effects of moniliformin in presence of cyclohexadepsipeptides on isolated mammalian tissue and cells
TRANSCRIPT

Toxicology in Vitro 20 (2006) 1284–1291www.elsevier.com/locate/toxinvit
EVects of moniliformin in presence of cyclohexadepsipeptideson isolated mammalian tissue and cells
M.R. Kamyar, K. Kouri ¤, P. Rawnduzi, C. Studenik, R. Lemmens-Gruber
Department of Pharmacology and Toxicology, University of Vienna, Pharmacy Centre, Althanstr. 14, A-1090 Vienna, Austria
Received 5 August 2005; accepted 1 March 2006Available online 8 March 2006
Abstract
Secondary metabolites produced by Fusarium spp. including beauvericin, enniatin and moniliformin are mycotoxins identiWed incereal samples. The two cyclohexadepsipeptide mycotoxins beauvericin and enniatin have cytotoxic, antibiotic, insecticidal and iono-phoric properties, while moniliformin primarily acts as a cardiotoxic mycotoxin. In this study, we examined the electromechanical andelectrophysiological eVects of moniliformin and moniliformin with ionophoric mycotoxins on cells (ventricular myocytes, Caco-2 cells)and in multicellular preparations (papillary muscles and terminal ilea of the guinea pig). Additionally, we investigated the inXuence ofmoniliformin on cell homeostasis in absence and presence of the cyclodepsipeptide mycotoxins (ventricular myocytes, Caco-2 cells).Experiments were performed using isometric measurements of contractility, intracellular microelectrode and patch-clamp techniques, andXuorescence imaging. While ionophoric cyclohexadepsipeptides aVect action potential parameters and cell homeostasis, moniliformin didnot change spontaneous rates of activity or cardiac action potentials. Furthermore, moniliformin had no eVect on intracellular concentra-tions of ions and ATP, and did not aVect pH. Moniliformin reduced contractility in papillary muscle, terminal ileum, the aorta and thepulmonary artery. However, moniliformin did not alter beauvericin and enniatin induced eVects. From our studies, we conclude thatmoniliformin is not solely a cardiotoxic secondary metabolite, but also exerts its eVects on smooth muscle. Moreover, there is no synergis-tic relationship between moniliformin and the concurrently produced cyclohexadepsipeptide mycotoxins beauvericin and enniatin.© 2006 Elsevier Ltd. All rights reserved.
Keywords: Moniliformin; Beauvericin; Enniatin; Cyclohexadepsipeptides; Mycotoxins
1. Introduction eVects of these three mycotoxins have been investigated.
Under certain environmental conditions Fusarium spp.concurrently produce several secondary metabolites such asbeauvericin (Krska et al., 1996), enniatins (Herrmann et al.,1996) and moniliformin (Logrieco et al., 1993; Plattner andNelson, 1994; Lew et al., 1996), which can be identiWed incereal samples (Logrieco et al., 2002; Jestoi et al., 2004). Todate, some of the electrophysiological and mechanical
Abbreviations: APA, action potential amplitude; APD20, APD50,APD90, action potential duration at 20-, 50- and 90% repolarization;MRP, membrane resting potential; BEA, beauvericin; ENN, enniatin; f,spontaneous rate of frequency; fc, force of contraction; MON, monilifor-min; Vmax, upstroke velocity
* Corresponding author.E-mail address: [email protected] (K. Kouri).
0887-2333/$ - see front matter © 2006 Elsevier Ltd. All rights reserved.doi:10.1016/j.tiv.2006.03.001
However, despite their natural concurrence, little is knownabout the interactions between moniliformin and beauveri-cin and enniatin.
Beauvericin and enniatins are cyclohexadepsipeptideswith cytotoxic (Ojcius et al., 1991; Calo et al., 2004; Jowet al., 2004), antibiotic (Dobler et al., 1969), and insecticidaleVects (Grove and Pople, 1980; Gupta et al., 1991), whichare probably related to their ionophoric properties(Dorschner and Lardy, 1968; Shemyakin et al., 1969;Ivanov et al., 1973; Ovchinnikov et al., 1974; Prince et al.,1974; Benz, 1978; Ojcius et al., 1991). Experiments in iso-lated ventricular myocytes and unilamellar vesicles havedemonstrated channel-forming activity of beauvericin(Kouri et al., 2003) and enniatins (Kamyar et al., 2004) atlow concentrations. DiVerences in single channel kineticswere observed, dependent on the mycotoxin and the cell

M.R. Kamyar et al. / Toxicology in Vitro 20 (2006) 1284–1291 1285
membrane properties. Besides its ionophoric activity, beau-vericin can also aVect physiological ion channels (Wu et al.,2002; Tang et al., 2005). In isolated terminal ilea and heartmuscles of guinea pigs the two cyclohexadepsipeptidemycotoxins decrease contractility, shorten action potentialduration and depolarize the membrane resting potential(Lemmens-Gruber et al., 2000; Kamyar et al., 2004).
The cytotoxicity for moniliformin has been demon-strated in various cells (Dombrink-Kurtzman et al., 1993;Wu et al., 1995; Reams et al., 1996a). At low doses, chromo-somal aberrations in primary cultures of rat hepatocyteshave been shown (Knasmüller et al., 1997), althoughNorred et al. (1992) could not verify these Wndings. Monili-formin is thought to be a cardiotoxic mycotoxin, but itsactions and mechanisms remain controversial (Reams et al.,1996a,b, 1997; Gathercole et al., 1986; Chen et al., 1990;Engelhardt et al., 1989; Fan et al., 1991; Zhang and Li, 1989;Nagaraj et al., 1996). Electrocardiographic alterations wereinterpreted as result of altered myocardial metabolism.Thus, we studied whether moniliformin directly aVects elec-trophysiological properties. Since the gastrointestinal tractis the Wrst organ contacted by ingested mycotoxins, we alsotested the eVects of moniliformin, beauvericin and enniatinon isolated terminal ilea of guinea pigs and Caco-2 cells.
Studies with combined mycotoxin administration arerare. Additive or less than additive toxicity was found whenchicks were fed diets containing moniliformin and aXatox-ins (Kubena et al., 1997). A sudden death syndrome wasobserved in chicks and poults with moniliformin, irrespec-tive of the presence or absence of the sodium ionophoremonensin (Reams et al., 1996a). However, no in vitro dataare available about combined eVects of the concurrentlyidentiWed mycotoxins moniliformin, beauvericin and ennia-tin. In this study, we address the relationship between thethree concurrent secondary metabolites of Fusarium spp.
2. Material and methods
2.1. Chemicals
Beauvericin (BEA), enniatin (ENN) and moniliformin(MON) were supplied by Sigma–Aldrich GmbH, Vienna,Austria. While MON is water-soluble, BEA and ENN arepoorly soluble in water, therefore a stock solution withTween 80: MeOHD1:2 (v:v) was prepared. This solutionwas further diluted with the appropriate solution andadded to the bathing solution at the Wnal concentration.
Fluorescent dyes and Pluronic were procured fromMolecular Probes (Leiden, The Netherlands). All otherchemicals were purchased from Sigma (Sigma–AldrichGmbH, Vienna, Austria).
2.2. Isometric measurement of contractility and spontaneous activity
The eVect of the compounds was studied in isolated ter-minal ilea and heart preparations of the guinea pig (Insti-
tute for Laboratory Animal Science and Genetics, MedicalUniversity of Vienna, A-2325 Himberg, Austria). The ani-mals (either sex, 320–430 g) were kept in automatically con-trolled temperature conditions (23§2 °C) in 12 h dark-lightcycles, with food and water “ad libitum”. Twenty-fourhours prior to the experiment, they were deprived of food.The guinea pigs were killed by a blow on the neck. Afterquick removal, the intestine was cleaned by Xushing withnutrient solution, and then it was placed in oxygenatednutrient solution. Spontaneously beating right atria, papil-lary muscles from the right ventricle, and rings of terminalilea (1 cm) were dissected from the guinea pig and immersedinto a Krebs–Henseleit bathing solution (37 °C, 95%O2–5%CO2). The force of contraction was measured isometri-cally with a force transducer (Transbridge™, 4-ChannelTransducer AmpliWer, Sarasota, FL, USA). The papillarymuscles were stimulated with rectangular pulses of 3 msduration 10% above threshold voltage at a frequency of1 Hz. The terminal ilea were precontracted with 60 mM KClcontaining nutrient solution, and the aorta and pulmonaryartery with 90 mM KCl containing nutrient solution. Rest-ing tension was set at 3.9 mN for papillary muscles, 10.9 mNfor right atria, 4.9 mN for terminal ilea, 19.6 mN for aortaand 9.8 mN for pulmonary artery in order to allow for thedevelopment of maximal contractility. After a controlperiod of 30 min, the test compound was added cumula-tively every 40 min until a steady state was reached. Signalswere recorded with a chart recorder (BD 112 Dual Chan-nel, Kipp & Zonen, NL).
2.3. Intracellular microelectrode technique
Papillary muscles were Wxed in a lucite chamber andsuperfused with modiWed Tyrode’s solution (in mM: NaCl136.9, KCl 5.4, MgCl2 1.05, NaHCO3 11.9, NaH2PO4 0.42,CaCl2 1.8, glucose 5.55; pH at 7.2–7.4; 37 °C; gassed with95%O2–5%CO2). Preparations were stimulated with rect-angular pulses of 1 ms duration 10% above threshold volt-age at a constant frequency of 1 Hz. Action potentialswere recorded diVerentially. Glass microelectrodes with aresistance of 10–20 M� were Wlled with 3 mM KCl solu-tion. Signals were electrically diVerentiated to measure themaximum rate of rise of the action potential. For record-ing, two microprobe systems (Model M 701, WP-Instru-ments, Hamden, CT, USA) were used. The signal wasWltered at 10 kHz and sampled at 2 kHz. Data acquisitionwas carried out with the pCLAMP 6 software (AxonInstruments).
2.4. Patch clamp experiments on ventricular myocytes
Electrical activity was studied on freshly isolated ventric-ular myocytes of guinea pigs. The heart was excised andcannulated to a LangendorV apparatus via the aorta. Cellswere isolated by retrograde perfusion with a protease/colla-genase containing modiWed Tyrode’s solution, and theywere stored in modiWed Tyrode’s solution.

1286 M.R. Kamyar et al. / Toxicology in Vitro 20 (2006) 1284–1291
Electrodes pulled from borosilicate capillaries had a tipresistance of 5–10 M�. Potassium currents were recordedwith the patch clamp technique in the inside-out mode(Hamill et al., 1981) under symmetrical conditions regard-ing pipette and bathing solutions. The used pipette andbathing solutions contained 140 mM potassium aspartate,2 mM CaCl2, 2 mM MgCl2, 2 mM Na2ATP, 10 mMHEPES, titrated to pH 7.4 with KOH. Potassium currentsoriginated when the concentration of Na2ATP in the bath-ing solution was reduced. All experiments were performedat room temperature (19–22 °C). Five minutes after forma-tion of an inside-out patch, the control period of 10 minstarted before MON (10, 30, 100 and 300�M) was added.Experiments lasted at least 30 min.
Electrophysiological measurements were carried outwith an Axopatch-1D patch clamp ampliWer (Axon Instru-ments, CA) at a cut-oV frequency (¡3 dB) of 2 kHz. Cur-rents were Wltered at 5kHz with a dual variable Wlter (VBF 8,Kemo Ltd, Beckenham, Kent, UK), digitized via an ADconverter (TL-1 interface, Axon Instruments, CA) andsampled at 5–10 kHz. Data acquisition and storage wereprocessed directly to a PC equipped with pCLAMP 6 soft-ware (Axon Instruments, CA). Single channel analysis wasperformed with the ASCD software (G. Droogmans,Leuven, Belgium).
2.5. Fluorescence ratio imaging
Ventricular myocytes and Caco-2 cells were loaded inmodiWed Tyrode’s solution with either 1–25 �M of the Ca2+
indicator FURA 2AM, 0.5–25�M of the Xuorescent Na+
indicator SBFI AM, 0.5–25 �M of the K+ indicator PBFIAM, or 0.5–20 �M of the Mg2+ indicator MagFura 2AM,and an equivalent concentration of Pluronic 20% DMSO atroom temperature (19–22 °C) for 20–30 min. After washout,cells were allowed to hydrolyze the dye for 30 min to 1 hbefore they were added to the bath of the set-up. After acontrol period of 10 min, the test compounds were added tothe bathing solution. Control experiments with the appro-priate amount of solvent were performed additionally.Intracellular pH was measured in cardiac myocytes andCaco-2 cells using the pH-sensitive Xuorescent dye BCECFAM. Cells were loaded with 1–2.5 �M dye for 30 min. Cali-bration of pH was performed with 50 �M nigericin andhigh K+ Tyrode’s solutions with pH ranging from 5 to 7.5.
The bath was located on the stage of an inverted epi-Xuorescence microscope (Axiovert 100, Zeiss) equippedwith a Fluar £40/1.30 NA oil immersion objective. Excit-atory light was provided by a xenon arc lamp light source.The dyes were excited alternatively at 340 nm and 380 nmwavelengths (Omega Optical, Inc., VT) by means of anoptical Wlter changer (Lambda 10-2, Sutter Instruments,CA). Emitted light was Wltered at 510 nm bandpass andacquired by an intensiWed CCD camera (Extended ISIS,Photonic Science Ltd, UK) connected to a PC through aninterface card (Axon Image Lightning). Ratiometric mea-surements were realized following background substractionwith the Axon Imaging Workbench 2.2 software (AxonInstruments, CA) averaging two frames. Results are pre-sented as the F340/F380 signal ratio. For the detailed experi-mental protocol see Kouri et al. (2005).
2.6. Statistical analysis
Values are expressed as means§S.E. of n experiments.SigniWcance was calculated with the Student’s t-test forpaired observations. Statistical signiWcance was determinedat a level of p < 0.05.
3. Results
3.1. Electromechanical eVects of MON
Under our experimental conditions in papillary musclesof guinea pigs, the force of contraction (fc) was reduced in aconcentration-dependent manner by MON with an IC50 of78§13 �M (nD3). However, the rate of activity of sponta-neously beating right atria (nD 3) was not changed in con-centrations of MON up to 100 �M. At 300 �M, a small, butnot signiWcant decrease in activity rate was observed,whereas fc was markedly reduced in the right atria(IC50D 30§ 8 �M) (Table 1).
The eVect on contractility of vascular smooth muscleswas weaker than in heart muscle preparations. The decreaseof fc at 300�M MON was 40§ 4% in the aorta (nD 3), andonly 18§3% in the pulmonary artery (nD3). The reductionof fc was found to occur in a similar concentration range(IC50: 69§9 �M, nD 3) as in papillary muscles (Table 1).
The decrease in fc was independent of extracellular cal-cium concentrations in the range of 3.2 to 10.4 mM CaCl2
Table 1IC50-values (�M) for eVects on contractility (fc) and spontaneous rate of activity (f)
Data for BEA, ENN and MON alone as well as for MON in the presence of BEA or ENN are presented; n D 3 for each preparation and each mycotoxin(combination). In papillary muscles and terminal ilea the IC50-values of MON are not signiWcantly diVerent (p > 0.05) from the IC50-values of MON inpresence of BEA and ENN.
Mycotoxin Papillary muscle (fc) Terminal ileum (fc) Right atrium (fc/f)
ENN 8 § 1 3 § 0.3 7 § 2/2 § 0.4BEA 18§ 4 1 § 0.2 0.5 § 0.2/>30MON 78§ 13 69 § 9 30§ 8/>300MON (BEA) 70§ 11 50 § 11 –MON (ENN) 85§ 9 76 § 2 –

M.R. Kamyar et al. / Toxicology in Vitro 20 (2006) 1284–1291 1287
in terminal ilea (nD 6), as well as in papillary muscles(nD 7). In addition, we exposed MON-treated preparationsto glibenclamide, a blocker of the ATP-sensitive potassiumchannel. The MON-induced reduction of fc could not beantagonized by glibenclamide in smooth and heart musclepreparations (nD 2 and nD3, respectively).
3.2. Electrophysiological eVects of MON
The eVects of MON on the action potential of isolatedguinea pig papillary muscles were studied at 10 (nD3), 30(nD 3) and 100�M (nD3). The membrane resting potential,action potential amplitude, upstroke velocity, and actionpotential duration were not aVected signiWcantly by MONin papillary muscles (Table 2). In spontaneously beatingPurkinje Wbres maximum diastolic potential, action poten-tial amplitude, upstroke velocity, action potential duration,slope of slow diastolic depolarization and rate of activitywere not changed by 100 �M MON (nD3) as well (Fig. 1).
It was previously reported that MON may interfere withthe cellular energy production (Nagaraj et al., 1996). Dur-ing depletion of intracellular ATP, an ATP-sensitive potas-sium channel is opened, resulting in a shortening of actionpotential duration. However, even at a concentration of300 �M, MON did not have any eVects on open probability,current amplitude, and single channel kinetics of the ATP-dependent potassium current in inside-out patches of ven-tricular myocytes (nD3).
3.3. EVects of MON on intracellular ion concentrations
For studying the eVect of MON on intracellular calciumconcentrations, the calcium ionophore ionomycin (50 �M,nD3) was used as a reference compound. Although MONexerted a negative inotropic eVect in isolated papillary mus-cles and a decrease of fc in terminal ilea, it did not changethe intracellular calcium concentration in cardiomyocytesand Caco-2 cells. The eVect was absent at the high concen-tration of 300 �M MON and in extended incubation peri-ods of up to 45 min (nD3) (Fig. 2). Furthermore, MONalso did not change the intracellular sodium and potassiumconcentrations of Caco-2 cells (100 �M, nD2).
Table 2EVects of MON on papillary muscle action potential
No signiWcant diVerences (p > 0.05) between the control values and thevalues for each parameter at 30 and 100 �M MON were estimated.Abbreviations: action potential (AP), membrane resting potential (MRP),action potential amplitude (APA), upstroke velocity (Vmax), action poten-tial duration at 20-, 50-, and 90% repolarization (APD20, APD50, APD90);n D 3.
AP parameter Control 30 �M MON 100 �M MON
MRP (mV) 88.1 § 2.4 90.0 § 2.2 87.3 § 2.0APA (mV) 115.1 § 4.0 112.5 § 4.6 116.6 § 4.8Vmax 217.5 § 10.6 222.5 § 10.7 220.4 § 17.5APD20 79.3 § 8.1 77.3 § 0.5 81.9 § 6.1APD50 138.6 § 8.7 127.6 § 11.0 142.6 § 9.0APD90 157.8 § 11.1 144.5 § 12.5 170.3 § 9.2
3.4. EVects of MON on cellular ATP concentrations and intracellular pH
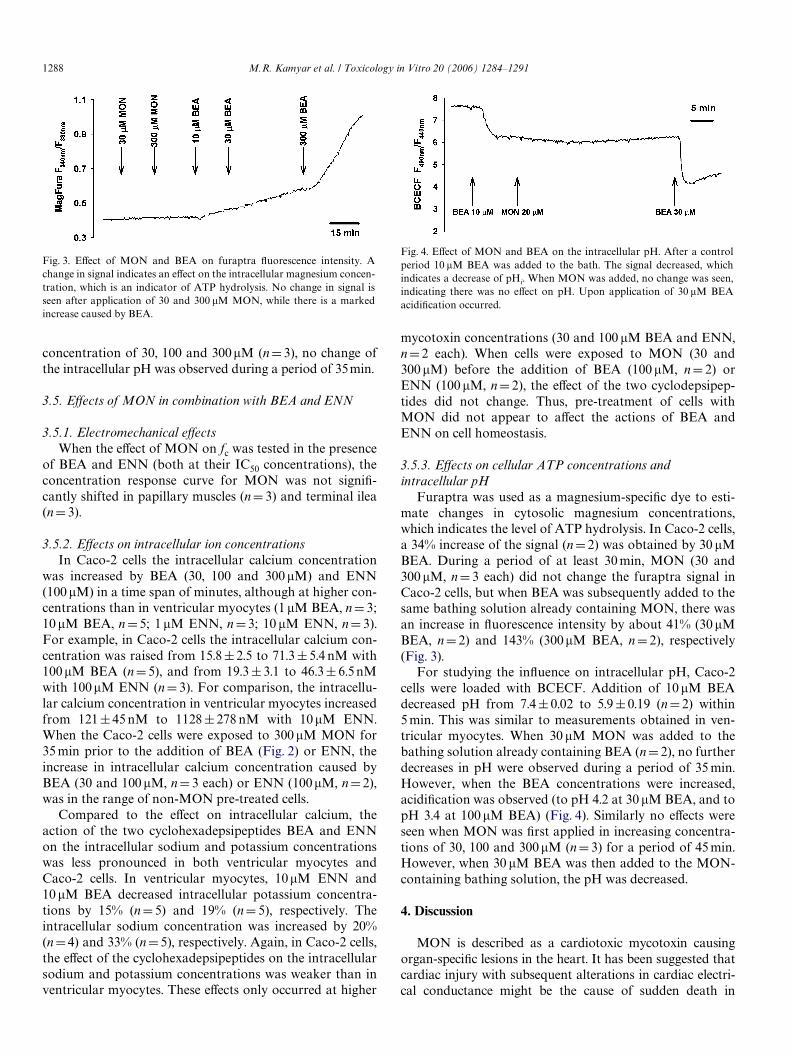
When ATP synthesis is limited, an increase in intracellu-lar magnesium concentration occurs due to the unbindingof ATP to magnesium (Leyssens et al., 1996). MagFura 2AM (furaptra) was used as a magnesium-speciWc dye toestimate changes in its cytosolic concentration. MON (30and 300�M, nD3 each) did not change the furaptra signalin cardiomyocytes and Caco-2 cells (Fig. 3), indicating thatsigniWcant hydrolysis of ATP did not occur.
Caco-2 cells were loaded with BCECF to study theeVect on intracellular pH. When MON was added at a
Fig. 1. Original recordings of action potentials (upper beam) and Vmax(lower beam) from an isolated spontaneously beating guinea pig PurkinjeWbre during control (A) and 60 min after addition of 100 �M MON (B).
Fig. 2. The eVect of MON and BEA on the intracellular calcium concen-tration of Caco-2 cells. The addition of 300 �M MON and 100 �M BEA isindicated with arrows.
1288 M.R. Kamyar et al. / Toxicology in Vitro 20 (2006) 1284–1291
concentration of 30, 100 and 300�M (nD3), no change ofthe intracellular pH was observed during a period of 35min.
3.5. EVects of MON in combination with BEA and ENN
3.5.1. Electromechanical eVectsWhen the eVect of MON on fc was tested in the presence
of BEA and ENN (both at their IC50 concentrations), theconcentration response curve for MON was not signiW-cantly shifted in papillary muscles (nD 3) and terminal ilea(nD3).
3.5.2. EVects on intracellular ion concentrationsIn Caco-2 cells the intracellular calcium concentration
was increased by BEA (30, 100 and 300�M) and ENN(100 �M) in a time span of minutes, although at higher con-centrations than in ventricular myocytes (1 �M BEA, nD 3;10 �M BEA, nD5; 1 �M ENN, nD3; 10 �M ENN, nD 3).For example, in Caco-2 cells the intracellular calcium con-centration was raised from 15.8§2.5 to 71.3§5.4 nM with100�M BEA (nD5), and from 19.3§3.1 to 46.3§ 6.5 nMwith 100�M ENN (nD 3). For comparison, the intracellu-lar calcium concentration in ventricular myocytes increasedfrom 121§45 nM to 1128§278 nM with 10�M ENN.When the Caco-2 cells were exposed to 300 �M MON for35 min prior to the addition of BEA (Fig. 2) or ENN, theincrease in intracellular calcium concentration caused byBEA (30 and 100 �M, nD 3 each) or ENN (100�M, nD2),was in the range of non-MON pre-treated cells.
Compared to the eVect on intracellular calcium, theaction of the two cyclohexadepsipeptides BEA and ENNon the intracellular sodium and potassium concentrationswas less pronounced in both ventricular myocytes andCaco-2 cells. In ventricular myocytes, 10�M ENN and10 �M BEA decreased intracellular potassium concentra-tions by 15% (nD5) and 19% (nD 5), respectively. Theintracellular sodium concentration was increased by 20%(nD4) and 33% (nD5), respectively. Again, in Caco-2 cells,the eVect of the cyclohexadepsipeptides on the intracellularsodium and potassium concentrations was weaker than inventricular myocytes. These eVects only occurred at higher
Fig. 3. EVect of MON and BEA on furaptra Xuorescence intensity. Achange in signal indicates an eVect on the intracellular magnesium concen-tration, which is an indicator of ATP hydrolysis. No change in signal isseen after application of 30 and 300 �M MON, while there is a markedincrease caused by BEA.
mycotoxin concentrations (30 and 100 �M BEA and ENN,nD2 each). When cells were exposed to MON (30 and300 �M) before the addition of BEA (100 �M, nD 2) orENN (100 �M, nD 2), the eVect of the two cyclodepsipep-tides did not change. Thus, pre-treatment of cells withMON did not appear to aVect the actions of BEA andENN on cell homeostasis.
3.5.3. EVects on cellular ATP concentrations and intracellular pH
Furaptra was used as a magnesium-speciWc dye to esti-mate changes in cytosolic magnesium concentrations,which indicates the level of ATP hydrolysis. In Caco-2 cells,a 34% increase of the signal (nD2) was obtained by 30 �MBEA. During a period of at least 30 min, MON (30 and300 �M, nD 3 each) did not change the furaptra signal inCaco-2 cells, but when BEA was subsequently added to thesame bathing solution already containing MON, there wasan increase in Xuorescence intensity by about 41% (30 �MBEA, nD 2) and 143% (300�M BEA, nD 2), respectively(Fig. 3).
For studying the inXuence on intracellular pH, Caco-2cells were loaded with BCECF. Addition of 10 �M BEAdecreased pH from 7.4§ 0.02 to 5.9§0.19 (nD2) within5 min. This was similar to measurements obtained in ven-tricular myocytes. When 30 �M MON was added to thebathing solution already containing BEA (nD 2), no furtherdecreases in pH were observed during a period of 35 min.However, when the BEA concentrations were increased,acidiWcation was observed (to pH 4.2 at 30 �M BEA, and topH 3.4 at 100 �M BEA) (Fig. 4). Similarly no eVects wereseen when MON was Wrst applied in increasing concentra-tions of 30, 100 and 300�M (nD 3) for a period of 45 min.However, when 30 �M BEA was then added to the MON-containing bathing solution, the pH was decreased.
4. Discussion
MON is described as a cardiotoxic mycotoxin causingorgan-speciWc lesions in the heart. It has been suggested thatcardiac injury with subsequent alterations in cardiac electri-cal conductance might be the cause of sudden death in
Fig. 4. EVect of MON and BEA on the intracellular pH. After a controlperiod 10 �M BEA was added to the bath. The signal decreased, whichindicates a decrease of pHi. When MON was added, no change was seen,indicating there was no eVect on pH. Upon application of 30 �M BEAacidiWcation occurred.

M.R. Kamyar et al. / Toxicology in Vitro 20 (2006) 1284–1291 1289
chronically poisoned poultry (Reams et al., 1997). Indeed,cardiotoxicity with ultrastructural damage (Zhao et al., 1993;Bermudez et al., 1997; Morgan et al., 1998, 1999) consists ofdegeneration and necrosis of the heart (Engelhardt et al.,1989), and the development of myocardial hypertrophy(Reams et al., 1996b, 1997). The mechanism underlyingMON toxicity is described as inhibition of mitochondrialpyruvate and �-ketoglutarate oxidation with subsequentreduction of oxidative phosphorylation (Thiel, 1978). Reamset al. (1996b) did not observe any eVects of MON on therate and the force of atrial contraction in guinea pigs. Thus,they attributed hypertrophy to MON-induced hypoxia andaltered functions of cardiac cell enzymes rather than in termsof direct cardiostimulation. In contrast, Fan et al. (1991)observed a decrease in myocardial contractile force in rats,and ventricular arrhythmia. This latter eVect was alsoreported by Zhang in ducks (1989). Additionally, electrocar-diograph measurements on broiler chickens have beenshown to be inXuenced by MON (Nagaraj et al., 1996).
However, no direct evidence of the electrophysiologicaleVects of MON was available. No data about the eVect onaction potential parameters are published for MON,although MON is reported to be a cardiotoxic mycotoxin.Thus, we tested the action of MON on the cardiac actionpotential of isolated papillary muscles and Purkinje Wbresof the guinea pig. No signiWcant eVects on action potentialparameters were observed on papillary muscles and Pur-kinje Wbres up to a high concentration of 300 �M of MON.Furthermore, the rate of activity of spontaneously beatingPurkinje Wbres was not changed, and no early or delayedafter-depolarizations were found from which extrasystolescould originate. Therefore, our experiments support thehypothesis that altered cardiac metabolism is the maincause of MON-induced lesions, and this deWcit manifestsinto electrophysiological disorders such as arrhythmia.
Intracellular sodium, potassium and calcium concentra-tions were not inXuenced by MON. These observations cor-roborate our Wndings that MON does not alter maximumupstroke velocity, or action potential amplitude and dura-tion in isolated papillary muscles and Purkinje Wbres. Thus,a direct interaction of MON with ion channels and subse-quent changes in cell homoestasis can be excluded as thecause of its cardiac eVects. Since MON did not changeintracellular calcium concentrations, its interaction with anintermediate component of calcium release is also unlikelyto be the reason for negative inotropy. However, the nega-tive inotropic eVect and reduction of fc in smooth musclepreparations was evident. These Wndings illustrate thateVects on contractility are not induced by electrophysiolog-ical eVects of MON, but are due to morphological changesas previously suggested by Engelhardt et al. (1989), Zhaoet al. (1993), Reams et al. (1997), Bermudez et al. (1997) andMorgan et al. (1998, 1999).
It has been proposed that MON may interfere with cellu-lar energy production and induce hypoxia (Nagaraj et al.,1996). Upon decrease of the intracellular ATP concentrationan ATP-sensitive potassium channel is opened which results
in a shortening of the action potential duration. We did notsee such an eVect on the repolarization phase of actionpotentials recorded from papillary muscles and PurkinjeWbres. ATP hydrolysis, as was revealed by Xuorescence imag-ing, was not aVected, and ATP-sensitive potassium channelswere not inXuenced by MON. Thus, our Wndings agree withdata reported by Reams et al. (1996a), who found no eVectsof MON on ATP production in myoblasts.
Additionally, our experiments show that eVects of MONare not organ-speciWc, since the reduction of fc occurred atthe same concentration range in both papillary muscles andterminal ilea. Furthermore, a decrease of fc was also foundin the aorta and pulmonary artery, although this eVect wasof a lesser magnitude.
BEA, ENNs and MON are Fusarium mycotoxins thatare concurrently identiWed in cereal samples (Logriecoet al., 2002; Jestoi et al., 2004). This raises the question as towhether the mycotoxins might interact. Due to the iono-phoric properties of the two cyclohexadepsipeptides, aneVect on cell homeostasis is likely. This action is accompa-nied by ATP hydrolysis and acidiWcation, which ultimatelycontributes to cell death. MON itself did not show theseeVects in isolated cells, but is reported to interfere with cellmetabolism (Reams et al., 1996a). Although MON signiW-cantly decreases fc in isolated muscle preparations, none ofthe eVects of the cyclodepsipeptides BEA and ENN wereampliWed. The concentration response curves for the eVectof BEA and ENN on contractility were not shifted byMON, and the BEA- and ENN-induced changes in cellhomeostasis were not aVected in presence of MON. TheseWndings support the results reported by Reams et al.(1996a), who found that MON does not predispose L6myoblasts to monensin toxicosis when studied in combina-tion with the sodium ionophore. BEA and ENN are notselective sodium ionophores, but also conduct potassium,calcium and magnesium ions. These Wndings suggest thatMON-induced eVects are apparently not modulated by ion-ophore mycotoxins.
From our studies, we concluded that MON (i) is not aspeciWc cardiotoxic secondary metabolite, but also aVectssmooth muscle preparations, (ii) aVects contractility only atrather high concentrations, (iii) exerts no electrophysiologi-cal eVects, (iv) does not inXuence intracellular ion concen-trations, ATP and pH and (v) does not act synergisticallywith concurrently occurring cyclohexadepsipeptide myco-toxins BEA and ENN.
Acknowledgement
This work and M.R. Kamyar were supported by theAustrian Science Fund (FWF project P16045-B05).
References
Benz, R., 1978. Alkali ion transport through lipid bilayer membranes med-iated by enniatin A and B and beauvericin. Journal of Membrane Biol-ogy 43, 367–394.

1290 M.R. Kamyar et al. / Toxicology in Vitro 20 (2006) 1284–1291
Bermudez, A.J., Ledoux, D.R., Rottinghaus, G.E., Bennett, G.A., 1997. Theindividual and combined eVects of the Fusarium mycotoxins monilifor-min and fumonisin B1 in Turkeys. Avian Diseases 41, 304–311.
Calo, L., Fornelli, F., Ramires, R., Nenna, S., Tursi, A., CaiaVa, M.F.,Macchia, L., 2004. Cytotoxic eVects of the mycotoxin beauvericin tohuman cell lines of myeloid origin. Pharmacogical Research 49, 73–77.
Chen, L.Y., Tian, X.L., Yang, B., 1990. A study on the inhibition of ratmyocardium glutathione peroxidase and glutathione reductase bymoniliformin. Mycopathologia 110, 119–124.
Dobler, M., Dunitz, J.D., Krajewski, J., 1969. Structure of the K+ complexwith enniatin B, a macrocyclic antibiotic with K+ transport properties.Journal of Molecular Biology 28, 603–606.
Dombrink-Kurtzman, M.A., Javed, T., Bennett, G.A., Richard, J.L.,Cote, L.M., Buck, W.B., 1993. Lymphocyte cytotoxicity and erythro-cyte abnormalities induced in broiler chicks by fumonisins B1 and B2and moniliformin from Fusarium proliferatum. Mycopathologia 124,47–54.
Dorschner, E., Lardy, H., 1968. SpeciWcity of ion transport induced bybeauvericin. Antimicrobial Agents and Chemotherapy 8, 11–14.
Engelhardt, J.A., Carlton, W.W., Tuite, J.F., 1989. Toxicity of Fusariummoniliforme var. subglutinans for chicks, ducklings, and Turkey poults.Avian Diseases 33, 357–360.
Fan, L.L., Li, J., Sun, L.H., 1991. EVect of moniliformin on myocardialcontractility in rats. Biomedical and Environmental Sciences 4, 290–294.
Gathercole, P.S., Thiel, P.G., Hofmeyr, J.H., 1986. Inhibition of pyruvatedehydrogenase complex by moniliformin. Biochemical Journal 233,719–723.
Grove, J.F., Pople, M., 1980. The insecticidal activity of beauvericin andthe enniatin complex. Mycopathologia 70, 103–105.
Gupta, S., KrasnoV, S.B., Underwood, N.L., Renwick, J.A.A., Roberts,D.W., 1991. Isolation of beauvericin as an insect toxin from Fusariumsemitectum and Fusarium moniliforme var. subglutinans. Mycopatholo-gia 115, 185–189.
Hamill, O.P., Marty, A., Neher, E., Sakmann, B., Sigworth, F.J., 1981.Improved patch-clamp techniques for high-resolution current record-ing from cells and cell-free membrane patches. PXügers Archiv 391, 85–100.
Herrmann, M., Zocher, R., Haese, A., 1996. EVect of disruption of theenniatin synthetase gene on the virulence of Fusarium avenaceum.Molecular Plant–Microbe Interactions 9, 226–232.
Ivanov, V.T., Evstatov, A.V., Sumskaya, L.V., Melnik, E.I., Chumbridze,T.S., Portnova, S.L., Balashova, T.A., Ovchinnikov, Y.A., 1973. Sand-wich complexes as a functional form of the enniatin ionophores. FEBSLetters 36, 65–71.
Jestoi, M., Rokka, M., Yli-Mattila, T., Parikka, P., Rizzo, A., Peltonen, K.,2004. Presence and concentrations of the Fusarium-related mycotoxinsbeauvericin, enniatins and moniliformin in Wnnish grain samples. FoodAdditives and Contaminants 21, 794–802.
Jow, G.M., Chou, C.J., Chen, B.F., Tsai, J.H., 2004. Beauvericin inducescytotoxic eVects in human acute lymphoblastic leukemia cells throughcytochrome c release, caspase 3 activation: the causative role of cal-cium. Cancer Letters 216, 165–173.
Kamyar, M.R., Rawnduzi, P., Studenik, C., Kouri, K., Lemmens-Gruber,R., 2004. Investigation of the electrophysiological properties of ennia-tin. Archives of Biochemistry and Biophysics 429, 215–223.
Knasmüller, S., Bresgen, N., Kassie, F., Mersch-Sundermann, V., Gelderblom,W., Zohrer, E., Eckl, P.M., 1997. Genotoxic eVects of three Fusariummycotoxins, fumonisin B1, moniliformin and vomitoxin in bacteriaand primary cultures of rat hepatocytes. Mutation Research 391,39–48.
Krska, R., Schuhmacher, R., Grasserbauer, M., Scott, P.M., 1996. Determi-nation of the Fusarium mycotoxin beauvericin at micrograms/kg levelsin corn by high-performance liquid chromatography with diode-arraydetection. Journal of Chromatography A 746, 233–238.
Kouri, K., Lemmens, M., Lemmens-Gruber, R., 2003. Beauvericin inducedchannels in ventricular myocytes and artiWcial membranes. Biochimicaet Biophysica Acta - Biomembranes 1609, 203–210.
Kouri, K., Duchen, M., Lemmens-Gruber, R., 2005. EVects of beauvericinon the metabolic state and ionic homeostasis of ventricular myocytesof the guinea pig. Chemical Research in Toxicology 18, 1661–1668.
Kubena, L.F., Harvey, R.B., Buckley, S.A., Edrington, T.S., Rottinghaus,G.E., 1997. Individual and combined eVects of moniliformin present inFusarium fujikuori culture material and aXatoxin in broiler chicks.Poultry Science 76, 265–270.
Lemmens-Gruber, R., Rachoy, B., Steininger, E., Kouri, K., Saleh, P.,Krska, R., Josephs, R., Lemmens, M., 2000. The eVect of the Fusariummetabolite beauvericin on electromechanical and physiological proper-ties in isolated smooth and heart muscle preparations of guinea pigs.Mycopathologia 149, 5–12.
Lew, H., Chelkowski, J., Pronczuk, P., Edinger, W., 1996. Occurrence of themycotoxin moniliformin in maize (Zea mays L.) ears infected by Fusar-ium subglutinans (Wollenw & Reinking) Nelson et al. Food Additivesand Contaminants 13, 321–324.
Leyssens, A., Nowicky, A.V., Patterson, L., Crompton, M., Duchen, M.R.,1996. The relationship between mitochondrial state, ATP hydrolysis,[Mg2+]i and [Ca2+]i studied in isolated rat cardiomyocytes. Journal ofPhysiology 496, 111–128.
Logrieco, A., Moretti, A., Altomare, C., Bottalico, A., Carbonell Torres, E.,1993. Occurrence and toxicity of Fusarium subglutinans from Peruvianmaize. Mycopathologia 122, 185–190.
Logrieco, A., Rizzo, A., Ferracane, R., Ritieni, A., 2002. Occurrence ofbeauvericin and enniatins in wheat aVected by Fusarium avenaceumhead blight. Applied Environmental Microbiology 68, 82–85.
Morgan, M.K., Bursian, S.J., Rottinghaus, G.E., Bennett, G.A., Render,J.A., Aulerich, R.J., 1998. Subacute and reproductive eVects in minkfrom exposure to Fusarium fujikuori culture material (M-1214) con-taining known concentrations of moniliformin. Archives of Environ-mental Contamination and Toxicology 35, 513–517.
Morgan, M.K., Fitzgerald, S.D., Rottinghaus, G.E., Bursian, S.J., Aulerich,R.J., 1999. Toxic eVects to mink of moniliformin extracted from Fusar-ium fujikuori culture material. Veterinary and Human Toxicology 41,1–5.
Nagaraj, R.Y., Wu, W., Will, J.A., Vesonder, R.F., 1996. Acute cardiotoxic-ity of moniliformin in broiler chickens as measured by electrocardio-graphy. Avian Diseases 40, 223–227.
Norred, W.P., Plattner, R.D., Vesonder, R.F., Bacon, C.W., Voss, K.A.,1992. EVects of selected secondary metabolites of Fusarium monili-forme on unscheduled synthesis of DNA by rat primary hepatocytes.Food and Chemical Toxicology 30, 233–237.
Ojcius, D.M., Zychlinsky, A., Zheng, L.M., Young, J.D., 1991. Ionophore-induced apoptosis: a role of DNA fragmentation and calcium Xuxes.Experimental Cell Research 197, 43–49.
Ovchinnikov, Y.A., Ivanov, V.T., Evstranov, A.V., Mikhaleva, I.I., Bystrov,V.F., Portnova, S.L., Balashova, T.A., Meshcheryakova, E.N., Tulchin-sky, V.M., 1974. The enniatin ionophores. Conformation and ion bind-ing properties. International Journal of Peptide and Protein Research6, 465–498.
Plattner, R.D., Nelson, P.E., 1994. Production of beauvericin by a strain ofFusarium proliferatum isolated from corn fodder for swine. Appliedand Environmental Microbiology 60, 3894–3896.
Prince, R.C., Crofts, A.R., Steinrauf, L.K., 1974. A comparison of beau-vericin, enniatin and valinomycin as calcium transporting agents in lipo-somes and chromatophores. Biochemical and Biophysical ResearchCommunications 59, 697–703.
Reams, R., Thacker, H.L., Novilla, M., Laska, D., Horn, J., Harrington, D.,Greenlee, W., Vesonder, R., 1996a. Development of an L6 myoblastin vitro model of moniliformin toxicosis. Mycopathologia 133, 105–114.
Reams, R.Y., Thacker, H.L., Harrington, D.D., Novilla, M.N., Wilson, B.,1996b. PuriWed moniliformin does not aVect the force or rate of con-traction of isolated guinea pig atria. Mycopathologia 133, 115–118.
Reams, R.Y., Thacker, H.L., Harrington, D.D., Novilla, M.N., Rotting-haus, G.E., Bennett, G.A., Horn, J., 1997. A sudden death syndromeinduced in poults and chicks fed diets containing Fusarium fujikuroiwith known concentrations of moniliformin. Avian Diseases 41, 20–35.

M.R. Kamyar et al. / Toxicology in Vitro 20 (2006) 1284–1291 1291
Shemyakin, M.M., Ovchinnikov, Y.A., Ivanov, V.T., Antonov, V.K.,Vinogradova, E.I., Shkrob, A.M., Malenkov, G.G., Evstratov, A.V.,Laine, I.A., Melnik, E.I., Ryabova, I.D., 1969. Cyclodepsipeptides aschemical tools for studying ionic transport through membranes. Jour-nal of Membrane Biology 1, 402–430.
Tang, C.Y., Chen, Y.W., Jow, G.M., Chou, C.J., Jeng, C.J., 2005. Beauveri-cin activates Ca2+-activated Cl¡ currents and induces cell deaths inXenopus Oocytes via inXux of extracellular Ca2+. Chemical Researchin Toxicology 18, 825–833.
Thiel, P.G., 1978. A molecular mechanism for the toxic action of monili-formin, a mycotoxin produced by Fusarium moniliforme. BiochemicalPharmacology 27, 483–486.
Wu, S.N., Chen, H., Liu, Y.C., Chiang, H.T., 2002. Block of L-type Ca2+
current by beauvericin, a toxic cyclopeptide, in the NG108-15 neuronalcell line. Chemical Research in Toxicology 15, 854–860.
Wu, W., Liu, T., Vesonder, R.F., 1995. Comparative cytotoxicity of fumon-isin B1 and moniliformin in chicken primary cell cultures. Mycopatho-logia 132, 111–116.
Zhang, H., Li, J.L., 1989. Study on toxicological mechanism of monilifor-min. Wei Sheng Wu Hsueh Pao 29, 93–100 (Abstract in English).
Zhao, D., Feng, Q., Yan, X., Li, C., Pan, Y., Cui, Q., 1993. Ultrastructuralstudy of moniliformin induced lesions of myocardium in rats and mice.Biomedical and Environmental Sciences 6, 37–44.