eight cycles of selection for drought tolerance in lowland tropical maize. iii. responses in...
TRANSCRIPT

Field Crops Research, 31 (1993) 269-286 269 Elsevier Science Publishers B.V., Amsterdam
Eight cycles of selection for drought tolerance in lowland tropical maize. III. Responses in drought-
adaptive physiological and morphological traits
J. Bolafios, G.O. Edmeades and L. Martinez CIMMYT, Mexico D.F., Mexico
(Accepted 29 January 1992)
ABSTRACT
Bolafios, J., Edmeades, G.O. and Martinez, L.M., 1993. Eight cycles of selection for drought tolerance in tropical maize. III. Responses in drought-adaptive physiological and morphological traits. Field Crops Res., 31: 269-286.
Selection for grain yield under severe drought stress has often been considered inefficient because the estimate of heritability of grain yield has been observed to decline as yields fall. Under these conditions secondary traits may increase selection efficiency, provided they have adaptive value, high heritability, and are easy to measure. Increased relative stem and leaf elongation rate (RLE), delayed foliar senescence, reduced canopy temperatures and reduced anthesis-silking interval (ASI) were used to augment efficiency of selection for grain yield under drought during eight cycles of recurrent full- sib selection in the lowland tropical maize (Zea mays L.) population, 'Tuxpefio Sequla'. Six cultivars comprising Cycles 0, 2, 4, 6 and 8 of Tuxpefio Sequta, and a check cultivar were grown for two con- secutive years at Tlaltizap~tn, Mexico, under three moisture regimes that provided a well-watered control, a severe moisture stress during flowering, and a severe stress during grain-falling. Previous reports have documented significant improvements in grain yield and ASI in this population. When observed under drought, no significant differences were detected among cultivars in RLE or canopy- air temperature differentials, nor in chlorophyll per unit leaf area during grain-filling (an indicator of foliar senescence). Cultivars did not differ in seasonal predawn or diurnal courses of leaf water po- tential, in leaf osmotic potential, in capacity to adjust osmotically, nor in their seasonal profiles of soil water content with depth to 140 era. Selection significantly altered final plant height, total leaf num- ber, and tassel primary branch number by -0.9%, -0.5% and -2.6% cycle-~, respectively. Obser- vations on root growth in 2-m deep pots showed that eight cycles of selection had reduced root bio- mass in the upper 50 cm by 33%, consistent with a significant change of - 1.2% cycle -~ in vertical root-pulling resistance. The lack of direct and correlated changes in traits related to plant water status due to selection suggests that in this population heritabflities of such traits are low, or that the traits are only weakly associated with grain yield under severe moisture stress. The present study indicates that improved drought tolerance in Tuxpeiio Sequla was due to increased partitioning of biomass towards the developing ear during a severe drought stress that coincided with flowering, rather than to a change in plant water status.
Correspondence to: G.O. Edmeades, CIMMYT, Apdo. Postal 6-641, 06600 Mexico D.F., Mexico.
0378-4290/93/$05.00 © 1993 Elsevier Science Publishers B.V. All fights reserved.

270 J. BOLAI~IOS ET AL.
I N T R O D U C T I O N
Selection for grain yield under environmental stress, as compared to selec- tion under unstressed conditions, has often been considered less efficient be- cause of reductions in the estimate of heritability of grain yield as environ- mental variance rises and observed genetic variance falls (Rosielle and Hamblin, 1981; Blum 1988; Johnson and Geadelmann, 1989). The use of secondary traits can increase selection efficiency under these conditions, pro- vided traits have a clear adaptive value under stress, relatively high heritabil- ity, have a significant genetic correlation with grain yield, and are easy to measure (Falconer, 1960 ). Physiologists and ideotype breeders have long ad- vocated the judicious incorporation of secondary traits within breeding pro- grams (Donald, 1968; Edmeades et al., 1987; Blum, 1988; Ludlow and Mu- chow, 1990; Rasmusson, 1991 ).
Desirable secondary traits associated with improved performance under drought include those which allow plants to gain access to and absorb a greater volume of soil water, to reduce rates of water loss, and to maintain high phys- iological activity at low water potentials (Turner, 1986; Ludlow and Mu- chow, 1990). For example, the capacity to maintain leaf expansion and to retain green leaf area under drought stress is expected to increase light inter- ception and radiation-use efficiency, and result in increased productivity. Several reports have shown that the expansion of leaves is very sensitive to moisture deficits and responds rapidly to changes in leaf water status (Acev- edo et al., 197 I, 1979; Michelena and Boyer, 1982). In tropical maize culti- vars, Sobrado (1986) found a strong relationship between leaf expansion rate under drought and pre-dawn leaf turgor potential, elongation ceasing at tur- gor potentials of less than - 0.2 MPa. Water stress accelerates the senescence of lower leaves in maize, and cultivars with increased capacity for osmotic adjustment have been shown to have delayed leaf senescence under drought (Wright et al., 1983; Hsiao et al., 1984). Other studies comparing the re- sponse of different maize cultivars to drought have shown that those able to maintain higher leaf water potential (leaf ~u) under drought also produced more biomass and grain yield (Ackerson, 1983; Lorens et al., 1987). These reports suggest that selection for an increased rate of leaf and stem extension (RLE) and delayed senescence under drought would improve crop water sta- tus via increased rooting volume and increased capacity for osmotic regulation.
Transpiration consumes energy and cools the leaf, so under conditions of similar energy influx, differences in canopy temperature (CT) reflect varia- tion in rates of transpiration (Jackson, 1982; O'Toole et al., 1984; Blum, 1988 ). Transpiration rate may itself be an indirect measure of the volume of plant-available soil moisture. Portable hand-held infrared thermometers have been used to measure CT and thus provide an indirect field measure of de- hydration avoidance and plant water status in several crop breeding programs (Jackson, 1982; Blum, 1988 ). Selection for reduced CT under drought there-

SELECTION FOR DROUGHT TOLERANCE IN LOWLAND TROPICAL MAIZE III 271
fore could be expected to result in an increase in root length density and root- ing depth.
Recurrent selection for improved performance under drought in the tropi= cal maize population 'Tuxpefio Sequia' was begun in 1975. The superior frac= tion of the population was identified by a selection index comprising in part increased grain yield and a reduced anthesis=silking interval (ASI) under drought. Other traits included in the index were increased RLE, reduced CT and delayed foliar senescence, observed under drought stress. Correlations between grain yield under drought and these secondary traits were usually significant among full=sib families in the early cycles of selection (Fischer et al., 1989), suggesting that they had a positive adaptive value under drought. A comparison of experimental varieties selected from within Cycle 0 for an index of these traits plus grain yield, or for grain yield alone, confirmed the value of the multi=trait selection procedure. Evaluation of the first three selec- tion cycles under severe stress showed that RLE, retention of green leaves and CT had significantly improved with selection, suggesting that these traits have a relatively high heritability (Fischer et al., 1989).
Previous papers in this series described direct and correlated responses to eight cycles of recurrent selection for improved performance under drought in this same population (Bola~os and Edmeades, 1993a,b). Selection re= suited in significant increases in grain yield in droughted environments vary- ing in yield potential from 0.3 to 8 Mg ha- 1. Drought was generally timed to coincide with flowering and grain-filling, and yield gains were associated with improved partitioning toward the developing ear at flowering. This in turn resulted in a marked reduction in ASI under drought and an increase in har- vest index over a range of moisture stress conditions. It seemed likely that reduced ASI was partly the consequence of higher plant ~ during flowering, as silk extrusion is extremely sensitive to plant water deficits (Westgate and Boyer, 1986). Although selection did not significantly affect total biomass production, it resulted in a statistically significant but quantitatively unim- portant improvement in intercepted radiation use efficiency under moisture stress (Bola~os and Edmeades, 1993a,b), perhaps due to delayed foliar senescence.
The objective of this aspect of the study was to evaluate the direct and cor- related responses in drought-adaptive physiological and morphological traits to eight cycles of recurrent selection for drought tolerance in Tuxpefio Sequia. The traits examined relate mainly to plant water status and the maintenance of canopy function, but correlated morphological responses to selection are also reported.
M A T E R I A L S A N D M E T H O D S
Selection procedures for improving drought tolerance of Tuxpefio Sequia have been described in detail elsewhere (Edmeades et al., 1987; Fischer et al.,

272 J. BOLAI~OS ET AL.
1989; Bolafios and Edmeades, 1993a). In each cycle 30-40% of the full-sib families were selected using an index which aimed to: maintain maturity and grain yield constant under well-watered conditions; increase grain yield, RLE and green leaf longevity under drought; and decrease ASI and CT under drought (Fischer et al., 1989).
Prior to evaluation, fresh seed of bulks of Tuxpefio Sequia Cycle 0, 2, 4, 6 and 8 (Co, C2, C4, C6 and C8), and the check, Pop. 21 C6, were prepared (Bolafios and Edmeades, 1993a). These six cultivars were then evaluated in four replications of a randomized complete block design under three mois- ture regimes during the rain-free winter seasons (November-April) of 1986/ 87 ('87 season) and 1987/88 ('88 season) on the CIMMYT Experiment Sta- tion at TlaltizapAn, Mexico. Site characteristics, crop husbandry, experimen- tal design, and data analysis procedures have been described previously (Bo- lafios and Edmeades, 1993a,b). The three water regimes used during evaluation were: (a) well-watered (WW), with water applied approximately every 10 days; (b) intermediate stress (IS) or stress during grain-filling, where irrigation was permanently suspended 65-75 days after planting (DAP); and (c) severe stress (SS), obtained by irrigating only until 44-53 DAP, and al- lowing the crop to complete its life cycle on stored soil water.
Measurements of secondary traits in field experiments. All measurements were taken on well-bordered plants. Leaf and stem elongation rate was measured over two sequential 7-day periods in the SS and WW treatments during both seasons, beginning 3 weeks prior to flowering. The youngest visible leaf in the whorl on eight plants per plot was marked by cutting 5 cm offthe leaf tip. The height from the ground to the cut tip of the vertically extended leaf was mea- sured, and one week later the measurement repeated on the same leaf. A sec- ond set of measurements was initiated immediately using the current young- est visible leaf in the whorl. The absolute increment in height under drought was divided by the corresponding increment under well-watered conditions for the same entry and replicate to give the relative leaf and stem extension rate (RLE). Analysis showed no significant interactions among seasons, mea- surement periods or cultivars, so data for RLE were averaged across measure- ment periods and years.
Canopy-to-air temperature differentials (CT d) were determined by taking 10 readings in the center of each plot with a hand-held infrared thermometer (Model AG-42, Telatemp Corp., Fullerton, CA) having a field of view of 2.5 ° Measurements were performed at 81, 97 and 108 DAP in '87, and at 76 and 92 DAP in '88 (i.e., when radiation interception > 80%), between 12:00- 15:00 h on hot, clear, windless days. The instrument was pointed away from the sun at an angle of depression of 20°and at an angle of incidence to the row of 20 °. The field of view included only fully exposed foliage, avoiding tassels when they were emerged. We attempted to remove time trends in the energy

SELECTION FOR DROUGHT TOLERANCE IN LOWLAND TROPICAL MA1ZE Ill 273
balance of the crop during the measurement period by averaging results from two observers, each of whom proceeded in opposite directions through the plots. Data were averaged across sampling dates and years for stressed (SS, IS) and WW environments.
Differences in leaf senescence were quantified by measuring lamina chlo- rophyll concentrations (#g c m - 2 ) near the center of 12 ear leaves per plot at three sampling times during mid- to late grain-filling. A portable chlorophyll photometer (Design Electronics, Palmerston North, New Zealand) de- scribed by Hardacre et al. (1984) was used. It was calibrated using leaves with a wide range of chlorophyll concentrations (20-70 gg c m - 2 ) , and instru- ment readings were regressed on values obtained from the same leaves by chemical extraction (Arnon, 1949). No cultivar×moisture environment or cultivar× sampling date interactions were observed, so data were averaged across moisture environments and sample dates.
Vertical root-pulling resistance, the maximum force required to pull a plant vertically from the soil (Beck et al., 1987), was measured at 50% anthesis on six plants per plot in each of the water regimes. Each measured plant was separated by at least one undisturbed plant. Measurements were made with a mechanical root puller connected to the lower stalk by a chain and a 200-kg capacity torsion balance. Values were averaged across moisture regimes.
Plant height (here defined as the distance from the ground to the ligule of the uppermost leaf) was measured after anthesis on 10 plants per plot in the WW treatment. During vegetative growth the fifth and tenth leaves of 10 plants per plot were identified by cutting off the leaf tip. Final leaf number (FLN) was recorded on these plants 3 weeks after flowering, and the number of pri- mary tassel branches (TBN) of l 0 additional plants per plot was determined at the same time. Cultivar × moisture environment interactions for TBN or FLN were non-significant, so data for each were averaged across environments.
Pre-dawn (05:00-07:00 h ) leaf ~ of Co, C4, and C8 under SS and WW treat- ments was measured several times 7-14 days apart, beginning at 80 DAP in '87 and at 56 DAP in '88. On four successive days ~was measured at 2-hourly intervals from 06:00-19:00 h to describe diurnal trends. In each plot four young fully exposed and fully expanded leaves were cut with scissors, and wrapped in moist cheesecloth (Turner, 1981 ). Within 1 rain they were sealed in a pressure chamber (Soil Moisture Corporation, Santa Barbara, Califor- nia) by the mid-rib and their balancing pressure determined to the nearest 0.05 MPa.
In '87, water potential components were determined from pressure-volume curves obtained from the youngest fully expanded and fully exposed leaf from each of six plants of each cultivar in SS and WW treatments, using the method described by Cutler et al. (1979). Leaves were sampled during pre-dawn (05:00-07:00 h) at anthesis and were hydrated in water for 4 h before mea-

274 J. BOLAI~OS ET AL.
surement. For each leaf, 1/~ was plotted versus the cumulative volume of expressed sap, V, expressed in terms of leaf relative water content. A linear regression of 1 /~ on V was fitted to the data points (minimum of four) that comprised the linear portion of each graph, i.e., that portion assumed to rep- resent leaf osmotic potential (leaf ~'s). The regression equation was solved for leaf ~/s for each measured value of V, and at a relative leaf water content of 100% to provide an estimate of ~s at full turgor. The difference between the extrapolated value of leaf ~'s and leaf ¢/in the non-linear portion of the plot was considered leaf turgor potential.
In '88, leaf ~,s was determined at 50, 61, 71, 83, 96, and 106 DAP in a different manner, using vapor pressure osmometry and methods described by Bolafios and Edmeades ( 1991 ). A total of 10-12 lamina discs 5 cm in diam- eter were punched from uppermost fully expanded leaves on 10-12 plants per plot during the pre-dawn period (05:00-07:00 h). Discs were immediately wrapped in plastic bags and frozen in dry ice ( - 7 8 ° C ) or in a freezer ( - 18°C). Samples were thawed 24-48 h later, and their sap mechanically expressed as a single bulk sample per plot. No attempt was made to correct for apoplastic dilution. Osmolality (mol kg- 1 ) of this sap was immediately measured with a Wescor Vapor Pressure Osmometer, Model 5500 (Wescor Inc., Logan, Utah). Osmolality was converted to leaf ~ by assuming that 1.0 mol kg- ~ was equal to - 2.48 MPa.
Soil water extraction profiles were determined by the neutron scattering method. A 5-cm diameter aluminum access tube was installed to a depth of 1.6 m in the center of each plot immediately prior to planting. Volumetric soil water contents (SWC) of each plot were determined at regular intervals throughout the season (26, 39, 54, 67, 77, 90, 102, 115, and 125 DAP in '87; and 55, 62, 76, 90, and 104 DAP in '88) using a neutron moisture meter (Model 3300, Troxler Electronic Laboratories, Research Triangle Park, NC ). Measurements were at depths of 30, 52, 74, 96, 118, and 140 cm, and were taken the day immediately before irrigation, so that they reflected the mini- mum SWC experienced by the crop. The neutron moisture meter was cali- brated by regression of instrument measurement on gravimetric determina- tions of SWC from cores obtained during the installation of access tubes. Bulk density ( 1.27 g cm-3) estimated from core measurements were used to con- vert from gravimetric to volumetric SWC.
In the combined analysis of variance, gains from selection were tested for significance by excluding the check cultivar and partitioning the cultivar sum of squares into linear and quadratic contrasts. In all cases the quadratic com- ponent was not significant. Rate of gain per cycle was taken as the linear regression coefficient of trait values on cycle number. Percentage gain per cycle was obtained by dividing gain per cycle by the mean value for the trait across cycles of selection.

SELECTION FOR DROUGHT TOLERANCE IN LOWLAND TROPICAL MAIZE I11 2 7 5
Studies of roots in pots. Because of the difficulties in recovering root systems in field studies, separate experiments were established to study the root sys- tems of Co, C4, and Cs in the summer season of 1987 (May-October) and the winter season of 1989/90 (November-May). Pots comprising PVC tubes 2 m long and 30 cm in diameter (soil volume approximately 135 1) were filled with a 2:1 mixture of alluvial soil:sand and fertilized adequately. Experimen- tal design was a split plot in six replications. Main plots were two moisture treatments, well-watered and stressed, where the stress was induced by with- drawing irrigation 2 weeks prior to 50% anthesis and allowing plants to com- plete their life cycles on soil water stored in the pot. Each plot consisted of two plants in a single pot. In '87, roots were sampled only at harvest, but in '89 roots were sampled at 50% anthesis, mid-grain-filling, and at harvest. Roots were recovered by splitting the pot longitudinally, removing the entire soil core, carefully washing away the soil, and dividing the root system in 25-cm deep horizons. Samples were oven-dried at 80°C for 72 h prior to weighing. During analysis of root weights, no cultivar × harvest date interaction for root weight was detected, so data were averaged for the two experiments and four sample times. Within each horizon the total length of a l-g subsample of roots was estimated by counting the number of intersections in a 2 × 2-cm grid (Newman, 1966), and root dry weight per unit length was calculated.
R E S U L T S
Field experiments. Mean stem and leaf elongations under SS and WW condi- tions were 22.2 and 33.8 cm week- 1 respectively, so that RLE averaged 0.665 (Table 1 ). A non-significant increase of 0.0017 (0.26%) cycle-1 in RLE was observed, but this resulted from a greater decrease in absolute elongation in the WW treatment ( - 0 .20 cm week-1 cycle-1, NS) than in the SS treatment ( - 0.08 cm week- 1 cycle- 1, NS ). These results were consistent with a signif- icant change of - 1.5 cm (0.87%) cycle -1 in plant height (Table 1 ).
Canopy-air temperature differentials averaged 0.47 ° C and - 7.04 ° C under SS and WW conditions, respectively (Table 1 ), and neither cultivar effects nor rate of change with selection were significant. Under Tlaltizap~in condi- tions there were no differences among cultivars in rate of leaf senescence, as measured by chlorophyll concentration during the last half of grain-filling. This result was consistent with visual scores of rates of leaf senescence made in the field during the same period (data not shown).
Significant responses to selection occurred in FLN and TBN, where changes of -0 .11 leaf ( - 0 . 5 % ) cycle -1 and -0 .45 primary branches plant -1 ( - 2.6%) cycle- 1 were observed (Table 1 ). Cultivar × stress level interac- tions for these two traits were not significant. Vertical root-pulling resistance, when measured near anthesis, also responded significantly to selection at a rate of - 1.02 kg plant -1 ( - 1.2%) cycle -1 (Table 1 ). Values of the check
276 J. BOLAI~OS ET AL.
TABLE1
Leaf and stem vertical elongation per week under severe stress (SS) and well-watered (WW) treat- ments, relative leaf and stem elongation rate (RLE), air-to-canopy temperature differentials (CT d) under SS and WW treatments, ear leaf chlorophyll concentration (CHL), plant height (PLHT), final leaf number (FLN), number of tassel primary branches (TBN), and vertical root pulling resistance (RPR) of six maize cultivars when grown in 1986/87 and 1987/88 winter seasons in Tlaltizapfin, Mexico
Cultivar Elongation" RLE" CTa b CHL c PLHT a FLN e TBN e RPR f (cm) (°C) (/tg (cm) (kg)
cm -2) SS WW SS WW
Tux. Seq. Co 22.2 33.9 0.656 0.55 -7 .11 47.6 178.5 21.8 19.1 86.0 Tux. Seq. C2 22.3 33.7 0.661 0.32 -7 .08 49.0 173.5 21.5 17.5 84.6 Tux. Seq. C4 22.4 3.6 0.666 0.23 -6 .96 48.6 169.3 21.7 16.0 83.6 Tux. Seq. C6 21.6 3.0 0.653 0.78 -7 .34 49.5 167.0 20.8 17.0 80.4 Tux. Seq. Cs 21.8 32.2 0.675 0.56 - 6 . 7 0 48.5 166.8 21.0 14.8 77.9 Pop. 21 C6 23.8 36.1 0.658 0.38 -7 .01 48.0 176.5 21.4 18.5 86.5 Mean 22.3 33.8 0.665 0.47 -7 .04 48.5 171.9 21.4 17.2 83.2
Cultivar NS NS NS NS NS NS * * ** NS Sd 0.75 0.52 0.024 0.38 0.39 0.92 8.1 0.24 0.42 4.2 Cycle NS NS NS NS NS NS * * ** NS Rateofchange s - 0 .08 - 0 . 2 0 0.0017 0.02 0.03 0.12 -1 .5"* -0 .11" -0.45** -1 .02"
aAverage weekly stem and leaf vertical elongation. Data are averages of two consecutive weeks and two seasons. RLE = Elongation (SS)/Elongation (WW). bCTd = ( T~opy = Tai,), is the mean of 2-3 sampling dates in each of two years. CCHL is the mean of three sampling dates under stress (SS, IS) across years. dPLHT is for WW treatments across years. eFLN and TBN are means across water regimes and years. fRPR is mean across all water regimes at 50% anthesis for 1987/88 only. SLeast squares estimate of rate of change per cycle. *, ** and NS refer to P< 0.05, P< 0.01 and P> 0.05, respectively.
cultivar for each of these traits were usually similar to those of Tuxpefio Se- quia Co, C2 or C4.
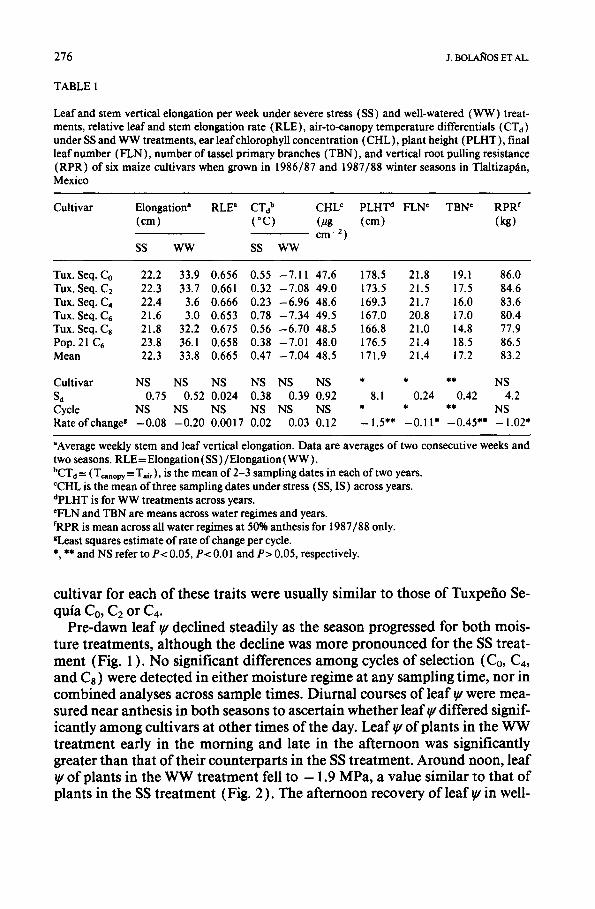
Pre-dawn leaf ¥ declined steadily as the season progressed for both mois- ture treatments, although the decline was more pronounced for the SS treat- ment (Fig. 1 ). No significant differences among cycles of selection (Co, C4, and Ca) were detected in either moisture regime at any sampling time, nor in combined analyses across sample times. Diurnal courses of leaf ~u were mea- sured near anthesis in both seasons to ascertain whether leaf ~ differed signif- icantly among cultivars at other times of the day. Leaf ~u of plants in the WW treatment early in the morning and late in the afternoon was significantly greater than that of their counterparts in the SS treatment. Around noon, leaf ~u of plants in the WW treatment fell to - 1.9 MPa, a value similar to that of plants in the SS treatment (Fig. 2 ). The afternoon recovery of leaf ~u in well-
SELECTION FOR DROUGHT TOLERANCE IN LOWLAND TROPICAL MAIZE II1 277
0,0.
a- - 0 . 2
9- -0.4
- 0 . 6 t-"
-0.8-
17 -1 .0 - .e n
-1 .2 7O
1987
z I I I I
t q
90 110 50
X 1988
t I I 70 90 110 130
Days after planting
Fig. 1. Seasonal course of pre-dawn leaf q/for Co, C4, and C8 of the maize population Tuxpefio Sequia grown under droughted and well-watered conditions during the winter seasons of 1986/ 87 (1987) and 1987/88 (1988) at Tlaltizap~tn, Mexico. Each data point is the mean of 16 observations. Open symbols (O, A and [] ) and closed symbols (O, • and • ) represent Co, C2 and Cs under droughted and well-watered conditions, respectively. Vertical bars represent the Sd among cultivars across water regimes at each sample date.
v
9-
G) ._1
0.0-
- 0 .4 -
- 0 .8 .
- 1 .2 .
-1 .6 .
- 2 . 0
-2 .4
1987
I I I I
8 12 16
1988
I I = I l l I
8 12 16
Hour of day
Fig. 2. Diurnal course of leaf ~u for Co, C4, and Cs of the maize population Tuxpefio Sequia grown under droughted and well-watered conditions during the winter seasons of 1986/87 ( 1987 ) and 1987/88 (1988 ) at Tlaltizap~in, Mexico. Each data point is the mean of 16 obser- vations. Open symbols ( O, A and [] ) and closed symbols (O, • and • ) represent Co, C2 and C8 under droughted and well-watered conditions, respectively. Vertical bars represent the Sd among cultivars across water regimes at each sample date.
watered plants was considerably more rapid than that of the droughted plants. Cultivar effects and cultivar×moisture level interactions for diurnal leaf were not significant at any sample time.
Analysis of internal water relations and osmotic adjustment by pressure-
278 J. BOLAI~IOS ETAL.
0.0
- 0 . 5 -
-1.0-
Q'- -1.5
-3- -2.0
-2.5
- 5 . 0
-3.5 50
Cycle 0
oo
60 70 80 90 O0
Cycle 8 % ~ o °
o~ °~" qL
0 • III I I I I
60 70 80 90 100 Relative water content (%)
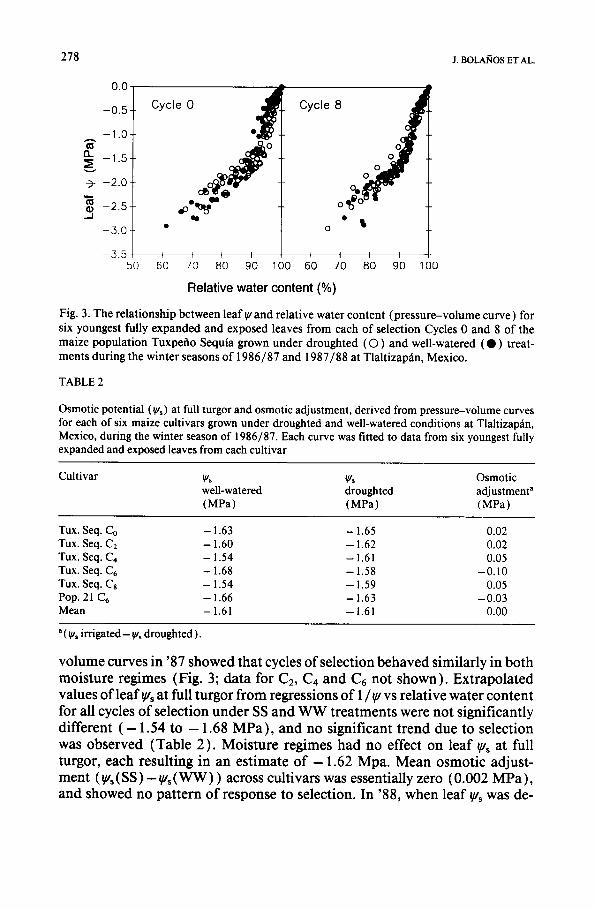
Fig. 3. The relat ionship between leaf ~, and relative water conten t (p res su re -vo lume curve) for six youngest fully expanded and exposed leaves f rom each of selection Cycles 0 and 8 of the maize popula t ion Tuxpefio Sequia grown under droughted ( O ) and well-watered ( • ) treat- ments dur ing the winter seasons of 1986/87 and 1987/88 at Tlaltizapfin, Mexico.
TABLE 2
Osmotic potential (~u,) at full turgor and osmotic adjustment, derived from pressure-volume curves for each of six maize cultivars grown under droughted and well-watered conditions at Tlaltizapfin, Mexico, during the winter season of 1986/87. Each curve was fitted to data from six youngest fully expanded and exposed leaves from each cultivar
Cultivar V, ~& Osmotic well-watered droughted adjustment" (MPa) (MPa) (MPa)
Tux. Seq. Co - 1.63 - 1.65 0.02 Tux. Seq. C2 - 1.60 - 1.62 0.02 Tux. Seq. C4 - 1.54 -1 .61 0.05 Tux. Seq. C6 - 1.68 - 1 . 5 8 - 0 . 1 0 Tux. Seq. Ca - 1.54 - 1.59 0.05 Pop. 21 C6 - 1.66 - 1.63 - 0 . 0 3 Mean - 1.61 - 1.61 0.00
"(~, i r r iga ted- ~u, droughted ).
volume curves in '87 showed that cycles of selection behaved similarly in both moisture regimes (Fig. 3; data for C2, Ca and C6 not shown) . Extrapolated values o f leaf ~s at full turgor from regressions of 1/~u vs relative water content for all cycles o f selection under SS and W W treatments were not significantly different ( - 1.54 to - 1.68 MPa) , and no significant trend due to selection was observed (Table 2) . Moisture regimes had no effect on leaf ~us at full turgor, each resulting in an estimate o f - 1 . 6 2 Mpa. Mean osmotic adjust- ment ( ~'s (SS) - ~u~ ( W W ) ) across cultivars was essentially zero (0.002 MPa ), and showed no pattern of response to selection. In '88, when leaf ~, was de-
SELECTION F O R D R O U G H T T O L E R A N C E IN L O W L A N D T R O P I C A L MAIZE II1 2 7 9
T A B L E 3
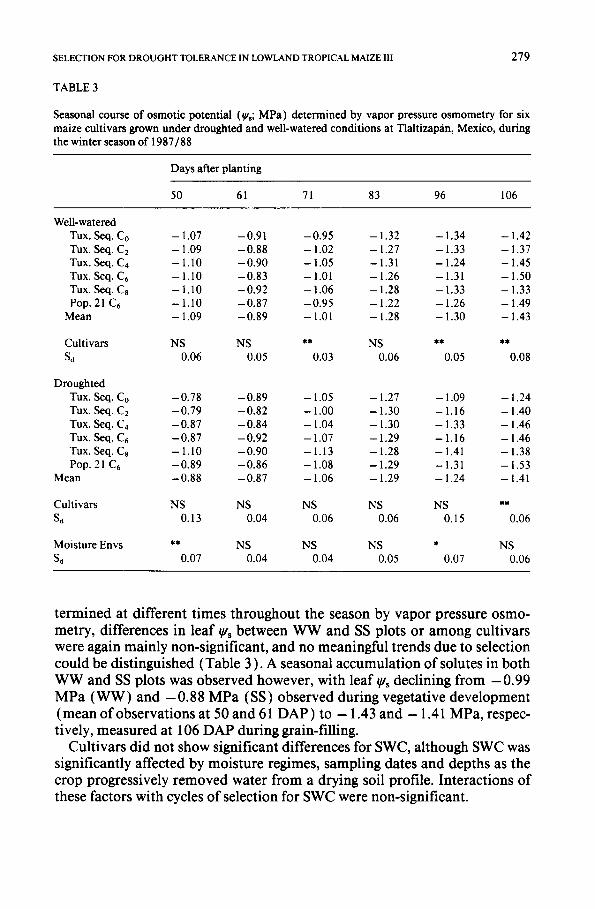
Seasonal course of osmotic potential (~/,; M P a ) determined by vapor pressure osmometry for six maize cultivars grown under droughted and well-watered conditions at Tlaltizap~in, Mexico, during the winter season of 1 9 8 7 / 8 8
Days after planting
50 61 71 83 96 106
Well-watered T u x . S e q . Co - 1 .07 - 0 . 9 1 - 0 . 9 5 - 1 .32 - 1 . 3 4 - 1 .42
T u x . S e q . C2 - 1 .09 - 0 . 8 8 - 1 . 0 2 - 1 .27 - 1 .33 - 1 .37
T u x . S e q . C4 - 1 .10 - 0 . 9 0 - 1 .05 - 1.31 - 1 . 2 4 - 1 .45
T u x . S e q . C6 - 1 . 1 0 - 0 . 8 3 - 1.01 - 1 .26 - 1.31 - 1 .50
T u x . S e q . Ca - 1 . 1 0 - 0 . 9 2 - 1 .06 - 1 .28 - 1 .33 - 1 .33
Pop. 21 C6 - 1 . 1 0 - 0 . 8 7 - 0 . 9 5 - 1 .22 - 1 .26 - 1 .49
Mean - 1 .09 - 0 . 8 9 - 1.01 - 1 .28 - 1 . 3 0 - 1 .43
Cultivars N S N S ** N S ** **
Sd 0 . 0 6 0 . 0 5 0 . 0 3 0 . 0 6 0 . 0 5 0 . 0 8
Droughted T u x . S e q . Co - 0 . 7 8 - 0 . 8 9 - 1 .05 - 1 .27 - 1 .09 - 1 .24
T u x . S e q . C2 - 0 . 7 9 - 0 . 8 2 - 1 . 0 0 - 1 . 3 0 - 1 .16 - 1 .40
T u x . S e q . C4 - 0 . 8 7 - 0 . 8 4 - 1 .04 - 1 . 3 0 - 1 .33 - 1 .46
T u x . S e q . C6 - 0 . 8 7 - 0 . 9 2 - 1 .07 - 1 .29 - 1 .16 - 1 .46
T u x . S e q . C8 - 1 . 1 0 - 0 . 9 0 - 1 .13 - 1 .28 - 1.41 - 1 .38
Pop. 21 C6 - 0 . 8 9 - 0 . 8 6 - 1 .08 - 1 .29 - 1.31 - 1 .53
Mean - 0 . 8 8 - 0 . 8 7 - 1 .06 - 1 .29 - 1 .24 - 1.41
Cultivars N S N S N S N S N S **
Sa 0 . 1 3 0 . 0 4 0 . 0 6 0 . 0 6 0 . 1 5 0 . 0 6
Moisture Envs ** N S N S N S * N S
Sd 0 . 0 7 0 . 0 4 0 . 0 4 0 . 0 5 0 . 0 7 0 . 0 6
termined at different times throughout the season by vapor pressure osmo- merry, differences in leaf ~'s between WW and SS plots or among cultivars were again mainly non-significant, and no meaningful trends due to selection could be distinguished (Table 3 ). A seasonal accumulation of solutes in both WW and SS plots was observed however, with leaf ~gs declining from - 0.99 MPa (WW) and - 0.88 MPa (SS) observed during vegetative development (mean of observations at 50 and 61 DAP) to - 1.43 and - 1.41 MPa, respec- tively, measured at 106 DAP during grain-filling.
Cultivars did not show significant differences for SWC, although SWC was significantly affected by moisture regimes, sampling dates and depths as the crop progressively removed water from a drying soil profile. Interactions of these factors with cycles of selection for SWC were non-significant.
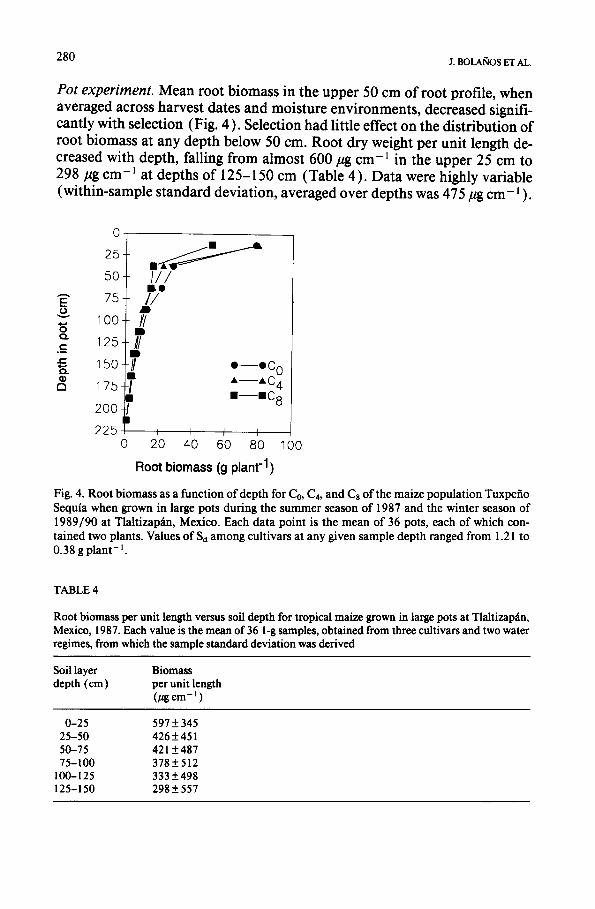
280 J. BOLAROS ET AL.
Pot experiment. Mean root biomass in the upper 50 cm of root profile, when averaged across harvest dates and moisture environments, decreased signifi- cantly with selection (Fig. 4 ). Selection had little effect on the distribution of root biomass at any depth below 50 cm. Root dry weight per unit length de- creased with depth, falling from almost 600/~g cm-1 in the upper 25 cm to 298 ~tg cm -~ at depths of 125-150 cm (Table 4). Data were highly variable (within-sample standard deviation, averaged over depths was 475/tg c m - I ).
E O
O ¢-j
¢ -
e -
K fD
t21
0
25
50
75
100
D
175 ~.~
0
/ / / I • / /
m f~
m
• •C O • AC 4 u - - m C 8
I I I I 2O 40 60 8O 100
Root biomass (g plant "1)
Fig. 4. Root biomass as a function of depth for Co, C4, and Ca of the maize population Tuxpefio Sequia when grown in large pots during the summer season of 1987 and the winter season of 1989/90 at Tlaltizap~tn, Mexico. Each data point is the mean of 36 pots, each of which con- tained two plants. Values of Sd among cultivars at any given sample depth ranged from 1.21 to 0.38 g plant- ~.
TABLE4
Root biomass per unit length versus soil depth for tropical maize grown in large pots at Tlaltizap~in, Mexico, 1987. Each value is the mean of 36 l-g samples, obtained from three cultivars and two water regimes, from which the sample standard deviation was derived
Soil layer Biomass depth (cm) per unit length
(/tg cm - l )
0-25 597 + 345 25-50 426 + 451 50-75 421 +487 75-100 378 + 512
100-125 333+498 125-150 298 + 557

SELECTION FOR DROUGHT TOLERANCE IN LOWLAND TROPICAL MAIZE 111 281
No significant differences were detected in weight per unit length of root be- tween moisture regimes nor among cultivars.
D I S C U S S I O N
Direct responses to selection. No significant progress towards increased RLE, reduced rates of leaf senescence, or reduced canopy temperatures under mois- ture stress was observed (Table 1 ), even though these traits were included in the selection index used to improve the performance of Tuxpefio Sequia un- der drought. All are indirect measures of improved plant water status under moisture stress. This contrasts with an earlier comparison of Co and C3 in this population which, under severe drought, showed significant improvements in RLE (from 0.57 to 0.66), in retention of green leaf area during grain-filling (from 58% to 74%), in CT (from 34.7°C to 32.5°C), and in ASI (from 7.1 days to 5.0 days). In the same study grain yield, measured under drought, increased from 0.62 to 1.03 Mg ha - ' (Fischer et al., 1989). In the present study these improvements in yield and ASI have been demonstrated again (Bolafios and Edmeades, 1993a,b), but the previously observed changes in secondary traits used during selection could not be confirmed. The lack of observed progress in RLE, CTd and delayed canopy senescence in this recent evaluation may be because C3 was not included, or because the level of drought was more severe than that used previously. Grain yields in the SS treatment were only 0.26-0.41 Mg ha- 1, versus 0.62-1.03 Mg ha- 1 in the earlier study, and this increase in severity may have masked gains made in secondary traits.
Other explanations for the lack of direct selection responses in RLE, CTd and canopy senescence include a lack of causal association between them and grain yield, and low heritability or limited genetic variability for the trait (Blum, 1988 ). Although Fischer et al. (1989) reported significant linear cor- relations between these traits and grain yield of full-sib families from C2 and C3 of Tuxpefio Sequia when grown under drought, similar studies among $1 or full-sib progeny from a wider array of tropical maize germplasm have failed to show consistent confirmation of these relationships. Of the secondary traits it is only ASI which has been reliably and significantly correlated with grain yields in maize progenies grown under drought in small single-row plots (Bo- lafios and Edmeades, 1990). This lack of consistency in degree of association may be due to the sensitivity of linear correlation coefficients to outlying val- ues. Another reason for lack of progress in secondary traits could be a low effective heritability, which can arise because additive genetic variation is low (Falconer, 1960), or because of inaccurate measurement of the trait. For example, RLE is determined from the difference in distance from leaf tip to ground over a 7-day period. A small change in the position from which the measurements are made, especially easy when plants are grown on ridges, can introduce serious errors in the measurement of incremental height change.

282 J. BOLAI~OS ET AL.
Canopy temperature measurements are also subject to error because of inad- equate sampling of the target, background interference, and the influence of cloud and wind during measurement. Fischer et al. (1989), using an infrared thermometer with an angle of acceptance of 28 °, reported linear correlation coefficients between CT and grain yield ranging from - 0.40 to - 0.73 for 250 families growing under severe drought stress. The instrument used in the present evaluation had an angle of acceptance of only 2.5 °, and the smaller surveillance area could have been, on occasion, made up entirely of back- ground rather than target. Similarly, selection for reduced leaf senescence rate was usually by means of a weekly visual score subject to observer error.
Correlated responses in morphological traits. Plant height, FLN and TBN were not under direct selection during the development of Tuxpefio Sequia, yet all were significantly reduced as selection progressed. The decline in height and FLN can be directly associated with a reduction in days to flower (Bolafios and Edmeades, 1993b), caused by the tendency of early-flowering families to escape drought stress that intensified during the flowering period. The de- crease in TBN of 2.6% cycle-~ paralleled an observed reduction in tassel bio- mass of 2.6% cycle-~ (Bolafios and Edmeades, 1993b). In the closely related population, 'Tuxpefio Crema I', direct selection for reduced TBN led to a sig- nificant increase in female inflorescence biomass at 50% silking, a reduction in tassel biomass, and an increase in harvest index (Fischer et al., 1987). These results, and those of the present study (Bolafios and Edmeades, 1992b), suggest that reduced tassel size in tropical maize is directly associated with increases in ear growth at flowering and in harvest index.
Correlated responses in measures of plant water status. An objective of selec- tion was to improve the water relations of the crop under drought by selecting for simple physiological traits that were indirect indicators of improved water status (Edmeades et al., 1987; Fischer et al., 1989). Other studies comparing cultivars under drought have shown that increases in grain yield, biomass production and harvest index were correlated with the ability of the cultivars to maintain a higher leaf ~u or leaf turgor potential under drought, either be- cause of improved soil water uptake or through osmotic adjustment (Acker- son, 1983; Bennett et al., 1986; Lorens et al., 1987). Selection in Tuxpefio Sequia had no effect on total biomass production (Bolafios and Edmeades, 1993a), nor on soil water extraction patterns at depths down to 140 cm at any time during the season in either moisture environment. Since soil ¢/and leaf ~u would normally be in equilibrium by early morning, we also used pre-dawn leaf ~u as an indirect measure of the cultivar's ability to explore the soil vol- ume, but found it unaffected by selection. These observations suggest that the total volume of water transpired by the crop was little affected by selection,

SELECTION FOR DROUGHT TOLERANCE IN LOWLAND TROPICAL MAIZE Ill 283
and imply that changes, if any, in rooting depth, root length density and in osmotic adjustment played a relatively minor role in explaining the observed improvements in grain yield. This was examined by more direct measure- ments of these traits.
The negligible role played by osmotic adjustment in explaining changes in grain yield and ASI was confirmed by direct measurement. Seasonal and diur- nal courses of leaf ~ (Figs. 1,2 ) and capacity for osmotic adjustment (Tables 2, 3 ) showed no significant response to selection. Maize has been reported to adjust osmotically in response to drought (Acevedo et al., 1979; Sobrado, 1986; Westgate and Boyer, 1986; Bolafios and Edmeades, 1991 ), thus main- taining cell turgor and turgor-related metabolic processes in the face of declin- ing ~u (Ludlow and Muchow, 1990 ). Limited capacity for osmotic adjustment was observed in 204 maize cultivars, mainly tropical in origin, grown under drought at the same site (BolaSos and Edmeades, 1991 ). This suggests that the extent of genetic variability for this trait in tropical germplasm may be small. Alternatively, it is possible that well-watered plants are fully adjusted in this arid winter environment where leaf ~, of control plants may fall to - 1.9 MPa at noon (Fig. 2 ) and CTa of well-watered plots may reach - 7 °C (Table 1 ). Although almost no osmotic adjustment was observed in Tuxpefio Sequia, a seasonal decline in leaf ~us was detected (Table 3 ), suggesting that a mechanism for accumulating solutes remained functional. The seasonal de- cline in pre-dawn leaf ~u observed in well-watered plots (Fig. 1 ) is consistent with other field reports of maize (Fereres et al., 1978; Wolfe et al., 1988 ), and was probably due to an increase in resistance to water uptake as the crop aged.
Lack of progress in traits indicative of plant water status may also be a consequence of using small, unbordered plots (one row 2.5 m in length ) dur- ing selection. Traits that provide a comparative advantage in single-row plots during selection, such as aggressive rooting, rapid canopy expansion and lax leaves, may be of no advantage in large plots. Conversely, families which grow slowly and conserve soil water early in the growth cycle for later use could be penalized by their more competitive neighbors. It seems unlikely however that this entirely explains the observed lack of progress in osmotic adjustment.
Roots. Rooting patterns observed in large pots indicated a loss of superficial roots with selection (Fig. 4) and an overall reduction in root biomass. This is consistent with the observed change of - 1.02 kg plant-1 cycle-~ in vertical root-pulling resistance (Table 1 ). Root pulling resistance has been shown to depend proportionately more on the characteristics of the upper than the lower root layers (Kevern and Hallauer, 1983 ). The loss of root biomass with selec- tion was surprising, since it was believed that selection favored families with deep rooting and a high root length density (Fischer et al., 1989). The selec- tive loss of roots in the upper 50 cm probably reflects the fact that during selection soil water deficits decreased in severity downwards from the soil

284 j. BOLAI~OS ET AL.
surface, leaving the upper 50 cm of soil too dry for active root growth. New root growth in this this zone, if it were possible, would have little adaptive value. Had selections been made under frequent but light sprinkler irriga- tions, it seems probable that superficial root biomass may have increased at the expense of deeper roots. Because shoot water status and biomass produc- tion were unaffected by selection (Bola~os and Edmeades, 1993a), these changes in root distribution imply that the effect of root signals on shoot growth (Davies and Zhang, 1991 ) was also unchanged.
Differences in rooting biomass with depth may provide a guide to the amount and timing of water availability to the crop, although they do not necessarily estimate the ability ofcultivars to extract water (Passioura, 1986). Root length densities of 1.0 c m cm -3 are generally considered sufficient for complete extraction of soil water, although for most crops these are larger than 3.0 cm era- 3 near the surface (O'Toole and Bland, 1987 ). The root length densities of C8 and Co in the upper 25 cm of the pot, admittedly an artificial rooting environment, were 5.11 and 7.58 c m c m -3 , respectively. There was no evidence that the loss of roots accompanying selection resulted in inade- quate water extraction in the upper part of the profile. On the other hand, the reduction in root biomass may partially account for the observed increase in ear growth in this tropical maize population (Bolafios and Edmeades, 1993b).
In conclusion, these data suggest that responses to selection in grain yield, ASI and harvest index (Bolafios and Edmeades, 1993a,b) were not a conse- quence of improved water relations, but rather were due to improved parti- tioning ofbiomass toward the female inflorescence at flowering. This was ac- companied by a reduction in biomass in organs (tassels and upper roots) whose growth may compete with that of the developing ear. Drought, when managed to coincide with flowering, apparently provided an effective selec- tion environment for distinguishing families that partitioned more assimi- lates toward the ear, and the subsequent changes in grain yield, ASI and har- vest index due to selection were virtually independent of changes in plant water status.
ACKNOWLEDGEMENTS
We thank our field staff, P. Galvez, J.C. Bahena, H. Corrales, C. Figueroa and E. Bahena, for plot management and assistance with data collection. We also thank J.A. Deutsch, H.R. Lafitte, S.C. Chapman and M. Reynolds for helpful discussions of earlier drafts of this paper.
REFERENCES
Acevedo, E., Hsiao, T.C. and Henderson, D.W., 1971. Immediate and subsequent growth re- sponses of maize leaves to changes in water status. Plant Physiol., 48: 631-636.

SELECTION FOR DROUGHT TOLERANCE IN LOWLAND TROPICAL MAIZE Iii 285
Acevcdo, E., Fereres, E., Hsiao, T.C. and Henderson, D.W., 1979. Diurnal growth trends, water potential, and osmotic adjustment of maize and sorghum leaves in the field. Plant Physiol., 64: 476-480.
Ackerson, R.C., 1983. Comparative physiology and water relations of two corn hybrids during water stress. Crop Sci., 23: 278-283.
Arnon, D.I., 1949. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vul- garis. Plant Physiol., 24: 1-5.
Beck, D.L., Darrah, L.L. and Zuber, M.S., 1987. An improved technique for measuring resis- tance to roo t pulling in maize. Crop Sci., 27: 356-358.
Bennett, J.M., Jones, J.W., Zur, B. and Hammond, L.C., 1986. Interactive effects of nitrogen and water stresses on water relations of field-grown corn leaves. Agron. J., 78:273-280.
Blum, A., 1988. Plant Breeding for Stress Environments. CRC Press, Boca Raton, Florida, 223 PP.
Bolafios, J. and Edmeades, G.O., 1990. CIMMYT's strategies in breeding for drought tolerance in tropical maize. In: P.W. Unger, W.R. Jordan, T.V. Sneed and R.W. Jensen (Editors), Challenges in Dryland Agriculture - A Global Perspective. Proc. of the Int. Conf. on Dryland Farming. Texas Agric. Exp. Stations, College Station, TX, pp. 752-754.
Bolafios, J. and Edmeades, G.O., 1991. Value of selection for osmotic potential in lowland trop- ical maize. Agron. J. 83: 948-956.
Bolafios, J. and Edmeades, G.O., 1993a. Eight cycles of selection for drought tolerance in low- land tropical maize. I. Responses in grain yield, biomass, and radiation utilization. Field Crops Res., 31: 233-251.
Bolaflos, J. and Edmeades, G.O., 1993b. Eight cycles of selection for drought tolerance in low- land tropical maize. II. Responses in reproductive behavior. Field Crops Res., 31: 258-268.
Cutler, J.M., Shahan, K.W. and Steponkus, P.L., 1979. Characterization of the internal water relations of rice by a pressure-volume method. Crop Sci., 19:681-685.
Davies, W.J. and Zhang, J., 1991. Root signals and the regulation of growth and development of plants in drying soil. Annu. Rev. Plant Physiol. Plant Mol. Biol., 42: 55-76.
Donald, C.M., 1968. The breeding of crop ideotypes. Euphytica, 17: 385-403. Edmeades, G.O., Fischer, K.S. and Islam, T.M.T., 1987. Improvement of maize yield under
drought stress. In: J.M. Menyonga, T. Bezuneh and A. Youdeowei (Editors), Food Grain Production in Semi-Arid Africa. OUA/STRC-SAFGRAD, Ouagadougou, Burkina Faso, pp. 191-210.
Falconer, D.A., 1960. Introduction to Quantitative Genetics. Ronald Press, New York, 380 pp. Fereres, E., Acevedo, E., Henderson, D.W. and Hsiao, T.C., 1978. Seasonal changes in water
potential and turgor maintenance in sorghum and maize under water stress. Physiol. Plant., 44: 261-267.
Fischer, K.S., Edmeades, G.O. and Johns0n~'E.C., 11987. Recurrent selection for reduced tassel branch number and reduced leaf area density above the ear in tropical maize populations. Crop Sci., 27:1150-1156.
Fischer, K.S., Edmeades, G.O. and Johnson, E.C., 1989. Selection for the improvement of maize yield under moisture-deficits. Field Crops Res., 22: 227-243.
Hardacre, A.K., Nicholson, H.F. and Boyce, M.L.P., 1984. A portable photometer for the mea- surement of chlorophyll in intact leaves. N.Z.J. Exp. Agric., 12: 357-362.
Hsiao, T.C., O'Toole, J.C., Yambao, E.B. and Turner, N.C., 1984. Influence of osmotic adjust- ment on leaf rolling and tissue death in rice (Oryza sativa L. ). Plant Physiol., 75: 338-341.
Jackson, R.D., 1982. Canopy temperature and crop water stress. Adv. Irrig., 1: 43-85. Johnson, S.S. and Geadelmann, J.L., 1989. Influence of water stress on grain yield response to
recurrent selection in maize. Crop Sci., 29: 558-565. Kevern, T.C. and Hallauer, A.R., 1983. Relation of vertical root-pull resistance and flowering
in maize. Crop Sci., 23: 357-363.

286 J. BOLAI~OS ET AL.
Lorens, G.F., Bennett, J.M. and Loggale, L.B., 1987. Differences in drought resistance between two corn hybrids. I. Water relations and root length density. Agron. J., 79: 802-807.
Ludlow, M.M. and Muchow, R.C., 1990. A critical evaluation of traits for improving crop yields in water-limited environments. Adv. Agron., 43:107-153.
Michelena, V.A. and Boyer, J.S., 1982. Complete turgor maintenance at low water potentials in the elongation region of maize leaves. Plant Physiol., 69:714-718.
Newman, E.I., 1966. A method for estimating the total length of root in a sample. J. Appl. Ecol., 3: 139-145.
O'Toole, J.C. and Bland, W.L., 1987. Genotypic variation in crop plant root systems. Adv. Agron., 41: 91-145.
O'Toole, J.C., Turner, N.C., Namuco, O.P., Dingkuhn, M. and Gomez, K.A., 1984. Compari- son of some crop water stress measurement methods. Crop Sci., 24:112 l-1128.
Passioura, J.B., 1986. Resistance to drought and salinity: avenues for improvement. Aust. J. Plant Physiol., 13:191-201.
Rasmusson, D.C., 1991. A plant breeder's experience with ideotype breeding. Field Crops Res., 26: 191-200.
Rosielle, A.A. and Hamblin, J., 1981. Theoretical aspects of selection for yield in stress and non-stress environments. Crop Sci., 21: 943-946.
Sobrado, M.A., 1986. Tissue water relations and leaf growth of tropical corn cultivars under water deficits. Plant Cell Environ., 9:451-457.
Turner, N.C., 1981. Techniques and experimental approaches for the measurement of plant water status. Plant Soil, 58: 339-366.
Turner, N.C., 1986. Crop water deficits: a decade of progress. Adv. Agron., 39:1-51. Westgate, M.E. and Boyer, J.S., 1986. Reproduction at low silk and pollen water potentials in
maize. Crop Sci., 26:951-956. Wright, G.C., Smith, R.C.G. and Morgan, J.M., 1983. Differences between two sorghum geno-
types in adaptation to drought stress. III. Physiological responses. Aust. J. Agric. Res., 34: 637-651.
Wolfe, D.W., Henderson, D.W., Hsiao, T.C. and Alvino, A., 1988. Interactive water and nitro- gen effects on senescence of maize. I. Leaf area duration, nitrogen distribution, and yield. Agron. J., 80: 859-864.