electrolytic stimulation of bacteria enterobacter dissolvens by a direct current
TRANSCRIPT

Biochemical Engineering Journal 28 (2006) 23–29
Electrolytic stimulation of bacteriaEnterobacter dissolvensby a direct current
Peng Shea, Bo Songb, Xin-Hui Xing a, Mark van Loosdrechtc, Zheng Liua,∗a Department of Chemical Engineering, Tsinghua University, Beijing 100084, China
b Institute of Rare Precious Animals and Plants, China West Normal University, Sichuan 637002, Chinac Department of Biotechnology, Delft University of Technology, Julianalaan 67, 2628 BC Delft, The Netherlands
Received 16 June 2005; received in revised form 22 July 2005; accepted 16 August 2005
Abstract
The effects of a weak electric field on the growth and metabolic behavior ofEnterobacter dissolvens were investigated using glucose as the solecarbon source. A direct current (DC) was applied using salt bridge electrodes and platinum electrodes. The best stimulating effects in terms of cellgrowth and the dehydrogenase activity (DHA) were obtained when a DC of 10 mA was applied for 12 h via the platinum electrodes. In this case,the electrolysis of water was the major electrode reaction, as determined by cyclic voltammetry. The presence of the hydrogen generated a strongr sumptioni ng the laters ilsf oil.©
K
1
f(fcmaarebtdtio
ble forativeofcellsectorainsttolyticifiedavior
lyticup
ousedd inand
e soled its
1d
eductive environment and led to a reduction of NAD/NADH ratio from 7 to 3. The specific activity of dehydrogenase and glucose conncreased 2- and 1.5-fold, respectively. The application of the DC via the platinum electrodes also led to accelerated cell death duritationary phase. This is possibly due to the presence of anodic intermediates including H2O2, OH• and O2
•. These results provide more detaor understanding the effect of a DC onE. dissolvens, a strain with potential application in the electro-remediation of PAHs contaminated s
2005 Elsevier B.V. All rights reserved.
eywords: Electrolytic stimulation;Enterobacter dissolvens; Direct current; Dehydrogenase activity; Glucose; Redox potential
. Introduction
Electrolytic stimulation refers to a microbial process per-ormed in the presence of electrolysis by a weak direct currentDC). The exposure to a weak DC may lead to an enhancedermentation of yeast[1] and the reduction of nitrate[2,3]. Theurrent intensity for the electrolytic stimulation process is nor-ally kept below 20 mA to avoid the bactericidal effect observedt high density[4], However, negative effects of weak DC havelso been reported, such as the inactivation ofThiobacillus fer-ooxidans and Acidiphilium SJH in liquid culture [5] and thenhancement of antibiotic efficiency against a wide variety ofiofilms [6–8]. These diverse results indicate the complexity of
he electrolytic stimulation process and the growing attentionirected to the fundamental study of the mechanism of elec-
rolytic stimulation. Nakanishi et al.[1] have suggested that themproved yeast growth and ethanol production is due to thexygen generated at the anode. Beschkov et al.[3] assumed
∗ Corresponding author. Tel.: +86 10 6277 9876; fax: +86 10 6277 0304.
that the hydrogen generated at the cathode was responsithe enhanced bacterial denitrification. Regarding the negeffects of a DC, Jackman et al.[5] assumed the deactivationacidophilic bacteria was caused by the interaction of thewith the electrode surface. Stewart et al.[8] confirmed that thgeneration of oxygen by electrolysis is one important fathat leads to the enhancement of Tobramycin efficacy agPseudomonas aeruginosa biofilm. Despite the many effortsincrease the understanding of the mechanism of electrostimulation, a more comprehensive investigation on diversmicrobiological systems that focuses on cell metabolic behin an electric field is still needed.
The objective of the present study is to extend the electrostimulation process toEnterobacter species, an important groof degraders for xenobiotic compounds[9–11] that are widelydistributed in the natural environment. For this, an indigenEnterobacter dissolvens strain isolated from soil contaminatby polycyclic aromatic hydrocarbons (PAHs) was cultivatethe presence of a DC electric field applied via salt bridgeplatinum electrodes, respectively. Glucose was used as thcarbon source due to its explicit metabolic pathway an
E-mail address: [email protected] (Z. Liu). importance as a basic substrate. The electrolytic reaction was
369-703X/$ – see front matter © 2005 Elsevier B.V. All rights reserved.oi:10.1016/j.bej.2005.08.033

24 P. She et al. / Biochemical Engineering Journal 28 (2006) 23–29
followed by cyclic voltammetry. The effects of the DC on thespecific activity of dehydrogenase, glucose consumption andcell death were examined. These results are of fundamentalimportance for the further study of an electric field enhancedbioremediation of PAHs usingE. dissolvens.
2. Experimentals
2.1. Bacterium and chemicals
A gram-negative bacillus strain capable of utilizing phenan-threne as the sole carbon source was isolated from PAHs-contaminated soil sampled in Tianjin, China. The strain wasidentified asE. dissolvens according to its 16 rRNA sequence,which showed a 99% identity with the sequence previousreported for strainE. dissolvens LMG 2683[12]. 2,3,5-Triphenyltetrazolium chloride (TTC) with a purity of 99.0% was pur-chased from Alfa Aesar (Massachusetts, USA). Other reagentsused were of analytically pure grade purchased from standardsuppliers.
2.2. Medium and pre-cultivation
Prior to the electrolytic stimulation experiments, the strainwas cultivated in shake flasks. The culture medium was pre-pared by mixing glucose with basal mineral medium to afi 15),KF di in a3 0 rpma po-n es tione
2.3. Electrolytic stimulation
The electrolytic stimulation experiments were conducted intwo identical vessels with a working volume of 100 ml, as shownin Fig. 1. Two kinds of electrodes were applied in the experi-ments. The first kind was a platinum (Pt) electrode of 0.3 mmin diameter. The second was a salt bridge composed of 1.7%agar and 30% KCl in a basal medium. Prior to the electrolyticexperiments, the two vessels were both autoclaved and filledwith 70 ml of pre-cultivated cell culture. An electric field wasthen applied to one vessel and the other vessel was used as acontrol. During the experiments, the temperature of the waterjacket was maintained at 30◦C. Samples were taken periodi-cally from each vessel and immediately analyzed. The checkingof colony features by agar plating indicates that a pure cultureof E. dissolvens was well maintained during the cultivation.
2.4. Analysis
2.4.1. Cyclic voltammetryThe electrolytic reactions of the Pt electrodes were charac-
terized in the pre-cultivated cell culture by cyclic voltamme-try under an aerobic condition at room temperature. A three-electrode system (EG&G Instruments) was used for measure-ment and the sweep rate was set at 50 mV/s. Pt wires wereused as both the counter electrode and working electrode. As ef-e DHEs d ona furica
2as a
t . 2 ml
of the
nal concentration (in g/l deionized water) of glucose (2HPO4 (0.8), KH2PO4 (0.2), NH4NO3 (0.8), MgSO4 (0.25),eSO4·7H2O (0.09), and CaCl2 (0.032). 0.1 ml of acclimatize
noculum was added into 150 ml of filter-sterilized medium00 ml shake flask and shaken with a rotation speed of 15t 30◦C for 10 h. When cell growth entered the early exential phase (0.2–0.3 in OD600 against fresh medium), thtrain was collected and subjected to electrolytic stimulaxperiments.
Fig. 1. Schematic diagram
aturated calomel electrode (E◦ = 0.2412 V) was used as the rrence electrode. All the potentials are reported on thecale. Before use, the work electrode was carefully polishegold polishing paper, then sonicated for 10 min in 98% sulcid and rinsed with deionized water.
.4.2. Cell dehydrogenase activityThe dehydrogenase activity was measured using TTC
erminal acceptor of protons with glucose as the substrate
CSTR-type electrolytic vessel.
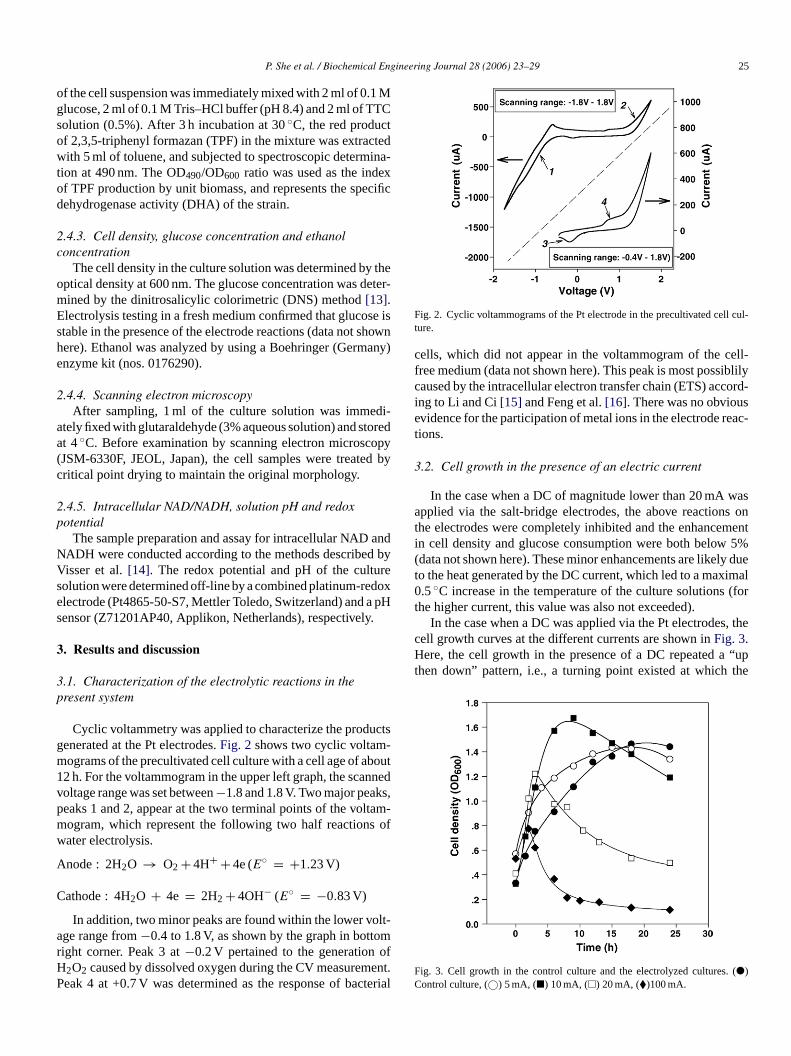
P. She et al. / Biochemical Engineering Journal 28 (2006) 23–29 25
of the cell suspension was immediately mixed with 2 ml of 0.1 Mglucose, 2 ml of 0.1 M Tris–HCl buffer (pH 8.4) and 2 ml of TTCsolution (0.5%). After 3 h incubation at 30◦C, the red productof 2,3,5-triphenyl formazan (TPF) in the mixture was extractedwith 5 ml of toluene, and subjected to spectroscopic determina-tion at 490 nm. The OD490/OD600 ratio was used as the indexof TPF production by unit biomass, and represents the specificdehydrogenase activity (DHA) of the strain.
2.4.3. Cell density, glucose concentration and ethanolconcentration
The cell density in the culture solution was determined by theoptical density at 600 nm. The glucose concentration was deter-mined by the dinitrosalicylic colorimetric (DNS) method[13].Electrolysis testing in a fresh medium confirmed that glucose isstable in the presence of the electrode reactions (data not shownhere). Ethanol was analyzed by using a Boehringer (Germany)enzyme kit (nos. 0176290).
2.4.4. Scanning electron microscopyAfter sampling, 1 ml of the culture solution was immedi-
ately fixed with glutaraldehyde (3% aqueous solution) and storedat 4◦C. Before examination by scanning electron microscopy(JSM-6330F, JEOL, Japan), the cell samples were treated bycritical point drying to maintain the original morphology.
2p
andN d byV res doxe a pHs y.
3
3p
uctsg -m bout1 nnev s,p ltamm s ofw
A
C
olt-a omr ofH entP teri
Fig. 2. Cyclic voltammograms of the Pt electrode in the precultivated cell cul-ture.
cells, which did not appear in the voltammogram of the cell-free medium (data not shown here). This peak is most possiblilycaused by the intracellular electron transfer chain (ETS) accord-ing to Li and Ci[15] and Feng et al.[16]. There was no obviousevidence for the participation of metal ions in the electrode reac-tions.
3.2. Cell growth in the presence of an electric current
In the case when a DC of magnitude lower than 20 mA wasapplied via the salt-bridge electrodes, the above reactions onthe electrodes were completely inhibited and the enhancementin cell density and glucose consumption were both below 5%(data not shown here). These minor enhancements are likely dueto the heat generated by the DC current, which led to a maximal0.5◦C increase in the temperature of the culture solutions (forthe higher current, this value was also not exceeded).
In the case when a DC was applied via the Pt electrodes, thecell growth curves at the different currents are shown inFig. 3.Here, the cell growth in the presence of a DC repeated a “upthen down” pattern, i.e., a turning point existed at which the
F (C
.4.5. Intracellular NAD/NADH, solution pH and redoxotential
The sample preparation and assay for intracellular NADADH were conducted according to the methods describeisser et al.[14]. The redox potential and pH of the cultuolution were determined off-line by a combined platinum-relectrode (Pt4865-50-S7, Mettler Toledo, Switzerland) andensor (Z71201AP40, Applikon, Netherlands), respectivel
. Results and discussion
.1. Characterization of the electrolytic reactions in theresent system
Cyclic voltammetry was applied to characterize the prodenerated at the Pt electrodes.Fig. 2shows two cyclic voltamograms of the precultivated cell culture with a cell age of a2 h. For the voltammogram in the upper left graph, the scaoltage range was set between−1.8 and 1.8 V. Two major peakeaks 1 and 2, appear at the two terminal points of the voogram, which represent the following two half reactionater electrolysis.
node : 2H2O → O2 + 4H+ + 4e (E◦ = +1.23 V)
athode : 4H2O + 4e = 2H2 + 4OH− (E◦ = −0.83 V)
In addition, two minor peaks are found within the lower vge range from−0.4 to 1.8 V, as shown by the graph in bottight corner. Peak 3 at−0.2 V pertained to the generation2O2 caused by dissolved oxygen during the CV measuremeak 4 at +0.7 V was determined as the response of bac
d
-
.alig. 3. Cell growth in the control culture and the electrolyzed cultures.�)ontrol culture, (©) 5 mA, (�) 10 mA, (�) 20 mA, (�)100 mA.

26 P. She et al. / Biochemical Engineering Journal 28 (2006) 23–29
Fig. 4. Morphological changes of cells at different cultivation time (magnification: 10,000×).
cell density started to decline. When the current was lower than10 mA, a significant increase in the cell density in the expo-nential phase was achieved. In the stationary phase, however,the application of the DC led to a significant reduction in thecell density, as compared to those obtained without the DC. Inthe case when high-density currents, i.e., 20 and 100 mA, wereapplied, the increase in cell density was obtained within the first2–3 h and then started to decline in the early stage of the exponen-tial phase. Compared to the cell culture at 5- or 10-mA current,the implementation of high-density currents shifted the turningpoint of cell growth to an earlier time, which was most possiblycaused by the accumulation of anodic intermediates includingH2O2 and OH•, as discussed later in this section.
The results discussed above suggested that the effect of waterelectrolysis on cell growth, as a function of the current density,can be dramatically different. The choice of the DC density isessential for a specific purpose. For the present study, 10 mA isoptimal for enhancing the growth ofE. dissolvens.
Fig. 4gives the surface morphology of electrolyzed cells cul-tivated at 10 mA as well as those in the controlled culture that wasperformed without applying an electric field. Here, cell integritywas maintained up to 12 h cultivation while cell disruption wasobserved when the DC was applied for 24 h. These results mayexplain the reduction in the cell density in the stationary phaseshown inFig. 3.
Wabner and Grambow[17] have usedp-nitrosodimethyl-aniline (RNO) as a selective scavenger to investigate the reactiveintermediates from anodic water oxidation on a Pt electrode. Thesame method was also applied in the present study, and showedthe existence of OH•, H2O2 and O2
• as the products of elec-trolysis, in addition to oxygen and hydrogen. OH• and O2
• hadbeen reported to be potentially harmful to bacterial cells[18].H2O2 and its destructive effects on cell membranes were alsoreported[19]. Thus, we suggested here that these three com-pounds were responsible for the cell damage in the presence of aDC.
3.3. Characterization of the electrolytic stimulation process
To establish a comprehensive picture of the effect of a DCon the cell behavior ofE. dissolvens, the time course of celldensity, specific activity of DHA used as an index of microbialactivity [20], glucose consumption, ethanol production, solutionpH, and both the extracellular and intracellular redox states weredetermined.
3.3.1. Effect of the DC on cell growth and itsdehydrogenase activity (DHA)
As shown inFig. 5A, the cell density and its DHA exhib-ited a similar response to the application of the DC. Within
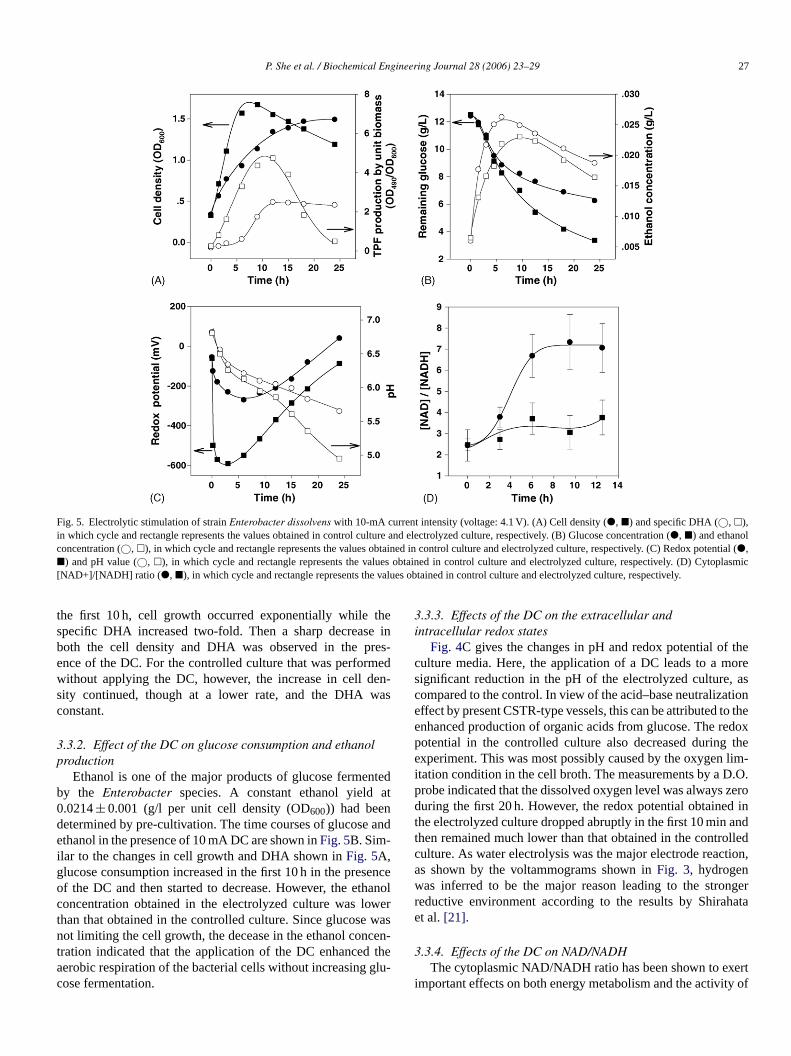
P. She et al. / Biochemical Engineering Journal 28 (2006) 23–29 27
Fig. 5. Electrolytic stimulation of strainEnterobacter dissolvens with 10-mA current intensity (voltage: 4.1 V). (A) Cell density (�, �) and specific DHA (©, �),in which cycle and rectangle represents the values obtained in control culture and electrolyzed culture, respectively. (B) Glucose concentration (�, �) and ethanolconcentration (©, �), in which cycle and rectangle represents the values obtained in control culture and electrolyzed culture, respectively. (C) Redox potential (�,�) and pH value (©, �), in which cycle and rectangle represents the values obtained in control culture and electrolyzed culture, respectively. (D) Cytoplasmic[NAD+]/[NADH] ratio ( �, �), in which cycle and rectangle represents the values obtained in control culture and electrolyzed culture, respectively.
the first 10 h, cell growth occurred exponentially while thespecific DHA increased two-fold. Then a sharp decrease inboth the cell density and DHA was observed in the pres-ence of the DC. For the controlled culture that was performedwithout applying the DC, however, the increase in cell den-sity continued, though at a lower rate, and the DHA wasconstant.
3.3.2. Effect of the DC on glucose consumption and ethanolproduction
Ethanol is one of the major products of glucose fermentedby the Enterobacter species. A constant ethanol yield at0.0214± 0.001 (g/l per unit cell density (OD600)) had beendetermined by pre-cultivation. The time courses of glucose andethanol in the presence of 10 mA DC are shown inFig. 5B. Sim-ilar to the changes in cell growth and DHA shown inFig. 5A,glucose consumption increased in the first 10 h in the presenceof the DC and then started to decrease. However, the ethanolconcentration obtained in the electrolyzed culture was lowerthan that obtained in the controlled culture. Since glucose wasnot limiting the cell growth, the decease in the ethanol concen-tration indicated that the application of the DC enhanced theaerobic respiration of the bacterial cells without increasing glu-cose fermentation.
3.3.3. Effects of the DC on the extracellular andintracellular redox states
Fig. 4C gives the changes in pH and redox potential of theculture media. Here, the application of a DC leads to a moresignificant reduction in the pH of the electrolyzed culture, ascompared to the control. In view of the acid–base neutralizationeffect by present CSTR-type vessels, this can be attributed to theenhanced production of organic acids from glucose. The redoxpotential in the controlled culture also decreased during theexperiment. This was most possibly caused by the oxygen lim-itation condition in the cell broth. The measurements by a D.O.probe indicated that the dissolved oxygen level was always zeroduring the first 20 h. However, the redox potential obtained inthe electrolyzed culture dropped abruptly in the first 10 min andthen remained much lower than that obtained in the controlledculture. As water electrolysis was the major electrode reaction,as shown by the voltammograms shown inFig. 3, hydrogenwas inferred to be the major reason leading to the strongerreductive environment according to the results by Shirahataet al.[21].
3.3.4. Effects of the DC on NAD/NADHThe cytoplasmic NAD/NADH ratio has been shown to exert
important effects on both energy metabolism and the activity of
28 P. She et al. / Biochemical Engineering Journal 28 (2006) 23–29
several enzymes including dehydrogenases[22–24]. Here, thecytoplasmic levels of NAD and NADH were determined in thecell extracts. As shown inFig. 5D, a rapidly increased differ-ence between the magnitude of NAD/NADH was obtained inthe presence and the absence of the DC. The presence of a DCled to a reduction of the NAD/ NADH from 7 to 3, indicat-ing a shift in the intracellular redox equilibrium towards thereduced state. This reduction in NAD/NADH was less thanthat obtained onEscherichia coli and other bacterial species[24] where the cell growth experienced a complete transitionfrom aerobic condition to anaerobic condition. Thus the culti-vation of E. dissolvens in the presence of a DC seems aboutmidway between the aerobic condition and anaerobic condi-tion. This is probably attributable to the simultaneous presenceof hydrogen and the oxygen from air and generated by waterelectrolysis.
3.4. Effects of anodic and cathodic reaction on the cultureof E. dissolvens
It is suggested by the minor shift in the internal redox potentialthat oxygen and hydrogen may simultaneously influence cellbehavior as the two products of water electrolysis. In order todistinguish their individual effects, the electrolytic cultivationof E. dissolvens was conducted using Pt electrode as the anodeo g thee callya olledc turesl en int thec
ares cellD tlyi d to
F lturesrw reset
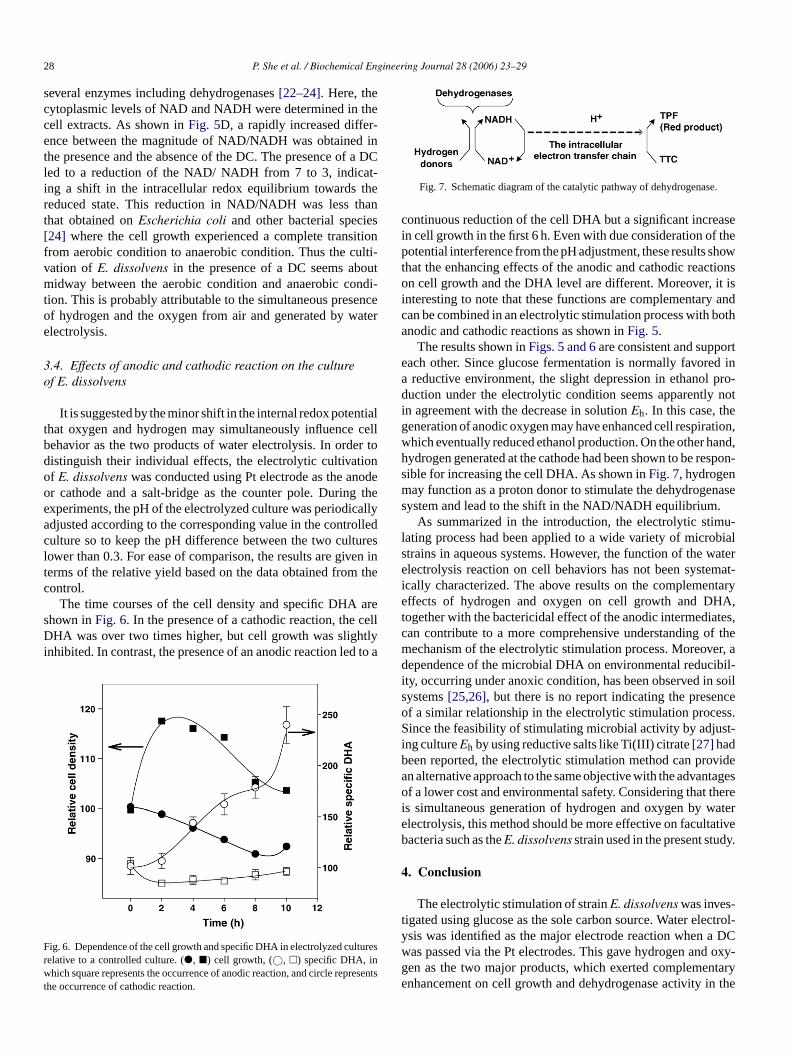
Fig. 7. Schematic diagram of the catalytic pathway of dehydrogenase.
continuous reduction of the cell DHA but a significant increasein cell growth in the first 6 h. Even with due consideration of thepotential interference from the pH adjustment, these results showthat the enhancing effects of the anodic and cathodic reactionson cell growth and the DHA level are different. Moreover, it isinteresting to note that these functions are complementary andcan be combined in an electrolytic stimulation process with bothanodic and cathodic reactions as shown inFig. 5.
The results shown inFigs. 5 and 6are consistent and supporteach other. Since glucose fermentation is normally favored ina reductive environment, the slight depression in ethanol pro-duction under the electrolytic condition seems apparently notin agreement with the decrease in solutionEh. In this case, thegeneration of anodic oxygen may have enhanced cell respiration,which eventually reduced ethanol production. On the other hand,hydrogen generated at the cathode had been shown to be respon-sible for increasing the cell DHA. As shown inFig. 7, hydrogenmay function as a proton donor to stimulate the dehydrogenasesystem and lead to the shift in the NAD/NADH equilibrium.
As summarized in the introduction, the electrolytic stimu-lating process had been applied to a wide variety of microbialstrains in aqueous systems. However, the function of the waterelectrolysis reaction on cell behaviors has not been systemat-ically characterized. The above results on the complementaryeffects of hydrogen and oxygen on cell growth and DHA,together with the bactericidal effect of the anodic intermediates,c f them er, ad ibil-i soils nceo ss.S st-ib videa tageso therei atere ativeb dy.
4
-t ctrol-y a DCw d oxy-g ntarye in the
r cathode and a salt-bridge as the counter pole. Durinxperiments, the pH of the electrolyzed culture was periodidjusted according to the corresponding value in the contrulture so to keep the pH difference between the two culower than 0.3. For ease of comparison, the results are giverms of the relative yield based on the data obtained fromontrol.
The time courses of the cell density and specific DHAhown inFig. 6. In the presence of a cathodic reaction, theHA was over two times higher, but cell growth was sligh
nhibited. In contrast, the presence of an anodic reaction le
ig. 6. Dependence of the cell growth and specific DHA in electrolyzed cuelative to a controlled culture. (�, �) cell growth, (©, �) specific DHA, inhich square represents the occurrence of anodic reaction, and circle rep
he occurrence of cathodic reaction.
a
nts
an contribute to a more comprehensive understanding oechanism of the electrolytic stimulation process. Moreovependence of the microbial DHA on environmental reduc
ty, occurring under anoxic condition, has been observed inystems[25,26], but there is no report indicating the presef a similar relationship in the electrolytic stimulation proceince the feasibility of stimulating microbial activity by adju
ng cultureEh by using reductive salts like Ti(III) citrate[27] hadeen reported, the electrolytic stimulation method can pron alternative approach to the same objective with the advanf a lower cost and environmental safety. Considering that
s simultaneous generation of hydrogen and oxygen by wlectrolysis, this method should be more effective on facultacteria such as theE. dissolvens strain used in the present stu
. Conclusion
The electrolytic stimulation of strainE. dissolvens was invesigated using glucose as the sole carbon source. Water elesis was identified as the major electrode reaction whenas passed via the Pt electrodes. This gave hydrogen anen as the two major products, which exerted complemenhancement on cell growth and dehydrogenase activity

P. She et al. / Biochemical Engineering Journal 28 (2006) 23–29 29
case of a low current intensity, e.g., 10 mA. On the other hand,the presence of a 10 mA current led to cell dissociation during itsstationary phase. Cell growth was also inhibited when a higherDC at 20 or 100 mA was applied.
Acknowledgements
The authors are very grateful to R. Kleerebezem and A.M.Proll in The Deparatment of Biotechnology, TU Delft for theiradvices and supports. The authors would extend their thanks toProf. Dezheng Wang at Department of Chemical Engineering,Tsinghua University for his critical reading and helps in prepar-ing this manuscript.
References
[1] K. Nakanishi, H. Tokuda, T. Soga, T. Yoshinaga, M. Takeda, Effectof electric current on growth and alcohol production by yeast cells, J.Ferment. Bioeng. 85 (1998) 250–253.
[2] A.M. Hayes, J.R.V. Flora, J. Khan, Electrolytic stimulation of denitrifi-cation in sand columns, Water Res. 32 (1998) 2830–2834.
[3] V. Beschkov, S. Velizarov, S.N. Agathos, V. Lukova, Bacterial denitri-fication of waste water stimulated by constant electric field, Biochem.Eng. J. 17 (2004) 141–145.
[4] M. Diao, X.Y. Li, J.D. Gu, H.C. Shi, Z.M. Xie, Electron microscopicinvestigation of the bactericidal action of electrochemical disinfection incomparison with chlorination, ozonation and Fenton reaction, ProcessBiochem. 39 (2004) 1421–1426.
ts ofhilic
ment
the014.B.R.rical
4,6-ol868.
[actor), J.
[ lon,zoic
acid esters (parabens) and their aerobic transformation into phenol bythe resistantEnterobacter cloacae strain EM, Appl. Environ. Microbiol.67 (2001) 2404–2409.
[12] L. Hauben, E.R.B. Moore, L. Vauterin, M. Steenackers, J. Mergaert, L.Verdonck, J. Swings, Phylogenetic position of phytopathogens withinthe Enterobacteriaceae, Syst. Appl. Microbiol. 21 (1998) 384–397.
[13] G.L. Miller, R. Blum, W.E. Glennon, A.L. Burton, Measurement ofcarboxy- methylcellulose activity, Anal. Biochem. 1 (1960) 127–132.
[14] D. Visser, G.A. van Zuylen, J.C. van Dam, M.R. Eman, A. Proll, C. Ras,L. Wu, W.M. van Gulik, J.J. Heijnen, Analysis of in vivo kinetics ofglycolysis in aerobicSaccharomyces cerevisiae by application of glucoseand ethanol pulses, Biotechnol. Bioeng. 88 (2004) 157–167.
[15] H.N. Li, Y.X. Ci, Electrochemical method for analyzing intracellularredox activity changes of the etoposide-induced apoptosis in HL-60 cells,Anal. Chim. Acta 416 (2000) 221–226.
[16] J. Feng, Y.X. Ci, J.L. Lou, X.Q. Zhan, Voltammetric behavior of mam-malian tumor cells and bioanalytical applications in cell metabolism,Bioelectrochem. Bioenerg. 48 (1999) 217–222.
[17] D. Wabner, C. Grambow, Reactive intermediates during oxidation ofwater at lead dioxide and platinum electrodes, J. Electroanal. Chem.195 (1985) 95–108.
[18] T.Z. Liu, T.F. Lin, D.T.Y. Chiu, K.J. Tsai, A. Stern, Palladium or plat-inum exacerbates hydroxyl radical mediated DNA damage, Free Radic.Biol. 23 (1997) 155–161.
[19] N. Hoshino, T. Kimura, A. Yamaji, T. Ando, Damage to the cytoplasmicmembrane ofEscherichia coli by catechin-copper (II) complexes, FreeRadic. Biol. Med. 27 (1999) 1245–1250.
[20] J.C. Praveen-Kumar, Tarafdar, 2,3,5-Triphenyltetrazolium chloride(TTC) as electron acceptor of culturable soil bacteria, fungi and actino-mycetes, Biol. Fert. Soils 38 (2003) 186–189.
[21] S. Shirahata, S. Kabayama, M. Nakano, T. Miura, K. Kusumoto, M.zed-
DNA997)
[ fectsMol.
[ smicle, J.
[ tidecteria
[ s and.
[ ehy-with
7–52.[ anc-
996)
[5] S.A. Jackman, G. Maini, A.K. Sharman, C.J. Knowles, The effecdirect electric current on the viability and metabolism of acidopbacteria, Enzyme Microb. Technol. 24 (1999) 316–324.
[6] S.A. Blenkinsopp, A.E. Khoury, J.W. Costerton, Electrical enhanceof biocide efficacy againstPseudomonas aeruginosa biofilms, Appl.Environ. Microbiol. 58 (1992) 3770–3773.
[7] N. Wellman, S.M. Fortun, B.R. McLeod, Bacterial biofilms andbioelectric effect, Antimicrob. Agents Chemother. 40 (1996) 2012–2
[8] P.S. Stewart, W. Wattanakaroon, L. Goodrum, S.M. Fortun,McLeod, Electrolytic generation of oxygen partially explains electenhancement of tobramycin efficacy againstPseudomonas aeruginosabiofilm, Antimicrob. Agents Chemother. 43 (1999) 292–296.
[9] C.E. French, S. Nicklin, N.C. Bruce, Aerobic degradation of 2,trinitrotoluene by Enterobacter cloacae PB2 and by pentaerythrittetranitrate reductase, Appl. Environ. Microbiol. 64 (1998) 2864–2
10] I.B. Pudge, A.J. Daugulis, C. Dubois, The use ofEnterobacter cloacaeATCC 43560 in the development of a two-phase partitioning biorefor the destruction of hexahydro-1,3,5-trinitro-1,3,5-s-triazine (RDXBiotechnol. 100 (2003) 65–75.
11] N. Valkova, F. Lepine, L. Valeanu, M. Dupont, L. Labrie, J.G. BisailR. Beaudet, F. Shareck, R. Villemur, Hydrolysis of 4-hydroxyben
Gotoh, H. Hayashi, K. Otsubo, S. Morisawa, Y. Katakura, Electrolyreduced water scavenges active oxygen species and protectsfrom oxidative damage, Biochem. Biophys. Res. Commun. 234 (1269–274.
22] J.T. Barron, L.P. Gu, J.E. Parrillo, Cytoplasmic redox potential afenergetics and contractile reactivity of vascular smooth muscle, J.Cell. Cardiol. 29 (1997) 2225–2232.
23] J.T. Barron, L.P. Gu, J.E. Parrillo, Malate-aspartate shuttle, cytoplaNADH redox potential, and energetics in vascular smooth muscMol. Cell. Cardiol. 30 (1998) 1571–1579.
24] J.W. Wimpenny, A. Firth, Levels of nicotinamide adenine dinucleoand reduced nicotinamide adenine dinucleotide in facultative baand the effect of oxygen, J. Bacteriol. 111 (1972) 24–32.
25] M. Brzezinska, Z. Stepniewska, W. Stepniewski, Soil oxygen statudehydrogenase activity, Soil Biol. Biochem. 30 (1998) 1783–1790
26] J. Glinski, Z. Stepniewska, M. Brzezinska, Characterization of the ddrogenase and catalase activity of the soils of two natural sitesrespect to the soil oxygenation status, Pol. J. Soil Sci. 19 (1986) 4
27] G. Jin, A.J. Englande, Redox potential as a controlling factor in enhing carbon tetrachloride biodegradation, Water Sci. Technol. 34 (159–66.