effects of glomus intraradices on the growth and reactive oxygen metabolism of foxtail millet under...
TRANSCRIPT

ORIGINAL ARTICLE
Effects of Glomus intraradices on the growth and reactive oxygenmetabolism of foxtail millet under drought
Minggui Gong & Xiaoyan You & Qiaoming Zhang
Received: 5 September 2013 /Accepted: 2 April 2014# Springer-Verlag Berlin Heidelberg and the University of Milan 2014
Abstract Foxtail millet (Setaria italica L.) is an ancient do-mesticated crop grown in arid and semi-arid Africa and Eur-asian continent. This study investigated the effects ofarbuscular mycorrhizal (AM) fungi (Glomus intraradices)and water stress on the growth and reactive oxygen metabo-lism of foxtail millet under greenhouse pot conditions. Aftertwo soil water availability treatments [well-watered (WW)−0.11 MPa; water-stressed (WS) −0.68 MPa] for 31 days,G. intraradices could colonize well in the roots of foxtailmillet. Water stress inhibited AM colonization, plant growthand crop yield, and also increased reactive oxygenmetabolismand antioxidant enzyme activities of foxtail millet.G. intraradices-inoculated foxtail millet had greater plantheight, collar diameter, panicle heigh, panicle weight andgrain weight, higher enzyme activities of superoxide dismut-ase (SOD), catalase (CAT), peroxidase (POD) and glutathionereductase (GR), and lower concentration of malondialdehyde(MDA) , H2O2, and O2
·-, when compared with non-inoculatedfoxtail millet under both WWand WS conditions. Our resultsdemonstrated that G. intraradices-inoculated foxtail milletshowed higher drought-tolerance by improving their growthand productivity, reactive oxygen species (ROS) and antiox-idant enzyme activities, compared with non-inoculated foxtailmillet. This indicates that G. intraradices inoculation is apotential method for enhancing the performance of foxtailmillet under drought conditions.
Keywords Arbuscular mycorrhizal fungi . Foxtail millet(Setaria italicaL.) . Reactive oxygen species . Antioxidantenzyme .Water stress
Introduction
Drought is a major abiotic stress limiting crop growth andproductivity throughout the world due to climate change(Kramer and Boyer 1995; Omirou et al. 2013). Up to 45%of the agriculture lands on Earth are subjected to continuous orfrequent water deficiency, which cause about 50% loss inaverage grain yield (Celebi et al. 2010; Singh et al. 2012).This worldwide problem is increasingly serious as a conse-quence of global climate changes (Goicoechea et al. 2005).Drought stress results in a series of morphological, physiolog-ical and biochemical damage in plants (Gong et al. 2013).However, plants themselves evolve anti-drought mechanismsto cope with water stress, such as increasing root growth inorder to absorb more water, elevating antioxidant enzymeactivities to scavenge for free radicals, and accumulatingosmotic regulators to reduce osmotic pressure (Huang et al.2011). Plant drought-resistance could be also enhancedthrought other external methods, for example, the applicationof mycorrhizal inoculation.
Arbuscular mycorrhizal (AM) fungi exist in almost allterrestrial ecosystems, and symbiosis with AM fungi is com-monly considered to be highly beneficial for nearly all agri-cultural crops (Smith and Read 1997). Host plants provideAM fungi with soluble carbon sources, and the fungi enhancethe absorption and translocation of nutrient ions and waterfrom soil to plants (Subramanian and Charest 1997; Celebiet al. 2010). Improved adaptation of mycorrhizal plants todrought conditions is linked to several interrelated observa-tions andmechanisms (Omirou et al. 2013). Fungal arbusculesin root cells of host plants are the main sites of water exchange
M. Gong :X. YouCollege of Food and Bioengineering, Henan University of Scienceand Technology, Luoyang, Henan 471023, China
Q. Zhang (*)College of Forestry, Henan University of Science and Technology,Luoyang, Henan 471003, Chinae-mail: [email protected]
Ann MicrobiolDOI 10.1007/s13213-014-0895-y

between AM fungi and plants (Smith and Read 1997), AMexternal hyphae act as bridges between soil particles and plantroots which improve direct water uptake from sources inac-cessible to non-mycorrhizal roots (Porcel et al. 2003). Fur-thermore, mycorrhizal plants increase the activities of antiox-idant enzyme against the generation of reactive oxygen spe-cies (ROS) (Wu et al. 2006). ROS embraces not only freeradicals such as superoxide (O2
·-) and hydroxyl radicals (OH·),but also hydrogen peroxide (H2O2) and singlet oxygen (1O2),and these molecules often cause oxidative damage to biomol-ecules such as lipid peroxidation, protein denaturation andDNA mutation in plant cells (Porcel et al. 2003). Inductionof antioxidant enzymes [i.e., superoxide dismutase (SOD, EC1.15.1.1), catalase (CAT, EC 1.11.1.6), peroxidase (POD, EC1.11.1.9) and glutathione reductase (GR, EC 1.6.4.2)] is themost commonmechanism for detoxifying ROS synthesized inplant cellular and aerobic metabolisms during stress responses(Asada 1999; Lambais et al. 2003).
Increasing the drought tolerance of crop would be thecrucial economical approach to improve agricultural produc-tivity and to reduce the agricultural use of fresh water re-sources (Singh et al. 2012). Hence, an improved understand-ing of general protective mechanisms conferring enhanceddrought tolerance to crop becomes an important issue(Sreenivasulu et al. 2004). AM fungi improve drought toler-ance of cereal crops (Ruiz-Lozano and Azcón 1995). Elliset al. (1985) have found that AM colonization increases leafarea, biomass and grain yield of wheat under drought condi-tions. Ruiz-Lozano (2003) confirms that AM symbiosis en-hances the activities of antioxidant enzyme in the eliminationof active oxygen species during water stress. It is also reportedthat AM-inoculated maize has higher concentrations of glu-cose, fructose and amino acids than non-inoculated maizeunder water stress (Celebi et al. 2010). Other factors associat-ed with AM symbiosis may enhance the drought-resistance ofcrops, which include changing in leaf elasticity, improvingleaf water and turgor potentials and regulating stomatal tran-spiration (Al-Karaki et al. 2004).
Foxtail millet (Setaria italica L.) is one of ancient domes-ticated crops for staple foods in arid and semi-arid Africa andEurasian continent before the popularity of rice and wheat, it iscultivated widely for its drought tolerance, rapid growth andhigh grain yield (Sreenivasulu et al. 2004). Foxtail millet isone of important crops for hay, pasture, and food in theseregions today, and also serves as an important biofuel crop toassist future BioEnergy studies (Lata et al. 2011). In this study,we investigated the difference of growth and reactive oxygenmetabolism between Glomus intraradices inoculated andnon-inoculated foxtail millet under well-watered andwater-stressed conditions. The objective of this researchwas to evaluate the effects of G. intraradices on thegrowth, yield, ROS and antioxidant enzymes of foxtailmillet under well-watered and water-stressed conditions.
Furthermore, we attempted to confirm if AM symbiontsenhance the drought tolerance of foxtail millet undergreenhouse pot conditions.
Materials and methods
Soil and biological materials
Soil was collected from experimental farm in Henan Univer-sity of Science and Technology, Luoyang, China. The soilproperties were as follows: pH 7.7 (1:5 soil:water ratio),available nitrogen 33.61 mg kg−1, Olsen phosphorus11.34 mg kg−1, available potassium 94.26 mg kg−1, andorganic matter 16.42 g kg−1. Prior to use in experiments, soilwas sieved through a 2 mm sieve, mixed with fine sand andorganic matter (peat) (soil/sand/organic matter, 2:1:1, v/v/v),and autoclaved at 0.11 Mpa and 121 °C for 2 h.
Glomus intraradices BGC AH01 was provided by theInstitute of Plant Nutrition and Resources, Beijing Academyof Agriculture and Forestry Sciences, China. Mycorrhizalinocula consisted of sand, spores (spores density of 416–582per 100 g dry soil), mycorrhizal hyphae, and infected maizeroot fragments (average 78% colonization rate). Each pot wasinoculated with a 30 g inoculum of the mycorrhizal treatment,or a non-mycorrhizal control treatment that consisted of 30 gsterilized inoculum. Mycorrhizal inoculum was placed 5 cmbelow foxtail millet root at the time of transplantation.
Seeds of Setaria italica L. (Yugu-5) were kindly suppliedby Henan Academy of Agricultural Science (Zhengzhou,China), and stored at 4°C prior to use. Seeds of similar sizewere surface-sterilized using 75% alcohol for 10 min beforesowing, rinsed three times with sterile distilled water, imbibedin sterile distilled water for 24 h, and then germinated in sterilevermiculite at 28°C/80% relative humidity with a photoperiodof 14 h and a photosynthetic photon flux density of 500 μmolm−2 s−1. Ten days after sowing, each developed seedling wastransplanted into a plastic container (15 cm in diameter and15 cm in depth) with 1 kg soil mixture.
Growth conditions
Seedlings were grown in a greenhouse under natural lightbetween May and August 2012 at a temperature of 18–30°C, with 60–75% relative humidity. At 92 days aftertransplanting, all plants were allowed to grow in optimalconditions, and the soil moisture in all pots was maintainedat a field capacity of 75% (−0.12 MPa of soil water potential).A 31 day water-stressed phase followed the growth phase inAugust 2012. In this phase, half of the pots were maintained ina well-watered (WW) condition at 70% field capacity (−0.11MPa), whereas the other half were subjected to water-stressed(WS) conditions at 30% field capacity (−0.68 MPa) for one
Ann Microbiol

month. Prior to stress initiation, irrigation of the WS seedlingswas suspended until the soil moisture reached 30% fieldcapacity. Aluminum foil was used to cover the soil surfaceand the bottom of pots in order to limit water loss. Water losswas measured gravimetrically by weighing the pots and lostwater was replaced with fresh distilled water each day at 18:00in order to keep 70 and 30 % field capacity, respectively.
Root colonization and plant growth analysis
The AM colonization rate was determined by collecting rootsand washing them gently with distilled water. Root sectionsmeasuring 1 cm in length were cleared for 15 min in 10%KOH at 90 °C, bleached in alkaline hydrogen peroxide for 20min, acidified in 1% HCl, and stained with 0.05% (w/v)trypan blue in lactophenol. according to Phillips and Hayman(1970). The colonization rate was determined by the gridlineintersect method (Giovannetti and Mosse 1980).
Physiological response parameters of foxtail millet seed-lings were measured at the end of the water-stressed phase. Allseedlings were harvested and the growth substrate thoroughlyremoved from soil. At the end of growth period, seedlingswere collected and the plant height, root length (the distancebetween the collar and the most distant root apex), collardiameter, panicle height, single panicle weight and 1,000grain weight were separately recorded. Panicle weight andgrain weight was determined by oven drying at 70 °C for 72 h.
Analysis of malondialdehyde and ROS
Fresh leaves or roots (200 mg) were homogenized in 2 mL10 mM sodium phosphate buffer (pH 7.4) and centrifuged at4,000 g for 10 min at 4 °C. The concentration ofmalondialdehyde (MDA) was determined by 2-thiobarbituricacid (TBA) reaction as described by Sudhakar et al. (2001).The production rate of H2O2 and O2
·- were performed asdescribed by Wang and Luo (1990).
Determination of antioxidant enzymes
To analyze antioxidant enzyme activities, 0.2 g fresh leaves orroots were collected randomly from each pot, and then frozenin liquid nitrogen immediately. For extraction, the frozenleaves or roots were homogenized in 5mL 0.1 M cold phos-phate buffer (pH 7.8) containing 0.1 mM EDTA, 1 mM 1,4-dithiothreitol and 2% (w/v) polyvinylpyrrolidone (PVP). Af-ter centrifugation at 12,000 g for 10 min, the supernatant wasused for assays of antioxidant enzyme activity, and all steps ofthe extraction procedure were carried out at 4 °C.
SOD activity was measured according to the method ofGiannopolitis and Ries (1977). One unit of SOD was definedas the amount of enzyme that inhibited the 50% photoreduc-tion of nitro blue tetrazolium (NBT) measured at 560 nm.
CATactivity was measured spectrophotometrically by follow-ing degradation of H2O2 at 240 nm according to the method ofAebi (1984). One unit of CAT activity was expressed as μmolH2O2 oxidized mg−1 protein min−1.
POD activity was determined using the guaiacol oxidationmethod (Chance and Maehly 1955), and quantified spectro-photometrically as the amount of formed tetraguaiacol perminute per gram of fresh plant tissue. Protein was determinedfollowing the method of Bradford (1976) using bovine serumalbumin (BSA, Sigma, St. Louis, MO) as standard.
GR activity was determined by following the decrease inabsorbance at 340 nm due to the glutathione dependence ofNADPH for 10 min. The reaction mixture contained 1 mMEDTA, 0.2 mM NADPH and 0.5 mM GSSG, 100 mM phos-phate buffer (pH 7.8), and 0.2 mL enzyme extract (Chen andWang 2002).
Experimental design and statistical analysis
The experiment was arranged in a randomized block designwith two factors: (1) mycorrhizal treatments, i.e., Glomusintraradices (Schenck& Smith) and a non-mycorrhizal control;(2) soil water conditions, i.e., WWandWS conditions. The datawere subjected to two-factor ANOVA analysis with mycorrhi-zal colonization and soil water as treatments. Each of the fourtreatments had five replicates (pots) making a total of 20 pots(one seedling per pot); five blocks were applied in this study.
All experimental data also had five replications per treat-ment, and means ± SD were calculated. The least significantdifferences were evaluated using Tukey’s multiple range testat 5%. All statistical analyses were performed using SPSSsoftware (version 13.0 for Windows, SPSS, Chicago, IL).
Results
AM colonization
AMcolonizationwas easily found in the roots ofG. intraradices-inoculated foxtail millet, and all non-inoculated seedlings hadnot been infected by AM fungi. Water stress significantly de-creased AM colonization in the roots. AM colonization rate ofG. intraradices-inoculated foxtail millet was 70.8 % under WWconditions and 60.2% under WS conditions respectively(Table 1).
Growth parameters and yield
Water stress depressed plant height, collar diameter, panicleheight, single panicle weight and 1,000 grain weight in foxtailmillet (Table 1). G. intraradices-inoculated foxtail millet hadhigher plant height, collar diameter, panicle height, singlepanicle weight and 1,000 grain weight than non-inoculated
Ann Microbiol
seedlings under both WW and WS conditions (Table 1).G. intraradices-inoculated foxtail millet also had a higher rootlength compared with non-inoculated seedlings under WSconditions, but had no significant difference under WW con-ditions. The enhancement of panicle and grain weight inG. intraradices-inoculated foxtail millet was higher underWS conditions, compared with WW conditions (Table 1).
ROS response
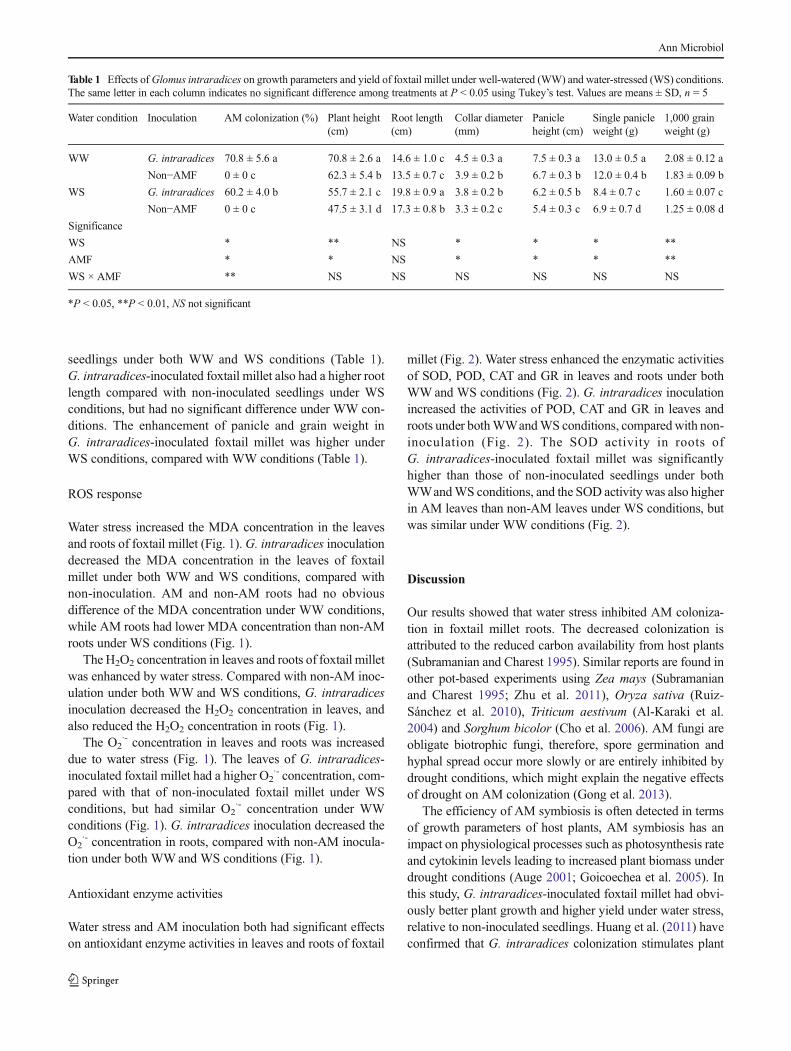
Water stress increased the MDA concentration in the leavesand roots of foxtail millet (Fig. 1).G. intraradices inoculationdecreased the MDA concentration in the leaves of foxtailmillet under both WW and WS conditions, compared withnon-inoculation. AM and non-AM roots had no obviousdifference of the MDA concentration under WW conditions,while AM roots had lower MDA concentration than non-AMroots under WS conditions (Fig. 1).
The H2O2 concentration in leaves and roots of foxtail milletwas enhanced by water stress. Compared with non-AM inoc-ulation under both WW and WS conditions, G. intraradicesinoculation decreased the H2O2 concentration in leaves, andalso reduced the H2O2 concentration in roots (Fig. 1).
The O2·- concentration in leaves and roots was increased
due to water stress (Fig. 1). The leaves of G. intraradices-inoculated foxtail millet had a higher O2
·- concentration, com-pared with that of non-inoculated foxtail millet under WSconditions, but had similar O2
·- concentration under WWconditions (Fig. 1). G. intraradices inoculation decreased theO2
·- concentration in roots, compared with non-AM inocula-tion under both WWand WS conditions (Fig. 1).
Antioxidant enzyme activities
Water stress and AM inoculation both had significant effectson antioxidant enzyme activities in leaves and roots of foxtail
millet (Fig. 2). Water stress enhanced the enzymatic activitiesof SOD, POD, CAT and GR in leaves and roots under bothWW and WS conditions (Fig. 2). G. intraradices inoculationincreased the activities of POD, CAT and GR in leaves androots under bothWWandWS conditions, comparedwith non-inoculation (Fig. 2). The SOD activity in roots ofG. intraradices-inoculated foxtail millet was significantlyhigher than those of non-inoculated seedlings under bothWWandWS conditions, and the SOD activity was also higherin AM leaves than non-AM leaves under WS conditions, butwas similar under WW conditions (Fig. 2).
Discussion
Our results showed that water stress inhibited AM coloniza-tion in foxtail millet roots. The decreased colonization isattributed to the reduced carbon availability from host plants(Subramanian and Charest 1995). Similar reports are found inother pot-based experiments using Zea mays (Subramanianand Charest 1995; Zhu et al. 2011), Oryza sativa (Ruiz-Sánchez et al. 2010), Triticum aestivum (Al-Karaki et al.2004) and Sorghum bicolor (Cho et al. 2006). AM fungi areobligate biotrophic fungi, therefore, spore germination andhyphal spread occur more slowly or are entirely inhibited bydrought conditions, which might explain the negative effectsof drought on AM colonization (Gong et al. 2013).
The efficiency of AM symbiosis is often detected in termsof growth parameters of host plants, AM symbiosis has animpact on physiological processes such as photosynthesis rateand cytokinin levels leading to increased plant biomass underdrought conditions (Auge 2001; Goicoechea et al. 2005). Inthis study, G. intraradices-inoculated foxtail millet had obvi-ously better plant growth and higher yield under water stress,relative to non-inoculated seedlings. Huang et al. (2011) haveconfirmed that G. intraradices colonization stimulates plant
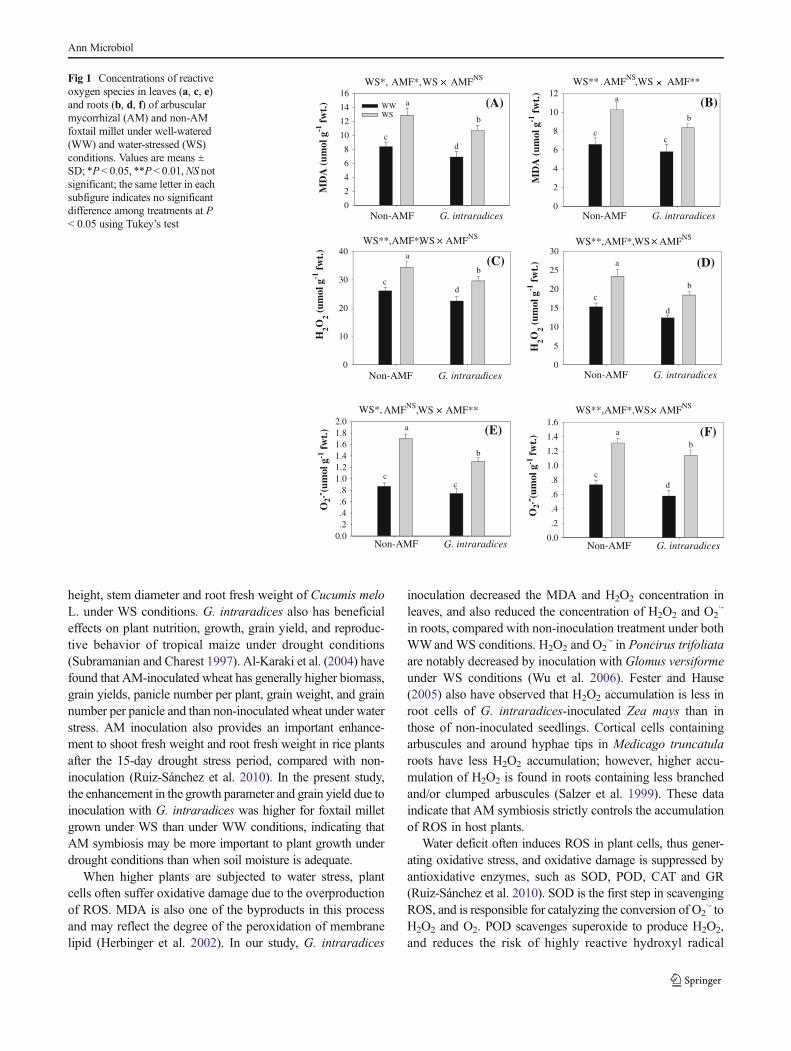
Table 1 Effects ofGlomus intraradices on growth parameters and yield of foxtail millet under well-watered (WW) and water-stressed (WS) conditions.The same letter in each column indicates no significant difference among treatments at P < 0.05 using Tukey’s test. Values are means ± SD, n = 5
Water condition Inoculation AM colonization (%) Plant height(cm)
Root length(cm)
Collar diameter(mm)
Panicleheight (cm)
Single panicleweight (g)
1,000 grainweight (g)
WW G. intraradices 70.8 ± 5.6 a 70.8 ± 2.6 a 14.6 ± 1.0 c 4.5 ± 0.3 a 7.5 ± 0.3 a 13.0 ± 0.5 a 2.08 ± 0.12 a
Non−AMF 0 ± 0 c 62.3 ± 5.4 b 13.5 ± 0.7 c 3.9 ± 0.2 b 6.7 ± 0.3 b 12.0 ± 0.4 b 1.83 ± 0.09 b
WS G. intraradices 60.2 ± 4.0 b 55.7 ± 2.1 c 19.8 ± 0.9 a 3.8 ± 0.2 b 6.2 ± 0.5 b 8.4 ± 0.7 c 1.60 ± 0.07 c
Non−AMF 0 ± 0 c 47.5 ± 3.1 d 17.3 ± 0.8 b 3.3 ± 0.2 c 5.4 ± 0.3 c 6.9 ± 0.7 d 1.25 ± 0.08 d
Significance
WS * ** NS * * * **
AMF * * NS * * * **
WS × AMF ** NS NS NS NS NS NS
*P < 0.05, **P < 0.01, NS not significant
Ann Microbiol
height, stem diameter and root fresh weight of Cucumis meloL. under WS conditions. G. intraradices also has beneficialeffects on plant nutrition, growth, grain yield, and reproduc-tive behavior of tropical maize under drought conditions(Subramanian and Charest 1997). Al-Karaki et al. (2004) havefound that AM-inoculated wheat has generally higher biomass,grain yields, panicle number per plant, grain weight, and grainnumber per panicle and than non-inoculated wheat under waterstress. AM inoculation also provides an important enhance-ment to shoot fresh weight and root fresh weight in rice plantsafter the 15-day drought stress period, compared with non-inoculation (Ruiz-Sánchez et al. 2010). In the present study,the enhancement in the growth parameter and grain yield due toinoculation with G. intraradices was higher for foxtail milletgrown under WS than under WW conditions, indicating thatAM symbiosis may be more important to plant growth underdrought conditions than when soil moisture is adequate.
When higher plants are subjected to water stress, plantcells often suffer oxidative damage due to the overproductionof ROS. MDA is also one of the byproducts in this processand may reflect the degree of the peroxidation of membranelipid (Herbinger et al. 2002). In our study, G. intraradices
inoculation decreased the MDA and H2O2 concentration inleaves, and also reduced the concentration of H2O2 and O2
·-
in roots, compared with non-inoculation treatment under bothWWand WS conditions. H2O2 and O2
·- in Poncirus trifoliataare notably decreased by inoculation with Glomus versiformeunder WS conditions (Wu et al. 2006). Fester and Hause(2005) also have observed that H2O2 accumulation is less inroot cells of G. intraradices-inoculated Zea mays than inthose of non-inoculated seedlings. Cortical cells containingarbuscules and around hyphae tips in Medicago truncatularoots have less H2O2 accumulation; however, higher accu-mulation of H2O2 is found in roots containing less branchedand/or clumped arbuscules (Salzer et al. 1999). These dataindicate that AM symbiosis strictly controls the accumulationof ROS in host plants.
Water deficit often induces ROS in plant cells, thus gener-ating oxidative stress, and oxidative damage is suppressed byantioxidative enzymes, such as SOD, POD, CAT and GR(Ruiz-Sánchez et al. 2010). SOD is the first step in scavengingROS, and is responsible for catalyzing the conversion of O2
·- toH2O2 and O2. POD scavenges superoxide to produce H2O2,and reduces the risk of highly reactive hydroxyl radical
Non-AMF G. intraradices0
2
4
6
8
10
12
14
16WWWS
a
cd
b
(A)
WS** , AMFNSAMFNS ,WS WS AMF**
Non-AMF G. intraradices
MD
A (
umol
g-1
fw
t.)
MD
A (
umol
g-1
fw
t.)
0
2
4
6
8
10
12(B)a
b
cc
Non-AMF G. intraradices
H2O
2 (um
ol g
-1 f
wt.
)
H2O
2 (um
ol g
-1 f
wt.
)
0
10
20
30
40(C)a
bc
d
WS**, AMF*,
AMF*,
WS AMFNS
Non-AMF G. intraradices0
5
10
15
20
25
30a
bc
d
Non-AMF G. intraradices
O2.-
(um
ol g
-1 f
wt.
)
O2.-
(um
ol g
-1 f
wt.
)0.0.2.4.6.8
1.01.21.41.61.82.0
(D)
(E)
WS*,
WS*,
AMFNS, WS AMF** WS**,AMF*, WS AMFNS
WS**,AMF*, WS AMFNS
a
b
cc
Non-AMF G. intraradices0.0
.2
.4
.6
.8
1.0
1.2
1.4
1.6a
b
cd
(F)
Fig 1 Concentrations of reactiveoxygen species in leaves (a, c, e)and roots (b, d, f) of arbuscularmycorrhizal (AM) and non-AMfoxtail millet under well-watered(WW) and water-stressed (WS)conditions. Values are means ±SD; *P < 0.05, **P < 0.01,NS notsignificant; the same letter in eachsubfigure indicates no significantdifference among treatments at P< 0.05 using Tukey’s test
Ann Microbiol
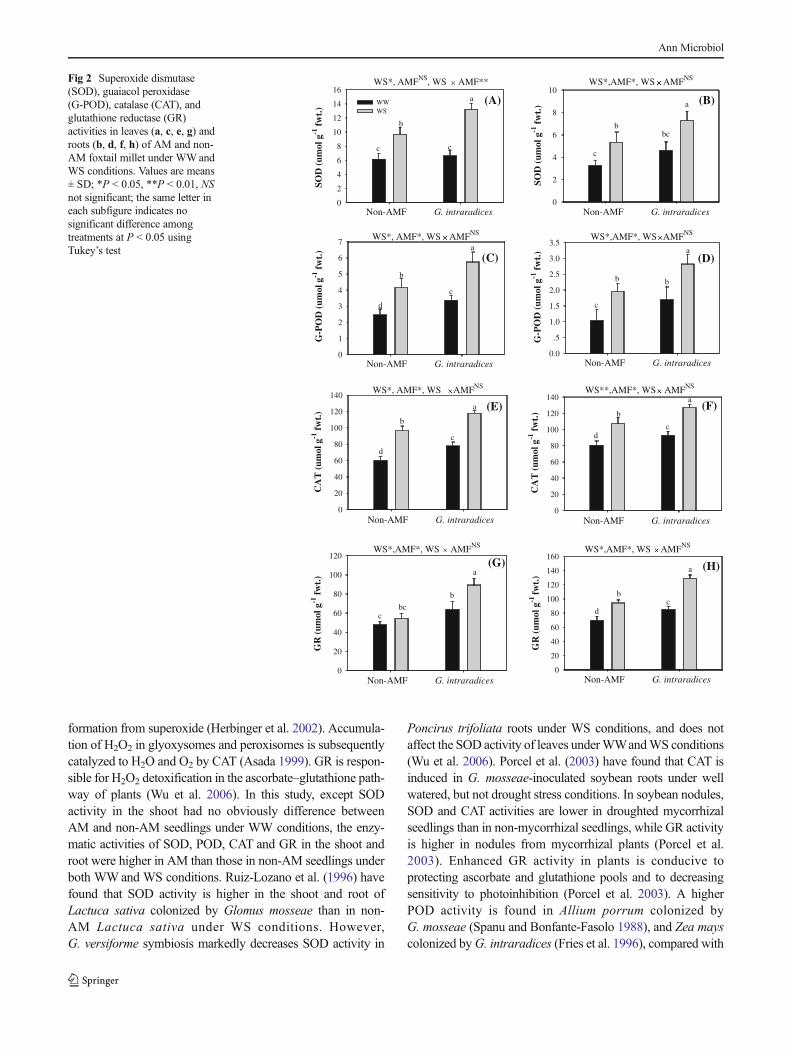
formation from superoxide (Herbinger et al. 2002). Accumula-tion of H2O2 in glyoxysomes and peroxisomes is subsequentlycatalyzed to H2O and O2 by CAT (Asada 1999). GR is respon-sible for H2O2 detoxification in the ascorbate–glutathione path-way of plants (Wu et al. 2006). In this study, except SODactivity in the shoot had no obviously difference betweenAM and non-AM seedlings under WW conditions, the enzy-matic activities of SOD, POD, CAT and GR in the shoot androot were higher in AM than those in non-AM seedlings underboth WWand WS conditions. Ruiz-Lozano et al. (1996) havefound that SOD activity is higher in the shoot and root ofLactuca sativa colonized by Glomus mosseae than in non-AM Lactuca sativa under WS conditions. However,G. versiforme symbiosis markedly decreases SOD activity in
Poncirus trifoliata roots under WS conditions, and does notaffect the SOD activity of leaves underWWandWS conditions(Wu et al. 2006). Porcel et al. (2003) have found that CAT isinduced in G. mosseae-inoculated soybean roots under wellwatered, but not drought stress conditions. In soybean nodules,SOD and CAT activities are lower in droughted mycorrhizalseedlings than in non-mycorrhizal seedlings, while GR activityis higher in nodules from mycorrhizal plants (Porcel et al.2003). Enhanced GR activity in plants is conducive toprotecting ascorbate and glutathione pools and to decreasingsensitivity to photoinhibition (Porcel et al. 2003). A higherPOD activity is found in Allium porrum colonized byG. mosseae (Spanu and Bonfante-Fasolo 1988), and Zea mayscolonized byG. intraradices (Fries et al. 1996), compared with
Non-AMF G. intraradices
SOD
(um
ol g
-1 f
wt.
)
0
2
4
6
8
10
12
14
16
WWWS
(A)
WS*, AMF*, WS AMFNS WS*,AMF*, WS AMFNS
WS*, AMFNS, WS AMF** WS*,AMF*, WS AMFNS
WS*, AMF*, WS AMFNS WS**,AMF*, WS AMFNS
b
c
a
c
Non-AMF G. intraradices
SOD
(um
ol g
-1 f
wt.
)
0
2
4
6
8
10
c
b
a
bc
Non-AMF G. intraradices
G-P
OD
(um
ol g
-1 f
wt.
)
0
1
2
3
4
5
6
7
WS*,AMF*, WS AMFNS WS*,AMF*, WS AMFNS
a
b
c
d
Non-AMF G. intraradices
G-P
OD
(um
ol g
-1 f
wt.
)
0.0
.5
1.0
1.5
2.0
2.5
3.0
3.5
c
b b
a
(B)
(C) (D)
Non-AMF G. intraradices
CA
T (
umol
g-1
fw
t.)
0
20
40
60
80
100
120
140
Non-AMF G. intraradicesC
AT
(um
ol g
-1 f
wt.
)
0
20
40
60
80
100
120
140a
b
c
d
d
b
c
a
Non-AMF G. intraradices
GR
(um
ol g
-1 f
wt.
)
0
20
40
60
80
100
120
Non-AMF G. intraradices
GR
(um
ol g
-1 fw
t.)
0
20
40
60
80
100
120
140
160
d
bc
a
c
b
a
bc
(F)(E)
(G) (H)
Fig 2 Superoxide dismutase(SOD), guaiacol peroxidase(G-POD), catalase (CAT), andglutathione reductase (GR)activities in leaves (a, c, e, g) androots (b, d, f, h) of AM and non-AM foxtail millet under WWandWS conditions. Values are means± SD; *P < 0.05, **P < 0.01, NSnot significant; the same letter ineach subfigure indicates nosignificant difference amongtreatments at P < 0.05 usingTukey’s test
Ann Microbiol

non-mycorrhizal seedlings. Under low P conditions, POD inbean roots colonized by Glomus clarum is induced at an earlyof symbiosis development, but suppressed at a late stage(Lambais et al. 2003). The above findings indicate that antiox-idative enzymes often have differential responses under thesame stress conditions. Determinating total activities in crudehomogenates may not adequately reflect the importance ofcompartment-specific changes (Noctor and Foyer 1998), sinceit does not consider activities at the sites where and when theantioxidative enzymes are synthesized and required (Porcelet al. 2003). However, it is certain that AM symbiosis inducesthe antioxidative enzymes to attenuate cellular photo-oxidativedamage in host plants under drought stress, and that this isperhaps one of the most important mechanisms by which AMsymbiosis increases the drought tolerance of host plants.
This study investigated the responses in terms of growth,grain yield, ROS and antioxidant enzyme activities of foxtailmillet when inoculated with G. intraradices under WW andWS conditions. In conclusion, G. intraradices-inoculated fox-tail millet showed higher drought-tolerance with improvedgrowth and yield, ROS and antioxidant enzyme activities com-pared with non-inoculated foxtail millet. In order to developsustainable agriculture in arid and semi-arid regions of theworld, AM fungal application should be taken into account asa means of alleviating the abiotic stress of agricultural crops.
Acknowledgments This study was supported by the National NaturalScience Foundation of China (Grant Nos. 31200035) and the DoctoralScientific Fund Project of Henan University of Science and Technolo-gy(090017048,09001704). We also thank anonymous reviewers for theirvaluable suggestion to enhance the manuscript.
References
Abei M (1984) Catalase in vitro. Methods Enzymol 105:121–126Al-Karaki G, McMichael B, Zak J (2004) Field response of wheat to
arbuscular mycorrhizal fungi and drought stress. Mycorrhiza 14(4):263–269
Asada K (1999) The water–water cycle in chloroplasts: scavenging ofactive oxygen and dissipation of excess photons. Annu Rev PlantPhysiol Plant Mol Biol 50:601–639
Auge RM (2001) Water relations, drought and vesicular arbuscular my-corrhizal symbiosis. Mycorrhiza 11:3–42
Bradford MM (1976) A rapid and sensitive method for quantitation ofmicrogram quantities of protein utilizing the principle of protein dyebinding. Ann Biochem 72:248–254
Celebi SZ, Demir S, Celebi R, Durak ED, Yilmaz IH (2010) The effect ofArbuscular Mycorrhizal Fungi (AMF) applications on the silagemaize (Zea mays L.) yield in different irrigation regimes. Eur JSoil Biol 4:302–305
Chance B,Maehly AC (1955) Assay of catalase and peroxidase. MethodsEnzymol 2:764–775
Chen JX, Wang XF (2002) Experimental Instruction of Plant Physiology.South China University of Technology Press, Guangzhou
Cho K, Toler H, Lee J, Ownley B, Stutz JC, Moore JL, Augé RM (2006)Mycorrhizal symbiosis and response of sorghum plants to combineddrought and salinity stresses. J Plant Physiol 163:517–528
Ellis JR, Larsen HJ, Boosalis MG (1985) Drought resistance of wheatplants inoculated with vesicular-arbuscular mycorrhizae. Plant Soil86:369–378
Fester T, Hause T (2005) Accumulation of reactive oxygen species inarbuscular mycorrhizal roots. Mycorrhiza 15:373–379
Fries LLM, Pacovsky RS, Safir GR (1996) Expression of isoenzymesaltered by both Glomus intraradices colonization and formononetinapplication in corn (Zea mays L.) roots. Soil Biol Biochem 28:981–988
Giannopolitis CN, Ries SK (1977) Superoxide dismutase. I. Occurrencein higher plants. Plant Physiol 59:309–314
Giovannetti M, Mosse B (1980) An evaluation of techniques formeasuring vesicular–arbuscular infection in roots. NewPhytol 84:489–500
Goicoechea N, Merino S, Sánchez-Díaz M (2005) Arbuscular mycorrhi-zal fungi can contribute to maintain antioxidant and carbon metab-olism in nodules of Anthyllis cytisoides L. subjected to drought. JPlant Physiol 162:27–35
Gong M, Tang M, Chen H, Zhang Q, Feng X (2013) Effects of twoGlomus species on the growth and physiological performance ofSophora davidii seedlings under water stress. New Forest 44:399–408
Herbinger K, Tausz M, Wonisch A, Soja G, Sorger A, Grill D (2002)Complex interactive effects of drought and ozone stress on theantioxidant defence systems of two wheat cultivars. Plant PhysiolBiochem 40:691–696
Huang Z, Zou ZR, He CX, He ZQ, Zhang ZB, Li JM (2011)Physiological and photosynthetic responses of melon (Cucumismelo L.) seedlings to three Glomus species under water deficit.Plant Soil 339:391–399
Kramer PJ, Boyer JS (1995) Water relations of plants and soils.Academic, San Diego
Lambais MR, Ríos-RuizWF, Andrade RM (2003) Antioxidant responsesin bean (Phaseolus vulgaris) roots colonized by arbuscular mycor-rhizal fungi. New Phytol 160:421–428
Lata C, Jha S, Dixit V, Sreenivasulu N, Prasad M (2011) Differentialantioxidative responses to dehydration-induced oxidative stress incore set of foxtail millet cultivars [Setaria italica (L.)]. Protoplasma248:817–828
Noctor G, Foyer CH (1998) Ascorbate and glutathione: keeping activeoxygen under control. Annu Rev Plant Physiol Plant Mol Biol 49:249–279
Omirou M, Ioannis MI, Constantinos E (2013) Mycorrhizal inoculationaffects arbuscular mycorrhizal diversity in watermelon roots, butleads to improved colonization and plant response under water stressonly. Appl Soil Ecol 63:112–119
Phillips JM, Hayman DS (1970) Improved procedures for clearingroots and staining parasitic and vesicular arbuscular mycorrhi-zal fungi for rapid assessment of infection. Trans Br Mycol Soc55:158–161
Porcel R, Barea JM, Ruiz-Lozano JM (2003) Antioxidant activities inmycorrhizal soybean plants under drought stress and their possiblerelationship to the process of nodule senescenc. New Phytol 157:135–143
Ruiz-Lozano JM (2003) Arbuscular mycorrhizal symbiosis and allevia-tion of osmotic stress. New perspectives for molecular studies.Mycorrhiza 13:309–317
Ruiz-Lozano JM, Azcón R (1995) Hyphal contribution to water uptake inmycorrhizal plants as affected by the fungal species andwater status.Physiol Plant 95:472–478
Ruiz-Lozano JM, Azcón R, Palma JM (1996) Superoxide dismutaseactivity in arbuscular mycorrhizal Lactuca sativa plants subjectedto drought stress. New Phytol 134:327–333. doi: 10.1111/j.1469-8137.1996.tb04637.x
Ruiz-Sánchez M, Aroca R, Muñoz Y, Polón R, Ruiz-Lozano JM (2010)The arbuscular mycorrhizal symbiosis enhances the photosynthetic
Ann Microbiol

efficiency and the antioxidative response of rice plants subjected todrought stress. J Plant Physiol 167:862–869
Salzer P, Corbiere H, Boller T (1999) Hydrogen peroxide accumu-lation in Medicago truncatula roots colonized by the arbuscularmycorrhiza-forming fungus Glomus intraradices. Planta 208:319–325
Singh S, Gupta AK, Kaur N (2012) Differential responses of antioxida-tive defence system to long-term field drought in wheat (Triticumaestivum L.) genotypes differing in drought tolerance. J Agron CropSci 198:185–189
Smith SE, Read DJ (1997) Mycorrhizal symbiosis, 2nd edn. Academic,London
Spanu P, Bonfante-Fasolo P (1988) Cell-wall-bound peroxidase activityin roots of mycorrhizal Allium porrum. New Phytol 109:119–124
Sreenivasulu N, Miranda M, Prakash HS, Wobus U, Weschke W(2004) Transcriptome changes in foxtail millet genotypes athigh salinity: Identification and characterization of a PHGPXgene specifically upregulated by NaCl in a salt-tolerant line. JPlant Physiol 161:467–477
Subramanian KS, Charest C (1995) Influence of arbuscular mycorrhizaeon the metabolism of maize under drought stress. Mycorrhiza 5:273–278
Subramanian KS, Charest C (1997) Nutritional, growth, and repro-ductive responses of maize (Zea mays L.) to arbuscular mycor-rhizal inoculation during and after drought stress at tasselling.Mycorrhiza 7:25–32
Sudhakar C, Lakshmi A, Giridarakumar S (2001) Changes in the antiox-idant enzymes efficacy in two high yielding genotypes of mulberry(Morus alba L.) under NaCl salinity. Plant Sci 161:613–619
Wang AG, Luo GH (1990) Quantitative relation between the reaction ofhydroxylamine and superoxide anion radicals in plants. PlantPhysiol Commun 6:55–57
Wu QS, Xia RX, Zou YN (2006) Reactive oxygen metabolism in my-corrhizal and non-mycorrhizal citrus (Poncirus trifoliata) seedlingssubjected to water stress. J Plant Physiol 163:1101–1110
Zhu XC, Song FB, Liu SQ (2011) Arbuscular mycorrhiza impacts ondrought stress of maize plants by lipid peroxidation, proline contentand activity of antioxidant system. J Food Agric Environ 9:583–587
Ann Microbiol