expression of ?cell-cycle-dependent? genes in regenerating rat liver
TRANSCRIPT

Cell Biology International Reports, Vol. 12, No. 6, June 1988 483
EXPRESSION OF "CELL-CYCLE-DEPENDENT" GENES
IN REGENERATING RAT LIVER
E. Biesiada and M.Choraky*
Department of Tumor Biology, Institute of bncology 44-100 Gliwice, Poland
ABSTRACT The expression of genes coding for the ATP/ADP
translocase, calcyclin, ornithine decarboxylase, vimentin, proto-onc genes ~53 and c-Ha-r-1 and also for two genes JE and KC with as yet unknown function was studied during regeneration of rat liver. Genes highly induced were: JE (2-8 h of regeneration), ATP/ADP translocase (8-18 h), c-Ha-r--l (6-48 h) and ~53 (6-12 h). Vimentin and KC gene transcripts were not detectable in the first 48 h of liver regeneration, whereas ornithine decarboxylase and calcyclin gene transcripts were present at constant levels. Our findings extend the list of genes expressed at the early stages of liver regeneration.
I NT'RODUCTION When quiescent mammalian cells are stimulated to
proliferate a number of genes called "cell-cycle dependent" (CCD) genes are activated and their transcripts (mRNAs) accumulate in abundant amounts. Various cell lines (mostly fibroblasts) grown in cell culture, and T lymphocytes have been used to study expression of genes during the course of the cell-cycle (for review see Kaczmarek,l986; Denhardt et a1.,1986).
Rat hepatocytes which rarely divide, start to proliferate in a synchronized way in response to partial hepatectomy (for review see Bucher et al., 1983 ; Sobczak and Duguet.1986; and ref. therein). Starting from about 8 h after hepatectomy UP to 18-20 hours, growth $n size of hepatocytes is accompaniend by active poly(A) RNA synthesis and assembly of polysomes.
* To whom correspondence should be addressed
030~1651/66/060463-1 O/$03.00/0 @ 1966 Academic Press Ltd.

484 Cell Biology International Reports, Vol. 12, No. 6, June 1988
DNA replication starts at about 12 h and reaches a peak at 22-24 h. The first burst of mitoses is observed at 32 h after surgery. In the initial 48 hours of the regeneration only hepatocytes and not other liver cell types exhibit a proliferative response to hepatectomy thus offering a unique model to study the expression mode of CCD genes in a living animal.
Regenerating rat hepatocytes exhibit a specific pattern and timing of gene induction which is similar to pattern observed in cells stimulated to proliferate in cell culture. Among the genes activated in the early Gi-GI/S period of liver regeneration there are at least four cellular oncogenes: c-fos, c-myc, ~53, C-rus (Goyette et a1.,1983; Goyette et a1.,1984; Makino et al., 1984; Kruijer et a1.,1986; Thompson et a1.,1986), and at 16 h activation of c-myb was reported (Sobczak i Duguet.1986). Other genes activated in regenerating liver include ct- and p-fibrinogen (Sobczak et al., 1987), actin and tubulin (Friedman et a1.,1984).
The aim of this work was to extend our former observation on expression of c-fos, c -rnyc and hsp70 genes in regenerating liver (Biesiada et al.,19871 by studying the activation of other genes. This includes genes coding for ATP/ADP translocase, ornithine decarboxylase (ODC) , vimentin, calcyclin, oncogenes c-Ha-r-1 and ~53 and for two PDGF-inducible genes KC and JE with as yet unknown function (Cochran et al., 1983).
MATER1 AI,!?. AND METHODS
Animals: Male albino rats (Wistar strain; 2-2.5 month old) were from our animal colony. Rats were partially hepatectomized (removal of approximately 70% of the liver mass) by the method of Higgins and Anderson (1931). To deplete liver glycogen, food was withdrawn 14 h before death. Rats were sacrificed between 9 and 11 a.m. Livers were frozen in liquid nitrogen, pulverized and stored at -7O*C. Three to five livers were pooled for each time point. m extraction and hybridization: Total RNA was extracted by the guanidinum isothiocyanate-hot phenol methods (Maniatis et a1.,19821, fractionated on 1% agarose-2.2 M formaldehyde gels and electroblotted onto GeneScreen filters. Prehybridization and hybridization steps were performed at 42OC essentially as described by Wahl et a1.(1979). The hybridization mixture contained 50% formamide, 5x ssc, 5x Denhardt's solution, 20 mM sodium phosphate (ph 6.51, 10% dextran sulphate, 250 pg/ml of denatured salmon sperm DNA and one of the radioactive probes described below. Filters were hybridized for 18 h then washed once with 2x ssc,

Cell Biology International Reports, Vol. 12, No. 6, June 1988 485
0.5% SDS at room temperature and three times with 0.1x ssc. 0.1% SDS at 55OC. Filters were covered with X-ray film and exposed at -7O*C with Kodak intensifying screen. Probes: The following plasmids were used as the hybridization probes: PBC-Nl carrying human c-Ha-z-as-1 gene sequences (gift of Dr. G.Sauer, Pulciani et al., 19821, PJE-3 and PKC-1 carrying mouse JE and KC gene sequences respectively (Cochran et a1.,1983), hp2Fl carrying human ATP/ADP translocase gene sequences (Battini et a1.,1987), p2A9 carrying human calcyclin gene sequences (Calabretta et al., 19861, P JODC42 carrying human ornithine decarboxylase gene sequences (Kaczmarek - unpublished data), pLSV53C-62 carrying human ~53 gene sequences (plasmid orginally obtained from Dr. M.Oren, see Zakut-Houri et al.,19851 (all these probes were a kind gift of Dr.L.Kaczmarekl, and p2AM carrying mouse P53 gene sequences (gift of Dr.P.Chumakov, see Chumakov et a1.,19821. Probes were labelled with a-[PazldATP (Amersham) by nick translatign to specific activity of approximately 10 cpm/pg DNA.
RESULTS AND DISCUSSION Northern blot analysis of RNA derived from
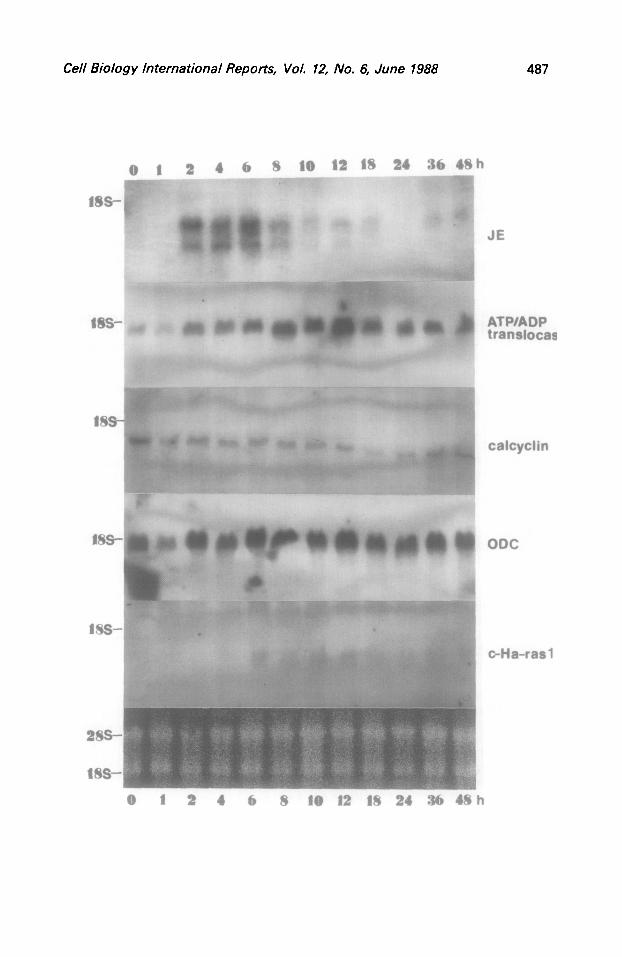
regenerating livers is shown on Fig.1. The most conspicuous observation is the expression pattern of JE gene. Hybridization with the plasmid pJE-3 revealed two RNA species of about 1.0 kb and 1.4 kb in size as it was estimated from their possition relative to that of rRNA. The lower (1.0 kb) band corresponds to the size of JE mRNA described by Cochran et al. (1983) . Both bands appeared at 2 h of regeneration and lasted up to 12 h with conspicious higher intensity at 6 h. At 18, 36 and 48 h of regeneration no hybridization of JE probe with the lower RNA band and at 24 h with both RNA bands was observed. The nature of the larger (" 1.4 kb) species of transcripts is at present unexplicable. RNA isolated from either control liver or 1 h after hepatectomy did not reveal detectable quantities of JE gene hybridizating transcripts. In Balb/c 3T3 cells stimulated by platelet-derived growth factor (PDGF), gene JE was induced 20 to 60-fold wihin l-4 h (Cochran et al ..19831. In contrast to JE gene we observed no detectable hybridization signals in Northern blots with KC, another PDGF-inducible gene (Cochran et al..19831 at any time of liver regeneration (not shown).
One more growth factor-inducible gene we tested was the one coding for the ATP/ADP translocase. Battini et a1.(1987) have shown that this gene is growth-regulated, and is activated when quiescent cells are stimulated

486 Cell Biology International Reports, Vol. 12, No. 6, June 1988
with platelet-derived growth factor or epidermal growth factor (EGF). The lensht of the ATP/ADP translocase gene transcript was reported as 1.5 kb by Battini et a1.(1987), and the RNA band detected by us is of similar size to this estimate (Fig.11. We detected an increase in the level of RNA hybridization with the ATP/ADP translocase gene sequences between 8 and 18 h of liver regeneration (Fis.1). This is roughly at the time of transition of Gi to S phase. and may reflect increased ATP demand at the time of active RNA and DNA synthesis. The drop in the intensity of hybridization signal at first hour after partial hepatectomy may be relevant, however no reasonable explanation of this phenomenon can be offered.
Hybridization of Northern blots with the plazmid pZA9 carrying gene for the calcium-binding protein (calcyclin) revealed a rather low signal intensity in regenerating liver compared to the signal from RNA of normal liver. During the replication of DNA (12-24 h after hepatectomyj the level of calcyclin RNA was still lower. This is in contrast to fibroblasts stimulated to proliferate by serum, PDGF or EGF (but not insulin) where the RNA level complementary to the calcyclin probe was elevated (Calabretta et a1.,1986). The size of calcyclin transcript is 0.6 kb (see also Calabretta et al., 1986).
Ornithine decarboxylase (ODC) is one of the critical enzymes involved in the control of cell growth. During the first few hours after partial hepatectomy the activity of ODC increases UP to 40 fold (Schrock et al., 1970). Translational regulation of ODC - regenerating liver was postulated by Faust0 (1971). Jskl and Htiltts (1974) and P8so (cit. after McConlogue et al., 1986) ; in 3T3 cells by Clark and Fuller (1985),
Fig.1. Levels of JE, ATP/ADP translocase, calcyclin, ornithine decarboxylase and c-Ha-1-as-l gene transcripts in rat liver at various times after partial hepatectomy. Rats were killed at the indicated times after the opration. Equal amounts of total liver RNA (20 pg per lane, equalized for rRNA - lower panel) were loaded onto identical asarose-formaldehyde gels: for each gene probe separate RNA samples have been elect blots were hybridized with the
Syhoresed. Northern P-labelled probes
described in MATERIALS AND METHODS. The 18s and 28s rRNA bands were visualised by staining with ethidium bromide of the same RNA preparation fractionated on the separate gel.
Cell Biology international Reports, Vol. 12, No. 6, June 7988 487
ms-
ms-
1 a 4 6 8 10 la 16 ar 34 481
ATPIADI trrnsloc
h
JE
calcyclil
ODC
*Ha-ras 1
0 4 6 8 10 12 18 24 .% 48 h

Cell Biology International Reports, Vol. 72, No. 6, June 1988
and in mouse S49 cell variants by McConlogue et a1.(19861. This supposition correlates with our finding. Northern blot analysis of RNA from regenerating rat liver probed with ODC gene sequences revealed a cosistent intensity of signals throughout the 2 day period (Fig.11. ODC gene transcript is about 2.0 kb in size (see also McConlogue et a1.,1986).
We have observed no detectable signal when total liver RNA was hybridized with vimentin gene probe (not shown). This gene was shown to be growth-regulated in mouse 3T3 cells and in various mesenchvme-derived cells (Ferrari et a1.,1986). The absence of vimentin gene activity in regenerating liver can be explained by the findings of Franke et al., (1979) that cytoskeletal intermediate filaments in epithelia-derived cells do not contain vimentin-type protein. The lack of vimentin gene transcripts corroborates with the finding of Franke et al., (1979) and also the fact that during the first two days of liver regeneration only hepatocytes are stimulated to divide.
In accordance with earlier reports (Goyette et al . ,1983; Goyette et a1..1984; Makino et al.,19841 we observed that expression of c-Ha-r--l gene (size transcript 1.4-1.6 kb) is extended over a broad period of regeneration but this becomes evident at 6 h (Fig.11.
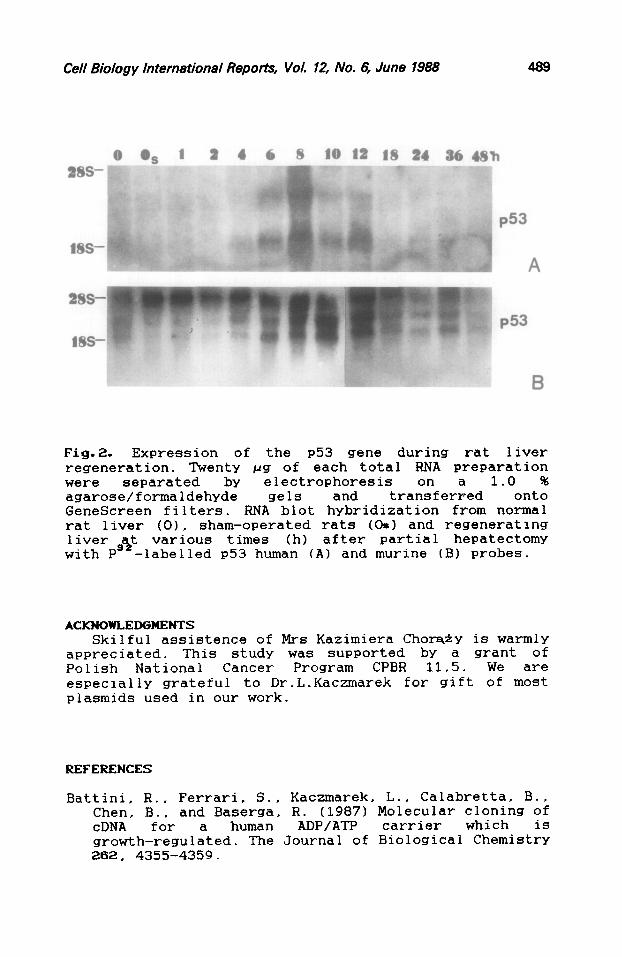
Finally, we confirmed the observation by Thompson et al., (1986) that the P53 gene is expressed at prereplicative stage (in our experiment from 6 to 12 h; Fig.2). Similar expression pattern was obtained both with human ~53 probe (Fig.2A) and with murine ~53 probe (Fig.2.B). Signals have been obtained with three band sizes: 1.8 kb (corresponding to ~53 mENA; estimation by Thompson et a1.,19861, 3.0 kb (unknown RNA species), and the largest of about 5.0 kb probably derived from cross-hybridization with 28s r-RNA.
Our findings increase the number of genes induced in regenerating hepatocytes. In the group of "early genes" most probably corresponding to "competence" genes besides c-fos. c-myc and P53 (Makino et a1.,1984; Goyette et al., 1984; Kruijer et a1.,1986; Thompson et al., 1986; Biesiada et al.19871 two other genes reported here have to be included. These are JE (with yet unknown function) and ATP/ADP translocase. The prompt but transient expression of JE gene after partial hepatectomy suggest its growth-regulatory role.
One may expect that the list of genes that take part in the control of hepatocyte proliferation will be extended.
Cell Biology International Reports, Vol. 12, No. 6, June 1988 499
P53
B
Fig.2. Expression of the ~53 gene during rat liver regeneration. Twenty pg of each total RNA preparation were separated by electrophoresis 1.0 % agarose/formaldehYde gels and tranq3nferFed onto GeneScreen filters. RNA blot hybridization from normal rat liver (01, sham-operated rats (OS) and regenerating liver est various times (h) after partial hepatectomy with P -1abelled ~53 human (A) and murine (B) probes.
ACKNOWLEDGMEXTS Skilful assistence of Mrs Kazimiera Chor&y is warmly
appreciated. This study was supported by a grant of Polish National Cancer Program CPBR 11.5. We are especially grateful to Dr.L.Kaczmarek for gift of most piasmids used in our work.
REFERENCES
Battini, R., Ferrari, S., Kaczmarek, L., Calabretta, B.. Chen, B., and Baserga. R. (1987) Molecular cloning of cDNA for a human ADP/ATP carrier which is growth-regulated. The Journal of Biological Chemistry 262, 4355-4359.

490 Cell Biology International Reports, Vol. 72, No. 6, June 1988
Biesiada. E.. Wi9niewski. J., Krawczyk, Z., and Chora,iy, M.(1987) Expression of c-fos, c-myc and hsp70 genes at early stages of the regeneration of rat liver. Bulletin of the Polish Academy of Sciences, Biological Sciences 35, 165-171.
Bucher. N.L.R., MacGowan, L.A., and Russel, W.E. (1983) Nerve, organ, and tissue regeneration; research perspectives ted. Seil, F.J.1. Academic Press. New York.
Calabretta, B., Battini, R., Kaczmarek. L., de Riel, J.K., and Baserga, R. (1986) Molecular cloning of the cDNA for growth factor-inducible gene with strong homology to S-100, a calcium-binding protein. The Journal of Biological Chemistry 261, 12628-12632.
Chumakov, P.M., Yotzova, W.S., and Georgiev, G.P. (1982) Isolation of recombinant clone containing mRNA sequences for mouse nonviral T-antigen. Proceedings of the Academy of Science USSR 267, 1272-1275.
Clark, J.L., and Fuller J. (1975) Regulation of ornithine decarboxylase in 3T3 cells by putrescine and spermidine: indirect evidence for translational control. Biochemistry 14, 4403-4409.
Cochran, B.H., Reffel, A-C., and Stiles, C.D. (1983) Molecular cloning of gene sequences regulated by platelet-derived growth factor. Cell 33, 939-947.
Denhardt, D.T., Edwards, D.R., and Parfett, C.L.J. (1986) Gene expression during the mammalian cell cycle. Biochimica et Biophysics Acta 865, 83-125.
Fausto, N. (1971) The control of ornithine decarboxylase activity during liver regeneration. Biochimica et Biophysics Acta 238, 116-128.
Ferarri, S., Battini, R., Kaczmarek, L.. Rittling. S.. Calabretta, B., de Riel, J.K., Philiponis. V.. Wei. J-F., and Baserga. R. growth regulation of
(;;:6) Coding sequences and human vimentin gene.
Molecular and Celluiar Biology 6. 3614-3620.
Franke, W.W.. Schmidt, E . , (1978) Different
@born, M., and Weber, K. intermediate-sized
distinguished by filaments
immunofluorescence microscopy. Proceedings of the National Academy of Sciences USA 75, 5034-5038.

Cell Biology International Reports, Vol. 12, No. 6, June 1988 491
Friedman, J.M., Chung, E.Y., and Darnell, J.E. (1984) Gene expression during liver regeneration. Journal of Molecular Biology 178. 37-53.
Goyette, M., Petropoulos, C.J., Shank, P.R., and Fausto, N. (1983) Expression of a cellular oncogene during liver regeneration. Science 218, 510-512.
Goyette, M., Petropoulos, C.J., Shank, P.R., and Fausto,N. (1984) Regulated transcription of c-Ki-rus and c-myc during compensatory growth of rat liver. Mollecular and Cellular Biology 4, 1493-1498.
Higgins, G.M., and Anderson, R.M. (1931) Experimental pathology of the liver. I. Restoration of the liver of the white rat following partial surgical removal. Archives of Pathology 12, 186-202.
JLinne. J., and Hijlttii, E. (1974) Regulation of ornithine decarboxylase activity by putrescine and spermidine in rat liver. Biochemical and Biophysical Research Communications 61, 449-456.
Kaczmarek, L. (1986) Protooncogene expression during the cell cycle. Laboratory Investigation 54. 365-376.
Kruijer, W., Skelly, H., Botteri, F., and van der Putten, H., Barbert, J.R., Verma, I.M., and Leffert, H.L. (1986) Proto-oncogene expression in regenerating liver is stimulated in cultures of primary adult rat hepatocytes. The Journal of Biological Chemistry 261.7929-7933.
Makino, R., Hayashi, K., and Sugimura, T. (1984) c-myc transcript is induced in rat liver at a very early stage of regeneration or by cycloheximide treatment. Nature 310, 697-698.
Maniatis, T., Fritsh. E.F., and Sambrook. J. (eds.) (1982) in: Molecular Cloning. Cold Spring Harbor Laboratory. Cold Spring Harbor, N.Y. PP. 194-195.
McConlogue, L., Dana, S-L.. and Coffiano. P. (1986) Multiple mechanisms are responsible for altered expression of ornithine decarboxylase in overproducing variant cells. Molecular and Cellular Biology 6, 2865-2871.
Pulciani, S., Santos, E., Lauver, V., Long, L.K., and Barbacid, M. (1982) Transforming genes in human tumors. Journal of Cellular Biochemistry 20, 51-61.

492 Cell Biology International Reports, Vol. 12, No. 6, June 1988
Schrock, T.R., Oakman, N.J.. and Buchaer, N.L.R. (1970) Ornithine decarboxylase activity in ralation of growth of rat liver. Bochimica et Biophysics Acta 204. 564-577.
Sobczak, J., and Duguet, M. (1986) Molecular biology of liver regeneration. Biochimie 68, 957-967.
Sobczak, J., Lotti, A-M., Taroux, P., and Duguet, M. (1987) Molecular cloning of mRNA sequences transiently induced during rat liver regeneration. Experimental Cell Research 169, 47-56.
Thompson. N.L., Mead, J.E., Braun, L., G0yette.M. Shank, P.R., and Fausto, N. (1986) Sequential protooncogene expression during rat liver regeneration. Cancer Research 4-6, 3111-3117.
Wahl. G.M., Stern, M., and Stark, G.R. (1979) Efficient transfer of long DNA fragments from agarose gels to diazobenzyloxymethyl-paper and rapid hybridization by using dextran sulfate. Proceedings of the National Academy of Science USA 76, 3683-3687.
Zakut-Houri, Z.. Bienz-Tadmor, B., Givo, D., and Oren, M. (1985) Human ~53 cellular tumor antigen: cDNA sequence and expression in COS cells. The EMBO Journal 4, 1251-1255.
Received: 9.12.87 Accepted: 25.4.88