endothelin-1 signaling promotes fibrosis in vitro in a ... journal of immunology endothelin-1...
TRANSCRIPT

of June 27, 2018.This information is current as
Coagulation CascadeModel by Activating the ExtrinsicVitro in a Bronchopulmonary Dysplasia Endothelin-1 Signaling Promotes Fibrosis In
Matthaios Speletas, George Kolios and Konstantinos RitisRafail, Stergios Vradelis, Ioannis Sigalas, You-Qiang Wu,Kourtzelis, Marianna Skordala, Ioannis Mitroulis, Stavros Konstantinos Kambas, Akrivi Chrysanthopoulou, Ioannis
http://www.jimmunol.org/content/186/11/6568doi: 10.4049/jimmunol.1003756April 2011;
2011; 186:6568-6575; Prepublished online 29J Immunol
MaterialSupplementary
6.DC1http://www.jimmunol.org/content/suppl/2011/04/29/jimmunol.100375
Referenceshttp://www.jimmunol.org/content/186/11/6568.full#ref-list-1
, 11 of which you can access for free at: cites 45 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved.Copyright © 2011 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on June 27, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on June 27, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

The Journal of Immunology
Endothelin-1 Signaling Promotes Fibrosis In Vitro in aBronchopulmonary Dysplasia Model by Activating theExtrinsic Coagulation Cascade
Konstantinos Kambas,*,1 Akrivi Chrysanthopoulou,*,1 Ioannis Kourtzelis,*,†
Marianna Skordala,‡ Ioannis Mitroulis,* Stavros Rafail,† Stergios Vradelis,*
Ioannis Sigalas,‡ You-Qiang Wu,† Matthaios Speletas,x George Kolios,{
and Konstantinos Ritis*
Neonatal respiratory distress syndrome can progress to bronchopulmonary dysplasia (BPD), a serious pulmonary fibrotic disorder.
Given the involvement of the extrinsic coagulation cascade in animal models of lung fibrosis, we examined its role in BPD. We
observed a higher number of neutrophils expressing tissue factor (TF) in bronchoalveolar lavage fluid (BALF) from infants with
BPD than from those with uncomplicated respiratory distress syndrome together with a parallel decrease in TF and connective
tissue growth factor (CTGF) in BALF supernatants during the disease course. The involvement of coagulation in the fibrotic process
associatedwith BPDwas further evaluated by treating primary human colonicmyofibroblasts with BALF supernatants from infants
with BPD. These human colonic myofibroblasts demonstrated an enhanced C5a- and thrombin-dependent migration. Moreover,
they expressed TF in an endothelin-1–dependent manner, with subsequent activation of the extrinsic coagulation cascade and
CTGF production mediated by protease-activator receptor-1 signaling. These data provide a novel mechanism for the develop-
ment of BPD and indicate that endothelin-1 signaling contributes to fibrosis by upregulating a TF/thrombin amplification loop
responsible for CTGF production, and offer novel and specific therapeutic targets for pulmonary fibrotic disease. The Journal of
Immunology, 2011, 186: 6568–6575.
Cross talk between inflammation and the activation ofthe extrinsic coagulation cascade has been described ina number of disease models (1). In addition to activated
endothelium and monocytes, neutrophils have been recentlyshown to express tissue factor (TF) (2–7), the in vivo initiator ofcoagulation. Importantly, the inflammatory mediators C5a (2–4,6–8), TNF-a (5, 6, 9), and IL-1b (10) are all inducers of TF.Furthermore, a recent study has demonstrated a correlation be-tween coagulation and the inflammatory response in patients withacute respiratory distress syndrome (ARDS), indicating that theexpression of TF by neutrophils in the alveolar microenvironmentis C5a- and TNF-a dependent (6).The coagulation system is actively involved in several fibrotic
models (11–15) through the production of thrombin. Moreover,thrombin has been linked to the production of procollagen (11)and the expression of connective tissue growth factor (CTGF),a crucial fibrotic factor, in lung fibroblasts via the protease-activated receptor 1 (PAR-1) (11, 12). One of the main celltypes that contribute to fibrosis via collagen production is myo-fibroblasts (16, 17). Although they originate from the differenti-ation of several cell populations in different tissues, they sharea common function (18), producing profibrotic mediators.In an attempt to further elucidate the interplay between in-
flammation and fibrosis mediated by the extrinsic coagulationsystem, we have used the pulmonary fibrotic disease broncho-pulmonary dysplasia (BPD) as a model. This is a life-threatening,rapidly progressing fibrotic disorder in infants that occurs as acomplication of neonatal respiratory distress syndrome (RDS)(19). BPD is thought to be the result of an excessive inflammatoryresponse (20), and earlier studies have established the significanceof neutrophils in the alveolar microenvironment in this process(21, 22). Furthermore, the levels of several mediators, such as IL-6(23–25), IL-8 (23, 25), TNF-a (25), and endothelin-1 (ET-1) (26),have been found to be increased in tracheal aspirates from infantswith BPD. Especially, ET-1 signaling, which is implicated in the
*First Department of Internal Medicine, Democritus University of Thrace, Alexan-droupolis 68100, Greece; †Department of Pathology and Laboratory Medicine, Uni-versity of Pennsylvania, Philadelphia, PA; ‡Neonatal Intensive Care Unit, GeneralUniversity Hospital of Alexandroupolis, Alexandroupolis 68100 Greece; xDepart-ment of Immunology and Histocompatibility, University of Thessaly, Larissa,Greece; and {Department of Pharmacology, Democritus University of Thrace, Alex-androupolis 68100, Greece
1K.K. and A.C. contributed equally to this work.
Received for publication November 12, 2010. Accepted for publication March 30,2011.
K.K. and A.C. performed the cell culture stimulation studies, real-time PCR, Westernblot experiments, immunocytochemistry, migration assay, and analyzed the data andwrote the manuscript; I.K. performed Western blot experiments and wrote the man-uscript; M.S. and I.S. diagnosed and followed the patients, and provided patients’samples and contributed to the writing of the manuscript; I.M. and S.V. contributed inthe isolation, culturing, and characterization of myofibroblast and had extensivescientific discussion for this study; I.M. also contributed to the writing of the man-uscript; S.R. optimized immunostaining conditions, analyzed data, and designed Fig.6; Y.-Q.W. synthesized and characterized C5a-GFP; M.S. performed immunostainingand ET-1 ELISA; G.K. contributed in the establishment of myofibroblast cultures andin the study design; and K.R. conceived and supervised the project and wrote themanuscript. All authors critically revised the manuscript and approved the finalversion.
Address correspondence and reprint requests to Dr. Konstantinos Ritis, General Uni-versity Hospital of Alexandroupolis, Dragana, Alexandroupolis 68100, Greece.E-mail address: [email protected]
The online version of this article contains supplemental material.
Abbreviations used in this article: ARDS, acute respiratory distress syndrome; ATIII,anti-thrombin III; BALF, bronchoalveolar lavage fluid; BPD, bronchopulmonarydysplasia; C5aR, C5a receptor; CTGF, connective tissue growth factor; ET-1, endo-thelin-1; HCMF, human colonic myofibroblast; PAR-1, protease-activated receptor 1;PMN, polymorphonuclear cell; RDS, respiratory distress syndrome; TF, tissue factor.
Copyright� 2011 by TheAmericanAssociation of Immunologists, Inc. 0022-1767/11/$16.00
www.jimmunol.org/cgi/doi/10.4049/jimmunol.1003756
by guest on June 27, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

pathogenesis of various pulmonary fibrotic models (27, 28), wasidentified as a target for antifibrotic therapeutic interventions(29). However, the possible mechanism linking ET-1 and the in-flammatory response to the development of fibrosis is not yetclear.The relatively rapid progression toward fibrosis during the
course of BPD makes this disease a particularly useful model foranalyzing the link between inflammation and extrinsic coagulation.In this report, we provide evidence that in BPD, the extrinsiccoagulation pathway, together with C5a, promotes the migrationof myofibroblasts to the alveolar microenvironment. These cellsproduce collagen and CTGF via PAR-1 signaling, thereby con-tributing to fibrosis. Moreover, ET-1 induces TF expression inmyofibroblasts, further amplifying the thrombin generation and thesubsequent progression to fibrosis.
Materials and MethodsPatient groups
The study subjects were 12 newborn infants admitted to the NeonatalDepartment and Neonatal Intensive Care Unit of University GeneralHospital of Alexandroupolis, Greece, who required mechanical ventilationat birth because of RDS. They were divided into two groups on the basis ofdisease course: 1) the RDS group consisted of six infants with acute re-spiratory failure and x-ray findings in the lungs consistent with RDS, whosubsequently recovered, were breathing freely, and had a normal chestradiograph on the 28th day of life; and 2) the BPD group consisted of sixinfants with RDS who subsequently experienced development of BPD,according to established clinical criteria (30), and an abnormal chest ra-diograph. The clinical parameters of all 12 patients are described inSupplemental Table I. Moreover, four healthy infants were also recruitedfor the collection of peripheral blood for the isolation of neutrophils. Allexperimental procedures were approved by the Institutional Review Boardand were in compliance with institutional guidelines and the HelsinkiDeclaration. Written consent was provided by the guardians of all infantsin this study.
Bronchoalveolar lavage fluid collection and preparation
Bronchoalveolar lavage fluid (BALF) was collected using a nonbron-choscopic technique, according to the recommendations of the EuropeanRespiratory Society Task Force committee (31). BALF collection wasperformed on day 3, because the highest level of cell migration into thealveolar microenvironment was observed at this point in the disease pro-cess, according to our preliminary data and previous findings (22). In thecase of the BPD infants, further BALF collection was performed on day12, the earliest day on which two of the BPD infants were extubated.BALF was processed as previously described (6). BALF supernatants fromthree patients with either BPD or RDS were pooled for further studies,after assessing the potential of all samples to induce collagen production inmyofibroblast cultures.
Cell preparation and culture
In vitro studies were performed on human colonic myofibroblasts (HCMFs)isolated, as previously described (32), from normal tissue in endoscopicbiopsy samples obtained from four healthy donors at the Academic Hos-pital of Alexandroupolis, Greece. Primary human lung myofibroblastswere also isolated and cultured from a single lung lobectomy sample.Samples were collected in compliance with institutional guidelines.
All experiments were carried out on cells at passage 2 to 3. Myofi-broblasts were characterized by immunocytochemical staining with thefollowing Abs: mouse anti-actin (smooth muscle; Invitrogen, Carlsbad,CA), mouse anti-vimentin (Invitrogen), and mouse anti-desmin mAbs(Invitrogen; Supplemental Fig. 1).
Peripheral polymorphonuclear cells (PMNs) from infants of the twogroups were isolated as previously described (6).
Stimulation and inhibition studies in cultured cells
Myofibroblasts were seeded into six-well plates (�0.5 3 106 cells/well;Corning) or in four-well chamber slides (�0.53 105 cells/well; Nunc Lab-Tek II Chamber Slide System; Sigma-Aldrich, St. Louis, MO) and culturedfor 7–10 d in complete DMEM (Life Technologies BRL, Carlsbad,CA). Cells were cultured in serum-free medium for 24 h before use instimulation/inhibition studies.
The majority of in vitro stimulation and inhibition studies were per-formed in HCMFs because of abundance of cells, whereas some key ex-perimental procedures were further reproduced in lung myofibroblasts.Myofibroblasts were incubated with 40 ml pooled BALF supernatant at37˚C in serum-free DMEM medium (Life Technologies BRL) in thepresence of various inhibitors or their respective controls. More specifi-cally, a neutralizing TF mAb (10 mg/ml; American Diagnostica, Green-wich, CT), monoclonal mouse IgG1 (DAKO, Denmark), neutralizingCTGF mAb (10 mg/ml; Santa Cruz Biotechnologies, Santa Cruz, CA), oranti-thrombin III ([ATIII] 0.5 U/ml; Kybernin ZLB Behring, Germany)was applied to inhibit crucial components of the extrinsic coagulationcascade. The latency-associated peptide mAb (TGF-b1, 1 mg/ml; R&DSystems, Minneapolis, MN) was used to inhibit TGF-b1 signaling inBALF supernatants. To block PAR-1, we pretreated HCMFs for 30 minwith the FLLRN PAR-1–specific antagonist peptide (500 mM final con-centration; Anaspec, Fremont, CA). HCMFs were treated with ET-1 pep-tide (70 pM final concentration; Sigma-Aldrich). TNF-a (Sigma-Aldrich)was used in a concentration of 5 ng/ml, whereas blocking of TNF-a inBALF was performed with recombinant anti-human TNF-a Ab(HUMIRA, 0.2 mg/ml; Abbott Laboratories, North Chicago, IL). ET-1receptors A and B were blocked with bosentan (10 mM; Actelion, Swit-zerland). Recombinant human C5a (10 nm) and C5a receptor a (C5aRa;5 mM) (PMX-53) were also applied. Pooled blood serum (2 ml) from adulthealthy donors (hereafter referred to as normal serum) was used in cellcultures requiring coagulation factors for thrombin generation. The finalvolume was adjusted to 2 ml in the assay using six-well plates and 300 mlin the assay that was carried out in four-well chamber slides. To isolatemRNA and protein samples, we harvested cells 3 and 4 h after incubation,respectively, because optimization experiments had yielded the highestexpression at these time points. All the reagents used in the study wereendotoxin-free (Limulus amebocyte assay; Sigma-Aldrich).
Relative quantification of TF and CTGF mRNA synthesis
Isolation of total RNA and cDNA synthesis were conducted as previouslydescribed (6). Real-time RT-PCR for the full-length TF isoform (referredhereafter as TF), CTGF, PAR-1, and C5aR1 (C5a receptor 1) was per-formed under the conditions described in Supplemental Table II. TheGAPDH housekeeping gene was used to normalize the expression levels oftarget genes, and the 22DDCt method (33) was applied to quantify theirrelative expression levels.
Western blot analysis
For TF detection, Western blotting was performed as previously described(6). To detect CTGF production, we used a polyclonal goat anti-human Ab(Santa Cruz Biotechnologies) at a 1/500 dilution, with incubation over-night at 4˚C. A CTGF blocking peptide was used (1.5 mg/ml; Santa CruzBiotechnologies) to ensure the specificity of Ab binding. For Westernblotting, protein precipitates from identical volumes of BALF supernatantwere used, and equal amounts of protein were loaded onto the gel for eachsample to obtain comparable results in the absence of a reference protein.In addition, TF and CTGF were blotted in parallel to eliminate any loadingerrors. Integrated OD analysis of the scanned films was performed usingGel Pro 3.1.
TF and CTGF immunostaining
Immunocytochemical staining was performed using a LSAB + System-APkit (DAKO), according to the manufacturer’s instructions. TF and CTGFwere detected using a streptavidin/biotin method, with an IgG1 mouseanti-human TF mAb (1/100 dilution; American Diagnostica) or IgG goatanti-human polyclonal CTGF Ab (1/100 dilution; Santa Cruz Biotech-nologies). An IgG1 anti-CD19 mAb (DAKO) was used as a negativecontrol.
ET-1 immunoreactivity assays
ET-1 levels in BALF supernatants from infants were determined using anELISA kit (R&D Systems), according to the manufacturer’s instructions.ET-1 was measured in three BALF supernatants from either BPD or RDSinfants; the supernatants were then pooled and used in stimulation studies.The detection range of the assay was ,1–200 pg/ml.
Collagen assays
The soluble collagen types released into culture medium by treated myo-fibroblasts (I–V) were measured using the Sircol Collagen Assay kit(Biocolor, Belfast, U.K.), according to the manufacturer’s instructions andas previously described (34). Collagen measurement was performed inboth cell lysates (100 mg total protein) and culture supernatants (after
The Journal of Immunology 6569
by guest on June 27, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

concentration of the collagen using the manufacturer’s isolation andconcentration reagent). Collagen production was measured in culturesupernatants and myofibroblast lysates after 24 and 12 h of stimulation,respectively, the time points at which the highest collagen production wasobserved. Our analysis produced similar results for culture supernatantsand cell lysates, and consequently only results from culture supernatantsare presented in this article.
Fluorescence microscopy
To detect the presence of C5aRs on HCMFs, we cultured cells on chamberslides and incubated them with 50 nM recombinant human C5a-GFP for 30min at 37˚C. To verify the specificity of the C5a-GFP binding, we pre-treated cells with a highly specific antagonist for the C5aR, C5aRa (PMX-53), or its inactive control (both at 5 mM, for 30 min at 37˚C before in-cubation with C5a-GFP). HCMFs were also stained with a PE-conjugatedanti-C5aR1 mAb (BD Pharmingen) in the presence of C5aRa or its in-active control. PE-conjugated mouse IgG1 was used as the control Ab.Cells were subsequently fixed in 4% PFA and visualized by reverse fluo-rescence microscopy (Axiovert 25, filter set 09, 488009; Zeiss).
HCMF migration assays
To assess the migration of myofibroblasts, an in vitro wound healing assaywas carried out according to the manufacturer’s instructions (Cell Biolabs,San Diego, CA). Concentrations of reagents were identical to the ones usedin the stimulation and inhibition studies section. Cells were visualized byreverse light microscopy (Axiovert 25; Zeiss). The migration rate wasmeasured according to the manufacturer’s instructions.
Statistical analysis
Data are presented as means 6 SD. Because of the limited sample size,a Mann–Whitney nonparametric test (unpaired, for six or more samples) ora Student t test (for fewer than six samples) was used to compare thedifferences in means. The level of significance was set to p # 0.05. Sta-tistical analysis was performed using the SPSS 8.0 statistical program forWindows.
ResultsTF and CTGF coexpression in BALF supernatants from infantswith BPD
To assess the inflammatory status of infants with BPD, we ex-amined the cell composition in BALF obtained at selected timepoints during mechanical ventilation (Supplemental Table III).PMNs were the main population present in the lavage from infantswith BPD (80.86 8.5%); in contrast, the percentage of PMNs waslower (47.8 6 6.0%) in the BALF from infants with RDS. Giventhe fact that inflammation and coagulation can be tightly cor-related in various diseases and our recent description of TF up-regulation in BALF neutrophils from patients with ARDS (6),an analogous condition in adults, we used immunostaining toexamine the expression of TF in BALF cells from infants. Neu-trophils from BPD BALF showed TF expression (Fig. 1AI),whereas those from infants with uncomplicated RDS did not (Fig.1AII). Most nonneutrophil BALF cells (e.g., atypical pneumo-nocytes, macrophages) from either group stained negative for TF(data not shown). The results of our real-time RT-PCR (Fig. 1B,bars 2 and 1, respectively) and Western blot analyses (Fig. 1Ca,lanes I and II, respectively) were consistent with the immuno-staining findings.The fibrotic nature of BPD and the previously reported corre-
lation between exogenous coagulation cascade and fibrosis (11, 12)led us to examine the levels of CTGF in BALF supernatants. Theprotein concentration of CTGF was higher in the BALF super-natants from the BPD group (Fig. 1Cb, lane II) than from the RDSgroup (Fig. 1Cb, lane I). TF protein levels were also higher inBALF supernatants from the BPD group (Fig. 1Cb, lanes I and II,respectively). TF expression in BALF cells decreased on the 12thday of life in three infants who experienced development of BPD(Fig. 1D), whereas both TF and CTGF in supernatants also de-creased over time (Fig. 1E), suggesting a linkage between them.
Moreover, BALF cells from BPD infants exhibited increased TFmRNA levels when compared with peripheral PMNs from the twoinfant populations (Supplemental Fig. 2), suggesting that its up-regulation is a localized pulmonary phenomenon.
C5a and thrombin induce HCMF migration into the BPDenvironment
Myofibroblasts are found in fibrotic lesions from BPD infants onthe fourth day of the disease course (35). Given that these cells arehighly involved in fibrosis, we asked how the possible interplaybetween the TF pathway and the fibrotic nature of CTGF in thesecells might contribute to the pathogenesis of BPD.To examine their ability to proliferate and migrate toward the
alveolar environment, we stimulated HCMFs with BALF super-natant from both infant groups. An increased proliferation andmigration rate was observed after incubation with BPD BALF (Fig.2AIII, 2B, bar III), as compared with BALF from infants withRDS (Fig. 2AII, 2B, bar II). Moreover, HCMFs treated with BPDBALF indicated increased a-smooth muscle actin levels, as dem-onstrated by immunostaining (Supplemental Fig. 1C).The previously described effect of thrombin on myofibroblast
migration (17) and its presence in BALF (36) prompted us toassess the effect of ATIII on the HCMF migration induced by BPDBALF. The migration rate was significantly reduced (Fig. 2AIV,2B, bar IV) after exposure to ATIII, indicating that the extrinsiccoagulation cascade plays a key role in this phenomenon.
FIGURE 1. TF and CTGF expression in BALF from infants with BPD
and RDS. A, TF immunostaining in BALF cells obtained from infants with
BPD (I) or RDS (II) (n = 3 for each group). An anti-CD19 Ab was used as
a negative control (III). Original magnification 31000. A single repre-
sentative experiment is shown. B, Relative expression of TF in BALF-
derived cells from infants with BPD (bar 2: TF, 15.996 0.871; *p, 0.01)
as compared with those with RDS (bar 1) during the third day of disease
manifestation. All experiments were performed in triplicate. Data are
presented as mean relative expression (fold increase) 6 SD of DCT values
using the 22DDCt equation (n = 6 for each group). C, Western blot analysis
of TF and CTGF protein levels in (a) BALF cell lysates and (b) BALF
supernatants from infants with RDS (lane I) or BPD infants (lane II) on the
third day of disease (n = 3). One representative experiment of three in-
dependent experiments is shown. D, Kinetic study of the relative quanti-
tation of TF mRNA in BALF cells obtained from infants with BPD (n = 3)
during the 3rd and 12th day after the onset of the disease. Expression levels
are shown as fold expression compared with the values for infants with
RDS on the third day. Squares, triangles, and circles represent the three
different infants used in this analysis. E, TF and CTGF protein levels in
BALF supernatants from infants with BPD, as detected by immunoblotting
on the 3rd (lane a) and 12th (lane b) day.
6570 ENDOTHELIN-1 AND TISSUE FACTOR IN FIBROSIS
by guest on June 27, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

In an attempt to identify other migratory stimuli, we also in-vestigated the effect of C5a anaphylatoxin, a strong chemo-attractant (37) that is involved in the development of ARDS inadults (6). We initially demonstrated the expression of C5aR onthe surface of HCMFs by detecting the binding of GFP-labeledrecombinant human C5a (Supplemental Fig. 3B); this observationwas verified by assessing mRNA and protein levels of C5aR(Supplemental Fig. 3A, 3C). We observed that C5a caused anincrease in the migration rate of HCMFs in the BPD environment,and this property was abolished by the addition of a C5aR an-tagonist (Fig. 2AV, 2B, bar V). The induction of migration inHCMFs treated with pure thrombin (Fig. 2AIX, 2B, bar IX) orrecombinant human C5a (Fig. 2AVIII, 2B, bar VIII) further sup-ported these findings. Moreover, thrombin and recombinant C5ahad a synergistic effect on the rate of HCMF migration (Fig. 2AX,2B, bar X).
Myofibroblasts produce CTGF in the BPD microenvironmentby activating the extrinsic coagulation pathway
Because myofibroblasts migrate and express CTGF (Fig. 3A, graybar 3, 3B, 3CIII, 3D, bar 1) in response to BPD BALF, wesearched for components of the BALF that could initiate this fi-brotic process. Given the reported involvement of the extrinsiccoagulation cascade in pulmonary fibrotic models (13–15), weincubated HCMFs with BPD BALF after treatment with eitherATIII or a neutralizing mAb against TF. The CTGF gene over-expression was abolished by treatment with ATIII (Fig. 3A, gray
bar 4, 3BIV, 3CVI, 3D, bar 4) and was partially inhibited by anti-TF Ab (Fig. 3A, gray bar 5, 3CVII, 3D, bar 5). These findingswere further observed in lung myofibroblast stimulations (Sup-plemental Fig. 4A, 4B). Moreover, a marked increase in collagenproduction was observed in culture supernatants from HCMFsstimulated with BPD BALF (Fig. 3E, bar 3), compared with cellstreated with RDS BALF (Fig. 3E, bar 2). Similar results were alsoobtained from lung myofibroblast cultures (Supplemental Fig.4C). Treatment with ATIII inhibited collagen production (Fig.3E, bar 4), whereas the anti-TF mAb had only a partial inhibi-tory effect (Fig. 3E, bar 5). Moreover, neutralization with CTGFpolyclonal Ab attenuated the production of collagen, addressingthe involvement of CTGF in collagen production in myofibro-blasts (data not shown). Blocking of the fibrosis-related factorTGF-b1 with neutralizing mAb in BALF supernatant from BPDinfants failed to abolish CTGF expression and collagen production(data not shown).To further verify the involvement of the extrinsic coagulation
cascade in the upregulation of CTGF in HCMFs, we blocked PAR-1 in cells before treating them with BALF supernatant from infantswith BPD. Pretreatment with the PAR-1 antagonist resulted in
FIGURE 2. HCMFs express C5aRs and migrate in response to C5a/
thrombin activity. C5a- and thrombin-dependent migration (A) and mi-
gration rate (B) of HCMFs in the BPD environment. In vitro migration, as
assessed by wound healing assay, of HCMFs treated with culture medium
(I); BALF supernatant from infants with RDS (II) or BPD infants (III);
BPD BALF supernatants preincubated with ATIII (0.5 U/ml) (IV); BPD
BALF supernatants after preincubation of cells with C5aRa (5 mM) (V) or
bosentan (10 mM) (VI); BPD BALF supernatants after simultaneous in-
hibition of C5aRs, ET-1 receptors, and thrombin (VII); C5a alone (10 nM)
(VIII); thrombin alone (5 U/ml) (IX); or simultaneous stimulation with C5a
and thrombin (X). Duplicate wells were prepared for each sample. Original
magnification 350. One representative experiment of three independent
experiments is shown in A. B, Data are presented as means 6 SD (n = 3).
FIGURE 3. CTGF expression and collagen levels after in vitro stimu-
lation of HCMF. A, Relative expression of CTGF and PAR-1 after in vitro
stimulation of HCMFs with BALF supernatant from infants with RDS or
BPD in the presence of extrinsic coagulation pathway inhibitors. HCMFs
were incubated with culture medium (bar 1), BALF supernatant obtained
from infants with RDS (bar 2) or BPD (bar 3), BALF supernatant from
BPD infants after pretreatment with ATIII (bar 4), the neutralizing anti-TF
Ab (bar 5), or PAR-1 antagonist (bar 6). Data are presented as mean
relative expression 6 SD (n = 4; *p , 0.05). B, CTGF expression, as
assessed by immunostaining (original magnification 3400). HCMFs were
treated with culture medium (I), BALF supernatant from infants with RDS
(II) or BPD (III), and BPD BALF supernatant pretreated with ATIII (IV). C
and D, CTGF protein expression on HCMFs, as detected by Western blot
analysis (D, CTGF/GAPDH ratio as obtained from integrated OD mea-
surement). HCMFs were treated with BALF supernatant from infants with
BPD (III, bar 1) or RDS (IV, bar 2), culture medium (V, bar 3), BPD BALF
supernatant pretreated with ATIII (VI, bar 4), or neutralizing anti-TF Ab
(VII, bar 5). One representative out of three independent experiments is
shown (B, C). Data are presented as means 6 SD (D). E, Detection of
the collagen content in supernatants after in vitro stimulation of HCMFs
treated with culture medium (bar 1), BALF supernatant from either RDS
(bar 2) or BPD (bar 3) infants, BPD BALF supernatant pretreated with
ATIII (bar 4) or neutralizing anti-TF Ab (bar 5), or BPD BALF super-
natant after pretreatment of cells with PAR-1 antagonist (bar 6). Data are
presented as means 6 SD (n = 3; *p , 0.05).
The Journal of Immunology 6571
by guest on June 27, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
a significant decrease in both CTGF mRNA levels in HCMFs (Fig.3A, gray bar 6) and collagen levels in culture supernatants (Fig.3E, bar 6). Furthermore, PAR-1 mRNA expression levels weresignificantly increased in HCMFs incubated with BALF frominfants with BPD (Fig. 3A, black bar 3), as compared with cellsstimulated with BALF from infants with RDS (Fig. 3A, black bars1 and 2, respectively). This effect was abolished by ATIII (Fig. 3A,gray bar 4) or the PAR-1 antagonist (Fig. 3A, black bar 6) andpartially inhibited by anti-TF treatment (Fig. 3A, black bar 5).
The BPD alveolar microenvironment induces TF expression inmyofibroblasts
The partial inhibitory effect of anti-TF Ab on CTGF and collagenproduction by HCMFs suggested a potential production of TF bythese cells. Indeed, HCMFs stimulated with BALF supernatantsfrom infants with BPD exhibited a significant increase in TFmRNA transcripts (Fig. 4A, bar 2) when compared with HCMFsstimulated with BALF supernatants from the RDS group (Fig. 4A,bar 3). This finding was further corroborated on the protein levelby Western blotting (Fig. 4B, lane II) and immunocytochemicalstaining (Fig. 4CII). TF immunostaining also verified these find-ings in lung myofibroblasts (Supplemental Fig. 4D). The pro-duction of TF by myofibroblasts suggests the existence of anamplification loop, which produces further activation of the TFpathway and subsequent CTGF induction.
ET-1 signaling mediates the induction of TF in myofibroblasts
To identify mediators that might be responsible for the upregula-tion of TF expression in myofibroblasts, we investigated the role ofC5a and TNF-a, well-established TF inducers that have previouslybeen described in analogous pulmonary models (ARDS) (6).Treatment of HCMFs with C5a or a neutralizing anti–TNF-a Abin BPD BALF failed to affect the TF expression levels (data notshown).Furthermore, because ET-1 and the extrinsic coagulation cascade
are involved in the fibrotic process and given that HCMFs express
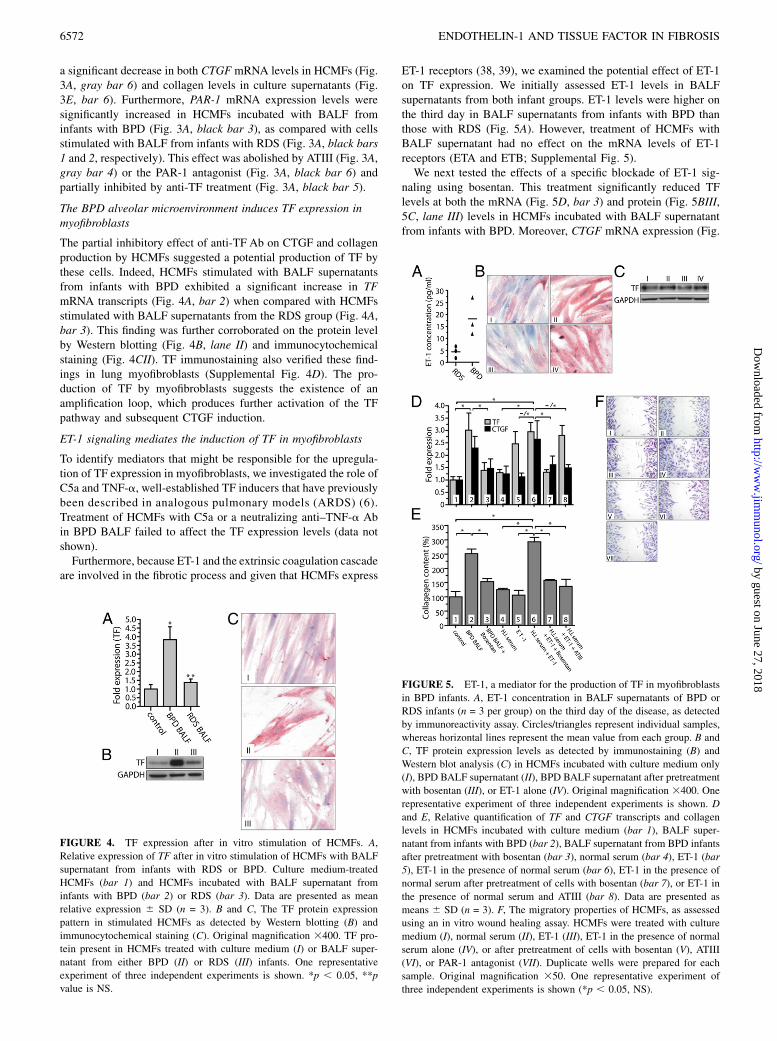
ET-1 receptors (38, 39), we examined the potential effect of ET-1on TF expression. We initially assessed ET-1 levels in BALFsupernatants from both infant groups. ET-1 levels were higher onthe third day in BALF supernatants from infants with BPD thanthose with RDS (Fig. 5A). However, treatment of HCMFs withBALF supernatant had no effect on the mRNA levels of ET-1receptors (ETA and ETB; Supplemental Fig. 5).We next tested the effects of a specific blockade of ET-1 sig-
naling using bosentan. This treatment significantly reduced TFlevels at both the mRNA (Fig. 5D, bar 3) and protein (Fig. 5BIII,5C, lane III) levels in HCMFs incubated with BALF supernatantfrom infants with BPD. Moreover, CTGF mRNA expression (Fig.
FIGURE 4. TF expression after in vitro stimulation of HCMFs. A,
Relative expression of TF after in vitro stimulation of HCMFs with BALF
supernatant from infants with RDS or BPD. Culture medium-treated
HCMFs (bar 1) and HCMFs incubated with BALF supernatant from
infants with BPD (bar 2) or RDS (bar 3). Data are presented as mean
relative expression 6 SD (n = 3). B and C, The TF protein expression
pattern in stimulated HCMFs as detected by Western blotting (B) and
immunocytochemical staining (C). Original magnification 3400. TF pro-
tein present in HCMFs treated with culture medium (I) or BALF super-
natant from either BPD (II) or RDS (III) infants. One representative
experiment of three independent experiments is shown. *p , 0.05, **p
value is NS.
FIGURE 5. ET-1, a mediator for the production of TF in myofibroblasts
in BPD infants. A, ET-1 concentration in BALF supernatants of BPD or
RDS infants (n = 3 per group) on the third day of the disease, as detected
by immunoreactivity assay. Circles/triangles represent individual samples,
whereas horizontal lines represent the mean value from each group. B and
C, TF protein expression levels as detected by immunostaining (B) and
Western blot analysis (C) in HCMFs incubated with culture medium only
(I), BPD BALF supernatant (II), BPD BALF supernatant after pretreatment
with bosentan (III), or ET-1 alone (IV). Original magnification 3400. One
representative experiment of three independent experiments is shown. D
and E, Relative quantification of TF and CTGF transcripts and collagen
levels in HCMFs incubated with culture medium (bar 1), BALF super-
natant from infants with BPD (bar 2), BALF supernatant from BPD infants
after pretreatment with bosentan (bar 3), normal serum (bar 4), ET-1 (bar
5), ET-1 in the presence of normal serum (bar 6), ET-1 in the presence of
normal serum after pretreatment of cells with bosentan (bar 7), or ET-1 in
the presence of normal serum and ATIII (bar 8). Data are presented as
means 6 SD (n = 3). F, The migratory properties of HCMFs, as assessed
using an in vitro wound healing assay. HCMFs were treated with culture
medium (I), normal serum (II), ET-1 (III), ET-1 in the presence of normal
serum alone (IV), or after pretreatment of cells with bosentan (V), ATIII
(VI), or PAR-1 antagonist (VII). Duplicate wells were prepared for each
sample. Original magnification 350. One representative experiment of
three independent experiments is shown (*p , 0.05, NS).
6572 ENDOTHELIN-1 AND TISSUE FACTOR IN FIBROSIS
by guest on June 27, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

5D, bar 3), collagen deposition (Fig. 5E, bar 3), and the migrationrate (Fig. 2AVI, 2B, bar VI) of these cells were also significantlyreduced after treatment with bosentan. Inhibition of ET-1 signal-ing also attenuated TF induction and collagen production in lungmyofibroblasts stimulated by BPD BALF (Supplemental Fig.4DIV). This effect of ET-1 receptor blockade could be attributed toan inhibition of the additional TF production in HCMFs and thesubsequent generation of thrombin.We further exposed HCMFs to a specific ET-1 peptide, which
resulted in an upregulation of TF protein (Fig. 5BIV, 5C, lane IV)and mRNA (Fig. 5D, bars 5 and 6) expression levels.Notably, in the presence of serum from healthy individuals
(normal serum), the ET-1 peptide induced HCMF migration (Fig.5FIV), CTGF expression (Fig. 5D, bar 6), and collagen deposition(Fig. 5E, bar 6); no such increases occurred in the absence ofserum (Fig. 5D, 5E, bar 5, 5FIII). Moreover, blocking withbosentan or pretreatment with ATIII abolished the cell migration(Fig. 5FV and 5FVI, respectively), CTGF induction (Fig. 5D, bars7 and 8, respectively), and collagen production (Fig. 5E, bars 7and 8, respectively) in HCMFs incubated with ET-1 and normalserum. Migration of HCMFs was also inhibited by the PAR-1antagonist (Fig. 5FVII), further supporting the involvement ofTF/thrombin axis in this process. Taken together, these findingssuggest a novel role for ET-1 in the fibrotic model of BPD bymediating the induction of TF in myofibroblasts (Fig. 6).
DiscussionThis study was designed to provide novel insight into the de-velopment of fibrosis in BPD patients as a result of cross talkbetween inflammation and coagulation. Our experimental datahave indicated that the inflammatory pulmonary environment inBPD patients contributes to the development of fibrosis by acti-vating the extrinsic coagulation cascade through ET-1 signaling.We have demonstrated that C5a and thrombin can promotemyofibroblast migration, and ET-1 signaling induces TF expressionin these cells. As a result, an amplification loop is generated thataugments the initial triggering of the extrinsic coagulation cascade,further activating myofibroblasts to produce CTGF and collagen.This course of events suggests that ET-1 plays a key role in theprogression to fibrosis.Other studies have reported an increased accumulation of PMNs
in the alveolar space (21, 22) and noted a correlation between thenumber of neutrophils present in the alveoli of preterm infants andthe development of BPD (21, 22). A role for these cells in theacute inflammatory response and tissue injury has also beendocumented (40). Similarly, neutrophils have been shown to bea significant source of TF expression in BALF from ARDS pa-tients (6). This study has demonstrated that neutrophils frominfants with BPD are the prevalent cell population in BALF, andthey contribute, at least in part, to the progression of the disorderby eliciting TF expression and subsequent thrombin generation.The increased expression of TF in the BPD BALF PMNs as ob-served by immunostaining and the similar TF mRNA levels inperipheral PMNs from control infants and infants with either RDSor BPD indicates a localized phenomenon in the alveolar micro-environment. However, we cannot overlook the possible partici-pation of microparticle-derived TF (1) produced by other cells andacquired by PMNs in the TF levels observed in PMNs from BPDBALF.The extrinsic coagulation cascade is able to induce CTGF ex-
pression and collagen deposition via PAR-1 signaling in nonhumanpulmonary models, either through thrombin generation (11, 12) orthrough other proteases of the system (26). CTGF has been im-
plicated in the pathogenesis of IPF through the activation offibroblasts and alveolar epithelial type II cells (41), and it alsoplays a major role in bleomycin-induced lung inflammation innonhuman models (12). Nevertheless, evidence for the invol-vement of CTGF in other human fibrotic respiratory syndromessuch as BPD is still scarce. The higher levels of TF and CTGF insupernatants collected on the third day of the disease provideevidence for a correlation between these two molecules. Sucha finding may be related to the previously reported accumulationof PMNs in the alveolar space and myofibroblasts in the fibroticlesions of BPD (35). Despite the variability of TF expression inBPD BALF cells at day 3 (which could be attributed to the dif-ferential expression of inflammatory mediators in the alveolarmicroenvironment), we found a parallel reduction of TF levels andneutrophil numbers in BALF during the course of the disease. Thisfinding is in accordance with previous studies that have indicateda decrease in thrombin concentration during the course of thedisease (36).The critical role of myofibroblasts in the fibrotic process (42) and
their previously documented presence in BPD lesions in infants(35) prompted us to select this cell type for our in vitro studies.Given that these cells share common functions, even when derivedfrom different tissues (18), we chose to perform the majority ofour in vitro studies in primary cultures of myofibroblasts fromhuman colon because of their availability, and to reproduce thekey findings in primary lung myofibroblasts because of the rarityof sample.Our assessment of the ability of BALF supernatants from infants
with BPD or RDS to induce migration, CTGF production, andcollagen deposition in HCMFs indicated that the myofibroblastsexhibit significantly higher migration in the presence of BALFsupernatant from BPD infants, and this migration is thrombin andC5a dependent. The involvement of complement was furtherverified by showing the functional presence of C5aR in these cells.The chemoattractant activity of thrombin has recently been re-ported (17). We suggest that the initial thrombin generation inBALF occurs as a result of the increased vascular permeabilitycaused by inflammation and TF expression mediated by neu-trophils and other alveolar cells, and that it constitutes a possibleearly chemoattractant stimulus for fibroblasts and myofibroblasts.
FIGURE 6. Proposed mechanism of fibrosis in BPD infants. After lung
injury (1), the inflammatory alveolar environment causes TF expression in
BALF cells and triggers the extrinsic coagulation cascade, resulting in
thrombin generation. 2, C5a and initial thrombin promote myofibroblast
migration. 3, ET-1 induces the expression of TF by the attracted myofi-
broblasts, creating an amplification loop for thrombin that induces further
CTGF production, through PAR-1, and eventually collagen deposition.
The Journal of Immunology 6573
by guest on June 27, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

Thrombin then activates the attracted cells to proliferate andproduce collagen.We found that in addition to their migratory capacity, myofi-
broblasts also express increased CTGF levels and produce collagenin the BPD environment, and that expression is mediated by the TF/thrombin/PAR-1 axis. This observation was consistent with ourhypothesis that the activation of the extrinsic coagulation cascadecontributes to the progression to fibrosis. The incubation of myo-fibroblasts with BALF supernatant from BPD infants also re-sulted in the production of significant amounts of TF. This novelfinding suggests that these cells participate in the activation ofthe extrinsic coagulation pathway, which may lead to excessthrombin generation. Signaling via PAR-1 further induced CTGFproduction and collagen deposition. To identify the key mediator(s)that stimulate myofibroblasts to produce TF in BALF from infantswith BPD, we examined candidate molecules that are known toinduce TF expression in various cell populations (29). Stimulationand inhibition studies failed to implicate C5a or TNF-a in theTF-mediated generation of thrombin.We also examined ET-1 because of its involvement in fibrotic
disorders and BPD. To our knowledge, our results demonstrate forthe first time that inhibition of both ET-1 receptors with bosentanin myofibroblasts abolished the BALF-dependent expression ofTF and subsequent cell migration, as well as CTGF and collagenproduction. Furthermore, the ET-1 peptide was able to induce TFproduction. Moreover, it induced the production of CTGF andcollagen, after adding normal serum, which provides coagulationfactors, thus supporting the involvement of thrombin in the pro-cess. The involvement of ET-1 in the fibrotic response of infantswith BPD was further supported by the presence of increasedamounts of the protein in their BALF. Our findings and the pre-viously reported ability of ET-1 signaling to induce the differen-tiation of lung fibroblasts into myofibroblasts suggest a role forET-1 in the fibrotic process of BPD (43). These data providefurther support for the importance of neutrophils in the alveolarspace in the development of this syndrome, because these cellsproduce endothelin-converting enzyme, which allows them toconvert big endothelin into the signaling ET-1 (44, 45).Based on our findings, we have proposed a possible patho-
physiological mechanism for BPD (Fig. 6). In this model, theinflammation-driven chemoattractant activity in the alveolar en-vironment (e.g., C5a- and IL-8–rich), in conjunction with endo-thelial damage to the pulmonary microvasculature, cause peri-pheral neutrophils to migrate into the alveolar space. It also in-duces the expression of TF, resulting in thrombin generation, andother mediators that further enhance the inflammatory response(Fig. 6, section 1). C5a and the initial thrombin generation pro-mote myofibroblast migration into the mesenchymal space (Fig.6, section 2). Leukocytes convert big endothelin into ET-1, leadingto further activation of the myofibroblasts via ET-1 signaling andthe subsequent production of TF (Fig. 6, section 3). This processresults in additional thrombin generation, initiating a vicious cycle(Fig. 6, section 3). TF/thrombin pathway proteases then stimulatemyofibroblasts through PAR-1 signaling to produce CTGF, whichinduces collagen production via an autocrine mechanism. Thisprocess could apply equally well to other human fibrotic disordersin which inflammation and coagulation coexist.In conclusion, this human ex vivo model demonstrates the in-
volvement of the extrinsic coagulation cascade in the developmentof fibrosis. Our study indicates a novel role for ET-1 in fibrosis andsuggests a mechanism for the amplification of thrombin generation.The development of molecules able to neutralize the mediatorsresponsible for this process may form the basis for novel ap-proaches for therapeutic intervention in fibrotic disorders.
AcknowledgmentsWe thank Marc Iglarz (Actelion, Allschwil, Switzerland) for providing
Bosentan. We also thank Victoria Tsironidou for technical assistance and
Dr. Deborah McClellan for help in editing the manuscript. Finally, we ac-
knowledge the contribution of John D. Lambris for the provision of vital
reagents and the revision of the manuscript.
DisclosuresThe authors have no financial conflicts of interest.
References1. Mackman, N. 2009. The many faces of tissue factor. J. Thromb. Haemost. 7
(Suppl 1): 136–139.2. Ritis, K., M. Doumas, D. Mastellos, A. Micheli, S. Giaglis, P. Magotti, S. Rafail,
G. Kartalis, P. Sideras, and J. D. Lambris. 2006. A novel C5a receptor-tissuefactor cross-talk in neutrophils links innate immunity to coagulation pathways. J.Immunol. 177: 4794–4802.
3. Maugeri, N., M. Brambilla, M. Camera, A. Carbone, E. Tremoli, M. B. Donati,G. de Gaetano, and C. Cerletti. 2006. Human polymorphonuclear leukocytesproduce and express functional tissue factor upon stimulation. J. Thromb.Haemost. 4: 1323–1330.
4. Redecha, P., R. Tilley, M. Tencati, J. E. Salmon, D. Kirchhofer, N. Mackman,and G. Girardi. 2007. Tissue factor: a link between C5a and neutrophil activationin antiphospholipid antibody induced fetal injury. Blood 110: 2423–2431.
5. Rafail, S., K. Ritis, K. Schaefer, I. Kourtzelis, M. Speletas, M. Doumas,S. Giaglis, K. Kambas, S. Konstantinides, and G. Kartalis. 2008. Leptin inducesthe expression of functional tissue factor in human neutrophils and peripheralblood mononuclear cells through JAK2-dependent mechanisms and TNFalphainvolvement. Thromb. Res. 122: 366–375.
6. Kambas, K., M. M. Markiewski, I. A. Pneumatikos, S. S. Rafail, V. Theodorou,D. Konstantonis, I. Kourtzelis, M. N. Doumas, P. Magotti, R. A. Deangelis, et al.2008. C5a and TNF-alpha up-regulate the expression of tissue factor in intra-alveolar neutrophils of patients with the acute respiratory distress syndrome. J.Immunol. 180: 7368–7375.
7. Redecha, P., C. W. Franzke, W. Ruf, N. Mackman, and G. Girardi. 2008. Neu-trophil activation by the tissue factor/Factor VIIa/PAR2 axis mediates fetal deathin a mouse model of antiphospholipid syndrome. J. Clin. Invest. 118: 3453–3461.
8. Kourtzelis, I., M. M. Markiewski, M. Doumas, S. Rafail, K. Kambas, I. Mitroulis,S. Panagoutsos, P. Passadakis, V. Vargemezis, P. Magotti, et al. 2010. Complementanaphylatoxin C5a contributes to hemodialysis-associated thrombosis. Blood 116:631–639.
9. Szotowski, B., S. Antoniak, W. Poller, H. P. Schultheiss, and U. Rauch. 2005.Procoagulant soluble tissue factor is released from endothelial cells in responseto inflammatory cytokines. Circ. Res. 96: 1233–1239.
10. Parry, G. C., and N. Mackman. 1995. Transcriptional regulation of tissue factorexpression in human endothelial cells. Arterioscler. Thromb. Vasc. Biol. 15: 612–621.
11. Howell, D. C., N. R. Goldsack, R. P. Marshall, R. J. McAnulty, R. Starke,G. Purdy, G. J. Laurent, and R. C. Chambers. 2001. Direct thrombin inhibitionreduces lung collagen, accumulation, and connective tissue growth factor mRNAlevels in bleomycin-induced pulmonary fibrosis. Am. J. Pathol. 159: 1383–1395.
12. Yu, C. C., M. J. Hsu, M. L. Kuo, R. F. Chen, M. C. Chen, K. J. Bai, M. C. Yu,B. C. Chen, and C. H. Lin. 2009. Thrombin-induced connective tissue growthfactor expression in human lung fibroblasts requires the ASK1/JNK/AP-1pathway. J. Immunol. 182: 7916–7927.
13. Imokawa, S., A. Sato, H. Hayakawa, M. Kotani, T. Urano, and A. Takada. 1997.Tissue factor expression and fibrin deposition in the lungs of patients with idi-opathic pulmonary fibrosis and systemic sclerosis. Am. J. Respir. Crit. Care Med.156: 631–636.
14. Kimura, M., K. Tani, J. Miyata, K. Sato, A. Hayashi, S. Otsuka, T. Urata, andS. Sone. 2005. The significance of cathepsins, thrombin and aminopeptidase indiffuse interstitial lung diseases. J. Med. Invest. 52: 93–100.
15. Scotton, C. J., M. A. Krupiczojc, M. Konigshoff, P. F. Mercer, Y. C. Lee,N. Kaminski, J. Morser, J. M. Post, T. M. Maher, A. G. Nicholson, et al. 2009.Increased local expression of coagulation factor X contributes to the fibroticresponse in human and murine lung injury. J. Clin. Invest. 119: 2550–2563.
16. Wick, G., A. Backovic, E. Rabensteiner, N. Plank, C. Schwentner, and R. Sgonc.2010. The immunology of fibrosis: innate and adaptive responses. TrendsImmunol. 31: 110–119.
17. Bogatkevich, G. S., A. Ludwicka-Bradley, and R. M. Silver. 2009. Dabigatran,a direct thrombin inhibitor, demonstrates antifibrotic effects on lung fibroblasts.Arthritis Rheum. 60: 3455–3464.
18. Hinz, B., S. H. Phan, V. J. Thannickal, A. Galli, M. L. Bochaton-Piallat, andG. Gabbiani. 2007. The myofibroblast: one function, multiple origins. Am. J.Pathol. 170: 1807–1816.
19. Erickson, A. M., S. M. de la Monte, G. W. Moore, and G. M. Hutchins. 1987.The progression of morphologic changes in bronchopulmonary dysplasia. Am. J.Pathol. 127: 474–484.
20. Jaarsma, A. S., M. A. Braaksma, W. B. Geven, W. Van Oeveren, andS. B. Oetomo. 2001. Early activation of inflammation and clotting in the pretermlamb with neonatal RDS: comparison of conventional ventilation and high fre-quency oscillatory ventilation. Pediatr. Res. 50: 650–657.
6574 ENDOTHELIN-1 AND TISSUE FACTOR IN FIBROSIS
by guest on June 27, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

21. Merritt, T. A., J. M. Puccia, and I. D. Stuard. 1981. Cytologic evaluation ofpulmonary effluent in neonates with respiratory distress syndrome and bron-chopulmonary dysplasia. Acta Cytol. 25: 631–639.
22. Ogden, B. E., S. A. Murphy, G. C. Saunders, D. Pathak, and J. D. Johnson. 1984.Neonatal lung neutrophils and elastase/proteinase inhibitor imbalance. Am. Rev.Respir. Dis. 130: 817–821.
23. Tullus, K., G. W. Noack, L. G. Burman, R. Nilsson, B. Wretlind, and A. Brauner.1996. Elevated cytokine levels in tracheobronchial aspirate fluids from ventilatortreated neonates with bronchopulmonary dysplasia. Eur. J. Pediatr. 155: 112–116.
24. Kotecha, S., L. Wilson, A. Wangoo, M. Silverman, and R. J. Shaw. 1996. In-crease in interleukin (IL)-1 beta and IL-6 in bronchoalveolar lavage fluidobtained from infants with chronic lung disease of prematurity. Pediatr. Res. 40:250–256.
25. Munshi, U. K., J. O. Niu, M. M. Siddiq, and L. A. Parton. 1997. Elevation ofinterleukin-8 and interleukin-6 precedes the influx of neutrophils in trachealaspirates from preterm infants who develop bronchopulmonary dysplasia.Pediatr. Pulmonol. 24: 331–336.
26. Niu, J. O., U. K. Munshi, M. M. Siddiq, and L. A. Parton. 1998. Early increase inendothelin-1 in tracheal aspirates of preterm infants: correlation with broncho-pulmonary dysplasia. J. Pediatr. 132: 965–970.
27. Hocher, B., A. Schwarz, K. A. Fagan, C. Thone-Reineke, K. El-Hag, H. Kusserow,S. Elitok, C. Bauer, H. H. Neumayer, D. M. Rodman, and F. Theuring. 2000.Pulmonary fibrosis and chronic lung inflammation in ET-1 transgenic mice. Am. J.Respir. Cell Mol. Biol. 23: 19–26.
28. Ross, B., P. D’Orleans-Juste, and A. Giaid. 2010. Potential role of endothelin-1in pulmonary fibrosis: from the bench to the clinic. Am. J. Respir. Cell Mol. Biol.42: 16–20.
29. Clozel, M., and H. Salloukh. 2005. Role of endothelin in fibrosis and anti-fibroticpotential of bosentan. Ann. Med. 37: 2–12.
30. Jobe, A. H., and E. Bancalari. 2001. Bronchopulmonary dysplasia. Am. J. Respir.Crit. Care Med. 163: 1723–1729.
31. de Blic, J., F. Midulla, A. Barbato, A. Clement, I. Dab, E. Eber, C. Green,J. Grigg, S. Kotecha, G. Kurland, et al; European Respiratory Society. 2000.Bronchoalveolar lavage in children. ERS Task Force on bronchoalveolar lavagein children. Eur. Respir. J. 15: 217–231.
32. Kouroumalis, A., R. J. Nibbs, H. Aptel, K. L. Wright, G. Kolios, and S. G. Ward.2005. The chemokines CXCL9, CXCL10, and CXCL11 differentially stimulateG alpha i-independent signaling and actin responses in human intestinal myo-fibroblasts. J. Immunol. 175: 5403–5411.
33. Livak, K. J., and T. D. Schmittgen. 2001. Analysis of relative gene expressiondata using real-time quantitative PCR and the 2(-Delta Delta C(T)) method.Methods 25: 402–408.
34. Huang, M., S. Sharma, L. X. Zhu, M. P. Keane, J. Luo, L. Zhang, M. D. Burdick,Y. Q. Lin, M. Dohadwala, B. Gardner, et al. 2002. IL-7 inhibits fibroblast TGF-beta production and signaling in pulmonary fibrosis. J. Clin. Invest. 109: 931–937.
35. Toti, P., G. Buonocore, P. Tanganelli, A. M. Catella, M. L. Palmeri, R. Vatti, andT. A. Seemayer. 1997. Bronchopulmonary dysplasia of the premature baby: animmunohistochemical study. Pediatr. Pulmonol. 24: 22–28.
36. Dik, W. A., L. J. Zimmermann, B. A. Naber, D. J. Janssen, A. H. van Kaam, andM. A. Versnel. 2003. Thrombin contributes to bronchoalveolar lavage fluidmitogenicity in lung disease of the premature infant. Pediatr. Pulmonol. 35: 34–41.
37. Ricklin, D., G. Hajishengallis, K. Yang, and J. D. Lambris. 2010. Complement:a key system for immune surveillance and homeostasis. Nat. Immunol. 11: 785–797.
38. Egidy, G., L. Juillerat-Jeanneret, P. Korth, F. T. Bosman, and F. Pinet. 2000. Theendothelin system in normal human colon. Am. J. Physiol. Gastrointest. LiverPhysiol. 279: G211–G222.
39. Kojima, N., M. Hori, T. Murata, Y. Morizane, and H. Ozaki. 2007. Differentprofiles of Ca2+ responses to endothelin-1 and PDGF in liver myofibroblastsduring the process of cell differentiation. Br. J. Pharmacol. 151: 816–827.
40. Gando, S. 2010. Microvascular thrombosis and multiple organ dysfunctionsyndrome. Crit. Care Med. 38(2 Suppl):S35–S42.
41. Allen, J. T., R. A. Knight, C. A. Bloor, and M. A. Spiteri. 1999. Enhancedinsulin-like growth factor binding protein-related protein 2 (connective tissuegrowth factor) expression in patients with idiopathic pulmonary fibrosis andpulmonary sarcoidosis. Am. J. Respir. Cell Mol. Biol. 21: 693–700.
42. Scotton, C. J., and R. C. Chambers. 2007. Molecular targets in pulmonary fi-brosis: the myofibroblast in focus. Chest 132: 1311–1321.
43. Shi-Wen, X., Y. Chen, C. P. Denton, M. Eastwood, E. A. Renzoni, G. Bou-Gharios, J. D. Pearson, M. Dashwood, R. M. du Bois, C. M. Black, et al. 2004.Endothelin-1 promotes myofibroblast induction through the ETA receptor viaa rac/phosphoinositide 3-kinase/Akt-dependent pathway and is essential for theenhanced contractile phenotype of fibrotic fibroblasts. Mol. Biol. Cell 15: 2707–2719.
44. Valdenaire, O., E. Rohrbacher, and M. G. Mattei. 1995. Organization of the geneencoding the human endothelin-converting enzyme (ECE-1). J. Biol. Chem. 270:29794–29798.
45. Massai, L., N. Volpi, P. Carbotti, M. Fruschelli, M. Mencarelli, A. Pecorelli,M. Muscettola, M. Agliano, C. Alessandrini, and G. Grasso. 2007. Endothelin-1and endothelin-converting enzyme-1 in human granulomatous pathology ofeyelid: an immunohistochemical and in situ hybridization study in chalazia.Histol. Histopathol. 22: 1343–1354.
The Journal of Immunology 6575
by guest on June 27, 2018http://w
ww
.jimm
unol.org/D
ownloaded from