encyclopedia of inland waters || algae of river ecosystems
TRANSCRIPT

Algae of River EcosystemsR J Stevenson, Michigan State University, East Lansing, MI, USA
ã 2009 Elsevier Inc. All rights reserved.
Introduction
Algae in rivers occupy two distinct habitats, the ben-thos and water column. Benthic algae are algaeattached to or associated with substrates in streamsand rivers. (The term benthic algae is used to refer tomicroscopic and macroscopic algae on or associatedwith substrates. Thus, periphyton and biofilms (micro-phytobenthos), macroalgae (microphytobenthos), andmetaphyton are benthic algae). Phytoplankton aresuspended in the water column. The amount of thesealgae and their function and biodiversity varies greatlyamong different types of streams and rivers and withtime. River ecosystems, broadly defined, include allflowing water sections within a watershed and maybe bordered or interrupted by lakes, reservoirs, andwetlands. Groundwater connections are also impor-tant parts of river ecosystems that affect algae. Algalbiomass is low on substrates inmany shadedmountainheadwater streams or when suspended in large riversduring high flow, but biomass can be very high inbenthos of open canopy streams draining fertilizedlands or as phytoplankton during slow flow, summerconditions. Correspondingly, photosynthesis, respira-tion, and nutrient transformation by algae increases onan areal or volumetric basis with increasing amountsof algae in the habitat, but cell-specific metabolismoften decreases with increasing algal biomass due tocompetition.Taxonomic and morphological diversity varies
among habitats and seasonally within rivers as well asamong rivers with different environmental conditions.Species composition varies in most river ecosystemsfrom diatom-dominated benthos and phytoplanktonduring spring to a varying diversity of green algae andCyanobacteria in warmer waters. However, a widerdiversity of almost all kinds of algae, euglenoids, reds(Rhodophyta), crytomonads, chrysophytes, xantho-phytes, dinoflagellates, and even browns (Phaeophyta)can be found in rivers. Size of these organisms variesfrom less than 5mm for small diatoms and chryso-phytes in the plankton to meters in length for somefilamentous green benthic macroalgae. Morphologi-cally, unicellular, colonial, and filamentous algaeoccur in both benthic and planktonic habitats. Bothunicellular and colonial flagellates use flagella tomove vertically in the plankton. Unicellular diatomsmay use their raphe, and filamentous cyanobacteriamove through sheaths in benthic habitats. Many
114
planktonic species rely simply on the water current orperiodic resuspensions to maintain sufficient exposureto light to survive.
Despite the great variation in algae of rivers andthe complexity of factors that affect them, we havelearned much about the environmental factors thatregulate the processes that control algal biomass,diversity, and function. For example, floods and gra-zing invertebrates can reduce algal biomass, reduceprimary production, and shift species composition toflood- or grazer-tolerant organisms. Whether or notfloods and grazing invertebrates are important inriver ecosystems is determined by the climate, geol-ogy, and resulting hydrology of the ecosystem. In thisarticle, I will use a hierarchical conceptual frameworkfor the factors that affect algae in rivers and streams.This framework will help organize the interrelation-ships among factors that regulate the complex pat-terns of algal biomass, function, and biodiversity, andthereby, will help predict patterns of spatial variationin these algal attributes among ecosystems and sea-sonal variation within these ecosystems. Ultimately,this broader conceptual model will help us predictlonger-term changes in algae of river ecosystemsand how human activities can be managed to solveand prevent environmental problems.
Algae are important elements of river ecosystemsand important determinants of the goods and servicesprovided by rivers. Algae are important in the foodweb for primary production, biogeochemical cycling,and habitat formation and alteration. Algae supporthuman well-being by producing food and cleaningwater for drinking. Excess algae in rivers cause manyproblems by altering physical habitat and depletingdissolved oxygen supplies. This affects biodiversityand productivity of rivers. Algae can also cause taste,odor, and toxicity problems in drinking water suppliesand foul the pipes and filters of water users. So under-standing algal ecology is important for managing riverecosystems to protect the goods and services thatrivers provide.
Factors Affecting Algae in Rivers
Both benthic algae and phytoplankton in rivers livein highly variable habitats. This makes relation ofpattern and process difficult. If we focus our attentionon algae at a specific location, five fundamental

Algae at alocationImmigration
Emigration
Death
GrazingReproduction
Figure 1 Five fundamental processes determine the biomass
and species composition of algae at location.
Algae (Incl. Cyanobacteria) _ Algae of River Ecosystems 115
processes affect how much algae occur in that loca-tion (Figure 1). Immigration of cells or groups of cellsinto the space provides the initial colonists as well asongoing replacement for cells that are continuouslyleaving the habitat via loss factors, such as grazing,emigration (drifting and sinking), and death. Repro-duction by cells within that location, as well as immi-gration, affects accrual positively. These fundamentalprocesses determine the structure of algal commu-nities in rivers, the biomass of individual species aswell as all algae at a location, and therefore speciescomposition. Many other biological processes, suchas photosynthesis, respiration, and nutrient uptake,are considered as functions. Many physical processes,such as eddy mixing and diffusion, affect nutrientavailability and sinking rates (equivalent to emigra-tion from plankton and immigration to benthos).Processes such as eddy mixing and diffusion are con-sidered as indirect processes because they regulatefundamental processes, in this case, reproductionand emigration rates. Finally, both abiotic and bioticfactors either directly or indirectly regulate the fivefundamental processes. These include light intensityand duration, nutrient concentration, and density ofgrazers, bacteria, and viruses that have direct effectsand flood frequency, climate, and geology that haveindirect effects. Many patterns are possible becausethey depend on the relative magnitude of each of thebiological processes and their many direct and indi-rect environmental determinants. Relating patternsin algal biomass and species composition to thebiological processes and environmental factors inrivers require, in most cases, application of nonequi-librium models in algal ecology.Factors that regulate algae in rivers operate at
different spatial and temporal scales (Figure 2). Theprocesses of accrual and losses of cells at a specificlocation operate at the cellular or local spatial scaleand reflect the biological responses to other ecosys-tem factors that are regulated at larger spatial andtemporal scales. Abiotic factors that directly affect
algae include both resources and stressors, such anutrients and light versus pH, salinity, shear stress,and heavy metals. Biotic factors can also be classifiedas positive or negative, and include commensalisticand mutualistic interactions as well as competition,predation, disease, and allelopathic interactions. Atthe habitat scale, riparian canopy, current velocity,and substrate presence and size affect algae, but pri-marily by mediating the direct biotic interactions,resources, and stressors. At watershed and regionalscales, climate and geology ultimately regulate landuse, hydrology, and geomorphology of rivers. Theyalso regulate species biogeography and their avail-ability to colonize rivers. The spatial and temporalhierarchy of these factors will regulate the complexityof possible interactions and facilitate prediction oflocal conditions and algal structure and functionin rivers.
Local Abiotic Factors
Resources
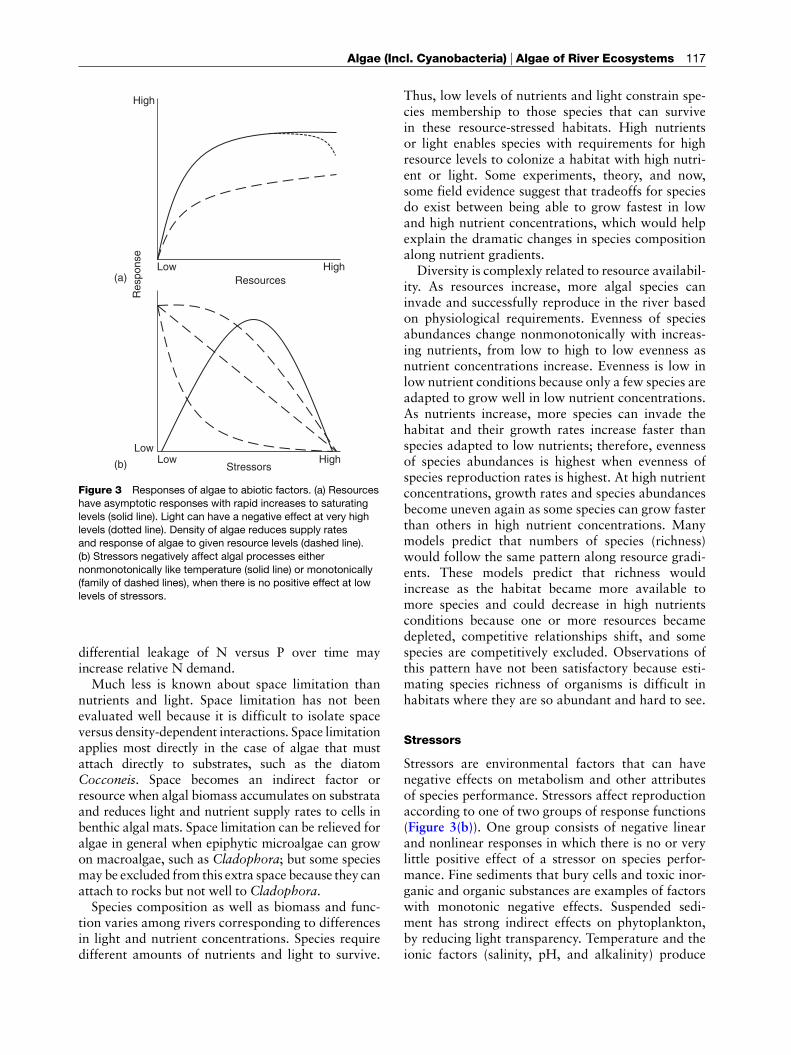
Algal reproduction can be regulated by light, nutri-ents, and space. A variety of metabolic processes havesimilar functional responses to light intensity andnutrient concentration. Metabolic rates increase rela-tively rapidly in response to light and nutrientincreases at low levels of resource availability, buteventually saturate at high levels of resources andrespond little to further increases (Figure 3). Lighthas the possibility of having an additional negativeeffect at very high intensities because of photoin-hibition processes. Light intensity strongly regulatesphotosynthetic ratesandcarbohydratesynthesis.Nutri-ent concentrations regulate their uptake rates and usein protein and lipid synthesis. The differential effects oflight and nutrients on algal metabolism have been usedto explain the paradox of how algae in thin biofilmscan reproduce in very low light as fast as in very highlight (10–1000mmquantam�2 s�1), yet photosyntheticrates of algae are known to respond greatly over thesame range in light conditions.
Variation in light and nutrients does affect algalbiomass, metabolism, and species composition instreams and rivers. High light intensity and nutrientconcentrations are required for accrual of high algalbiomass on substrata or in thewater column. Althoughreproduction of algal growth saturates at low light andnutrient levels, self-shading and reduced nutrient trans-port (because eddy mixing and diffusion are less thannutrient uptake rates) limit resource supply at the localscale as algae accumulate on substrates. For example,

Cel
lula
rLo
cal
Hab
itat
Rea
ch W
ater
shed
R
egio
n
StructureBiomassSpecies compositionDiversity
Species membership
ResourcesLightNutrientsSpace
Biotic subsidiesCommensalismMutualism
Biotic stressorsCompetitorsHerbivoresBioturbationDiseaseAllellopathy
SubstrateSizeComposition
ContaminantsSedimentsNutrients
RiparianCanopyDepth
Currentvelocity
Land use
Hydrology
Climate and geology
Biogeography
Flow stability
Geomorphology
Predators
Bio
logi
cal
Res
pons
e
Dire
ct
Pro
xim
ate
Indi
rect
fact
ors
Inte
rmed
iate
Ulti
mat
e
Factor levelsFactorsScale
Abiotic stressorspHSalinityTemperature
Shear stressAbrasionToxic subs.
FunctionsReproductionDeathImmigrationEmigration
PhotosynthesisRespirationNutrient uptake
Figure 2 Hierarchical interrelationships among proximate, intermediate, and ultimate determinants of benthic algal structure andfunction. The relative spatial scale is shown at the left. Modified from Stevenson RJ (1997) Scale-dependent causal frameworks and the
consequences of benthic algal heterogeneity. Journal of the North American Benthological Society 16: 248–262.
116 Algae (Incl. Cyanobacteria) _ Algae of River Ecosystems
phosphorus limitation of thin biofilms of diatomsrequires concentrations lower than 5mg l�1 PO4–P,whereas 30mg l�1 is required to saturate growth ofthicker mats. As algal biomass increases, more lightand nutrients are required to produce growth andaccrual of high biomasses in relatively short periodsof times. However, high algal biomasses can accumu-late over longer periods of times at lower light andnutrient concentrations in hydrologically stable habi-tats, such as some springs, where disturbance does notinterrupt accumulation processes. Thus, the process-based approach helps us understand how we can havehigh algal biomass at intermediate and high nutrientconcentrations.Experiments using nutrient diffusing substrates and
dosing in experimental streams show that both nitro-gen and phosphorus can limit algal growth in streams.
There is also some evidence that micronutrients canlimit growth of some algae that need large amounts ofan element for enzymes in critical processes, such asFe availability for nitrogen fixation by cyanobacteria.In several published meta-analyses of research withbenthic algae, the trend is for about half of all streamsbeing nutrient limited; and of those streams that arenutrient limited, a quarter are limited by P, a quarterare limited by N, and about half are colimited by bothN and P. In streams with high accumulations of dia-toms, Si may become limiting for both benthic andplanktonic algae. Nutrient ratios have been used suc-cessfully to predict which lakes are limited by N or P,but this approach has not been as successful in streamswhere nutrient concentrations in the water columnare highly variable, supply rates may differ with time,groundwater supplies are not often accounted for, and
Res
pons
e
High
(b)
(a)HighLow
Resources
LowHighLow
Stressors
Figure 3 Responses of algae to abiotic factors. (a) Resources
have asymptotic responses with rapid increases to saturating
levels (solid line). Light can have a negative effect at very highlevels (dotted line). Density of algae reduces supply rates
and response of algae to given resource levels (dashed line).
(b) Stressors negatively affect algal processes either
nonmonotonically like temperature (solid line) or monotonically(family of dashed lines), when there is no positive effect at low
levels of stressors.
Algae (Incl. Cyanobacteria) _ Algae of River Ecosystems 117
differential leakage of N versus P over time mayincrease relative N demand.Much less is known about space limitation than
nutrients and light. Space limitation has not beenevaluated well because it is difficult to isolate spaceversus density-dependent interactions. Space limitationapplies most directly in the case of algae that mustattach directly to substrates, such as the diatomCocconeis. Space becomes an indirect factor orresource when algal biomass accumulates on substrataand reduces light and nutrient supply rates to cells inbenthic algal mats. Space limitation can be relieved foralgae in general when epiphytic microalgae can growon macroalgae, such as Cladophora; but some speciesmay be excluded from this extra space because they canattach to rocks but not well to Cladophora.Species composition as well as biomass and func-
tion varies among rivers corresponding to differencesin light and nutrient concentrations. Species requiredifferent amounts of nutrients and light to survive.
Thus, low levels of nutrients and light constrain spe-cies membership to those species that can survivein these resource-stressed habitats. High nutrientsor light enables species with requirements for highresource levels to colonize a habitat with high nutri-ent or light. Some experiments, theory, and now,some field evidence suggest that tradeoffs for speciesdo exist between being able to grow fastest in lowand high nutrient concentrations, which would helpexplain the dramatic changes in species compositionalong nutrient gradients.
Diversity is complexly related to resource availabil-ity. As resources increase, more algal species caninvade and successfully reproduce in the river basedon physiological requirements. Evenness of speciesabundances change nonmonotonically with increas-ing nutrients, from low to high to low evenness asnutrient concentrations increase. Evenness is low inlow nutrient conditions because only a few species areadapted to grow well in low nutrient concentrations.As nutrients increase, more species can invade thehabitat and their growth rates increase faster thanspecies adapted to low nutrients; therefore, evennessof species abundances is highest when evenness ofspecies reproduction rates is highest. At high nutrientconcentrations, growth rates and species abundancesbecome uneven again as some species can grow fasterthan others in high nutrient concentrations. Manymodels predict that numbers of species (richness)would follow the same pattern along resource gradi-ents. These models predict that richness wouldincrease as the habitat became more available tomore species and could decrease in high nutrientsconditions because one or more resources becamedepleted, competitive relationships shift, and somespecies are competitively excluded. Observations ofthis pattern have not been satisfactory because esti-mating species richness of organisms is difficult inhabitats where they are so abundant and hard to see.
Stressors
Stressors are environmental factors that can havenegative effects on metabolism and other attributesof species performance. Stressors affect reproductionaccording to one of two groups of response functions(Figure 3(b)). One group consists of negative linearand nonlinear responses in which there is no or verylittle positive effect of a stressor on species perfor-mance. Fine sediments that bury cells and toxic inor-ganic and organic substances are examples of factorswith monotonic negative effects. Suspended sedi-ment has strong indirect effects on phytoplankton,by reducing light transparency. Temperature and theionic factors (salinity, pH, and alkalinity) produce

118 Algae (Incl. Cyanobacteria) _ Algae of River Ecosystems
nonmonotonic effects on species performance. Algalperformance is optimal at some intermediate levelof these factors because high and low levels of tem-perature, salinity, pH, and alkalinity limit speciesperformance for one reason or another. Temperature,for example, stimulates metabolism as temperatureincreases from relatively low to intermediate levels askinetic energy increases. However, high temperaturesdenature enzymes and reduce function. Salinity, pH,and alkalinity probably affect enzyme-mediated pro-cesses, and thereby create an intermediate conditionwhich is optimal for species.Because almost all algal species are negatively
affected by sediments and toxic substances, biomassof algae can be negatively affected by these stressors.Similarly, the negative effect of high temperatures(above 30 �C) is ‘toxic’ for most algae, except forsome cyanobacteria that tolerate temperatures as high55 �C in hot springs. So, high temperatures may reducealgal biomass, as low temperatures may. However,algal species are adapted to an unusually wide rangeof pH, alkalinity, and salinity, so that biomass is onlyaffected by the ionic factors in extreme conditions.Throughout much of the range of the ionic factors,species composition differs with varying ionic fac-tors, but functional redundancy is able to maintaincommunity-level reproduction and biomass.
Local Biotic Factors
Biotic factors can have positive or negative effects onalgae in rivers. We know relatively little about com-mensalistic and mutualistic interactions, but someexamples do exist. One example of commensalismwould be the attachment of some benthic diatomson stalks of other diatoms, which provides an advan-tage to species that can attach to stalks and has littlenegative effect on the stalked diatoms. Diatoms withendosymbiotic cyanobacteria provide an example ofmutualistic interactions. Of the negative interactions,much more is known about the herbivory and com-petition versus allellopathic interactions and disease-like effects of fungi, bacteria, and viruses. Many havehypothesized that the latter two biotic stressorsshould have great effects in dense microbial assem-blages like benthic algae. They are known to beimportant in lake and ocean phytoplankton. Unusualwhite circles in periphyton with high numbers ofbacteria and fungi indicate ‘disease,’ but little investi-gation has pursued this line of research.Competition is probably a more important deter-
minant of algal biomass, function, and species com-position for benthic algae than for phytoplankton.Phytoplankton seldom accumulate to sufficiently high
densities to deplete nutrient concentrations in riversbecause of the relatively short residence times of theseorganisms in their habitat. However, competition maybe important in very slow flowing, lake-like riverswhere residence time is sufficiently high to depletenutrients or light by biological uptake or shading.These processes are thought to be important in benthicalgal communities. If species membership is con-strained to diatom-dominated communities, peak bio-mass of communities may be constrained by light andnutrient depletion. We know that light and nutrientavailability within benthic algal mats decreases withincreasing density; less light penetration and decreasingnutrient transport rates and cell nutrient content hasbeen documented as diatom biomass increases on sub-strata. Per capita rates of metabolism and reproductiondecrease with increasing benthic algal biomass. Speciescomposition changes with increasing biomass of dia-toms on substrata. All indicate autogenic changes inenvironment that are consistentwith strong competitiveregulation of benthic algal communities.
Herbivory is also an important determinant ofalgae in rivers. It is more frequently important forbenthic algae than for phytoplankton because of thelackof time for zooplankton to accumulate in thewatercolumn of rivers. Low disturbance frequency by floodsand drought is important for determining whether her-bivory is important for benthic algae, too. When riverconditions allow herbivores to accumulate to high den-sities, they can regulate biomass, function, and speciescomposition of benthic algae in rivers. The importanceof zebra mussels in some rivers is a good example ofherbivores affecting river phytoplankton, but examplesof zooplankton regulation are not common. Filterfeeding invertebrates like blackflies and net-spinningcaddisflies may also be important regulators of sus-pended algal abundance in streams, but this is notwell understood.
Aquatic insects, snails, and some fish consume ben-thic algae, but aquatic insects and snails are the mostimportant in most situations. Protozoa have also beenshown to consume benthic algae, but their impor-tance does not seem to be as great as cased caddisflies,mayflies, and snails that graze algae from substrata.These invertebrates can constrain diatom biomassto very low levels. Many of them can consume fila-mentous green algae during early stages of growth,but not after they have exceeded the size that canbe controlled. They seem to avoid consumption offilamentous Cyanobacteria, but push it back fromactively grazed areas or may knock it off the sub-strate. Invertebrate herbivores can reduce algal bio-masses from 10 to 0.5 mg chl a cm�2. In addition,they selectively graze overstory versus understory dia-toms (stalked and filamentous forms versus tightly

Algae (Incl. Cyanobacteria) _ Algae of River Ecosystems 119
adnate and prostrate forms). Although grazers con-sume algae, not all are killed. Estimates of algaepassed alive and viable through guts of aquatic inver-tebrates often exceed 50%. In a sense, grazing of cellsremoves cells from the substratum and either causesdeath or emigration of the cells downstream whenthey are egested alive. Because of the importanceof grazing as a process of removing algae from alocation and conceptually within food webs, I haveincluded it as one of the five fundamental processes.Bioturbation is another process that affects benthic
algal biomass. Invertebrates, fish, and terrestrial ani-mals are common sources of disturbance of benthicalgae as they move through streams. Diel patternsin algal drift are observed in some stream that corre-spond to the dawn and dusk activity patterns of aquaticinvertebrates. Paths of disturbed benthic algae in shal-low riffles can be observed in deeper upstream–downstream channels where fish have moved frompool to pool. Movement of fish in pools disturbsthe development of periphyton and clouds the water.Raccoons and larger animals, such as manatees, croco-diles, hippopotamus, and humans, probably have greateffects on benthic algae when moving, but these effectshave not been quantified.
Habitat-Scale Factors
Riparian canopy, depth, substrate, and current veloc-ity vary among habitats within a reach and haveindirect effects on the five fundamental algal pro-cesses. Covariation among depth, substrate, and cur-rent velocity are regulated by the riffle-pool structurecarved in most rivers. Coarser substrates are foundin the shallow, high-velocity riffles. Finer substratesare found settling in the deeper, low-velocity pools.Riparian canopy and depth regulate light availability,whereas current velocity and substrate have relativelycomplex effects that warrant further discussion.Substrate is very important for algae as a stable
surface for attachment and growth and potentiallyas a source of nutrition. Some algae have specialmorphological adaptations for attaching to substrata,such as the raphe and mucilaginous stalks and tubesof diatoms. Many filamentous green algae producespecialized cells that attach to a substrate and thenform filaments when they divide. Thus, a major eco-logical division occurs in the algae about which weknow relatively little, except for these morphologicaladaptations. Benthic algae have morphological adap-tations that enable their attachment to substrata, andphytoplankton do not. Given the great differencesin habitat conditions when suspended in the waterversus attached to substrata, such as greater ranges
in current velocity and denser packing of cells forbenthic algae, great physiological differences mustaccompany these morphological adaptations.
For benthic algae, substrate size and nutritionalvalue may have great effects. The smallest substrates,silt and organic sediments, are smaller than the smal-lest algal cells. So organisms that live in this habitattend to be large and motile, such as the diatomsNitzschia and Surirella or the flagellated euglenoids.Sands, slightly larger and inert, are large enoughfor attachment by small diatoms in streams. Duringhydrologically stable periods, other algae can colo-nize sands, but only small diatoms can survive in thecrevices of individual sand grains when the sandstumble across the bottom of streams. As rock sub-strates become larger, they tend to become more resis-tant to hydrologic disturbance. Almost all pebble andlarger substrates can support diverse diatom assem-blages. As substrates become larger and more stable,they can support luxurious growths of filamentousgreen algae that require longer stable periods for colo-nization by spores and growth of filaments. Woodydebris and plants are other common substrates in riv-ers and both may have nutritional properties thataffect algae. Plants leak nutrients that become avail-able to epiphytic algae attached to them. Wood pro-vides nutrition to bacteria, which may actually havea negative effect on benthic algal colonization becauseof competition with bacteria for nutrients.
Current velocity has watershed- and habitat-scaleeffects on algae. For phytoplankton, current velocitydetermines residence time of water and algae in theriver and the time for cells to accumulate. For benthicalgae, increases in current velocity during rain eventsmay be sufficient to scour algae from stable substrates.Higher velocities can disturb the substrate in rifflesand cause severe scouring of algae from substrates.The time between scouring events provides the timefor recovery of benthic algal communities. Thus, var-iations in current velocity at the watershed scale deter-mine the frequency and intensity of disturbance andare important, ultimate determinants of nonequilib-rium ecological dynamics of algae in rivers.
The habitat-scale effects of current velocity onbenthic algae are relatively well understood com-pared to their effects on river phytoplankton. Eddymixing of the water column surely slows sinking ofphytoplankton and shear in the water column mayreduce nutrient depletion in waters surrounding cells.One of the important premises of current effects onalgae is that algae, in still waters, develop nutrient-poor shells of waters around cells. This shell developsbecause nutrient uptake rates from waters aroundcells exceed diffusion rates of nutrients into thosewaters. Water near surfaces, whether cell or substrate

Table 1 Illustrating the changing importance of immigration
versus reproduction during algal community development on
substrates
Day Density Immi Repro
1 1100 1000 100
2 2419 1000 3193 4001 1000 582
4 5897 1000 896
5 8168 1000 1271
6 10887 1000 17197 14139 1000 2252
8 18025 1000 2886
9 22663 1000 3638
10 28190 1000 452711 34766 1000 5576
12 42573 1000 6807
13 51820 1000 8247
14 62741 1000 992115 75595 1000 11854
16 90663 1000 14068
17 108242 1000 1657918 128636 1000 19394
19 152140 1000 22504
20 179023 1000 25883
21 209499 1000 2947622 243699 1000 33200
23 281636 1000 36937
24 323171 1000 40535
25 367984 1000 4381326 415561 1000 46577
27 465193 1000 48632
28 516004 1000 4981129 567001 1000 49997
30 617146 1000 49145
Accumulation of algal cells (density, cells cm�2) on substrates for 30days is
modeled with a spreadsheet. In this model, only immigration (Immi) and
reproduction (Repro) are processes affecting algal cell accumulation.
Grazing, death, and emigration are assumed to be 0. Immigration is
assumed to stay at 1000cellscm�2 day�1 throughout the colonization
period. Twenty percent of all cells from the last day and 10% of the
immigrating cells reproduce each day. A carrying capacity 1000000 cells
cm�2 was included to slow reproduction as density dependent competition
120 Algae (Incl. Cyanobacteria) _ Algae of River Ecosystems
surfaces, has different physical properties than wateraway from these surfaces. Although measurements ofnutrient concentrations in these 1–2-mm thick layersaround cells has not been practical, many observationsbetween current velocity, metabolic rates, and trans-port of nutrients through periphytonmats indicate thatthese shells do exist. Thus, as current velocity aroundcells increases, the shearing of layers of water aroundcells increases and disrupts this layer of nutrient-poor(and potential waste-rich) water from around cells.Habitat-scale effects of current velocity on benthic
algae have both positive and negative effects.Although the shear stress of current velocity decreasesimmigration rates and likely increases emigrationrates from habitats with moderate and fast currentvelocities, the increased physical mixing of waterthrough attached masses of microalgae or macroalgaestimulates metabolic rates. Concentration gradientsin micro- and macroalgal communities can causesevere nutrient depletion within this microhabitat.Assuming a common 10–30 cm s�1 range in velocitiesfrom slow (pool) to fast (riffle) current habitats,profound differences in current effects on immigra-tion and reproduction help relate the patterns ofbenthic algal colonization after flood disturbanceto these processes. Initially, algal biomass and accrualrates will be slower in riffles than in pools becauseimmigration is the dominant process affecting coloni-zation when algal biomass on substrates is low,for example (5–10)� 103 cells cm�2 (Table 1). Asmore cells accumulate, reproduction becomes moreimportant than daily accrual of cells and the positiveeffects of current outweigh the negative effects.So we eventually observe higher algal biomassesin riffles than in pools, even though that patternmay not be evident immediately after flood-relateddisturbances.
increased on substrates.
Reach to Regional Scale Factors
Although reach or segment scale factors (such ashydrology, channel geomorphology, flow stability,and stream size) interact to shape habitat structureand local abiotic conditions, regional processes asso-ciated with climate, geology, and biogeography regu-late the more general spatial and temporal patterns ofalgae in rivers (Figure 2). Climate and geology alsoaffect human activities in watersheds, which haveprofound effects on contaminants in streams, channelgeomorphology, and hydrology. Extensive discussionof human effects of algae in rivers is beyond thescope of this article and is covered elsewhere in thisencyclopedia. I want to synthesize the review ofalgal ecology in rivers by describing three common
patterns: seasonal and longitudinal patterns of phyto-plankton in rivers, algae and the river continuumhypothesis, and the effects of disturbance on benthicalgal–grazer interactions.
Phytoplankton in rivers have a distinct longitudinalpattern that varies in magnitude with seasonal varia-tions in discharge. Suspended algal densities are usu-ally low in headwater sections of rivers and largelycomposed of algae that have emigrated from sub-strates and are entrained in the water column. Aswater flows downstream, species better adapted tolife in the water column immigrate into a mass ofwater and stay in it as water moves downstream.Gradually, fewer benthic species occur in the waterthan do planktonic species. The longer that mass

Algae (Incl. Cyanobacteria) _ Algae of River Ecosystems 121
of water resides in the river before reaching a lake,estuary, or ocean, the higher the biomass of algae inthe water column will become. More phytoplanktonaccumulates in rivers of geologic regions where rivershave low gradients versus those with high gradients.Climate determines the time of year and the extent ofhydrologically stable periods when algae can accu-mulate in streams. In climates with periodic stormsduring the dry season, high discharge events maydisrupt the development of phytoplankton commu-nities. During long dry and hot periods, high andoften problematic biomasses of algae can developwith sufficient quantities that deoxygenation andaccumulation of toxic algae occurs.The river continuum hypothesis proposes that the
ecology of rivers is regulated by upstream–downstreampatterns in the hydrogeomorphology and connectivitywithin rivers. Although originally defined for free-flowing rivers in temperate climateswith forested land-scapes, the model has been adapted for dammed riversand many ecological regions. The original modeldescribed how narrow streams in forests would becovered by a riparian canopy that would limit light,and therefore, algal production. As streams becamewider downstream, lightwould becomemore availableand benthic algae would be able to accumulate. Corre-spondingly, the base of the food web was predicted toshift from allochthonous detritus in the headwatersto autochthonous primary production in the mid-reaches of the river. Farther downstream as algaebecame entrained in the water column and depthincreased, autochthonous productivity in rivers wouldshift from benthic to planktonic algae. Although thisupstream–downstream pattern in hydrogeomorphol-ogy and connectivity may vary greatly among regions,it tends to vary with climate and geologic conditions ofa region, and to some extent with local landscape con-ditions. Most differences occur in headwater regionswhere the river may not have accumulated sufficienterosional power to carve channels and broad flood-plains. Headwaters vary from springs arising in desertsto high gradient channels inmountains or low gradientchannels emerging out of wetlands. Each of theseprovides a different starting point in downstream regu-lation of the ecology of rivers and very different envir-onments for algae.One of the challenges for understanding the ecol-
ogy of algae in streams has been understanding therelationship between resource availability and distur-bance intensity. This challenge underpins: (1) the pre-diction of top-down or bottom-up regulation of riverfood-webs, (2) relationships between nutrients andalgal biomass for management of these important con-taminants of streams, and (3) biodiversity of algae instreams. Increases in nutrient concentrations should
result in an increase in algal biomass of streams.Increases in hydrologic stability also should result inincreases in algal biomass of streams because algaehave, on average, longer periods of time to accumulatebetween disturbances. Thus, in streams that are rela-tively hydrologically stable, we should have greaterresponses of algae to nutrients than in frequently dis-turbed streams. This is what we see in New Zealandstreams where hydrologic stability ranges from greatto average on the global scale. However, in the mid-dle of North America, we see the lower end of thedisturbance continuum. Low disturbance streams res-pond relatively little to increases in nutrient concentra-tions compared to relatively high disturbance streamsin this region. Here, accrual of algae is constrained byhigh grazing pressure in hydrologically stable streams.Flood and drought disturbances in relatively hydrolog-ically variable streams of this region constrain grazerabundances, so grazers no longer regulate algal bio-mass in the most hydrologically disturbed regions ofcentral North America – which compared to the globalrange of possibilities, is only intermediate on thehydrologic disturbance scale. Globally, we find com-plexity arising from the nonlinear and multitrophiclevel effects of disturbance on algal-resource inter-actions. Herbivores and disturbance constrain algalresponse to nutrients in the most hydrologically stablehabitats and the most hydrologically variable habitats,respectively; and algae respond most to nutrientsat intermediate levels of disturbance when they havesufficient time to recover from disturbance, but not somuch time that invertebrates can also recover.
Summary
Interactions between algae and their environment arecomplex, yet predictable based on accurate linkage ofprocesses, scale of determining factors, and algalattributes. Although much is yet to be understoodabout algae in rivers, we have learned much by amarriage of three basic scientific approaches: large-scale field observations from environmental monitor-ing projects, experiments, and process-based models.Study of algae in rivers provides a model for how tounderstand complex systems that are tightly coupledto human effects on the environment. Future work inrivers should address the importance of biodiversityand ecosystem function, how ecological systemsrespond to environmental change, and the impor-tance of algae in ecosystem goods and services. Thelessons that we have learned in the study of algae inrivers should be applied to studies of algae in otherhabitats as well, such as wetlands, terrestrial habitatsin rainforests, and the arctic tundra.

122 Algae (Incl. Cyanobacteria) _ Algae of River Ecosystems
See also: Algae; Aquatic Ecosystem Services; BiologicalInteractions in River Ecosystems; Climate and Rivers;Currents in Rivers; Ecology and Role of HeadwaterStreams; Ecology of Wetlands; Geomorphology ofStreams and Rivers; Groundwater Chemistry; Light,Biological Receptors; Nit rogen; Phosphorus;Photosynthetic Periphyton and Surfaces; RiparianZones; Streams and Rivers as Ecosystems; Streams;The Benthic Boundary Layer (in Rivers, Lakes andReservoirs); Wetlands of Large Rivers: Floodplains.
Further Reading
Biggs BJF (1996) Patterns in benthic algae of streams. In:
Stevenson RJ, Bothwell ML, and Lowe RL (eds.) Algal Ecology:Freshwater Benthic Ecosystems, pp. 31–56. San Diego, CA:
Academic Press.
Biggs BJF, Duncan MJ, Jowett IG, Quinn JM, Hickey CW,
Davies-Colley RJ, and Close ME (1990) Ecological characterisa-tion, classification, and modeling of New Zealand rivers: An
introduction and synthesis. New Zealand Journal of Marineand Freshwater Research 24: 277–304.
Descy JP (1987) Phytoplankton composition and dynamics in the
River Meuse (Belgium). Archiv fur Hydrobiologie 78(Suppl):225–245.
Hauer FR and Lamberti GA (2006) Methods in Stream Ecology,2nd edn. Amsterdam: Elsevier.
Reynolds CS Descy JP (1996) The production, biomass, and struc-ture of phytoplankton in large rivers. Archiv fur Hydrobiologie.113(Suppl): 161–187.
Reynolds CS, Descy JP, and Padisak J (1994) Are phytoplankton
dynamics in rivers so different from shallow lakes? Hydrobiolo-gia 289: 1–7.
Stevenson RJ (1997) Scale-dependent causal frameworks and the
consequences of benthic algal heterogeneity. Journal of the NorthAmerican Benthological Society 16: 248–262.
Stevenson RJ, Bothwell ML, and Lowe RL (1996) Algal EcologyFreshwater Benthic Ecosystems. San Diego: Academic Press.
Stevenson RJ, Rier ST, Riseng CM, Schultz RE, and Wiley MJ(2006) Comparing effects of nutrients on algal biomass in
streams in 2 regions with different disturbance regimes and
with applications for developing nutrient criteria.Hydrobiologia561: 140–165.
Vannote RL,Minshall GW, Cummins KW, Sedell JR, and Cushing CE
(1980) The river continuum concept. Canadian Journal of Fish-eries and Aquatic Sciences 37: 130–137.