not Altered by Genetic Incorporation of Glyphosate Tolerance. J. Nutr., 126: 717-727. Harrison, L.A., Bailey, M.R., Naylor, M.W., Ream, J.E., Hammond, B.G., Nida, D.L., Burnette, B.L., Nickson, T.E., Mitsky, T.A., Taylor, M.L., Fuchs, R.L. and Padgette, S.R. (1996). The Expressed Protein In Glyphosate-Tolerant Soybean, 5-Enolpyruvylshikimate-3-Phosphate Synthase from Agrobacterium sp. Strain CP4 Is Rapidly Digested in Vitro and is not Toxic to Acutely Gavaged Mice. The Journal of Nutrition, 126 (3), 728-740. (The American Institute of Nutrition). Halim, A.H., Wassom, C.E. and Mitchell, H.L., 1973. Trypsin inhibitor in corn (Zea mays L.) as influenced by genotype and moisture stress. Crop Sci. 13, 405-406. Kimber, I., Kerkvliet, N.I., Taylor, S.L., Astwood, J.D., Sarlo, K. and Dearman R.J. (1999). Toxicology of Protein Allergenicity: Prediction and Characterisation. Toxicological Sciences 48: 157-162. McElroy, D., Zhang, W., Cao, J. and Wu, R., (1990). Isolation of an Efficient Actin Promoter for Use in Rice Transformation. Plant Cell, 2, 163-171. Metcalfe, D.D., Astwood, J.D., Townsend, R., Sampson, H.A., Taylor, S.L. and Fuchs, R.L., (1996). Assessment of the allergenic potential of foods derived from genetically engineered crop plants. Critical Reviews in Food Science and Nutrition 36(S): S165-S186. Odell, J.T., Nagy, F. and Chua, N-H. (1985). Identification of DNA sequences required for activity of the cauliflower mosaic virus 35S promoter. Nature 313, 810-812. Organisation for Economic Cooperation and Development (OECD) (1999). Consensus Document on General Information Concerning the Genes and their Enzymes that Confer Tolerance to Glyphosate Herbicide. Series on Harmonisation of Regulatory Oversight in Biotechnology No. 10. OECD ENV/JM/MONO(99)9. Padgette, S. R., D. B. Re, D. B., Barry, G. F., Eichholtz D. A., Delannay X., Fuchs R. L., Kishore G. M., and R. T. Fraley. 1996. New Weed Control Opportunities: Development of Soybeans with a Roundup Ready™ Gene. In Herbicide Resistant Crops. Edited by S. O. Duke. C.R.C Press Inc. p. 53-84. Pichersky, E., Logsdon Jr, J.M., Mitchell McGrath, J. and Stasys, R.A. (1991). Fragments of plastid DNA in the nuclear genome of tomato: prevalence, chromosomal location, and possible mechanism of integration. Mol.Gen.Genet., 225, 453-458. Schultz, A., Kruper, A. and Amrhein, N. (1985). Differential sensitivity of bacterial 5-enolpyruvylshikimate-3-phosphate synthases to the herbicide glyphosate. FEMS Microbiology Letters, 28, 297-301. Wright, K.N., (1987). Nutritional properties and feeding value of corn and its by-products, pp 447-478. In Corn: Chemistry and Technology. S.A. Watson and P.E. Ramsted (Eds.), American Association of Cereal Chemists, Inc. St. Paul, Minnesota, USA.

33
Watson, S.A., (1982). Corn: Amazing Maize: General Properties. In CRC Handbook of Processing and Utilization in Agriculture, Volume II: Part 1 Plant Products. I.A. Wolff (ed.) CRC Press Inc., Florida, USA. Watson, S.A., (1987). Structure and Composition, pp. 53-82. In Corn: Chemistry and Technology. S.A. Watson and P.E. Ramstad, (Eds.), American Association of Cereal Chemists, Inc. St. Paul, Minnesota, USA. WHO (1991). Strategies for assessing the safety of foods produced by biotechnology. Report of a joint FAO/WHO Consultation. World Health Organization, Geneva, 59 pp. WHO (1993). Health aspects of marker genes in genetically modified plants. Report of a WHO Workshop. World Health Organization, Geneva, 32 pp. General references Australia New Zealand Food Authority (1999). Guidelines for the safety assessment of foods to be included in Standard A18 – Food Produced Using Gene Technology. Bargman, T.J., Taylor, S.L. and Rupnow, J.H. (1992). Food Allergies. In: Handbook of Natural Toxins Volume 7, Food Poisoning, 337-370. Published by: Marcel Dekker, New York, USA. Davis, B.D., Dulbecco, R., Eisen, H.N. and Ginsberg, H.S. (1980). Microbiology, 3rd Edition. Harper and Row Publishers, USA, 1355 pp. Frick, O.L., 1995. The potential for allergenicity in transgenic foods. In Genetically Modified Foods: Safety Aspects. Editors K.H. Engel, G.R. Takeoka and R. Teranishi. American Chemical Society, Washington DC. Jugenheimer, R.W., 1976. Corn Improvement, Seed Production and Uses. John Wiley and Sons, Inc. New York, New York, USA. Kimber, I., Kerkvliet, N.I., Taylor, S.L., Astwood, J.D., Sarlo, K. and Dearman, R.J., 1999. Toxicology of Protein Allergenicity: Prediction and Characterisation. Toxicological Sciences, 48, 157-162. Kuiper, H.A., Kleter, G.A., Noteborn, H.P.J.M. and Kok, E.J. (2001). Assessment of the food safety issues related to genetically modified foods. The Plant Journal, 27(6), 503-528. Novak, W.K. and Haslberger, A.G., 2000. Substantial Equivalence of Antinutrients and Inherent Plant Toxins in Genetically Modified Novel Foods. Food and Chemical Toxicology, 38, 473-483. World Health Organisation (WHO), 1995. Application of the Principles of Substantial Equivalence to the Safety Evaluation of Foods or Food Components from Plants Derived by Modern Biotechnology. Report of a WHO workshop.