fiziologie, fiziopatologie, biofizic -...
TRANSCRIPT

254
FIZIOLOGIE, FIZIOPATOLOGIE, BIOFIZICĂ
INFLUENŢA PROBELOR RESPIRATORII ASUPRA VARIABILITATEA RITMULUICARDIAC LA SUBIECŢI SANATOŞI CU ANXIETATE JOASĂ ŞI RIDICATĂ
Andrei Ganenco, Victor Vovc, Ion Moldovanu, Svetlana LozovanuCatedra Fiziologia Omului şi Biofizică, USMF “Nicolae Testemiţanu”
Institutul de Neurologie şi Neurochirurgie
SummaryThe influence of respiratory tests on the heart rate variability at healthy
subjects with high and low anxietyThe relationship of heart rate variability to thoracic and abdominal components of
breathing pattern, as well as to slow paced respiration, was studied in healthy volunteers withdifferent level of anxiety. Subjects with high anxiety showed lower levels of heart ratevariability, slow respiration increased the low frequency component of HRV in both groups,chest and abdominal respiration reduced HRV.
RezumatA fost studiată corelaţia dintre dintre variabilitatea ritmului cardiac (VRC) şi
componentele toracică şi abdominală a respiraţiei, precum şi corelaţia dinre VRC şi respiraţialentă dirijată, la subiecţi sănătoşi cu nivel diferit de anxietate. Subiecţii cu anxietate mai înaltă auprezentat valori mai joase a VRC, respiraţie lentă a mărit componenta frecvenţelor joase a VRCîn ambele grupuri, respiraţia abdominală sau toracică au redus VRC.
ActualitateaEste bine cunoscut faptul că Variabilitatea Rirmului Cardiac (VRC) prezintă un
parametru fundamental homeoctatic şi anumite modificări ale acestei variabilităţi, în specialreducerea ei, este un marker de posibilă moarte subită la pacienţii cu boli cardiace (Task Force ofthe European Society of Cardiology and theNorth American Society of Pacing andElectrophysiology, 1996). De asemenea se cunoaşte că respiraţia abdominală are un effectbenefic asupra organismului, de aceea pranayama (respiraţia în cadrul sistemului yoga)tradiţional este considerată ca fiziologică cu un efect catabolic major. Chiar există şi metode detraining respirator pentru a mări variabilitatea ritmului cardiac.
În alt context este bine cunoscut faptul, că anxietatea mărită a subiectului se asociază cutulburări vegetative şi sindromul psiho-vegetativ, descries în secolul trecut de autori germane(Fhiele, 1966) şi dezvoltat ulterior de şcoala profesorului F.Vein. Acest sindrom este consideratdrept un concept fundamental al vegetologiei moderne.
Există un anumit interes faţă de interacţiunea componentelor thoracic şi abdominal alepatternului rspirator (Minyaev, Davydov, 2000). Cercetări experimentale pe sănătoşi şi bolnavicare ar elucida importanţa patternului de respiraţie în diferitele lui variante (respiraţieabdominală, respiraţie toracică, hiperventilaţie) sunt în număr limitat.
Scopul cercetării noastre de a analiza variabilitatea ritmului cardiac la diferite modificărial patternului de respiraţie în funcţie de nivelul de anxietate la voluntarii sănătoşi.
MetodePersoanele incluse în studiul dat au fost selectate în baza scorului obţinut la completarea
testului de anxietate personală şi reactivă Spielberger. 12 subiecţi (6 bărbaţi, 6 femei, vîrsta
255
medie 20.59±0.43) au format lotul cu anxietate joasă (15-30 puncte), iar 13 subiecţi (6 bîrbaţi, 7femei, vîrsta medie 20.49±0.38) au format lotul cu anxietate înaltă (mai mult de 45 depuncte).ECG şi pneumotahografia au fost înregistrate cu unitatea de achiziţie a datelor MP35(BIOPAC systems).
Înregistrările au fost efectuate în următoarele condiţii: respiraţie spontană (3 min),respiraţie spontană cu limitarea mişcărilor toracice (sau respiraţie abdominală, 3 min), respiraţiespontană cu limitarea mişcărilor abdomenului (sau respiraţie toracică, 3 min) şi respiraţie dirijatăcu metronomul la frecvenţa respiraţiei 6/min (3 min). Simultan, presiunea CO2 în aerul expiratera controlată permanent cu ajutorul capnografului MEC-2000, cu scopul de-a menţine valorilelui constante.
Din ECG, perioada cardiac a fost calculată ca intervalul între 2 unde R succesive, a fostfolosită analiza Fourier pentru calcularea densităţii puterii spectrale şi determinaţi următoriiindici a variabilităţii ritmului cardiac: densitatea puterii spectrale, puterea spectrală absolută şinormalizată a frecvenţelor înalte (HF), puterea spectrală absolută şi normalizată a frecţvenţelorjoase (LF) (valorile normalizate reprezintă valoarea relativă a fiecărui component faţă de putereatotală fără componenta frecvenţelor foarte joase) şi raportul LF/HF.
Analiza statistică. T-testul a fost folosit pentru determinarea diferenţei statistice a puteriivariabilităţii cardiace şi componentelor sale în diferite condiţii. Valorile sunt reprezentate camedii±erori dacă nu se specifică altceva.
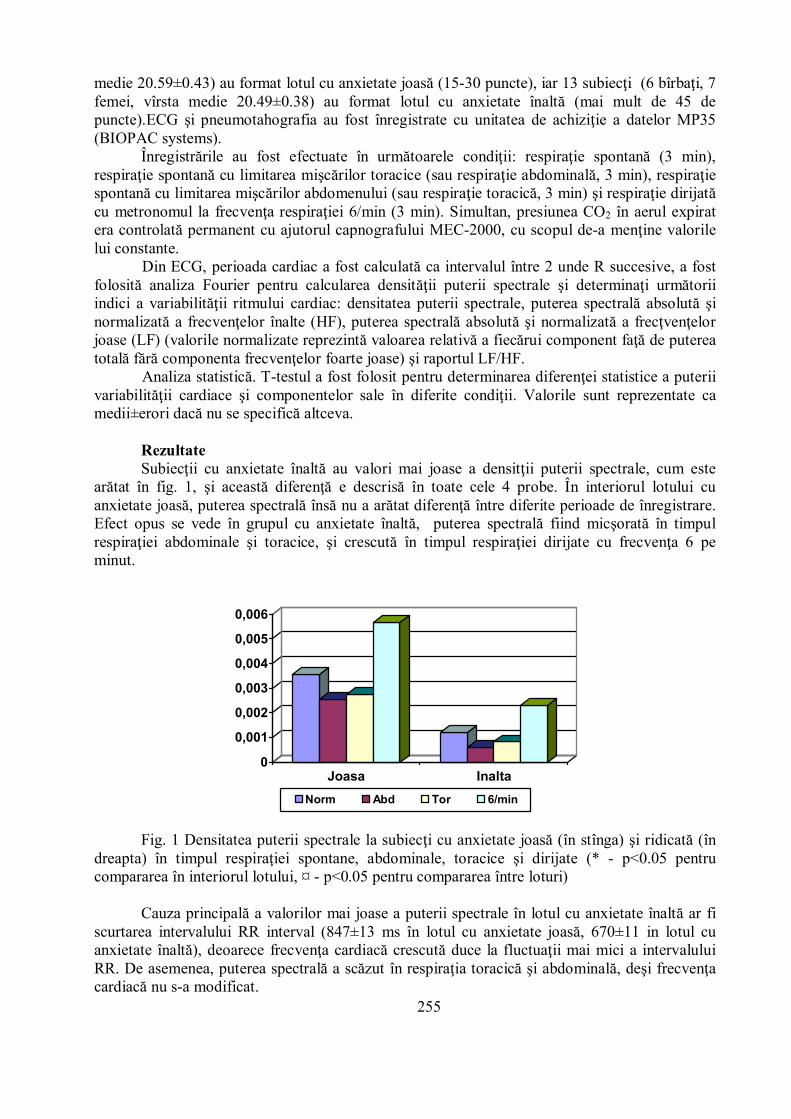
RezultateSubiecţii cu anxietate înaltă au valori mai joase a densitţii puterii spectrale, cum este
arătat în fig. 1, şi această diferenţă e descrisă în toate cele 4 probe. În interiorul lotului cuanxietate joasă, puterea spectrală însă nu a arătat diferenţă între diferite perioade de înregistrare.Efect opus se vede în grupul cu anxietate înaltă, puterea spectrală fiind micşorată în timpulrespiraţiei abdominale şi toracice, şi crescută în timpul respiraţiei dirijate cu frecvenţa 6 peminut.
0
0,001
0,002
0,003
0,004
0,005
0,006
Joasa InaltaNorm Abd Tor 6/min
Fig. 1 Densitatea puterii spectrale la subiecţi cu anxietate joasă (în stînga) şi ridicată (îndreapta) în timpul respiraţiei spontane, abdominale, toracice şi dirijate (* - p<0.05 pentrucompararea în interiorul lotului, ¤ - p<0.05 pentru compararea între loturi)
Cauza principală a valorilor mai joase a puterii spectrale în lotul cu anxietate înaltă ar fiscurtarea intervalului RR interval (847±13 ms în lotul cu anxietate joasă, 670±11 in lotul cuanxietate înaltă), deoarece frecvenţa cardiacă crescută duce la fluctuaţii mai mici a intervaluluiRR. De asemenea, puterea spectrală a scăzut în respiraţia toracică şi abdominală, deşi frecvenţacardiacă nu s-a modificat.
256
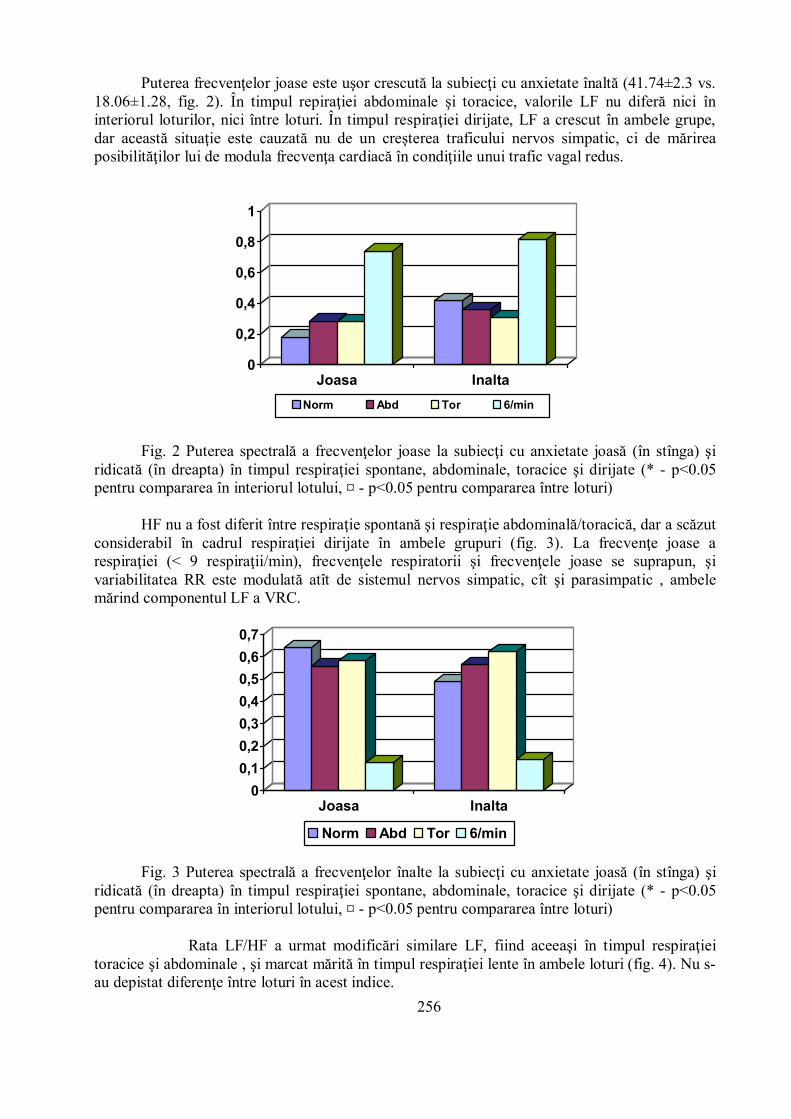
Puterea frecvenţelor joase este uşor crescută la subiecţi cu anxietate înaltă (41.74±2.3 vs.18.06±1.28, fig. 2). În timpul repiraţiei abdominale şi toracice, valorile LF nu diferă nici îninteriorul loturilor, nici între loturi. În timpul respiraţiei dirijate, LF a crescut în ambele grupe,dar această situaţie este cauzată nu de un creşterea traficului nervos simpatic, ci de mărireaposibilităţilor lui de modula frecvenţa cardiacă în condiţiile unui trafic vagal redus.
0
0,2
0,4
0,6
0,8
1
Joasa InaltaNorm Abd Tor 6/min
Fig. 2 Puterea spectrală a frecvenţelor joase la subiecţi cu anxietate joasă (în stînga) şiridicată (în dreapta) în timpul respiraţiei spontane, abdominale, toracice şi dirijate (* - p<0.05pentru compararea în interiorul lotului, ¤ - p<0.05 pentru compararea între loturi)
HF nu a fost diferit între respiraţie spontană şi respiraţie abdominală/toracică, dar a scăzutconsiderabil în cadrul respiraţiei dirijate în ambele grupuri (fig. 3). La frecvenţe joase arespiraţiei (< 9 respiraţii/min), frecvenţele respiratorii şi frecvenţele joase se suprapun, şivariabilitatea RR este modulată atît de sistemul nervos simpatic, cît şi parasimpatic , ambelemărind componentul LF a VRC.
00,10,20,30,40,50,60,7
Joasa Inalta
Norm Abd Tor 6/min
Fig. 3 Puterea spectrală a frecvenţelor înalte la subiecţi cu anxietate joasă (în stînga) şiridicată (în dreapta) în timpul respiraţiei spontane, abdominale, toracice şi dirijate (* - p<0.05pentru compararea în interiorul lotului, ¤ - p<0.05 pentru compararea între loturi)
Rata LF/HF a urmat modificări similare LF, fiind aceeaşi în timpul respiraţieitoracice şi abdominale , şi marcat mărită în timpul respiraţiei lente în ambele loturi (fig. 4). Nu s-au depistat diferenţe între loturi în acest indice.

257
01234567
Joasa Inalta
Norm Abd Tor 6/min
Fig. 4 Raportul LF/HF la subiecţi cu anxietate joasă (în stînga) şi ridicată (în dreapta) întimpul respiraţiei spontane, abdominale, toracice şi dirijate (* - p<0.05 pentru compararea îninteriorul lotului, ¤ - p<0.05 pentru compararea între loturi)
Concluzii1. Subiecţii cu anxietate mai îanltă prezintă valori mai joase a puterii densităţii spectrale.2. Respiraţia lentă (6/min) măreşte componeta de frecvenţe joase a variabilităţii ritmului
cardiac, deoarece acest component este modulat de interacţiunilie simpatico-vagale.
Bibliografie1. Task Force of the European Society of Cardiology and theNorth American Society of Pacing
and Electrophysiology, Heart rate variability: standards of measurement, physiologicalinterpretation and clinical use, Circulation, vol. 93, 1996, pp. 1043–1065
2. W. Fhiele, Psycho-vegetative syndrome, Ment. Welt., vol. 1, 1966, pp. 9-133. V. I. Minyaev, V. G. Davydov, The role of the thoracic and abdominal components of the
respiratory system during hyperventilation combined with chemoreceptor stimulation ofvarious intensities, Human Physiology, vol. 26, n. 4, 2000, pp. 451-455.
CORELAŢII DINTRE ACTIVITATEA SISTEMULUI CITOKINIC ŞI PROTEINAC–reactivă PE PARCURSUL ŞOCULUI HEMORAGIC
Victoria Rotaru, Eleonora Borş, Corneliu Hangan,Veronica CernitCatedra Fiziopatologie şi fiziopatologie clinică
SummaryCausal correlation between systemic activity of cytokines and C- reactive
protein during hemorrhagic shockHemorrhagic shock takes place by local and systemic inflammatory processes. A
pathogenetic factor of systemic inflammation is acute-phase response. During the acute-phaseresponse, the liver dramatically increases the synthesis of acute-phase proteins, such asfibrinogen and C-reactive protein (CRP). CRP is recognized as a true marker of local anddiffuse inflammatory process. Elevated levels of this protein of acute inflammation duringhemorrhagic shock are caused by elevated concentrations of cytokines IL-1, IL-6, TNF-α.
Rezumat

258
Şocul hemoragic se desfăşoară prin procese inflamatorii locale şi sistemice. Un factorpatogenetic al inflamaţiei sistemice este Răspunsul Fazei Acute (RFA). În timpul RFA în ficatcreşte corelativ sinteza proteinei C- reactive (PCR), fibrinogenului, precum şi nivelul citokinelorîn sânge. PCR este recunoscută drept un marker fidel al procesului inflamator local şi difuz.Valorile crescute ale acestei proteine a fazei acute a inflamaţiei pe perioada şocului hemoragicsunt determinate de concentraţiile elevate ale citokinelor IL-1-α, IL-6 şi TNF-α.
ActualitateaŞocul hemoragic se caracterizează prin modificarea concentraţiei proteinelor fazei acute
cauzate de alterările primare şi/sau secundare a celulelor, precum şi a structurilor extracelulare[1]. Proteina C reactivă (PCR) este o proteină plasmatică sintetizată de ficat şi de adipocite. Eaeste un marker al inflamaţiei sistemice a organismului, ce participă la eliminarea substanţelornecrotice din organism, fixând lipoproteinele cu densitate mică şi foarte mică. Manifestândfuncţie imunomodulatoare, interacţionează cu Fc-receptorii de pe suprafaţa celulelorimunocompetente - monocite, T-killeri, T-supresori şi participă la fixarea ligandului cu activareacomplementului. Proteina C-reactivă inhibă eliberarea citokinelor proinflamatorii prin mecanismde fixare a fosfolipidelor membranare.
Citokinele markeri ai inflamaţiei IL-1α, IL-6 şi TNF-α, sunt sintetizate preponderent decătre monocite şi macrofage [3]. IL-1 modulează diverse procese protective în organism, activateîn leziuni celulare, din care cauză este considerată mediator proinflamator, sintetizat primar lalocul leziunii. Sinteza IL-1 în inflamaţie creşte provocând reacţii sistemice, ceea ce o determinăca important mediator a reacţiei fazei acute [2].
IL-6 se consideră principalul stimulator al sintezei şi secreţiei proteinelor fazei acute decătre hepatocite. Cu toate că rolul IL-6 constă în activarea proceselor restabilirii homeostazieidereglate, sinteza exagerată a acesteea induce lezarea ţesuturilor. Există o corelaţie între gradulcreşterii nivelului IL-6 şi procesul inflamator sistemic [4].
TNF posedă un efect proinflamator marcat, ce se depistează iniţial în locul eliberăriiacestuia. Acesta activează leucocitele, expresează moleculele de adeziune celulară pe membranacelulelor endoteliale la nivelul patului microcirculator, astfel condiţionând migrarea leucocitelordin sânge în matricea extracelulară; stimulează elaborarea speciilor active de oxigen de cătreleucocite; induce secreţia citokinelor proinflamatoare, inclusiv IL-1, IL-8, IL-6 şi interferoni.Creşterea concentraţiei TNF-α induce sinteza proteinelor fazei acute [6].
Obiectivul actualei lucrări este estimarea corelaţiei dintre activitatea sistemului citokinicşi proteinei C -reactive pe parcursul şocului hemoragic.
Material şi metodeÎn experienţele proprii şocul hemoragic a fost reprodus la iepuri masculi de laborator
(masa corporală 2,3-2,5 kg), utilizând modelul clasic [5].Aplicând anestezie locală cu Sol. Novocaină 5%-10 ml, a fost preparată vena femurală.
Sângele a fost exfuzat în 3 prize (câte 15-20 ml la fiecare) la intervale de 15-20 min, volumul desânge pierdut fiind de circa 45-55 ml (5-7% din masa corpului), ceea ce constituie aproximativ40% din volumul total de sânge. Presiunea sanguină finală a fost menţinută la nivel de şoc sever- 40±5 mm Hg. Aceste valori au fost menţinute prin efuzii şi reinfuzii repetate de sânge pe operioadă până la 24 ore. Estimările biochimice ale sângelui s-au efectuat iniţial, la sfârşitulperioadelor de 90 min, 5 ore şi 24 ore ale evoluţiei şocului hemoragic.
Primele 5 ore animalele rămâneau fixate pe măsuţe. După aceasta animalele eraueliberate şi întreţinute în condiţii identice la temperatura de 25-30°C cu acces liber la apă şihrană pentru a studia durata vieţii animalelor din fiecare lot Pe tot parcursul vieţii se studiastarea generală a animalelor. La finele experienţelor animalele au fost sacrificate prin eutanazie.
259
Concentraţia proteinei C-reactive plasmatice ( mg/dl) a fost determinată prin metodaSlide-format (latex).
Concentraţia interleukinelor plasmatice (IL-1α, IL-6, TNFα- pg/ml) a fost cantitativdeterminată în ser, utilizând metoda imunoenzimatică ELISA (Enzyme-Linked ImmunoSorbantAssay).
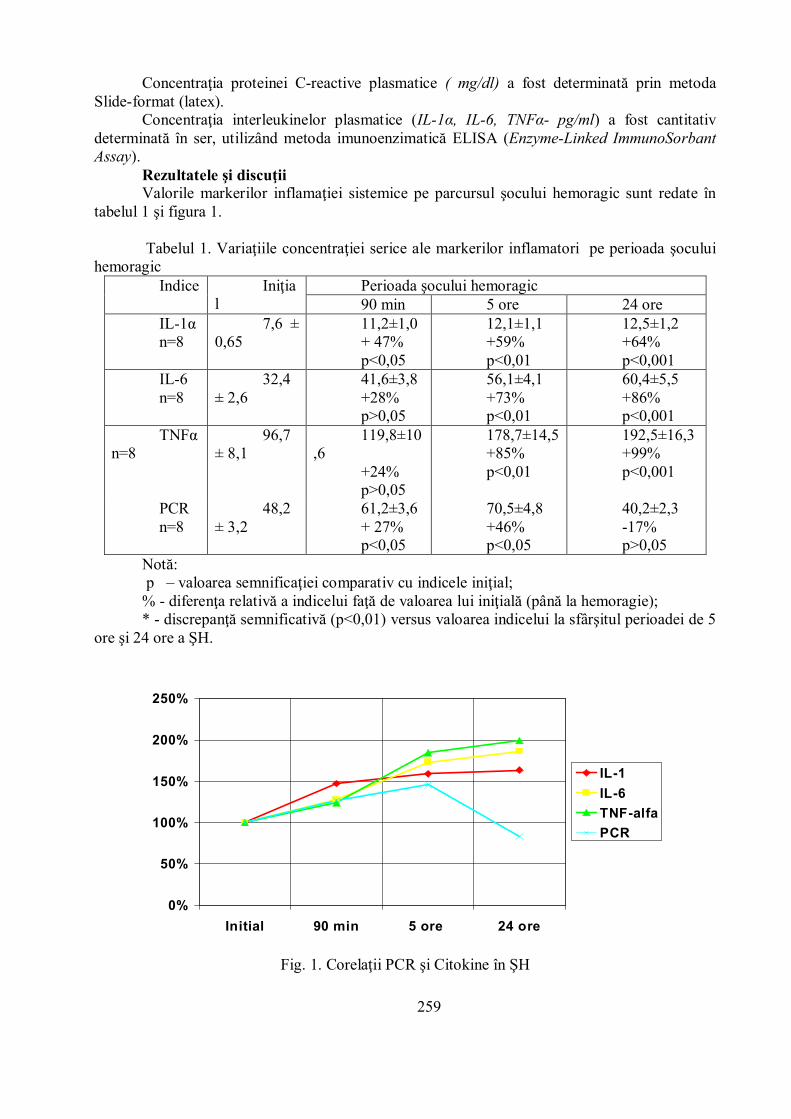
Rezultatele şi discuţiiValorile markerilor inflamaţiei sistemice pe parcursul şocului hemoragic sunt redate în
tabelul 1 şi figura 1.
Tabelul 1. Variaţiile concentraţiei serice ale markerilor inflamatori pe perioada şoculuihemoragic
Perioada şocului hemoragicIndice Iniţial 90 min 5 ore 24 ore
IL-1αn=8
7,6 ±0,65
11,2±1,0+ 47%p<0,05
12,1±1,1+59%p<0,01
12,5±1,2+64%p<0,001
IL-6n=8
32,4± 2,6
41,6±3,8+28%p>0,05
56,1±4,1+73%p<0,01
60,4±5,5+86%p<0,001
TNFαn=8
96,7± 8,1
119,8±10,6
+24%p>0,05
178,7±14,5+85%p<0,01
192,5±16,3+99%p<0,001
PCRn=8
48,2± 3,2
61,2±3,6+ 27%p<0,05
70,5±4,8+46%p<0,05
40,2±2,3-17%p>0,05
Notă: p – valoarea semnificaţiei comparativ cu indicele iniţial;% - diferenţa relativă a indicelui faţă de valoarea lui iniţială (până la hemoragie);* - discrepanţă semnificativă (p<0,01) versus valoarea indicelui la sfârşitul perioadei de 5
ore şi 24 ore a ŞH.
Fig. 1. Corelaţii PCR şi Citokine în ŞH
0%
50%
100%
150%
200%
250%
Initial 90 min 5 ore 24 ore
IL-1IL-6TNF-alfaPCR

260
La cea mai timpurie perioadă evaluată a ŞH (90 min) s-a constatat creşterea semnificativădoar a IL-1α. Incrementul relativ al acesteia în serul sanguin comparativ cu valorile
iniţiale a constituit în medie 47% (p<0,05). IL-6 s-a majorat cu 28%, iar TNF- α a atins cote cu24% peste valoarea iniţială. La perioada de 5 ore a ŞH creşterea sanguină a citokinelor acontinuat. IL-1α a realizat un spor cantitativ mai limitat cu 59% (p<0,01) mai mari vis-a-vis denivelul iniţial. IL-6 a crescut până la o diferenţă de 73% (p<0,01) comparativ cu valoareainiţială, iar TNF-α – şi mai considerabil, cu aproximativ 85% (p<0,01). Estimarea probelorsanguine la sfârşitul perioadei de 24 ore a ŞH a pus în evidenţă unele particularităţi ale dinamiciicitokinelor în ser. De menţionat în acest context o creştere neînsemnată a IL-1α faţă de valoareaatestată la 5 ore, care a măsurat 3,3%, iar comparativ cu nivelul iniţial decalajul a constituit înmedie 64%. Atât IL-6, cât şi
TNF-α au avut un spor de circa 7,7% pe perioada 5-24 ore a ŞH, ceea ce comparativ cuIL-1 este mai mult ca dublu. Faţă de valoarea iniţială IL-6 a crescut pe perioada de 24 ore a ŞHcu 86% (p<0,001), iar TNF-α cu 99% (p<0,01).
Aşadar, evaluarea dinamicii citokinemiei pe perioada de 24 ore de desfăşurare a ŞHevidenţiază creşterea mai rapidă în serul sanguin a IL-1α, aceasta fiind decelată semnificativă lasfârşitul perioadei de 90 min. Ulterior, până la perioada tardivă de 24 ore, IL-1α a elevat numaicu 11,6% şi a demonstrat către acest timp un increment de circa 64%.
IL-6 şi TNF-α s-au impus printr-o creştere concludentă după perioada de 90 min, acesteaatingând nivele circulante cu 73% şi, respectiv, cu 85% peste platoul iniţial la perioada de 5 ore aŞH. Această majorare este mai concludentă decât rata creşterii a IL-1α documentată la sfârşitulperioadei de 24 ore a ŞH. Către acest timp TNF-α a avut valori practic dublate vis-a-vismăsurărilor iniţiale.
Concentraţia proteinei C reactive a crescut statistic semnificativ cu 27% deja la perioadade 90 min a ŞH.
Incrementul acestei proteine a continuat până în perioada de 5 ore, demonstrând unsurplus de 46% comparativ cu valorile iniţiale. În continuare, către 24 ore de ŞH s-au consemnatvalori cantitative reduse, chiar comparativ cu platoul iniţial (-17%, p>0,05). În raport cu valoareaatestată la min 90 a ŞH s-a decelat un recul de circa 32%, iar faţă de valoarea perioadei de 5 orediscrepanţa a constituit 41%.
Dinamica acestui indice denotă modificări atât în geneza stimulilor necesari pentrusinteza proteinei C-reactive, cât şi modificări în capacitatea ficatului de a răspunde la aceştistimuli.
Din datele obţinute se evidenţiază următorul tablou. Iniţial hipoperfuzia generalizată aorganelor, indusă de hipotensiunea şocogenă a generat leziuni celulare şi a indus sinteza şielaborarea de factori stimulatori (fosfolipidele membranare, IL-6), care la nivelul ficatului audeclanşat sinteza de proteină C-reactivă. Ficatul a continuat să sintetizeze activ această proteinăpe parcursul perioadei de 5 ore a ŞH, când valoarea circulantă a acesteia s-a estimat cu 46%peste nivelul iniţial.
Declinul proteinei C-reactive la 24 ore de şoc poate fi explicat prin două fenomene: fie căficatul în această perioadă este compromis şi nu sintetizează proteina la nivel adecvat solicitării,fie că proteina sintetizată de ficat nu este lansată în circulaţie din cauza dereglărilormicrocirculatorii din organ, fiind stocată local în cantităţi considerabile. În acest caz stocul deproteină C-reactivă ar putea fi lansat în circulaţia sistemică pe parcursul resuscitării şi reperfuzieificatului, ceea ce ar avea impact în patogenia sindromului postinfuzional.
Concluzii1. Valorile crescute a PCR pe perioada de 5 ore a şocului hemoragic sunt determinate de
concentraţiile elevate ale IL-1-α, IL-6 şi TNF-α.

261
2. Pe parcursul a 5 ore de şoc hemoragic are loc diminuarea clearance-ului PCR, perioada deînjumătăţire a acesteea constituind 19 ore.
3. Declinul nivelului circulant al PCR depistat după 24 ore de ŞH se datorează afecţiuniiparenchimului hepatic, cauzat de hipoperfuzia hipovolemică.
4. Inflamaţia sistemică pe fondal de ŞH se caracterizează prin creşterea corelativă a citokinelorIL-1α, IL-6, TNF-α şi proteinei C reactive

262
Bibliografie1. DUTTON R. Current concepts of hemorrhagic shock. Anesthesiology Clinic, 2007, 25: 23-
34.2. CHAPMAN M. Hemorrhagic shock. Critical Care, 2004, 8: 373-84.3. KLABUNDE RE. The pathophysiology of hemorrhagic shock. PPT, 2004.4. LIU LL, Dubick MA. Hemorrhagic shock-induced vascular hyporeactivity in the rat:
Relationship to gene expression of nitric oxide synthase, endothelin-1, and select cytokines incorresponding organs. The Journal of surgical research. 2005, 125(2): 128-136.
5. MAURITZ JL, Renedo J, Barrion JP et al. Experimental models of hemorrhagic shock. NutrHosp, 2007, 22: 190-8.
6. HIROAKI S, Kentaro K, Toshiko T et al. Role of tumor necrosis factor-alpha andinterleukin-1beta on lung dysfunction following hemorrhagic shock in rats. Inter Med J ExpClinic Res, 2008, 14(5): 63-69.
REFLECTIONS ABOUT ESTIMATIVE METHOD OF THE ISOLATEPORTAL VEIN CONTRACTILITY
Aurel Saulea, Victor Rotaru, Victor OjogDepartment of Physiology and Biophysics, State University of Medicine and Pharmacy,
Chisinau, R. Moldova
SummaryThe current investigation intended to determine the conditions of portal vein contractions
in rats in isometric regime. The proceeding of object preparation as well as experimental phaseswas described. Main parameters of contractility were highlighted, also the importance of portalvein muscle reactivity at different concentrations of Ca ions. Hypotheses about the origin andmechanism of portal vein muscle contraction were enounced.
RezumatPrezenta investigaţie determină condiţiile contracţiilor venei porte la şobolan în regim
izometric. A fost descrisă desfăşurat manopera de preparare de obiectului de studiu precum şietapele experimentale. S-au evidenţiat indicii principali ai contractilităţii, cît şi importanţareactivitaţii muşchiului venos portal la diferite concentraţii a ionilor de Ca2+. Au fost expuseipoteze privind originea şi mecanismul contracţiei venei porte izolate.
Keywords - Cajal cells, portal vein, smooth muscle
The present study was done on a group of 200 white male rats, Wistar line with about180-200 g weight. All the painful manipulations were done under anesthesia, by using Nembutalin dose of 50 mg/kg of the animal’s weight or thiopental of Na (0,1g/kg).
In fixed terms and after finishing the experiments the animals were slaughtered throughbeheading. In anaesthetized rat the thoracic cavity has been opened, was found the vena portae,after which nearby of the liver’s entrance was applied the first tweezers which compressedplainly the vena (vein). In this way the vessel is stuffed with blood, becoming larger andcomfortable for the manipulations. Then the second tweezers is applied in the lower part of thevessel a little bit higher of confluence of one of its ramification in vena portae. Then withophthalmic scissors an excision is done below the first tweezers (just near by). The excisioncontinues along the vein’s route (longitudinal) making a concrete separation of the conjunctivetissue, without any harm to the vessel wall itself. The vein’s excision is done just after tweezers.

263
All these manipulations had to be done without any additional tension of the vena, in order not tochange its contractility, that is to keep the experimental purity.
So, a vena segment of about 4 mm was obtained. Once being extracted from de operatingfield the given isolated segment together with tweezers was removed into Petri’s box which hasbeen filled with Krebs-Henseleit solution, preventively being heated and oxygenated. Thisproceeding was done as quickly as possible with a maximal exclusion of unfavorable action ofthe air on the vena. It is necessary to underline this because vena portae in comparison withothers represents a vessel which possesses a pace-maker and the harmful influence of theatmospheric air could modify its property. Being put into Petri’s box with the given solution thesegment of the vena portae was again separated, this time more thoroughly, but keeping itsintegrity.
After that the separated vena portae has been transported together with Petri`s box to thedefinite station - the organ’s bath thermostatically isolated. The whole “pilgrimage” of the rat’svena portae, extraction and separation inclusively, with an intermittent station in Petri’s box mustlast as less as possible, for about 5-6 min. This term had to be kept strictly in each case in orderto respect the experimental purity and for a minimum exposure to the atmospheric air.
So, being in the isolated thermostatic organ’s bath an end of the vena portae together withthe tweezers is fixed to an immobile hook near the bath’s bottom. The second end with therespective tweezers is insert by the mechanotron`s arm 6MX1C, which serves as a translator offorce. The position of the vena portae was exactly perpendicular to bath’s bottom. Thethermostatic bath with a volume of 10 ml was perfused with Krebs-Henseleit solution with thefollowing content (mM/L): NaCl - 22,0; KCl - 4,7; NaHCO3 - 15,5; KH2PO4 - 1,2; CaCl - 2,5;MgCl2+6H2O - 1,2; glucoza-11,5; 37oC; the aeration being done with the gas composition O2 -95%; CO2 - 5%; pH-7,3 [1].
The perfusion was prepared just before the experiment from a concentrated solution ofthe above mentioned salts. NaHCO3 and glucose were added at the end. Until the Ca2+ wasadded preventing the sedimentation of the obtained solution CO2 was added.
The vascular isolated preparation was extended till the optimal power of contraction with4 mN. The signal of the mechanotron was transmitted to the amplifier UBP2-03. The recordingof the contractile function was done with autorecorder K 200 (RDG). The spontaneouscontractions of the isolated vena portae were determined in isometric regime [2, 3], thecalibration was done preventively. Before starting the experimental actions the vascularpreparation was stabilized for about 45 min till the complete adaptation “in vitro” conditions.Subsequently the recording was doing during 10 min with the film’s speed of 1min/cm, then itwas changed to 5sec/cm in order to proceed the respective with a 1 min length.
After the end of the recording during 2 min the solution replacement from the isolatedbath organs was done and used another solution; Krebs one, which previously was heated andaerated in the trickling column, the concentration of some ions was modified in dependence andexigency of the studies at every stage of it. In some cases the ingredient or the substance whichwas studied immediately was introduced in the bath of the isolated organs with Krebs solution,as in the group with trypsin “in vitro”. The introduced volume was corrected in such a way thatthe corresponding doze of the substance to be sufficient and the total volume from the bathisolated organs with the respective osmotic pressure to correspond exactly.
Just immediately after vena portae is put in the bath, the recording starts, the regime andexigencies being the same – all 15 min with the specific features which were describedpreviously.
The total time for an experiment from the beginning till the end of the study was about120 min; in such a way the possible metabolic tiredness being avoid in the vein’s segment. Therepeated apparatus calibration was inevitable at the end of all experiments. At last, the segment

264
of the vena portae was weighed in order that the contractility data to be reported to mg/mg ofvein’s weight.
The following indexes were studied (tab.1):- the amplitude contraction (mg/mg) through the contraction force report to the
contractile portion mass of the vena portae;- the contraction’s frequency (contraction/min);- the contraction’s surface (mm2/min);- the intensity of the structures functioning through the amplitude produce to the
contractions frequency (mg/mg/min);- the total contraction time (sec);- the contraction phase (sec);- without relaxations (sec);
Table 1. The contractile index value of the isolated vena portae.
Index Value
Amplitudine,N/mg 35,5±3,28
Frequency, contr/min 6,18±0,4IFS,Nmg/min 219,4±23,7
Surface,mm2/min 919,91±86,6
The total contraction time, sec 5,8±0,75
The contraction phase, sec 2,9±0,37
The relaxation phase, sec 2,9±0,37
Discussions and conclusions1. The mechanotron 6 MXIC, used for measuring the small movements, allows to
determine the phasic contractions of the vena portae and less the tonic contractions, because theartifacts (errors) would confuse the correct estimation in this regime. Minimalization of theelectric artifacts with the help of the electric transformer connected to the network which stopsthe tension’s fluctuation as well as the presence of the isolated organ bath in the screening roomcon not be absolute in order for keeping the experimental purity in estimating of the tonicestimation.
2. Changing the solutions in the isolated organ bath may reflect initial recording toniccontractions. It is necessary to keep a strictly liquid volume in isolated organ bath.
3. The amplitude of contractions characterizes the force of contraction and is indissolublebound by the Ca2+ ions reserve from outside, for the smooth vascular venous muscle indifferent Ca2+ ions concentration, is remarkable the change of the venous muscle reactivity of thevena portae in the groups of study in comparison with the control. Modification of thecontractions frequency tells us about pacemaker’s involvement, the source of the vena portaeautomatism – interstitial cells of Cajal [4, 5], which have been recently discovered.
4. The intensity of the structures functioning in the studies was changed in some cases onthe base of amplitude’s transformation in other only on the base of contractions frequency, butthere were noticed groups in which the transformation was depended by both index.

265
5. The other contractile index – the contraction’s surface, the total contraction’s time, thephase of contraction and of relaxation contribute to supplementation of the contractilitycharacteristic of the smooth vascular muscle of the vena portae.

266
References[1] R. Macinelli, P. Tonali, R. Romani, A. Tringali, R. Vargiu, G.B. Azzena, Mechanical
properties of smooth muscle portal vein in normal and dystrophin-deficient (mdx) mice,Experimental Physiology, vol. 84, 1999, pp. 929-940
[2] Е.Б. Манухина, Воротная вена и ее сократительная функция в норме и патологии,Успехи физиологических наук, том 19, № 3, 1988, стр. 45-66
[3] В.Г. Пинелис, М.О. Лоншамп, Влияние эндотелина-I на Na+/H+ обмен вгладкомышечных клетках сосудов, Бюллетень экспериментальной биологии имедицины, № 12, 1992, стр. 594-596
[4] T.B. Bolton, Calcium events in smooth muscles and their interstitial cells; physiological rolesof sparks, Journal of Physiology, vol. 570, no. 1, 2006, pp. 5–11.
[5] O.V. Povstyan, D.V. Gordienko, M.I. Harhun & T.B. Bolton, Identification of interstitialcells of Cajal in the rabbit portal vein, Cell Calcium, vol. 33, 2003, pp. 223–239.
EFECTELE NESPECIFICE ALE ESTROGENILORTatiana Cucu, Cristina Cucu
(Conducător ştiinţific: doctor în medicină, conferenţiar universitar, Nicolae Demişcan)Catedra Fiziologia Omului şi Biofizică, USMF “Nicolae Testemiţanu”
SummaryThe non-specific effects of estrogens
The effects of estrogens upon the cardiovascular system are: prevention ofatherosclerosis, improvement of the lipid profile and vasodilation. The effects of estrogens uponthe digestive system are: reduction of the intestinal peristalsis and increase of absorption.Estrogens also have neurotrophic and neuroprotective effects, along with the prevention of brainischemia, they have an antidiabetic action, increase the retention of water and sodium, protect theskeletal system and increase blood coagulation. Estrogens also maintain the normal structure ofskin in women by promoting keratinocytes and fibroblasts proliferation, and by stimulating itsvascularization.
RezumatLa nivelul sistemului cardiovascular, estrogenii au efect antiaterosclerotic, de ameliorare
a profilului lipidic şi de vasodilatare. La nivel digestiv, estrogenii micşorează peristaltismulintestinal, favorizînd absorbţia. Estrogenii mai posedă efect neurotrofic, neuroprotector şiantiischemic cerebral, au acţiune antidiabetică, favorizează retenţia de apă şi sodiu, au efectprotector asupra ţesutului osos, majorează coagulabilitatea sîngelui. La nivelul pielii, estrogeniimenţin structura ei normală la femei prin promovarea proliferării keratinocitelor, fibroblastelor,prin stimularea vascularizării pielii.
IntroducereEstrogenii sunt hormoni sexuali feminini, de origine steroidă, fiind reprezentaţi de
estradiol (cel mai activ), estronă şi estriol (mai puţin activi). Prin penetrarea estrogenilor încelulele-ţintă (din uter, vagin, glanda mamară, sistemul hipotalamo-hipofizar, oase, ficat etc.) şifiind transportaţi în nucleu, aceştia se cuplează cu receptorii estrogenici (ERα şi ERβ),interacţionînd ulterior cu ADN şi cu proteine celulare specifice, realizînd modularea expresiei şitranscripţiei unor gene şi inducînd sinteza proteinelor în celule.
Pe lîngă efectele specifice asupra organelor sexuale şi asupra dezvoltării sexuale la femei(stimularea proliferării şi dezvoltării uterului, vaginului şi glandelor mamare, proliferarea

267
mucoasei uterine, maturarea epiteliului vaginal, declanşarea menstruaţiei, maturarea organelorsexuale, dezvoltarea caracterelor sexuale secundare etc.), estrogenii mai posedă şi efectenespecifice, la nivelul altor organe şi sisteme de organe, acestea fiind expuse în cele ce urmează.
a) Efectele estrogenilor asupra sistemului cardiovascular:Estrogenii previn bolile cardiovasculare la femei, explicaţia posibilă fiind efectul lor
antiaterosclerotic. Efectele lor pozitive sunt atribuite ameliorării profilului lipidic şi, deasemenea, acţiunii directe a lor asupra endoteliului vascular şi musculaturii netede a vaselorsangvine. Estrogenii pot afecta şi ultimele stadii ale aterogenezei şi, nu în ultimul rind, potderegla reactivitatea vaselor aterosclerotice, ducînd astfel la vasodilataţie.
Efectele pozitive ale estrogenilor asupra profilului lipidic se manifestă prin mărireaproducerii de lipoproteine cu densitate înaltă (HDL) datorită diminuării activităţii lipazeiendoteliale hepatice şi reducerii catabolismului HDL. Dozele farmacologice de estrogeni deasemenea scad concentraţiile plasmatice de lipoproteine cu densitate joasă (LDL) – principaliitransportori de colesterol în circuitul sangvin- prin mărirea numărului şi sensibilităţii receptorilorhepatici pentru LDL. Conform unor studii, estrogenii modifică compoziţia LDL, mărindconţinutul de trigliceride şi fosfolipide, aceasta avînd ca rezultat formarea unei molecule maipuţin bogate în colesterol, ceea ce ar putea reduce absorbţia de colesterol. Cu toate acestea,estrogenii mai măresc şi sinteza lipoproteinelor cu densitate foarte joasă (VLDL), ducînd lamărirea trigliceridelor în sînge.
Acţiunea directă a estrogenilor asupra endoteliului vascular se explică prin diminuareainfluxului de calciu voltaj-dependent în aceste celule şi prin mărirea expresiei genelor implicateîn producerea vasodilatatorilor, cum ar fi prostaciclinele şi oxidul nitric (NO). Aceasta duce ladilatarea arterelor coronare, prevenind astfel ischemia ţesutului miocardic şi cardiopatiaischemică.
Unele studii au arătat că estrogenii inhibă proliferarea celulelor musculare netede dinpereţii vasculari, acest fapt putînd avea şi el un rol major în prevenirea aterosclerozei.
b) Efectele estrogenilor asupra sistemului digestiv:Celulele epiteliale intestinale conţin receptori estrogenici (ER), care sunt capabili de
reglarea transcripţiei genelor, astfel elucidîndu-se rolul estrogenilor în fiziologia tractuluigastrointestinal. Ca urmare a interacţiunii directe a estrogenilor cu ER intestinali, are locmicşorarea peristaltismului, ceea ce favorizează absorbţia intestinală, dar şi predispune laconstipaţii.
c) Efectele estrogenilor asupra sistemului nervos:Estrogenii influenţează direct funcţia cerebrală prin intermediul receptorilor estrogenici
localizaţi pe neuronii din mai multe regiuni ale creierului. Efectele lor sunt atît neuroprotective,cît şi neurotrofice. A fost demonstrat că estrogenii protejează neuronii izolaţi in vitro de cătrestresul oxidativ, leziunile ischemice, hipoglicemie, precum şi de către proteina amiloidă, careeste implicată în patogenia maladiei Alzheimer. În acelaşi timp, ei stimulează producereafactorilor de creştere a nervilor, promovînd astfel creşterea şi viabilitatea neuronală, reparaţianeuronilor lezaţi şi a ramificaţiilor dendritice.
La nivelul sinapselor neuronale, estrogenii măresc concentraţia de neurotransmiţători,cum ar fi cea de serotonină, dopamină şi norepinefrină, acţionînd asupra eliberării, recaptării şiinactivării lor enzimatice. În afară de aceasta, mai măresc şi numărul de receptori pentru aceştineurotransmiţători. Astfel, administrarea estradiolului induce o mărire semnificativă a captării deserotonină în cortexul frontal şi hipotalamus, mecanism implicat în reglarea dispoziţiei. Lafemeile de vîrstă reproductivă, a fost demonstrat că nivelele serice de estradiol corelează pozitivcu nivelele de serotonină, iar la femeile după menopauză, nivelul sangvin de serotonină estescăzut. În aceste studii, administrarea de estradiol ridica nivelul de serotonină la valoarea celeipremenopauzale. Estrogenii de asemenea sunt competitivi cu triptofanul, precursorul serotoninei,

268
pentru situsurile de cuplare de pe albuminele plasmatice, făcînd astfel triptofanul mai accesibilsistemului nervos central.
Prin inhibarea competitivă a enzimei ce inactivează norepinefrina, estrogenii provoacă unefect stimulator similar celui ale multor preparate antidepresante, iar prin reducerea activităţiiMAO măreşte nivelele de serotonină şi catecolamine în creier. Estrogenii de asemeneastimulează producerea de endorfine şi substanţe opioide de către hipotalamus.
Un alt efect semnificativ al estrogenilor asupra ţesutului cerebral este stimulareavascularizării cerebrale, posibil prin mecanisme similare celor ce au loc şi la nivelul arterelorcoronare (prin cuplare cu receptorii situaţi în endoteliu şi stimulării eliberării de oxid nitric, cecauzează vasodilatarea).
O altă acţiune asupra vascularizării cerebrale se manifestă prin efectul antiinflamator alestrogenilor la nivelul pereţilor vasculari, protejîndu-i de lezarea prin citokine şi radicali liberi, şiîmpiedicînd formarea plăcilor aterosclerotice, avînd ca rezultat prevenirea formării zonelorischemice ale ţesutului cerebral.
d) Efectele estrogenilor asupra sistemului endocrin:Cantităţi mici de estrogeni ameliorează metabolismul glucidic şi chiar reduc
hiperglicemia în diabetul zaharat insulinorezistent. Mecanismul ar fi modificarea sensibilităţiireceptorului insulinei sau, posibil, inhibiţia degradării insulinei.
Ingestia de estrogeni sintetici stimulează producerea hepatică de globuline, inclusiv aglobulinei ce fixează tiroxina. Astfel, sunt crescute nivelele totale de tiroxină cu o creştere afracţiei legate. Cu toate acestea, nivelul tiroxinei nelegate nu este influenţat de către ingestia deestrogeni.
Similar globulinei ce fixează tiroxina, estrogenii măresc producerea hepatică detranscortină, mărind nivelul corticosteroizilor transportaţi în sînge.
Estrogenii mai induc şi creşterea hepatică de angiotensinogen, ceea ce duce la mărireasecundară a angiotensinei şi, ca rezultat, a aldosteronului. Aceasta contribuie la retenţiacompensatorie a apei şi natriului de către rinichi, şi favorizează expansiunea lichiduluiintravascular în cel extracelular cu apariţia edemelor. Cu toate acestea, se consideră că acesteasunt efecte dozo-dependente.
În urma unei serii de cercetări pe animale a fost demonstrat că receptorii estrogenici ERαsituaţi în hipotalamus, şi anume în nucleul ventromedial, controlează ingestia de alimente,cheltuielile de energie şi distribuţia corporală a ţesutului adipos. Atunci cînd aceşti receptori eraudistruşi, animalele începeau imediat să mănînce mai multă mîncare, să cheltuie mai puţinăenergie şi să acumuleze ţesut adipos.
e) Efectele estrogenilor asupra sistemului osos:Estrogenii inhibă resorbţia osoasă, manifestînd în acest mod antagonism cu
parathormonul, datorită inducerii directe a apoptozei osteoclastelor printr-un mecanism mediatde receptorii estrogenici.
Conform unor studii recente, estradiolul stopează activarea enzimei caspaza-3 – enzimăce joacă un rol central în apoptoza osteoblastelor, favorizînd astfel formarea ţesutului osos.
f) Efectele estrogenilor asupra pielii:Este binecunoscut faptul că estrogenii sunt importanţi în menţinerea structurii normale a
pielii la femei.Activitatea mitotică înaltă în epidermul uman şi al rozătoarelor poate fi cauzată de
estrogeni. Unele studii au demonstrat faptul că keratinocitele expresează pe membrana sa unreceptor estrogenic (ER) şi că estrogenii sunt implicaţi în progresarea carcinoamelor de celulebazale şi scuamoase, la fel ca şi în cancerul mamar şi de prostată.
Există două tipuri distincte de ER intracelulari (ERα şi ERβ), care se cuplează cu 17β-estradiolul cu o afinitate similară. Mai mult decît atît, recent a fost demonstrat faptul că în uneleţesuturi estrogenii pot semnala şi printr-un receptor membranar. Efectele estrogenului asupra

269
keratinocitelor umane se realizează predominant prin ERβ, iar pînă nu demult se considera că înpiele există un singur receptor estrogenic ERα.
Rolul protector al estrogenilor asupra epidermului uman poate fi explicat prin aceea căestradiolul previne apoptoza indusă de către peroxidul de hidrogen în keratinocite prinpromovarea expresiei bcl-2. Mai mult decît atît, estradiolul inhibă producerea citokinelorimplicate în atragerea macrofagelor spre focarul de inflamaţie.
Pe lîngă influenţa lor asupra proliferării keratinocitelor, estrogenii influenţează şigrosimea pielii prin stimularea sintezei, maturaţiei şi metabolismului colagenului, fibroblasteledermale fiind identificate în unele studii drept celule-ţintă ale estrogenilor, ceea ce prezintăimportanţă în cicatrizarea plăgilor. Se pare că estrogenul reglează toate fazele cicatrizăriiepiteliale datorită efectelor sale asupra celulelor implicate în inflamaţie, asupra epitelizării,angiogenezei, depozitării de matrice extracelulară şi remodelării tisulare.
Pe lîngă aceasta, estrogenii stimulează vascularizarea pielii datorită acţiuniivasodilatatoare, au efect inhibitor asupra creşterii foliculului de păr, reduc dimensiunile şi inhibăsecreţia glandelor sebacee la ambele sexe, stimulează pigmentarea pielii, melanocitele fiindcelule sensibile la acţiunea estrogenilor, dar mecanismele prin care se realizează aceste acţiunisunt incomplet studiate.
g) Efectele estrogenilor asupra sîngelui:Una dintre primele complicaţii ale contraceptivelor orale pe bază de estrogeni a fost
creşterea incidenţei maladiilor tromboembolice. Explicaţia rezidă în faptul că estrogeniicontribuie la majorarea concentraţiei factorilor de coagulare II, VII, IX şi X, fibrinogenuluidatorită stimulării producerii lor hepatice şi, de asemenea, reduc nivelul antitrombinei III. Seconsideră că aceste modificări ale coagulabilităţii sangvine induse de estrogeni sunt dozo-dependente, deoarece dozele de mai puţin de 50 μg au un impact mic sau nu au deloc influenţăasupra factorilor de coagulare enumeraţi mai sus.
Concluzii1. Estrogenii, prin efectele lor nespecifice, influenţează asupra tuturor sistemelor funcţionale
ale organismului: sistemul cardiovascular, sistemul digestiv, sistemul nervos, sistemulendocrin, sistemul osos, piele, sînge.
2. De regulă, această acţiune este benefică pentru organism, efectele pozitive fiind: efectulantiaterosclerotic, de ameliorare a profilului lipidic, antiischemic, de favorizare aabsorbţiei intestinale, funcţia neurotrofică şi neuroprotectoare, reglarea stăriipsihoemoţionale, efectul antidepresant, antidiabetic, de control al ingestiei de alimente şia distribuţiei corporale a ţesutului adipos, de inhibiţie a resorbţiei osoase şi de stimulare aformării ţesutului osos, de menţinere a structurii normale a pielii la femei, de cicatrizareepitelială.
3. Însă este necesar de a lua în calcul şi efectele adverse ale estrogenilor, descrise înliteratură, acestea fiind: hipertrigliceridemia, predispunerea la constipaţii, mărireanivelelor de tiroxină şi corticosteroizi în sînge (fracţiile legate), de retenţie a apei şinatriului în organism, activitatea proliferativă înaltă a epidermului (ce poate duce, înunele cazuri, la progresarea cancerului de piele, mamar şi de prostată), şi, nu în ultimulrind, efectul de hipercoagulabilitate sangvină.
Bibliografie1. Ghicavîi V., Bacinschii N., Bumacova L., Guşuilă Gh., Podgurschi L. – Farmacologie clinică
– Chişinău,20102. Ramos, D, Stanczyk, F, et al - Metabolic and Endocrinologic Effects of Steroidal
Contraception - Glob. libr. women's med, 20093. Shepherd J., - Effects of Estrogen: Estrogen and the Brain– www.medscape.com

270
4. Stevenson S. ; Nelson L. D. ; Sharpe D. T. ; Thornton M. J. - 17 β-Estradiol regulates thesecretion of TGF-β by cultured human dermal fibroblasts - Journal of biomaterials science -2008, vol. 19, no8, pp. 1097-1109
5. Takashi Kameda, Hiroshi Mano et al. - Estrogen Inhibits Bone Resorption by DirectlyInducing Apoptosis of the Bone-resorbing Osteoclasts - (2010) -www.academic.research.microsoft.com
6. Thomas M., Xu X., Norfleet A.M., Watson C. - The presence of functional estrogen receptorsin intestinal epithelial cells - Endocrinology January 1, 1993 vol. 132 no. 1 426-430
7. Thornton M.J. - Oestrogen functions in skin and skin appendages - June 2005, Vol. 9, No. 3 ,Pages 617-629
8. Vischer U. - Estrogens and atherosclerosis: a direct protective effect on the IGH vascularwall?- European Journal of Endocrinology (1997) 137 343–344
9. www.glowm.com10. www.sciencedaily.com
PROTEINELE ŞOCULUI TERMICEduard Bobuţac, Irina Mîţu
Catedra Fiziopatologie şi Fiziopatologie clinică(Conducător ştiinţific Vasile Lutan, doctor habilitat, profesor universitar)
SummaryHeat shock proteins
The paper presents information on the history, discovery, chemical structure,classification and biological role of heat shock proteins (stress proteins) in non-specific cellresistance and the ability of these substances in pathological implications.
RezumatÎn lucrare sunt prezentate informaţii referitor la istoria descoperirii, structura chimică,
clasificarea şi rolul biologic al proteinelor şocului termic (proteine de stres) în rezistenţanespecifică celulară şi posibilitatea implicaţiilor acestor substanţe în patologie.
ActualitateaImportanţa proteinelor şocului termic pentru ştiinţa biologică şi medicină este
determinată de rolul acestor substanţe în menţinerea rezistenţei nespecificve a celulelor laacţiunea factorilor nocivi şi eventuala implicarea în practica medicală în tratamentul diferitorpatologii (insuficienţa circulatorie, cancer şi a.).
Proteinele şocului termic (Heat shock proteins, HSP) reprezintă un grup deproteine cu structură similară, sintetizate de celule în cazul expunerii acţiunii diferitorfactori nocifi din care cauză mai sunt denumite şi proteine de stres celular.
S-a demonstrat, că sinteza HSP reprezintă o reacţie universală adaptativă a celuleiîndreptată spre creşterea rezistenţei nespecifice în condiţii de stres.
IstoriaProteinele şocului termic au fost descoperite de către genetecianul Ferrucio Ritossa
în 1962 în cercetările cromozomilor politeni din glandele salivare ale larvelor deDrozofila. În una din experienţe, accidental a crescut temperatura în termostat ceea ce acondus la moartea insectelor. Din curiozitate acestea nu au fost aruncate, ci au fostcercetate. S-a observat, că în unele regiuni ale cromozomilor au apărut umflături (pufuri),

271
dovadă a activităţii înalte a genelor din locusurile respective. Concomitent în celule au fostdepistate şi nişte clase de proteine consecutiv activării acestor gene. Ulterior s-a presupus, căcauza activării genelor a fost temperatura înaltă, genele activate au fost numite “geneleraspunsului la şocul termic”, iar proteinele induse de acestea au fost numite “proteineleşocului termic”.
Investigaţiile ulterioare au demonstrat, că sinteza acestor proteine este o proprietateuniversală a tuturor celulelor organismelor vii – de la bacterii până la om. De asemenea s-aconstatat că sinteza „proteinelor şocului termic” este declanşată şi de alte tipuri de factorinocivi (stresogeni), cum ar fi:
- stresogeni biologici (infectarea cu viruşi, bacterii)- stresogeni fizici (radiaţii ionizante, tempertaura scăzută, hipoxie, inaniţie, radicali
liberi)- stresogeni chimici (alcooli, metale, deviaţâii de pH) şi a.
Fig. 1. „Pufurile” cromozomiale formate la acţiunea temperaturii înalte
Din aceste considerente proteinele şocului termic au mai fost numite şi proteinelede stres. Investigaţiile ulterioare au fost axate în direcţia stabilirii mecanismului induceriisintezei proteinelor de stres, mecanismul protecţiei celuleli în condiţii de stres celular şiposibilităţile implicării acestora în creşterea potenţialului antistres al celuleli şi utilizarea HŞP înterapia diferitor patologii.
Clasificarea proteinelor şocului termic.Proteinele şocului termic se găsesc în citosol, mitocondrii, reticul endoplasmatic şi
nucleu. La mamifere cele mai studiate proteine sunt HSP 60, 70, 90 şi 110 kDa, carejoacă un rol important în procesele fundamentale intracelulare - de la acţiuneaantiapoptotică pînă la circulaţia intracelulară de proteine («Тепловой шок» МагнушевскаяЕ.К., Умудова Э.И.)
În funcţie de masa moleculară HSP pot fi clasificate în:• HSP mici• HSP 60 kDa (GroEL)• HSP 70 kDa (DnaK)• HSP 90 kDa (HptG)• HSP 100 kDa (ATP Clp)
HSP cu 60 şi 70 kDa sunt numite chaperonine. HSP cu masa moleculară micăsunt ATP-independente, iar HSP cu masa moleculară mare sunt ATP-dependente.
Funcţiile proteinelor şocului termic.

272
Fiecare din aceste clase de proteine are o funcţie specifică.1. Clasa HSP70 (chaperonine) - în condiţii fiziologice interacţionează cu lanţurile de
proteine nou formate pe ribozomi prevenind plierea greşită a lanţului polipeptidic imaturşi transportă aceste lanţuri spre organitele celulei. Pe lângă această HSP70 sunt implicateîn transportul direcţional al mai multor tipuri de kinaze în regiunile, unde acestea îşi vordesfăşura activitatea. În aşa mod HSP previn denaturarea proteinelor celulare pe traseulintracellular de la ribozomi şi până la organitele de destinaţie, unde acestea pot fi supuse diferitoracţiuni fizico-chimice. Rolul HSP70 devine extrem de important în condiţii de stress cellular cumodificări grave ale mediului intracellular – hipoxie, hipo-hiperosmolaritate, acidiate, alcaloză,temperature scăzută sau ridicată, ceea ce ar conduce la denaturarea protreinelor nou formate.
Funcţiile HSP ca chaperonine pot fi rezumate după cum urmează:1. Coagularea lanţurilor polipeptidice imature;2. Facilitarea trecerii proteinelor prin compartimentele celulare;3. Modularea activităţii proteinei prin stabilizarea şi - sau maturizarea conformaţiei
funcţional competente;4. Suportul formării / distrugerii complexelor multiproteice;5. Corectarea proteinelor incorect conformate;6. Protejarea proteinelor de agregare;7. Direcţionarea proteinelor complet deformate spre distrugere;8. Organizarea agregatelor din fragmentele proteinelor distruse;9. Solubilizarea agregatelor de proteine pentru degradarea ulterioară.
2. Clasa HSP90 – în complex cu proteine auxiliare interacţionează cu receptoriicitoplasmatici ai hormonilor steroizi, asigurînd legarea efectivă a hormonului cu receptorulşi transportul complexului hormon - receptor spre nucleu. Aceasta asigură eficacitateabiologică a hormonilor steroizi vital necesari celulei în condiţii de stress.
4. Clasa HSP60 – participă în foldingul proteinelor complicate ce conţin mai multedomene, ceea ce asigură redarea structurii tridimensionale a moleculei proteice, conformismulmolecular şi activitatea biologică, dependentă de stereometria celuleli (de ex., a moleculelor deenzime). Înb caz de folding eronat, HSP efectuează re-foldingul moleculei.
De asemenea HSP participă la corecţia “greşelilor” în structura proteinelor parţialdenaturate, iar în caz de erori irecuperabile demarează liza proteinelor anomale, inţiind resintezaproteinelor normale. În cazul în care celula este inaptă de a sintetiza proteine normale, HSPiniţiază apoptoza celulei.
De exemplu, proteinele HSP70, HSP90 sunt prezente în concentraţii mari în celulenesupuse stresului, formând 1-1,5% din proteinele celulare totale, ceea ce indică onecesitate permanentă a celulei pentru a menţine homeostazia conformatională a acestorproteine.
Or, stresul celular induce sinteza proteinelor şocului termic. Expresia mărită aacestora protejează celula prin stabilizarea proteinelor normale precum şi prin refoldingulcelor afectate. Acumulîndu-se în diferite acţiuni dăunătoare, proteinele şocului termic ajutăcelula să menţină homeostazia în condiţii de stres. HSP asigură protecţia împotrivaalterării cromatinei.
Mecanismul general de sinteza al proteinelor şocului termic la eucariote constă îninducerea genelor proteinelor de soc termic (HSP) prin intermediul factorului termic deşoc (HSF). Familia HSF constă din 4 tipuri de HSF, iar la om se expresează doar HSF1,HSF2 şi HSF4. HSF1 - este un activator universal stres – receptiv, iar HSF2 este responsabilde procesele de diferenţiere celulară. În lipsa stresului aceşti factori se află în citoplasmăşi nucleu în formă de monomeri care nu se pot lega cu ADN-ul. Ca răspuns la stres,HSF se asamblează în trimeri (sunt posibili homotrimeri: HSF1-HSF1 sau heterotrimeri:
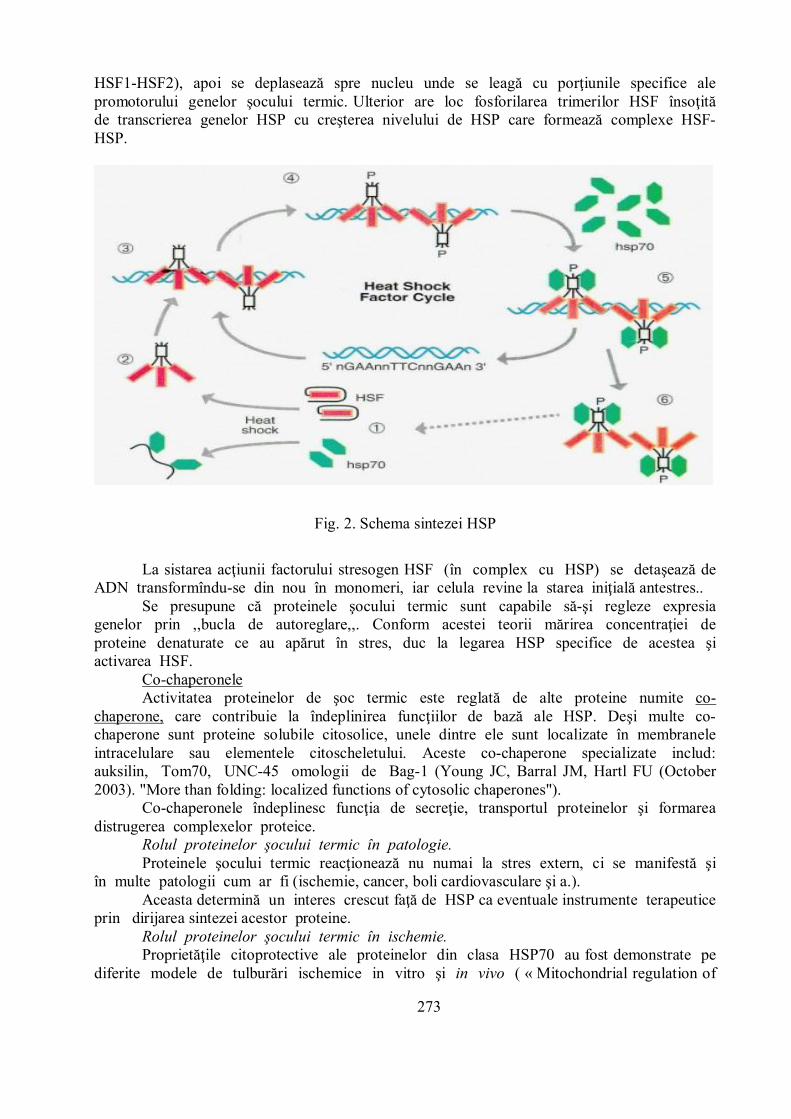
273
HSF1-HSF2), apoi se deplasează spre nucleu unde se leagă cu porţiunile specifice alepromotorului genelor şocului termic. Ulterior are loc fosforilarea trimerilor HSF însoţităde transcrierea genelor HSP cu creşterea nivelului de HSP care formează complexe HSF-HSP.
Fig. 2. Schema sintezei HSP
La sistarea acţiunii factorului stresogen HSF (în complex cu HSP) se detaşează deADN transformîndu-se din nou în monomeri, iar celula revine la starea iniţială antestres..
Se presupune că proteinele şocului termic sunt capabile să-şi regleze expresiagenelor prin ,,bucla de autoreglare,,. Conform acestei teorii mărirea concentraţiei deproteine denaturate ce au apărut în stres, duc la legarea HSP specifice de acestea şiactivarea HSF.
Co-chaperoneleActivitatea proteinelor de şoc termic este reglată de alte proteine numite co-
chaperone, care contribuie la îndeplinirea funcţiilor de bază ale HSP. Deşi multe co-chaperone sunt proteine solubile citosolice, unele dintre ele sunt localizate în membraneleintracelulare sau elementele citoscheletului. Aceste co-chaperone specializate includ:auksilin, Tom70, UNC-45 omologii de Bag-1 (Young JC, Barral JM, Hartl FU (October2003). "More than folding: localized functions of cytosolic chaperones").
Co-chaperonele îndeplinesc funcţia de secreţie, transportul proteinelor şi formareadistrugerea complexelor proteice.
Rolul proteinelor şocului termic în patologie.Proteinele şocului termic reacţionează nu numai la stres extern, ci se manifestă şi
în multe patologii cum ar fi (ischemie, cancer, boli cardiovasculare şi a.).Aceasta determină un interes crescut faţă de HSP ca eventuale instrumente terapeutice
prin dirijarea sintezei acestor proteine.Rolul proteinelor şocului termic în ischemie.Proprietăţile citoprotective ale proteinelor din clasa HSP70 au fost demonstrate pe
diferite modele de tulburări ischemice in vitro şi in vivo ( « Mitochondrial regulation of

274
apoptotic cell death. » Gogvadze V, Orrenius S. 1996, 1997, 1998, 2000, 2004, 2005). Iniţial,această protecţie se explica prin acţiunea HSP ca chaperonine (prin menţinereaconformaţiei normale a proteinelor şi prin prevenirea agregării lor), dar apoi s-a constatatcă HSP70 pot reactiona direct cu căile de apoptoză si necroză.
Ischemia cerebrală induce apoptoza prin diverse căi, iar HSP70 inhibă aceste căi.“Calea internă" a apoptozei constă în eliberarea substanţelor apoptogene mitocondriale,
deschiderea “porilor” mitocondriali şi activarea caspazelor.“Calea externă” a apoptozei constă în activarea receptorilor plasmatici membranari
(Fas şi TNFR) care induc apoptoza prin intermediul caspazei-8.Proteinele HSP70 pot inhiba eliberarea citocromului c (cyt c) din mitocondrii şi
translocarea factorului de inducere a apoptozei (FIA) în nucleu, reducând leziunilecerebrale ischemice şi, de asemenea, inhibînd eliberarea proteinelor proapoptotice (SMACDIABLO) din mitocondriile miocitelor.
În plus, proteinele HSP interacţionează cu topoizomeraza 1 (reglator al apoptozei)şi sunt efectori importanţi ai kinazei antiapoptotice Akt/PKB.
Activarea semnificativă de către proteinele de şoc termic a glutation peroxidazei siglutation reductazei este un element esenţial în mecanismul de acţiune citoprotectiv alHSP în ischemie.
Rolul proteinelor şocului termic în cancerProteinele de şoc termic sunt exprimate în mai multe tipuri de cancer uman şi
participă în proliferare, diferenţiere, metastazare, şi recunoaşterea celulelor tumorale decătre sistemul imun. Ele sunt biomarkeri utili ai carcinogenezei în anumite ţesuturi şisemnalează gradul de diferenţiere şi agresivitate a unor forme de cancer. În plus, nivelulde HSP circulante şi anticorpii anti-HSP pot fi utili în diagnosticul cancerului. Expresiamărită a HSP poate fi criteriu al răspunsului la tratamentul antitumoral. De exemplu, HSP27şi HSP70 formează rezistenţa la chimioterapie în cancerul mamar, iar creşterea niveluluide HSP27 prezice un răspuns slab la chimioterapia leucemiei. În acelaşi timp, expresiade HSP70 sugerează efectele bune ale chimioterapiei în osteosarcom. (“Heat shockproteins in cancer: diagnostic, prognostic, predictive, and treatment implications” Daniel R.Ciocca and Stuart K. Calderwood).
În dezvoltarea terapiei anticanceroase cu HSP a jucat rolul funcţia lor dublă înorganism: pe de o parte – funcţia citoprotectoare intracelulară anti-apoptotică, iar pe dealtă parte – efectul imunogen. Acest fapt a permis dezvoltarea a două strategii de bază înterapia cancerului:1) modificarea farmacologică a expresiei HSP şi activitatea lor în calitate de chaperonemoleculare;2) aplicarea HSP în vaccinuri anticancer bazate pe capacitatea acestora de a acţiona caun adjuvant imunologic. Astfel, în cadrul mai multor experienţe s-a dovedit ca animalelepot raspunde la “vaccinarea tumorală” cu sinteza de HSP în tesutul tumoral. Celuleletumorale atenuate si inoculate in cantitati mici la rozatoare induce acestora imunitate lainocularea repetată a celulelor tumorale. Se presupune, că HSP pot asocia fragmente proteicede la celulele maligne moarte si prezenta sistemului imun.
Rolul HSP în procesul îmbătrîniriiPe masură imbatrânirii organismul pierde capacitatea de a răspunde în mod adecvat
la diferite stresuri din exterior şi de a menţine homeostazia. Celulele bătrâne sunt maipredispuse la acţiuni nocive şi boli, şi, prin urmare, odată cu vârsta creşte şi sensibilitateala aceşti factori.
Pe parcursul vieţii unei proteine stabile în ea se produc numeroase modificări post-translaţionale. Stabilitatea proteinelor se perturbă din cauza numeroaselor acţiuni nocive –

275
oxidarea lanţurilor laterale, glicozilare, dezaminarea resturilor asparaginice şi glutaminice,ceea ce duce la formarea de legături izopeptidice.
Sensibilitatea la leziunile proteotoxice crește din cauza erorilor în transcripţie şitranslaţie și se manifestă prin defecte ale conformaţiei proteinelor. Pentru îmbătrînire estecaracteristică creşterea modificării proteinelor.
Funcţiile de chaperone se dereglează, crește necesitatea de degradare a proteinelor,dar activitatea aparatului proteolitic principal, scade cu vârsta, ceea ce duce la pericolulde glicare
Agregarea, de asemenea, este însoţită de inhibarea proteazomală şi oprirea cicluluicelular. Cu vârsta, dereglează şi degradarea lizozomală a proteinelor (probabil din cauzasuprimarii lipofuscinei). Acumularea de proteine incorect conformate şi slăbireamecanismelor de protecţie duce la formarea de agregate proteice care dereglează funcţiilecelulare şi sunt cauza a bolilor apărute cu vârsta.
Rolul HSP in bolile cardiovasculare- Proteinele de soc termic par sa aiba un rol semnificativ in maladiile
cardiovasculare, indeosebi hsp90, hsp70, hsp84, hsp27, hsp20 (“Influence of solublemediators upon the expression of different physiological and virulence hallmarks of bacteria.“ Roum.Biotech. Lett., vol. 13, 2, 3631-3642).
- Hsp90 leaga atât nitric oxid sintaza endoteliala, cât si guanilat ciclaza solubila.O kinaza a oxidului nitric din calea de semnalizare celulara, protein kinaza G,
fosforileaza o proteina de soc termic mica, hsp20. Fosforilarea hsp20 se coreleaza binecu relaxarea musculaturii netede si este o fosfoproteina implicata in proces, fiindsemnificativă in dezvoltarea fenotipului musculaturii netede .
Hsp de asemenea joaca un rol important in prevenirea agregarii trombocitare, infunctionarea miocitelor cardiace si prevenirea apoptozei dupa leziunile ischemice, infunctionarea musculaturii scheletice si raspunsul muscular la insulina.
Hsp27 este o fosfoproteina cu rol major pentru contractia tuturor grupelormusculare. Hsp27 functioneaza in migratia musculaturii netede si pare sa joace un rolintegral in dinamica filamentelor de actina si adeziunea focala. S-a emis ipoteza conformcareia hsp27 si hsp20 ar juca roluri in formarea puntilor incrucisate dintre actina simiozina (“Influence of soluble mediators upon the expression of different physiological andvirulence hallmarks of bacteria.“ Roum. Biotech. Lett., vol. 13, 2, 3631-3642).
Bibliografie1. Anckar J, Hietakangas V, Denessiouk K, Thiele DJ, Johnson MS, Sistonen L. “Inhibition
of DNA binding by differential sumoylation of heat shock factors.”Mol Cell Biol. 2006 Feb;26(3):955-64.
2. Anton Sandqvist, Johanna K. Bjork, Malin Åkerfelt, Zhanna Chitikova, Alexei Grichine,Claire Vourc, Caroline Jolly, Tiina A. Salminen, Yvonne Nymalm, Lea Sistonen“Heterotrimerization of heat-shock factors 1 and 2 provides a transcriptional switch inresponse to distinct stimuli.” Mol Biol Cell. 2009 Mar;20(5):1340-7. Epub 2009 Jan 7.
3. Daniel R. Ciocca and Stuart K. Calderwood “Heat shock proteins in cancer:diagnostic, prognostic, predictive, and treatment implications”
4. Didelot C, Lanneau D, Brunet M, Joly AL, De Thonel A, Chiosis G, Garrido C. “Anti-cancer therapeutic approaches based on intracellular and extracellular heat shockproteins.” Curr Med Chem. 2007;14(27):2839-47.
5. Didelot C, Schmitt E, Brunet M, Maingret L, Parcellier A, Garrido C. „Heat shockproteins: endogenous modulators of apoptotic cell death.” Handb ExpPharmacol. 2006;(172):171-98.

276
6. Gogvadze V, Orrenius S. “ Mitochondrial regulation of apoptotic cell death.“ 1996,1997, 1998, 2000, 2004, 2005
7. Hietakangas V, Ahlskog JK, Jakobsson AM, Hellesuo M, Sahlberg NM, Holmberg CI,Mikhailov A, Palvimo JJ, Pirkkala L, Sistonen L. “Phosphorylation of serine 303 is aprerequisite for the stress-inducible SUMO modification of heat shock factor 1.” MolCell Biol. 2003 Apr;23(8):2953-68.
8. Lanneau D, de Thonel A, Maurel S, Didelot C, Garrido C. “Apoptosis versus celldifferentiation: role of heat shock proteins HSP90, HSP70 and HSP27.” Prion. 2007Jan;1(1):53-60. Epub 2007 Jan 24.
9. Marko Kallio, Yunhua Chang, Martine Manuel, Tero-Pekka Alastalo1, Murielle Rallu,Yorick Gitton, Lila Pirkkala1, Marie-Thérèse Loones, Liliana Paslaru, Severine Larney,Sophie Hiard, Michel Morange, Lea Sistonen, and Valérie Mezger “Brain abnormalities,defective meiotic chromosome synapsis and female subfertility in HSF2 null mice.”EMBO J. 2002 Jun 3;21(11):2591-601.
10. Mayer MP, Bukau B. “Hsp70 chaperones: cellular functions and molecular mechanism.”Cell Mol Life Sci. 2005 Mar;62(6):670-84.
11. Магнушевская Е.К., Умудова Э.И “Тепловой шок”12. Schmitt E, Gehrmann M, Brunet M, Multhoff G, Garrido C. “Intracellular and
extracellular functions of heat shock proteins: repercussions in cancer therapy.” J LeukocBiol. 2007 Jan;81(1):15-27. Epub 2006 Aug 24.
13. Summers DW, Douglas PM, Ramos CH, Cyr DM. “Polypeptide transfer from Hsp40 toHsp70 molecular chaperones.” Trends Biochem Sci. 2009 May;34(5):230-3. Epub 2009Apr 7.
14. Van der Weerd L, Lythgoe MF, Badin RA, Valentim LM, Akbar MT, de Belleroche JS,Latchman DS, Gadian DG. “Neuroprotective effects of HSP70 overexpression aftercerebral ischaemia--an MRI study.” Exp Neurol. 2005 Sep;195(1):257-66.
15. Young JC, Barral JM, Hartl FU "More than folding: localized functions of cytosolicchaperones". (October 2003).
SCHIMBĂRILE FIZIOLOGICE ŞI PSIHOSOMATICEDUPĂ TRANSPLANTUL DE INIMĂ
Andrei Vilijînschii(Cond. şt. – Melniciuc Naina, asist.univ.)Catedra Fiziologia omului şi Biofizică)
SummaryPhysiological and psychosomatic changesafter heart transplantation
Heart transplantation is a current issue because each person has individual polymorphism.Antigenic structures of the donor heart are detected by the immune system recipient and,respectively, should be subject to particularly destruction and eliminated from the body. Theseprocesses are physiologically normal for the recipient, but they also kill him. To avoid thedestruction of the transplanted heart, this article will provide alternative methods of hearttransplantation.
RezumatTransplantul de inimă este o problemă actuală din cauza polimorfismului individual al
fiecărui om. Structuri antigenice ale inimii donatorului se detectează de către structuri specificeale sistemului imun ca fiind străine şi prin urmare trebuie să fie supuse distrugerii şi eliminării

277
din organismul recipientului. Aceste procese fiind fiziologic narmale pentru organismulrecipientului, îl omuară din momentul cînd a primit o inimă nouă. Pentru a evita aceste precesedistrugătoare în acest articol vor fi propuse metode alternative de transplant de inimă.
Cronologia transplantuluiPentru prima dată transplantul de inimă a fost efectut în 1905. Cîinele a trăit 2 ore. Cele
mai mari succese în acest domeniu au fost obţinute de către chirurgul rus V.P.Demihov. El aelabora peste 20 de variante de transplanturi de inima. Primul xenotransplant (transplant a unuiorgan de la animal la om) de inimă de la cimpanzeu la om a fost efectuat de către James Hardieîn 1967. Însă inima animalului după dimensiunea sa era mai mică decît cea a omul. Recipientul atrăit o oră şi 30 min. La 03 decembrie 1967 în Cape Town (Africa de Sud) tînărul chirurgChristian Barnard a efectua pentru prima dată alotransplantul (transplant a unui organ de la om laom) de inimă cu mare succes, folosind metodele lui Demihov. Primul recipient – un bărbat învîrstă de 55 de ani. Însă recipientul a trăit 18 zile. Cauza decesului – pneumonie bilaterală.Pentru prevenirea respingerii de transpant recipientului i s-au administrat preparate farmaceuticeimunosupresive – imuran şi prednizolon. De asemenea, a fost supus procedurilor de radiaţie.Asuprirea întregului sistem imun a favorizat dezvoltarea bolilor secundare. Necătînd la acesterezulate în următorii 2 ani numărul de transplanturi creştea. Oficial s-au efectuat 148 dealotransplanturi în acea perioadă. Nici un recipient nu a supravieţuit mai mult de 6 luni.Problema cotidiană a devenit – respingerea de transplant, sau mai corect – Complexul Principalde Histocompatibilitate. Primul alotransplant de inimă în Uniunea Sovietică a fost efectuat decătre celebrul cardiochirurg V.Shumakov în martie 1987. Primul xenotransplant de inimă (inimăde babuin) la sugar a fost efectut în SUA de către Leonard Bailey în centrul medical aluniversităţii Loma-Linda, statul California. Nou născută Stephanie Fay avea sindromulhipoplaziei părţilor stîngi ale inimii. Operaţia a avut succes, însă micul recipient a supravieţuit 21de zile. Cauza decesului – infecţia renală.
Problemele moral – eticeSe împart în 4 blocuri. 1. Relaţiile comerciale pe parcursul planificării operaţiei. Tehnica
şi tehnologiile operării ce ocupă partea financiară. 2. Constatarea morţii donatorului de inimădupă criteriile de moarte a creierului: (a) încetinirea totală a activităţii sistemului nervos centralcu starea inconştientă ireversibilă, cu condiţia susţinerii respiraţiei şi circulaţiei sangvine cuajutorul aparatajului corespunzător; (b) pe EEG se înregistrează o linie izoelectrică, nu semanifestă răspunsuri de reflex la stimuli externi; (c) diagnoză definitivă de moarte a creieruluise pune după 26 de zile; (d) pentru întreţinerea de apnee e necesar de controlat P(CO2)=40 mmHg pentru stimularea respiraţei. 3. Problema de explantare a organului din cadavru sau om viu.Nu este uman de a folosi în calitate de donatori de organe oamenii social vulnerabili, copii orfanişi vagabonzi, bolnavii clinicii psihiatrice şi oamenii ţărilor din Lumea a Treia. 4. Problemarepartizării transplanturilor. Criteriul principal de repartizare este gradul de compatibilitateantigenică a perechii donator-recipient. E de dorit ca difirenţa dintre antigenii complexului HLAsă nu fie mai mult de cît 3. O mare importanţă are severitatea bolii recipientului şi distanţa detransportare a organului, deoarece inima este organ cu durata cea mai mică de transportare invitro (aproximativ 5 ore).
Ultimele observaţii clinice în transplantologie au determinat formarea încă a unui bloc deprobleme moral-etice – schimbarea statutului psihofiziologic a recipientului. La mai mult de10% din pacienţi cu inimă nouă, după operaţie au observat apariţia unor noi trăsături de caracter,obiceiuri, talentelor, schimbarea comportamentului şi preferinţelor în alimentaţie, care anteriornu au fost practicate în viaţa cotidiană. Caz clinic: Pacientului (vîrsta 8 ani, sex feminin) i s-afăcut transplant de inimă. Donatorul (vîrsta 10 ani, sex feminin) – victima de asasinare brutală.După operaţie micul recipient a început să aibă des coşmaruri în carea ea era victima de

278
asasinare. Părinţii s-au adresat la psihiatru. Ceea ce a auzit el din povestirile pacientului, l-aconvins pe psihiatru că e vorba despre circumstanţele asasinării donatorului de inimă. Aceastăinformaţie a fost transmisă poliţiei şi era atît de exactă, încît asasinul a fost aresta. Asemeneacazuri sunt numeroase. Concluzia este:
Cauzele posibile care determină aceste schimbări: 1.”Memoria celulară” [Paul Pearsall].Această teorie este pînă ce în studiere. 2. Inima – ca un generator puternic a biocîmpuluimagnetic, mai puternic ca cel al creierului, care poate fi registrat la distanţa de 30 – 40 metri, peR-unda a EKG constituie 100 – 300 pT. Aceste biocîmpuri pot provoca o schimbare înfuncţionarea unor reflexe psihosomatice.
Imunitatea şi transplantul de inimă. HistocompatibilitateaCele mai profunde schimbări fiziologice după transplatare sunt în sistemul imun. Odată
cu apariţia în organismul recipientulu unui organ străin se induc reacţii de respingere atransplantului. Sunt de două tipuri care însoţesc unul pe altul: (1) Respingerea umorală şi (2)Respingerea celulară. Procesul de respingere se activează dătorită prezenţei a două complexe deantigeni: (1)HLA de clasa I (situaţi pe membranele cardiomiocitelor şi celulele endoteliale); (2)MICA (situaţi pe celulele endoteliale a vaselor coronariene şi participă la inducerea boliivasculare a transplantului). Antigenii HLA sunt prezenţi în organismul uman în număr de 150de tipuri de proteine şi peptide. Orice 6 antigeni din cei 150 sunt prezenţi pe fiecare celulă aorganismului. Astfel combinaţia antigenilor HLA poate depăşi un trillion. Respectiv acestordouă clase de antigeni se sintetizează anticorpii: anti-HLA şi anti-MICA (anti-HLA-MICA).Combinaţia de concentraţie a acestor două grupuri de anticorpi participă la patogeneza boliivasculare a transplantului. Boala se caracterizează prin mărirea grosimii pereţilor vaselorcoronariene şi ingustarea lumenului vaselor sangvine. Prin urmare – scăderea intensivităţiialimentării miocardului. Procesul poate bloca total circulaţia sangvină în vasele coronariene.Procesul pathologic se previne prin administrarea preparatului farmaceutic Sirolimus. Însăprocesul de respingere este dirijat în primul rînd de T-limfocite (cu influenţa citotoxică aT-killerilor). Semne de respingere pot fi urmărite pe EKG: tahicardie, extrasistole, scădereavoltajului undelor. În primele 4 – 5 zile organul se grefează. În acest timp celuleleimunocompetente identifică antigenul străin. Din a 5-a zi se produce perturbareamicrocirculaţiei, se dezvoltă edem şi se începe invazia organului transplantat cu celulelemononucleare şi se declanşează procesul de respingere. Astfel fiziologia omului dupătransplantare – este fiziologia respingerii organului transplantat.

279
Terapia imunosupresivăPentru a bloca reacţiile de respingere celulară şi umorală există imunosupresoare
specifice de ultima generaţie care în mare parte supresează T-limfocite care participă la reacţiilede respingere, decît pe cei care răspund de apărarea organismului contra agenţilor patogeni(viruşi, bacterii):
1. Steroizii (Hormonii glucocorticoizi, Prednizolon) inhibă proliferarea T-limfocitelor şisinteza anticorpilor. Inhibă toate procesele de inflamare.
2. Preparatele care au efect toxic asupra întregului sistem limfoid (ex. Azatioprin).3. Ciclosporină, Tacrolimus, Everolimus – specific inhibă formarea T-helperilor,
eliminarea selectivă a T-killerilor şi simultan stimulează T-supresori.4. Anticorpii policlonali la T-limfocite care stimulează eliminarea doar celulelor care
răspund de respingere. E o metodă cea mai puţin nocivă, ţintită, dar cea mai scumpă.Însă preparatele imunosupresive de ultima generaţie totuşi suprimează în oricare măsură şi
imunitatea antipatogenică. Imunitatea supresată dă posibilitate pentru dezvoltarea bolilorsecundare etiologia căror sunt agenţi patogeni, care se multiplică în starea imunităţii scăzute:Mediastinită, Leucocitoză, Toxoplasmoza, Pneumonie, Esofagita, Gastrită, Trombocitopenie,Sindromul limfoproliferativ, Pancreatită acută, Absces hepatic, Abces pulmonar, Insuficienţarenală şli riscul de intoxicaţie a organismului. De regulă cauza decesului sunt bolile secundare(mai des apar Insuficienţa renală şi Pneumoniile) menţionate mai sus, decît respingereatransplantului.
O altă complicaţie poate fi cancerul. Preparatele imunosupresive suprimează nu numaireacţiile imune, dar şi procesele de apoptoză, cauzînd apariţia polipilor. Caz clinic: RecipientulTony Hyuzman cu cea mai lungă durata a vieţii cu o inimă transplantată – 31 de ani. Cauzadecesului – cancerul de piele. Inima la momentul de moarte era în stare fiziologică normală şiputea încă să funcţioneze.
Alternativă la transplant1. Transplant de celule stem embrionare. Metoda cea mai eficientă pentru regenerarea
miocardului.2. O nouă descoperire a medicului şi histologului din R. Moldova – Sava Costin
(actualmente activează în Germania). Teolocite – celulele care au proprietate de aregenera miocardul afectat în urma infarctului.
3. Clonarea unei inimi noi cu structuri antigenice 100/100 compatibile cu cele alerecipientului.
4. Amprentarea genelor care codifică structura peptidică a complexului HLA (în special de clasa I)pe cromozomul 6.
Bibliografie1. Oliver, Ansel. Surgeon Bailey reflects 25 years after ‘Baby Fae. Adventist News
Network. — 2009. — October 21.2. L.B.Liausch. Catedra bioetică din RGMU (Moscova).3. Institutul de Medicină Biologică RAMS. G.T.Suhih.4. Studiile laboratorului biomagnetic IRE RAS (Moscova).5. ”Chirurgie plastică şi transplantul de organe”. Universitatea de Stat de Medicină din
Tiumeni (Rusia).

280
ETIOPATOGENIA HEMANGIOAMELORMariana Obreja
(Cond. şt.: N. Demişcan, dr. în boil., conf. univ.,E. Borş, dr. în medicină, conf. univ.,)
Catedra de fiziologie, Catedra de fiziopatologie USMF “N. Testemiţanu”
SummaryPathogenesis of hemangiomas
Today, many people worldwide suffer from a vascular disease, one of these diseases isthe hemangioma. A hemangioma is a vascular tumor which, in many cases, requires surgicalintervention. Aproximatively 30% of all hemangiomas are visible at birth. It occurs 5 time moreoften in females than in males, 83% occur on the head and neck area, the remaining 17 % appearthroughout to the rest of the body, externally and internally. The biggest problem is theappearance of this tumor. Could we understand how a hemangioma forms, we should be able toaccelerate its disappearance, or prevent its growth in the first place.
RezumatÎn prezent, multe persoane din lumea întreagă suferă de o maladie vasculara, una din
aceste maladii este hemangiomul. Hemangiomul este o tumoare vasculară care, de multe ori,necesită intervenţie chirurgicală. Aproximativ 30% din toate hemangioamele sunt vizibile lanaştere. Ele apar de 5 ori mai des la femei decît la bărbaţi, 83% apar în regiunea capului şi agîtului, restul 17% apar pe intreg corpul, pe suprafaţa externă, dar şi pe organele interne. Cea maimare problemă rămîne a fi, totuşi, apariţia acestei tumori. Dacă am putea înţelege cum seformează un hemangiom, am putea accelera dispariţia lui, sau măcar am putea preveni crestereasa.
IntroducereHemangiomul este o anomalie vasculară, care constă dintr-o aglomerare de vase capilare
nou-formate şi care se prezintă ca o pată roşie-violacee, de dimensiuni diferite. De obicei disparepînă la vîrsta de 10 ani. Potrivit diverselor surse, hemangiomul apare în 1:600 cazuri, în întregalume, ceea ce înseamnă 0,2% din populaţie. În literatura de specialitate s-a observat o diferenţăîntre incidenţa hemangioamelor la bărbaţi şi femei, astfel, raportul femei:bărbaţi este de 5:1.Nou-născuţii cu greutate mică au o şansă de 20% de a dezvolta un hemangiom.
Hemangiomul se caracterizează prin două stadii de dezvoltare. Primul este stadiul deevolutie, care se manifestă printr-o proliferare iniţială intensă, autonomă, a celulelor endoteliale,rezultînd cu un ghem de vase, pline cu sînge (poate apărea fenomenul de furt volemic). Al doileastadiu este cel de involuţie, decurge, în majoritatea cazurilor, cu resorbţia totală ahemangiomului.
A fost realizată o clasificare a tumorii, după mai multe criterii:1. după localizare:· cutanate – care afectează pielea· extracutanate – care afectează alte organe decît pielea (ficatul, SNC etc)2. după evoluţie:· care regresează spontan· care evoluează cu complicaţii (infecţii, ulceraţii, hemoragii)3. după expansiune (suprafaţă/profunzime):· plane – apar ca o pată violacee şi dispar la presiune· tumorale – de dimensiuni variabile, proemină la exterior, dar se întinde şi în
profunzime, nu se modifică la digitopresiune

281
· cavernoase – tumora este mai redusă la exterior, dar bine reprezentată în hipoderm; vaselesanguine se prezintă ca nişte caverne care comunică între ele
· cirsoide – vasele se prezintă ca nişte cordoane sinuoase, se întîlnesc într-un anevrism; lapalpare se percep pulsaţii.
4. după forme şi dimensiuni:· mici, medii, mari, cu aspect de fagure, pată, zmeură, strugure de poamă etc.
Cauzele care duc la apariţia acestei tumori încă nu sunt pe deplin elucidate, însă sepropun cîteva teorii. Astfel, nu a fost descris nici un model de transmitere familială sau genetică,există studii care demonstrează prezenţa hemangiomului la femei din trei generaţii succesive.Unii autori consideră că prezenţa tumorii la sexul feminin poate fi determinată de expresia uneigene specifice sau de sinteza hormonală crescută. Au fost raportate şi la femei însărcinate care aubeneficiat de tratament de stimulare ovariană cu gonadotropină corionică umană. O altă teoriesusţine că terapia cu steroizi estrogeni şi sarcina pot determina creşterea tumorii preexistente. Deasemenea, se suspectează că ar fi vorba şi de influenţa mediului ambiant, şi anume, gradul depoluare a aerului, utilizarea pesticidelor, un mod de viaţă nesănătos. Însă aceste idei deocamdatărămîn doar la etapa de suspiciuni.
Hemangioamele pot fi parte componentă a unor sindroame clinice bine definite, cum arfi:· Sindromul Kasabach-Merritt – prezenţa hemangioamelor hepatice gigante, asociate cu
trombocitopenie şi coagulare intravasculară diseminată.· Sindromul Osler-Rendu-Weber – prezenţa hemangioamelor mici şi numeroase ale feţei,
nărilor, buzelor, limbii, mucoasei orale, tractului gastro-intestinal şi ficatului.· Sindromul Von Hippel-Lindau – asociere de angioame cerebeloase şi ale retinei, asociate cu
leziuni hepatice şi pancreatice.· Sindromul Klippel-Trenaunay-Weber – manifestat prin edeme ale membrelor inferioare.· Lupusul eritematos sistemic etc.
Hemangiomul hepaticEste o afecţiune a ficatului care, din toate hemangioamele extracutanate, se întîlneşte cel
mai frecvent, adesea este descoperită întîmplător. După structură este asemănătorhemangioamelor din alte regiuni. Reprezintă o structură vasculară neregulată, cavernoasă şidispusă pe o stromă fibroasă, este localizat preponderent în lobul hepatic drept. Hemangiomulhepatic are culoare violet-albăstruie şi este bine delimitat de ţesutul înconjurător, de obicei, esteasimptomatic, însă tumorile multiple şi voluminoase prezintă anumite simptome. Acest tip dehemangiom face parte din tumorile congenitale care, de obicei, atrag atenţia purtătorului abiadupă jumătatea vieţii.
Manifestările cliniceAm menţionat anterior că tumorile voluminoase se pot manifesta prin anumite simptome.
De obicei, cel mai precoce simptom este o senzaţie vagă de greutate în epigastru sau hipocondruldrept; bolnavul nu-i acordă importanţă de la început şi o poate tolera 1-2 ani pînă ce să fiealarmat de apariţia simptomelor locale. Urmează senzaţia de durere, cu sediul destul de imprecis,adeseori subcostală, iradiind rareori în umăr sau subscapular drept. Durerea poate fi determinatăde mai multe cauze, ca tromboza sau infarctizarea leziunii, ruptura unui vas în interiorul tumorii,compresia ţesuturilor ş.a. Deosebit de necaracteristic şi derutînd pe medici şi bolnav poatesurveni cefaleea, cu origine necunoscută şi rezistentă la tratamentul obişnuit. În simptomatologiabolnavului pot fi descrise şi tulburări digestive variate, fie greaţă şi vomă, scăderea apetitului şisaţietate precoce, prin urmare, se observă o scădere ponderală. Aceste semne din urmă seînregistrează în cazul în care este comprimat stomacul.
De asemenea, hemangiomul se poate manifesta atipic, cu:· Insuficienţă cardiacă, datorată unor şunturi arterio-venoase masive, intratumorale

282
· Icter, apărut ca urmare a comprimării ductelor biliare· Hemobilie· Anemie· Trombocitopenie· Ruptură spontană sau posttraumatică cu hemoperitoneu· Edeme ale membrelor inferioare· Necroze· Infecţii, ulceraţii ş.a.
DiagnosticDe obicei, pacientul se adresează la medic cu dureri sau cu oricare altă maladie şi,
întîmplător, i se depistează un hemangiom extracutanat, de cele mai multe ori, hepatic. Există oserie de metode care ne pot ajuta în diagnosticarea corectă a tumorii. Aceste metode fac parte dinexamenul local primar.
Inspecţia – dacă tumora este mică, sau dacă ocupă o poziţie posterioară, regiuneasubcostală apare nemodificată; pe cînd în cazul hemangioamelor mijlocii şi mari situateanterosuperior, observăm deformarea asimetrică a peretelui abdominal anterior pe aria sfertuluisuperior drept, aici se vede o bombare care se deplasează odată cu fazele respiraţiei.
Palparea – se poate găsi fie un lob mărit, aparent regulat ca suprafaţă, fie mai ales sepoate individualiza pe suprafaţa ficatului una sau mai multe tumori cu unele caractereparticulare: de consistenţă mai puţin fermă decît parenchimul hepatic înconjurător, nu sînt netdureroase şi totuşi bolnavul se plînge în timpul examenului de senzaţii de durere.
Percuţia – nu arată altceva decît că formaţiunea tumorală este mată, asemenea ficatului.Auscultaţia este o metodă simplă de explorare, dar prea puţin utilizată în examenul
viscerelor abdominale.Tabloul clinic frust şi reducerea valorii diagnostice a simptomelor, determină sporire
importanţei aplicării metodelor auxiliare de explorare pentru confirmarea sau excludereapatologiei.
Examenul radiologic capătă o importanţă crescîndă odată cu perfecţionarea tehnicii. Înpoziţii standard, poate arăta creşterea de volum sau deformarea ficatului. Iradierea determină oregresie completă a hemangiomului, cu morbiditate minimă. Ligatura vasului central , ligaturaselectivă a vasului ce alimentează tumora reduce cu succes şuntul intratumoral, care altfel arduce la insuficienţă cardiacă cronică.
Tomografia computerizată şi rezonanţa magnetică nucleară. În prezent tomografiacomputerizată (CT) şi rezonanţa magnetică nucleară (RMN) sunt tot mai frecvent utilizate pentrudiagnosticarea hemangioamelor extracutanate, în cazuri dificile, îndeosebi la bolnavii cuobezitate concomitentă. Drept semn patologic este considerată depistarea unei zone centralehipodensă şi o captare inelară la periferie. Principalele avantaje ale acestor metode includ lipsade subiectivitate, adica au o specificitate înaltă, dependenţa mai redusă a datelor de investigator,reproductibilitatea mai bună a rezultatelor examinării de la clinică la clinică etc.
Scintigrafia – va arăta conturul şi localizarea hemangiomului ca o lacună în ficat.În ceea ce priveşte diagnosticul de laborator, probele nu arată nici o alterare; totuşi, în
literatura de specialitate, s-a descris un caz de hemangiom voluminos al lobului drept în care,paralel cu accentuarea hepatomegaliei, s-a instalat intoleranţă la novocaină; cum novocaina esteinactivată în ficat putem deduce de aici că peste o limită de creştere a tumorii vasculare potapărea unele limitări ale capacităţii funcţionale hepatice.
Concluzii1. Aşadar, ţinînd seama de caracterul progresiv al tumorii, dar şi de existenţa perioadelor, de
evoluţie lentă, este greu de cuprins un prognostic într-o formulă cu valabilitate generală.

283
2. Purtătorul unui hemangiom de dimensiuni mari este atenţionat să evite traumatismele şieforturile fizice, ce ar putea favoriza ruptura tumorii şi, în consecinţă, ar putea duce la efectemult mai grave.
3. În ceea ce priveşte tehnicile de depistare şi tratament, întrucît hemangioamele prezintă o marevarietate de forme, tehnicile trebuie adaptate la fiecare caz în parte, am putea spune, inventate.
Bibliografie1. Angelescu N., Tratat de patologie chirurgicală, Editura Medicală, Bucureşti, 20012. Grigorescu M., Tratat de hepatologie, Editura Medicală Naţională, Bucureşti, 20043. Popescu I., Chirurgia ficatului, vol. I, Editura Universitară “Carol Davila”, Bucureşti, 20044. Popescu I., Bertan M., Manual de chirurgie, vol. I, Editura Universitară “Carol Davila”,
Bucureşti, 20075. Tehnici de chirurgie vasculară, Editura Medicală Bucureşti, 2002
NEUROBIOLOGIA SOMNULUIEcaterina Chiriac
(Conducător ştiinţific – Nicolae Demişcan, doctor în biologie, conferenţiar)Catedra Fiziologia Omului şi Biofizica
SummaryThe humoral theory of sleep regulation proposed by Ishimori and Pieron postulates that
the homeostatic sleep drive leads to the accumulation of somnogens during wakefulness, and thatthis homeostatic need is discharged during sleep. Prostaglandin D2, adenosine, and cytokineshave been proposed as effective sleep substances. Also, several interactions between sleep- andwake-active neurons have been proposed at the systems level in many models of sleep–wakeregulation. In this review, we describe briefly the neurobiology of sleep.
RezumatTeoria umorală a reglării somnului propusă de Ishimori şi Pieron postulează că în timpul
stării de veghe în organism se acumulează substanţe somnogene, care asigură homeostazasomnului. Prostaglandina D2, adenozina şi citokinele au fost propuse drept substanţe somnogeneefective. De asemenea, s-a descoperit faptul că reglarea ciclului somn-veghe este realizată prininteracţiunea dintre diverşi neuroni ce controlează starea de veghe şi somnul. În această lucrare,realizăm o scurtă trecere în revistă a neurobiologiei somnului.
ActualitateaLa începutul secolului 20, japonezul Kuniomi Ishimori şi francezul Henri Pieron au
descoperit independent că un cîine căruia i-a fost injectat intracerebral lichidului cerebrospinal alaltui cîine ţinut în stare de veghe pentru o perioadă lungă de timp a adormit, presupunînd astfelexistenţa unor „substanţe ce induc somnul” şi care se acumulează în creier în timpul stării deveghe.
Conceptul factorului somnogenO substanţă poate fi somnogenă atunci cînd îndeplineşte următoarele condiţii:
- concentraţia ei să fie mai înaltă în timpul stării de veghe comparativ cu cea de somn;- să se micşoreze în timpul somnului şi să se mărească treptat în timpul stării de vegheîndelungată;- să aibă capacitatea de a inhiba activitatea neuronală responsabilă de reglarea stării de veghe(6).

284
Neurofiziologia ciclului somn-vegheStarea de veghe şi conştiinţă depinde de activitatea neuronilor sistemului ascendent
activator reticular al creierului. Aceşti neuroni au proiecţii la nivelul talamusului,hipotalamusului, porţiunii bazale a prozencefalului (PB) şi cortexului. O serie deneurotransmiţători, precum catecolaminele, acetilcolinele, histamina şi aspartatul sînt elaboraţi lanivelul substanţei reticulare şi au un rol important în activarea corticală.
Neurofiziologia somnului NREMPorţiunea anterioară a hipotalamusului, aria adiacentă a prozencefalului, talamusul, rafeul
dorsal şi nucleul tractului solitar sînt structurile reponsabile de reglarea somnului NREM.Neurotransmiţătorii precum serotonina şi GABA sînt localizaţi în aceste regiuni şi joacă un rolimportant în reglarea somnului.
Serotonina (5-HT), localizată la nivelul neuronilor rafeului, poate fi implicată îninducerea somnului. S-a demonstrat apariţia insomniei la lezarea celulelor serotoninergice alerafeului dorsal. Inhibitorii MAO (inhibă şi degradarea 5-HT), substanţele care participă labiosinteza serotoninei, precum triptofanul şi vitamina B6, facilitează inducerea somnuluifiziologic (9).
Neurotransmiţătorul inhibitor GABA (acidul gama-aminobutiric) este eliberat în cantităţimari pe parcursul somnului NREM. Neuronii gabaergici sunt localizaţi pretutindeni în creier,inclusiv la nivelul prozencefalului, hipotalamusului, talamusului, trunchiului cerebral şicortexului. GABA blochează neuronii activatori ai substanţei reticulare şi inhibă transmisia prinproiecţiile lor corticale şi talamice.
Neurofiziologia somnului REMAcetilcolina este localizată în neuronii tegmenului pontin şi este implicată în reglarea
somnului REM. S-a demonstrat faptul că neuronii care induc somnul REM sînt colinergici,localizaţi în ariile reticulare laterale şi mediale pontine şi trimit eferenţe talamusului,hipocampului şi hipotalamusului. Aceşti neuroni prezintă o activitate maximă în timpul somnuluiREM şi o activitate joasă sau nulă în timpul somnului NREM. Experimental, s-a demonstrat căfizostigmina (anticolinesterazic), precipitează apariţia somnului REM în timpul somnuluiNREM. Injectarea carbacolului (agonist muscarinic) la nivelul tegmentului pontin induce somnulREM, iar blocarea receptorilor muscarinici întîrzie apariţia acestuia (5).
Neuronii ce blochează somnul REM sînt noradrenegici şi serotoninergici, localizaţi lanivelul rafeului şi locusului ceruleus. Aceşti neuroni prezintă o activitate minimă sau nulă întimpul somnului REM. Astfel, modificarea nivelului norepinefrinei şi serotoninei poate afectasomnul REM (13).
Neuroanatomia ciclului somn-vegheCiclul somn-veghe este reglat de către următoarele structuri cerebrale:
- Trunchiul cerebral – aici sînt localizaţi neuronii glutamatergici, colinergici şi gabaergici ceinduc somnul REM, neuronii serotoninergici ce controlează starea de veghe şi neuroniiacetilcolinici ce controlează atît starea de veghe, cît şi somnul REM;- Hipotalamusul posterior şi lateral - neuronii orexinici şi noradrenergici ce reglează starea deveghe;- Aria preoptică - neuronii gabaergici inductori ai somnului localizaţi la nivelul porţiuniiventro-laterale a ariei preoptice hipotalamice şi în nucleul preoptic median;- Porţiunea bazală a prozencefalului - neuronii colinergici activatori ai stării de veghe, neuroniigabaergici ce induc somnul (6).
Substanţele ce reglează ciclul somn-vegheProstaglandina D2 (PGD2)În cadrul unui studiu asupra funcţiei PGD2 la nivelul SN, a fost descoperită acţiunea
acesteia de inducere a somnului. S-a descoperit faptul că nivelul PGD2 în lichidul cefalo-

285
rahidian prezintă o fluctuaţie circadiană, odată cu ciclul somn-veghe, ridicîndu-se în timpulprivării de somn.
Au fost identificaţi 2 receptori ai PGD2: receptorul DP1, identificat drept omolog al altorreceptori prostaglandinici şi DP2, identificat drept receptor chemoatractant pentru PGD2.Receptorii DP1 sunt localizaţi preponderent inferior de porţiunea rostrală a PB lateral de chiasmaoptică, în apropierea porţiunii posterioare a hipotalamusului, care conţine centrii de reglare aciclului somn-veghe. Distribuţia celulară a DP2 în creier încă nu a fost clarificată (11).
Mecanismul molecular al somnului indus de PGD2 - somnogenul endogen PGD2 esteprodus de L-PGDS (prostaglandin-sintetaza tip lipocainic), circulă prin lichidul cefalorahidian,stimulează receptorii DP1 ai suprafeţei ventrale cuprinsă între PB şi hipotalamus, şi măreşteconcentraţia adenozinei. Adenozina difuzează în parenchimul creierului jucînd rolul unuisomnogen secundar, inhibă neuronii excitatori ai PB prin intermediul receptorilor A1 şi activeazăneuronii somnului prin intermediul receptorilor A2A inducînd astfel somnul (12).
Inhibiţia centrului de excitaţie histaminergic în timpul somnului indus de agoniştiisistemului PGD2-A2A – studiile experimentale au demonstrat că în timpul somnului indus deagoniştii PGD2 sau a receptorilor adenozinei A2A, numărul neuronilor histaminergici activi dinnucleul tuberomamilar se micşorează. In vivo, microdializa experimentală a agonistuluireceptorilor A2A (CGS21680) la nivelul PB a inhibat eliberarea de histamină în cortexul frontalşi porţiunea medială a ariei preoptice, şi a mărit eliberarea GABA în nucleul tuberomamilar (7).
AdenozinaCreşterea concentraţiei de adenozină sugerează intensificarea consumului de energie,
reflectată de hidroliza ATP-ului.În SNC adenozina este un neuromodulator inhibitor al neuronilor excitatori (precum cei
colinergici sau glutamatergici). Rata formării adenozinei este reglată de către activitateaneuronală: nivelul ei începe să crească cînd aprovizionarea cu oxigen este diminuată, înasemenea circumstanţe precum ischemia, hipoxia şi hipoperfuzia. Astfel, se presupune căadenozina participă la minimalizarea leziunilor tisulare, micşorînd activitatea celulelor şi, drepturmare, necesitatea acestora de oxigen (8).
Receptorii adenozinei – dintre subtipurile receptorilor adenozinei (A1, A2A, A2B şi A3),receptorii A1 şi A2A sînt implicaţi în procesul reglării somnului. Receptorii A1 sînt largrăspîndiţi în creier şi inhibă neuronii stării de veghe, precum cei colinergici localizaţi în PB şihistaminergici localizaţi în porţiunea posterioară a hipotalamusului, iar receptorii A2 sîntlocalizaţi în PB şi activează doar neuronii somnului (12).
Nivelul adenozinei în timpul stării de veghe de lungă durată – în primele experimente pepisici, concentraţia adenozinei a crescut constant în PB pe parcursul primelor 6 ore de veghe,atingînd dublul cantităţii măsurate la începutul acestei perioade. În timpul somnului, niveluladenozinei a scăzut treptat în următoarele 2-3 ore. În talamus, nucleul dorsal al rafeului şitegmental pedunculo-pontin, şi aria preoptică adenozina nu a crescut în timpul celor 6 ore deveghe, deşi ele de asemenea sînt cunoscute ca zone ce reglează somnul. Creşterea regională anivelului adenozinei în timpul stării de veghe sugerează faptul că adenozina induce somnul prininhibiţia neuronilor corticali cu proiecţie în PB, responsabili de menţinerea ei (3).
Originea creşterii nivelului adenozinei în timpul perioadei de veghe de lungă durată –creşterea activităţii neuronale este semnalul de bază pentru creşterea nivelului adenozinei, însădeoarece astrocitele furnizează energie neuronilor în timpul activităţii maxime a acestora,adenozina poate proveni fie din înşişi neuronii sau din astrocitele învecinate. S-a demonstratfaptul că homeostaza somnului poate fi dereglată la animalele cu deficienţa genetică agliotransmisiei, nivelul adenozinei la acestea fiind diminuat, sugerînd astfel faptul că nivelulcrescut de adenozină în timpul stării de veghe îşi are originea din astrocite (1).
Mecanismele de acţiune ale adenozinei:

286
- PB – numeroase studii au demonstrat faptul că adenozina în PB influenţează somnul prinintermediul receptorilor A1: dacă creşterea nivelului adenozinei la nivelul PB în timpul perioadeiprelungită de veghe este suprimată de către antagoniştii receptorilor A1, somnul nu este indus.Efectul adenozinei în PB se bazează pe inhibiţia neuronilor colinergici cu proiecţie corticală.- Hipotalamusul – adenozina inhibă neuronii veghii prin intermediul receptorilor A1 şi inducesomnul NREM prin inhibiţia neuronilor histaminergici din nucleul tuberomamilar. Comparativcu alte regiuni ale creierului, agoniştii receptorilor adenozinei A2A de asemenea induc somnulprin inhibarea eliberării de histamină în nucleul tuberomamilar, efect mediat de eliberareaGABA. În hipotalamusul lateral, este prezentă o altă zonă responsabilă de reglarea stării deveghe-somn, la nivelul căreia agoniştii şi antagoniştii receptorilor adenozinei A1 induc sausuprimă somnul NREM/REM.- Alte zone ale creierului – administrarea adenozinei în cortexul prefrontal, de asemeneamodifică statusul de vigilenţă. Ca şi în alte zone ale creierului, agoniştii A1 induc somnul(micşorînd eliberarea de acetilcolină). În plus, un antagonist specific al receptorilor A1, cofeina,măreşte eliberarea de acetilcolină şi induce veghea, pe cînd antagoniştii A2A sînt inefectivi,sugerînd faptul că cofeina la nivel cortical măreşte excitabilitatea acestuia prin intermediulreceptorilor A1. Regiunea punţii, care este reglatorul central al somnului REM, este influenţatăde adenozină: agoniştii adenozinici injectaţi în regiunea formaţiunii reticulare pontine oral-caudale induc somnul REM de lungă durată (8).
Peptidele ce induc somnul deltaSînt peptide localizate la nivelul neuronilor, organelor periferice şi plasmei. Aceste
neuropetide induc somnul delta la mamifere. În afară de influenţa asupra somnului, s-ademonstrat faptul că ele afectează activitatea electrofiziologică şi nivelul neurotransmiţătorilor lanivelul creierului, ritmul circadian, nivelul hormonilor, performanţa fizică şi activitateapreparatelor medicamentoase neurotrope.
PGE2Fiind administrată la nivelul hipotalamusului posterior, PGE2 induce starea de veghe. S-
a determinat faptul că PGE2 creşte sinteza şi eliberarea histaminei la nivelul nucleuluituberomamilar. Dintre agoniştii celor 4 subtipuri de receptori ai PGE2 testaţi, doar agonistulreceptorului EP4 mimează efectul excitator al PGE2. Prin hibridizare in situ s-a observat faptulcă mRNA-ul receptorului EP4 este exprimat la nivelul nucleului tuberomamilar. Mai mult ca atît,perfuzia agonistului EP4 la acest nivel induce starea de veghe. Aceste descoperiri demonstreazăfaptul că PGE2 induce starea de veghe prin intermediul sistemului histaminergic (14).
HISTAMINANeuronii histaminergici sînt localizaţi în nucleul tuberomamilar, la nivelul porţiunii
posterioare a hipotalamusului. Ei trimit ramuri eferente tuturor regiunilor creierului. Histaminajoacă un rol important în reglarea ciclului somn-veghe, prin intermediul altor mediatori aicreierului precum PG2, PGE2, orexina/hipocretina şi adenozina (2).
Orexina/hipocretinaNeuronii orexin sunt localizaţi exclusiv la nivelul ariei laterale a hipotalamusului şi au
fibre ce se proiectează la nivelul întregului sistem nervos central, inclusiv nucleultuberomamilar, unde receptorii orexinei au o densitate mai mare (10). Perfuzia orexinei A lanivelul acestui nucleu la şoareci, induce prompt starea de veghe, concomitent reducînd somnulREM şi NREM. Studiile au arătat că orexina A măreşte eliberarea de histamină din porţiuneamedială a ariei preoptice şi cortexul frontal. Mai mult ca atît, infuzia orexinei A la nivelulventricolului lateral al şoarecilor sănătoşi induce starea de veghe, pe cînd la cei fără genareceptorului histaminic H1, nu o induce. Aceste descoperiri demonstrează faptul că orexina Aactivează neurotransmisia histaminergică, inducînd astfel starea de veghe prin intermediulreceptorului histaminic H1 (4).

287
Concluzii1. Structurile creierului implicate în reglarea ciclului somn-veghe sînt: trunchiul cerebral,
hipotalamusul posterior şi lateral, aria preoptică şi porţiunea bazală a prozencefalului;2. Substanţele chimice ce influenţează apariţia somnului sunt: serotonina, GABA, PGD2,
adenozina, peptidele delta;3. Substanţele somnogene se acumulează în organismul uman în timpul stării de veghe,
inducînd astfel somnul.
Bibliografie1. Bertil B Fredholm. Physiological and pathophysiological roles of adenosine. Sleep and
Biological Rhythms 9 (Suppl. 1): 24–28, 20112. Haas HL, Sergeeva OA, Selbach O. Histamine in the nervous system. Physiol. Rev. 88:
1183–241, 20083. Jiang-Fan Chen and Michael A. Schwarzschild. Adenosine A2A receptors and
neuroadaptation: Possible implications in sleep-wake physiology. Sleep and BiologicalRhythms 2: S19–S20, 2004
4. Junko HARA and Takeshi Sakurai. Interaction between sleep mechanisms and orexinneurons. Sleep and Biological Rhythms 9 (Suppl. 1): 38–43, 2011
5. Larry D. Sanford, Xiangdong Tang. GABAergic Regulation of REM Sleep in ReticularisPontis Oralis and Caudalis in Rats. J Neurophysiol 90:938-945, 2003
6. Martha S. Rosenthal. Physiology and Neurochemistry of Sleep. American Journal ofPharmaceutical Education Vol. 62, 1998
7. Osamu Hayaishi. Humoral mechanisms of sleep–wake regulation: Historical review ofprostaglandin D2 and related substances. Sleep and Biological Rhythms 9 (Suppl. 1): 3–9,2011
8. Porkka-Heiskanen T, Alanko L, Kalinchuk A, Stenberg D. Adenosine in sleep regulation.Sleep Med. Rev 6: 321–32, 2002
9. Reidun Ursin. Serotonin and sleep. Sleep Medicine Reviews, Vol. 6, No. 1, pp 57–69, 200210. Seiji Nishino and Yasushi Yoshida. History and perspectives of hypocretin/orexin research in
sleep medicine. Sleep and Biological Rhythms 1: 43–54, 200311. Ueno R, Ishikawa Y, Nakayama T, Hayaishi O. Prostaglandin D2 induces sleep when
microinjected into the preoptic area of conscious rats. Biochem. Biophys. Res. Commun 109:576–82, 1982
12. Yoshihiro Urade. Prostaglandin D2 and adenosine as endogenous somnogens. Sleep andBiological Rhythms 9 (Suppl. 1): 10–17, 2011
13. Yuichi Tamakawa, Akihiro Karashima. A Quartet Neural System Model Orchestrating Sleepand Wakefulness Mechanisms. J Neurophysiol 95:2055-2069, 2006
14. Z-L Huang, W-M Qu. Histaminergic role in sleep-wake cycle of orexin, adenosine, andprostaglandin E2 and D2. Sleep and Biological Rhythms 2: S21–S22, 2004