expression of proto-ret mrna in embryonic and adult rat tissues
TRANSCRIPT

Vo1.153, No. 3,1988
June 30,1988
BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
Pages 1290-1295
EXPRESSION OF PROTO-ret mRNA IN EMBRYONIC AND ADULT RAT TISSUES
Tomoko T~hira, Yukihito Ishizaka, Takashi Sugimura and Minako Nagao
Carcinogenesis Division, National Cancer Center Research Inst i tute, I - I , Tsukij i 5-chome, Chuo-ku, Tokyo 104, Japan
Received May 23, 1988
SUMMARY: The expression of proto-ret mRNA in adult and embryonic rat tissues were studied. Very low levels of proto-ret transcripts were found in adult rat tissues such as brain, thymus and testis. The sizes of these transcripts were almost the same as those found in human neuroblastema, SK-N-SH cells. High levels of proto-ret transcripts vmre found in the rat conceptus on days 9 to 11 of gestation, but not at later stages ef development. The level of transcripts in the conceptus on day 10 was about 20-50 times that in adult rat thymus. These results suggest that the proto-ret product, which is possibly a receptor-type tyrosine kinase, has special functions during embryonic development, o 19BSA=~i° P~e,~. zn°.
The ret oncogene was f i r s t isolated as a dominant transforming gene of
NIH3T3 cells after transfection with human T-cell lymphoma DNA (1). We
isolated re t - I I from an NIH3T3 transformant induced by DNA of a human colon
cancer (2). These two transforming genes, ret and re t - I I , have the 3' portion
of the proto-ret in common, but di f fer in the non-proto-ret sequences of their
5' parts. These two recembinants seem to be produced during the transfection
process. The biological significance of proto-ret has not been c lar i f ied,
except for the demonstration of the transforming act iv i t ies of these two
recombinants.
Recently, the nucleotide sequence of proto-ret cDNA was determined. The
deduced proto-ret product has a transmembrane domain followed by a tyrosine
kinase domain, and was proposed to be a receptor-type tyrosine kinase
(Takahashi et a l . , personal communication). The expression of proto-re_t mRNA
was detected in SK-N-SH neuroblastoma, HL-60 promyelocytic leukemia and THP-1
monocytic leukemia cell lines among 28 human tumor cell lines tested (3).
To elucidate the biological significance of proto-ret, we examined i ts
expression in various tissues of adult rats. We also studied the levels of
proto-ret expression during embryonic development of rats.
0006-291X/88 $1.50 Copyright © 1988 by Academic Press, Inc. All rights of reproduction in any form reserved. 1290

Vol. 153, No. 3, 1988 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
MATERIALS AND METHODS
Tissues and cells Adult rat tissues were obtained from 6-week-old F344 rats (Charles River Japan, Atsugi, Kanagawa). The conceptus and uterus were obtained from pregnant F344 rats (Shizuoka Laboratory Animal Center). The day of plug formation was defined as day 0 of gestation by the suppliers.
SK-N-SH neuroblastoma cells (4) were cultured in Dulbecco's modified Eagle's medium supplemented with 10% heat-inactivated fetal bovine serum and non-essential amino acids.
RNA blot analysis Total RNA was extracted by the guanidinium/cesium chloride method, and poly(A)+RNA was isolated on an oligo(dT)-cellulose column essentially as described elsewhere (5). RNA concentrations were determined by measurements of the optical density at 260 nm. Samples of 5 ~g of poly(A)+RNA were fractionated in 0.8% formaldehyde/agarose denaturing gel and blotted onto nitrocellulose f i l t e rs (6). Blots were hybridized with 32p-labeled probes in 50% formamide/O.65 M NaCl/O.1 M sodium Pipes (pH 6.8)/0.1% sodium dodecyl- sulfate/5 mM EDTA/IO% dextran sulfate/salmon testis DNA (100 ~g/ml) at 42°C for 16 hr, and washed at 50°C for 80 min with four changes of 2 x SSC/O.2% sodium pyrophosphate/O.1% sodium dodecylsulfate.
Probes Fragments of re t - I I and ret cDNAs were used to detect proto-ret mRNA. ~ e fragments were a 0.4 kb E~I-NcoI fragment of re t - I I cDNA, which was cloned from a cDNA l ibrary of a secondary transformant ( ~ i z a k a et al, in preparation) and a 0.2 kb BamHI fragment of ret cDNA generously provided by Dr. M. Takahashi (3). The'-f-ormer corresponds to a portion of the region encoding the proto-ret kinase domain, and the lat ter includes the region encoding the transme-m-brane domain. A 1.3 kb HpaII-EcoRI fragment of the rat B-actin gene, which specif ical ly detects cytoplasmic B-actin (7) and a 1.3 kb rat glyceraldehyde-3-phosphate dehydrogenase cDNA (8) were used as controls for loaded RNA. These fragments were gel-purif ied and labeled with 32p by nick translation. The specific act iv i t ies of the probes were 1-2 x 109 dpm/ug DNA.
RESULTS
Expression of proto-ret mRNA in adult rat tissues RNA blot analysis o f
poly(A)+RNAs from various rat tissues was carried out with the 0.4 kb EcoRI-
NcoI fragment of re t - I I cDNA as a probe (Fig. 1). This probe detected a 6.9
kb EcoRI fragment in human DNA and an 8.2 kb EcoRI fragment in rat DNA by
Southern blot hybridization under the same conditions as those used in RNA
blot hybridization. The intensity of the bands in rat DNA was less than 20%
of that in human DNA, reflecting the divergence due to species differences in
the proto-ret gene (data not shown). Four species of transcripts (approx-
imately 7.0, 6.0, 4.5 and 3.9 kb in size) were detected with this probe in
poly(A)+RNAs of certain human cel ls, such as SK-N-SH as reported by Takahashi
et al. (3).
The levels of proto-ret transcripts were very low in the rat tissues
examined and could be detected only after long exposure to X-ray fi lm (Fig.
1). Relatively strong signals were observed in samples of brain, thymus and
test is. Lung, heart, spleen and small intestine also expressed detectable
levels of proto-ret transcripts. We could not detect proto-ret transcripts in
tissues such as bone marrow, l iver and kidney, which showed definite bands
with probes for GAPDH and/or 8-actin.
I291
Vol. 153, No. 3, 1988 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
k" I 7 . 0 - - 6.0 4 . 5
3 . 9
proto-ret
: 'v.: :}~ : i i~',',.'~i ~P:H{~-~ *':~:'~" " " ~ ; h i l ; ' ~ i 1 .3 i~i j""=~* " "~;.~.~ii~*~L. • ~i'~ - ~ ,!~ i iw, . . . . . . . . . N , ,
. . . . . . . . • - . -@ ~.i .... ,:~ . . . . . . . . . . . . ~... . . 2 . 1 ~ -~ ~ : , , - . . : ~ ; ~ = ,~ 13-actin
i ! ~ ! ; ! ? ! " ~ ' ~ ! ~ r ~ ! 4 ~ ° ! !' ~!~;~!~:~ ':~:~:~:~: !;~:::i
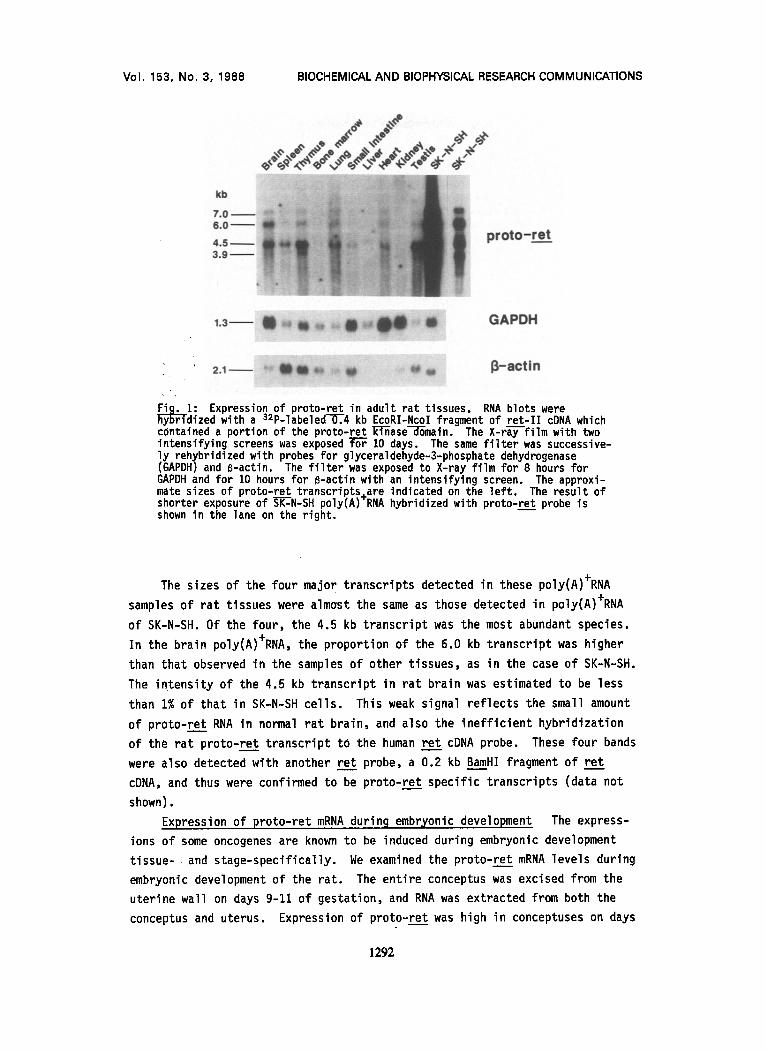
~ r 1: Expression of proto-ret in adult rat tissues. RNA blots were dized with a S2p-labeled--O'~.4 kb EcoRI-Ncol fragment of ret-II cDNA which
contained a portion of the proto-ret ~ITi-6ase~main. The X-ra-y-film with two intensifying screens was exposed ~ 10 days. The same f i l ter was successive- ly rehybridized with probes for glyceraldehyde-3-phosphate dehydrogenase (GAPDH) and B-actin. The f i l ter was exposed to X-ray film for 8 hours for GAPDH and for 10 hours for B-actin with an intensifying screen. The approxi- mate sizes of proto-ret transcripts±are indicated on the left. The result of shorter exposure of ~-RT-N-SH poly(A)~RNA hybridized with proto-ret probe is shown in the lane on the right.
The sizes of the four major transcripts detected in these poly(A)+RNA
samples of rat tissues were almost the same as those detected in poly(A)+RNA
of SK-N-SH. Of the four, the 4.5 kb transcript was the most abundant species.
In the brain poly(A)+RNA, the proportion of the 6.0 kb transcript was higher
than that observed in the samples of other tissues, as in the case of SK-N-SH.
The intensity of the 4.5 kb transcript in rat brain was estimated to be less
than 1% of that in SK-N-SH cells. This weak signal reflects the small amount
of proto-ret RNA in normal rat brain, and also the ineff icient hybridization
of the rat proto-ret transcript tO the human ret cDNA probe. These four bands
were also detected with another ret probe, a 0.2 kb BamHI fragment of ret
cDNA, and thus were confirmed to be proto-ret specific transcripts (data not
shown).
Expression of proto-ret mRNA durin 9 embryonic development The express-
ions of some oncogenes are known to be induced during embryonic development
t i ssue-and stage-specifically. We examined the proto-ret mRNA levels during
embryonic development of the rat. The entire conceptus was excised from the
uterine wall on days 9-11 of gestation, and RNA was extracted from both the
conceptus and uterus. Expression of proto-ret was high in conceptuses on days
1292
Vol. 153, No. 3, 1988 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
k~ ~:!i !iii " . i !: ili i.i i:i~.. ~ i 7 .0 ~ !iii~ ::.i~ii~:!?~i:.ii:i~!i)~i:,:i immL.
3.94"5 i..!ii ):ii ~ ~:!!i~, ! il ,-:~(iiiii?~ii! "i~::; :.~ihi~L " :~I .
®
proto-ret
i3-actin
kb
7 . 0 ~ 6.0
4 .5 3.9
2.1
/
OOmlooee-oe
proto-re_.tt
(3-actin
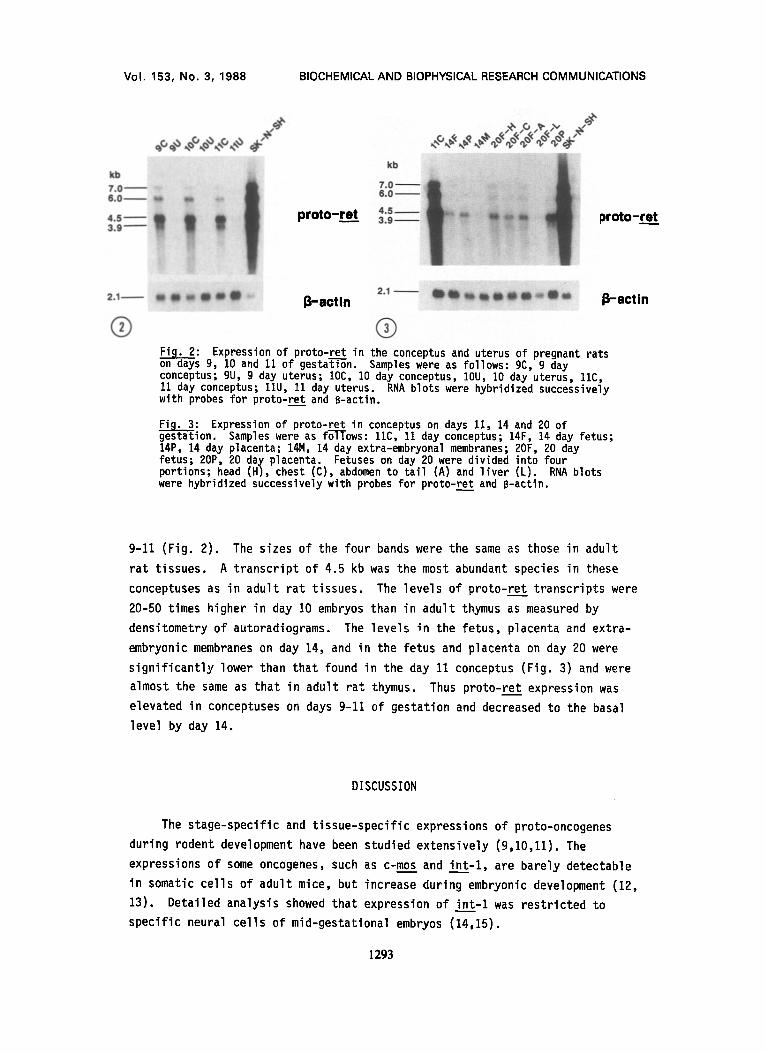
® Fi__~: Expression of proto-ret in the conceptus and uterus of pregnant rats on nays g, 10 and 11 of gestat-Ton. Samples were as follows: gC, g day conceptus; gU, 9 day uterus; IOC, 10 day conceptus, IOU, 10 day uterus, 11C, 11 day conceptus; 11U, 11 day uterus. RNA blots were hybridized successively with probes for proto-ret and B-actin.
Fig. 3: Expression of proto-ret in conceptus on days 11, 14 and 20 of gestation. Samples were as fo--TTows: 11C, 11 day conceptus; 14F, 14 day fetus; 14P, 14 day placenta; 14M, 14 day extra-embryonal membranes; 2OF, 20 day fetus; 20P, 20 day placenta. Fetuses on day 20 were divided into four portions; head (H), chest (C), abdomen to tail (A) and liver (L). RNA blots were hybridized successively with probes for proto-ret and B-actin.
9-11 (Fig. 2). The sizes of the four bands were the same as those in adult
rat tissues. A transcript of 4.5 kb was the most abundant species in these
conceptuses as in adult rat tissues. The levels of proto-ret transcripts were
20-50 times higher in day IO embryos than in adult thymus as measured by
densitometry of autoradiograms. The levels in the fetus, placenta and extra-
embryonic membranes on day 14, and in the fetus and placenta on day 20 were
signif icantly lower than that found in the day 11 conceptus (Fig. 3) and were
almost the same as that in adult rat thymus. Thus proto-ret expression was
elevated in conceptuses on days 9-11 of gestation and decreased to the basal
level by day 14.
DISCUSSION
The stage-specific and tissue-specific expressions of proto-oncogenes
during rodent development have been studied extensively (9,10,11). The
expressions of some oncogenes, such as c-mos and int-1, are barely detectable
in somatic cells of adult mice, but increase during embryonic development (12,
13). Detailed analysis showed that expression of int-1 was restricted to
specific neural cells of mid-gestational embryos (14,15).
1293

Vol. 153, No. 3, 1 9 8 8 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
We found that the expression of proto-ret was very low in adult rat
tissues but that in conceptuses i t was significantly increased on days 9-11
and decreased to the basal level by day 14 of gestation.
I t is not yet known which embryonic and extra-embryonic tissues express
proto-ret. We are now carrying out in situ hybridization studies to locate
organs expressing proto-~et.
Various hormones and growth factors are produced in conceptus on days
9-11. Recently Han et al. (16) reported that expression of rat TGF~ in the
maternal decidua reaches a peak on day 8, and discussed the role of TGFe in
cellular proliferation through an autocrine, paracrine or endocrine mechanism.
A similar situation was proposed for c-fms, which is a receptor-type tyrosine
kinase (17). c-fms and i ts ligand, CSF-1, are highly expressed during fetal
development (18,19). Proto-ret may be involved in cellular proliferation or
differentiation at this stage of development.
We examined the expression of proto-ret in three human choriocarcinoma
cell lines, BeWo (20), JEG-3 (21) and JAR (22), and in the mouse terato-
carcinoma F9 (23) cell line by RNA blot analysis. No proto-ret transcripts
were detected in the total RNAs of these cell lines (data not shown).
Proto-ret expressed four species of transcripts in all samples tested.
The mechanisms of formation of these four transcripts are unknown, ret and
re t - I I , which have non-proto-ret sequences in their 5' halves, also expressed
four transcripts with the same intervals of size. Therefore these four
transcripts may be produced by alteration in the 3' halves. Our preliminary
data suggested that some of these are produced by alternative splicing
(Ishizaka et al . , in preparation). The proportions of the intensities of each
band differed sl ightly in brain and other tissues including the conceptus. We
are now studying whether these four transcripts have different biological
significances.
ACKNOWLEDGEMENT
We are grateful to Dr. M. Takahashi for cDNA clones of ret, Dr. S. Sakiyama for a subclone of the rat cytoplasmic B-actin gene,'-b-r. Ph. Jeanteur for a cDNA clone for rat glyceraldehyde-3-phosphate dehydrogenase mRNA, and Dr. M. Akiyama for SK-N-SH cells.
This study was supported by a grant-in-aid from the Ministry of Health and Welfare for a Comprehensive 1D-Year Strategy for Cancer Control, Japan.
REFERENCES
1. Takahashi, M., Ritz, J . , and Cooper, G. M. (1985) Cell 42, 581-588. 2. Ishizaka, Y., Tahira, T., Ochiai, M., Ikeda, I . , Sugimura, T., and Nagao,
M. (1988) Oncogene Res. in press.
1294

Vol. 153, No. 3, 1988 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
3. Takahashi, M., and Cooper, G. M. (1987) Mol. Cell. Biol. 7, 1378-1385. 4. Biedler, J. L., Helson, L., and Spengler, B. A. (1973) Cancer Res. 33,
2643-2652. 5. Maniatis, T., Fritsch, E. F., and Sambrook, J. (1982) Molecular Cloning:
Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY.
6. Thomas, P. S. (1980) Proc. Natl. Acad. Sci. USA 77, 5201-5205. 7. Nudel, U., Zakut, R., Shani, M., Neuman, S., Levy, Z., and Yaffe, D.
(1983) Nucl. Acids Res. 11, 1759-1771. 8. Fort, Ph., Marty, L., Piechaczyk, M., El Sabrouty, S., Dani, Ch.,
Jeanteur, Ph., and Blanchard, J. M. (1985) Nucl. Acids Res. 13, 1431- 1442. MUller, R., Slamon, D. J. , Tremblay, J. M., Cline, M. J. , and Verma, I. M. (1982) Nature 299, 640-644. MSller, R., Tremblay, J. M., Adamson, E. D., and Verma, I. M. (1983)
, 454-456. J. , and Cline, M. J. (1984) Proc. Natl. Acad. Sci. USA, 81,
Nature 304 Slamon, D. 7141-7145. Propst, F., (1987) Mol. Jakobovits, (1986) Proc. Wilkinson, D 79-88. Schackleford Han, V. K. M D. C. (1987) Sherr, C. J.
,
10.
I I .
12.
13.
14.
15. 16.
17.
18.
19.
20. 21. 22.
23.
Rosenberg, M. P., lyer, A., Kaul, K., and Vande Woude, G. F. Cell. Biol. 7, 1629-1637. A., Shackleford, G. M., Varmus, H. E., and Martin, G. R. Natl. Acad. Sci. USA 83, 7806-7810.
. G., Bailes, J. A., and McMahon, A. P. (1987) Cell 50,
, G. M., and Varmus, H. E. (1987) Cell 50, 89-95. ., Hunter I I I , E. S., Pratt, R. M., Zendegui, J. G., and Lee, Mol. Cell. Biol. 7, 2335-2343.
, Rettenmier, C. W., Sacca, R., Roussel, M. F., Look, A. T., and Stanley, E. R. (1985) M~ller, R., Slamon, D. J. Cline, M. J. , and Verma, Pollard, J. W., Bartocci, and Stanley, E. R. (1987)
Cell, 41, 665-676 , Adamson, E. D., Tremblay, J. M., M~ller, D., I . M. (1983) Mol. Cell. Biol . , 3, 1062-1069 A., Arceci, R., Orlofsky, A., Ladner, M. B., Nature 330, 484-486.
Pat t i l lo , R. A., and Gey, G. O. (1968) Cancer Res. 28, 1231-1236 Kohler, P. 0., and Bridson, W. E. (1971) J. Clin. Endocrinol. 32, 683-687 Pat t i l lo , R. A., Ruckert, A., Hussa, R., Bernstein, R., and Delfs, E. (1971) In Vitro 6, 398-399 Bernstine, E. G., Hooper, M. L., Grandchamp, S., and Ephrussi, B. (1973) Proc. Natl. Acad. Sci. USA 70, 3899-3903
1295