effects of ghrelin on hypothalamic glucose responding neurons in rats
TRANSCRIPT
www.elsevier.com/locate/brainres
Brain Research 1055
Research Report
Effects of ghrelin on hypothalamic glucose responding neurons in rats
Xi Chena, Yin-Lin Gea, Zheng-Yao Jianga,*, Chang-Qin Liua,
Inge Depoortereb, Theo L. Peetersb
aDepartment of Physiology, Qingdao University School of Medicine, Qingdao 266021, P.R. ChinabCentre for Gastroenterological Research, Catholic University of Leuven, Leuven, Belgium
Accepted 30 June 2005
Available online 9 August 2005
Abstract
Ghrelin is an endogenous ligand of the growth hormone secretagogue receptor (GHS-R) with potent stimulatory effects on food intake.
The aim of the present study was to investigate the effects of ghrelin on neuronal activity of hypothalamic glucose responding neurons.
Single unit discharges in the lateral hypothalamic area (LHA), the ventromedial hypothalamic nucleus (VMH), and the parvocellular part of
the paraventricular nucleus(pPVN) were recorded extracellularly by means of four-barrel glass micropipettes in anesthetized rats. The activity
of glucose-sensitive neurons (GSNs) in the LHA, pPVN, and of glucoreceptor neurons (GRNs) in the VMH modulated by administration of
ghrelin was analyzed. In the LHA, the majority of GSNs (17/25) increased in frequency due to ghrelin. Whereas the majority of VMH-GRNs
(27/33) and pPVN-GSNs (9/13) was inhibited. The responses to ghrelin were abolished by pretreatment of [d-Lys-3]-GHRP-6, ghrelin
receptor antagonist. These data indicate that the glucose responding neurons in the LHA, VMH, and pPVN are also involved in the
orexigenic actions of ghrelin in the hypothalamic circuits, although AgRP/NPY neurons in the arcuate nucleus (ARC) are the primary targets
of ghrelin.
D 2005 Elsevier B.V. All rights reserved.
Theme: Neural basis of behavior
Topic: Central control of ingestion
Keywords: Ghrelin; Glucose-sensitive neuron; Glucoreceptor neuron; LHA; VMH; pPVN
1. Introduction
Ghrelin is a recently identified endogenous ligand of the
growth hormone secretagogue receptor (GHS-R) [15]. It
was originally isolated from the stomach [5,6], but has also
shown to be present in the rat hypothalamus [4,18]. Recent
data have led to the recognition that ghrelin plays an
important role in body-weight regulation and energy
0006-8993/$ - see front matter D 2005 Elsevier B.V. All rights reserved.
doi:10.1016/j.brainres.2005.06.080
Abbreviations: AgRP, agouti-related peptide; CART, cocaine- and
amphetamine-regulated transcript; POMC, proopiomelanocortin; GSN,
glucose-sensitive neuron; GRN, glucoreceptor neuron; ARC, arcuate
nucleus; LHA, lateral hypothalamic area; pPVN, parvocellular part of
paraventricular nucleus; VMH, ventromedial hypothalamic nucleus; GHS-
R, growth hormone secretagogue receptor
* Corresponding author. Fax: +86 532 83801449.
E-mail address: [email protected] (Z.-Y. Jiang).
homeostasis because its administration increases food intake
and causes fat and weight gain in rodents [23,31]; the
orexigenic effect of ghrelin seems to be independent of its
GH-releasing activity [14]. It has been found that circulating
levels of ghrelin increase following a 48-h fast, and infusion
of glucose into the stomach decreases plasma ghrelin
concentration [22,31].
Information accumulated over the past decade has
revised our views on the hypothalamic control of appetite.
Hypothalamic areas including the paraventricular nucleus
(PVN), perifornical area (PFA), and the lateral hypothalamic
area (LHA) are richly supplied by axons from the arcuate
nucleus (ARC) NPY/AgRP and POMC/CART neurons
[8,29]. The recent studies have shown that injection of
ghrelin into the cerebrospinal fluid (CSF) induces c-fos
expression in the PVN, dorsomedial (DMH), VMH, and
(2005) 131 – 136

X. Chen et al. / Brain Research 1055 (2005) 131–136132
ARC of the hypothalamus, as well as in the nucleus of the
solitary tract (NTS) and area postrema (AP) of the brain
stem [17]. It has been established that the glucose-sensitive
neurons (GSNs) in the LHA, pPVN, and glucoreceptor
neurons (GRNs) in the VMH are involved in the control of
food intake [25]. Furthermore, it has been shown that the
activity of GSNs in the LHA was suppressed by leptin,
whereas the activity of GRNs in the VMH was facilitated; in
contrast, orexin-A had opposite effects [30]. The present
study was undertaken to examine the effects of ghrelin on
the glucose responding neurons in the LHA, VMH, and
pPVN.
Table 1
Effects of ghrelin on hypothalamic neurons
Decrease Increase No effect
LHA
25 GSNs 3 17 5
47 Non-GSNs 8 2 37
VMH
33 GRNs 27 2 4
48 Non-GRNs 5 19 24
pPVN
13 GSNs 9 1 3
36 Non-GSNs 5 9 22
2. Materials and methods
2.1. Animals
Adult Wistar rats (Qingdao Institute for Drug Control) of
either sex, weighing 220–280 g, were used. They were
housed under conditions of controlled illumination (12:12-h
light/dark cycle, lights on/off: 8:00 a.m./8:00 p.m.), humid-
ity, and temperature (22 T 2 -C) for at least 7 days prior to
the experiments. Standard laboratory chow pellets and tap
water were available ad libitum. All animal experiments
were carried out in accordance with the ethic guidelines of
Qingdao University for animal care.
2.2. Electrophysiological recordings
Rats were anesthetized with urethane (1.0 g/kg, i.p.) and
a maintenance dose of anesthetics was given whenever
necessary. Anesthetized animals were positioned in a
stereotaxic apparatus (Narishige SN-3, Tokyo, Japan) with
the incisor bar 3.3 mm below the center of ear bars, the
dorsal surface of the brain was exposed. Stereotaxic
coordinates were as follows: LHA (1.8–2.3 mm posterior
to the bregma, 1.5–2.5 mm lateral to the sagittal sinus, 7.5–
9.0 mm ventral from the dura); VMH [P: 2.8–3.3 mm,
L(R): 0.2–1.0 mm, H: 9.3–10 mm]; pPVN [P: 1.8–2.3
mm, L(R): 0.1–0.4 mm, H: 7.7–8.4 mm] [26]. Rectal
temperature was maintained at 36–38 -C.Four-barrel glass microelectrode (total tip diameter 3–10
Am, resistance 5–20 MV) was used for electrophysiological
recording and micro-pressure injection. The recording glass
microelectrode was filled with 0.5 M sodium acetate and 2%
Pontamine sky blue. The other three barrels connected with 4-
channel pressure injector (PM2000B,Micro Data Instrument,
Inc., USA) were filled with 2 M solution of glucose (pH 7.4),
15 nM solution of ghrelin, and 28 nM solution of [d-Lys-3]-
GHRP-6 (each was dissolved in 0.9%NaCl) and 0.5MNaCl,
respectively. The barrel filled with 0.5 M NaCl was used to
rule out the osmotic effects and any neurons that responded to
Na+ or Cl� applications were omitted from the results.
Rat ghrelin and the ghrelin receptor antagonist ([d-Lys-
3]-GHRP-6) were generously supplied by Dr. T.L. Peeters
(Gut Hormone Laboratory, Leuven, Belgium). Drugs were
ejected on the surface of firing cells with short pulse gas
pressure (1500 ms, 5.0–15.0 psi) [13]. The intrabarrel drug
concentrations were chosen on the basis of their efficacy to
reliably alter cell firing. Volumes less than 1 nl of ghrelin
were applied to the firing cells during extracellular recording.
The recorded electrical signals were amplified and
displayed on a Memory Oscilloscope (VC-11, Nihon
Kohden), the analog signals were fed into a signal analyzer
and computer which incorporated a signal discriminator to
allow unitary data to be stored on-line.
2.3. Histological verification
To check the position of the recording electrode, at the
end of each experiment a direct current (10 AA, 20 min)
was passed through the electrode to form an iron deposit
of Pontamine sky blue. The rats were perfused trans-
cardially with 0.9% saline, followed by 10% buffered
Formalin solution. The brains were removed, 50-Am frozen
coronal sections were cut through the regions of the
hypothalamus, stained with Neutral red, cleared with
xyline, and coverslipped.
2.4. Data analysis
Data were expressed as means T standard error of the
mean (SEM). Comparisons of agents induced responses
before (pre-) and after (post-) treatment were made by
Student’s t test; the differences of the percentages between
GSNs and non-GSNs responding to ghrelin or [d-Lys-3]-
GHRP-6 or not responding on LHA, VMH, and pPVN
neurons were tested by means of the v2 test. Differences
were considered to be significant at P < 0.05.
3. Results
Results of ghrelin on hypothalamic GSNs and non-GSNs
are summarized in Table 1. 25 (35%) GSNs in 72 LHA
neurons were identified by their suppression in response to
applied glucose. Of 25 LHA-GSNs tested with ghrelin, 17
(68%) GSNs were excited. 33 (40%) GRNs in 81 VMH
neurons were identified by their facilitation in response to
X. Chen et al. / Brain Research 1055 (2005) 131–136 133
applied glucose. Of 33 VMH-GRNs tested with ghrelin, 27
(81.88%) GRNs were inhibited. 13 (26%) pPVN-GSNs
were identified by their suppression in response to glucose.
Of 13 pPVN-GSNs tested with ghrelin, 9 (70%) pPVN-
GSNs were suppressed.
3.1. Effects of ghrelin on LHA GSNs
The effects of pressure-ejected application of glucose and
ghrelin were studied in a total of 72 LHA neurons in 85 rats.
Neurons were said to be glucose-sensitive if the activity of
the neuron is decreased by application of glucose [25].
Glucose inhibited about 35% (25/72) neurons in LHA,
which were identified as GSNs. In LHA, 17 out of 25 (68%)
GSNs and 2 out of 47 (4.3%) non-GSNs showed an
excitation in response to the administration of ghrelin,
whereas 1.2% (3/25) GSNs and 17% (8/47) non-GRNs were
inhibited. The changes in the firing rate in response to
micro-pressure injection of glucose and ghrelin are illus-
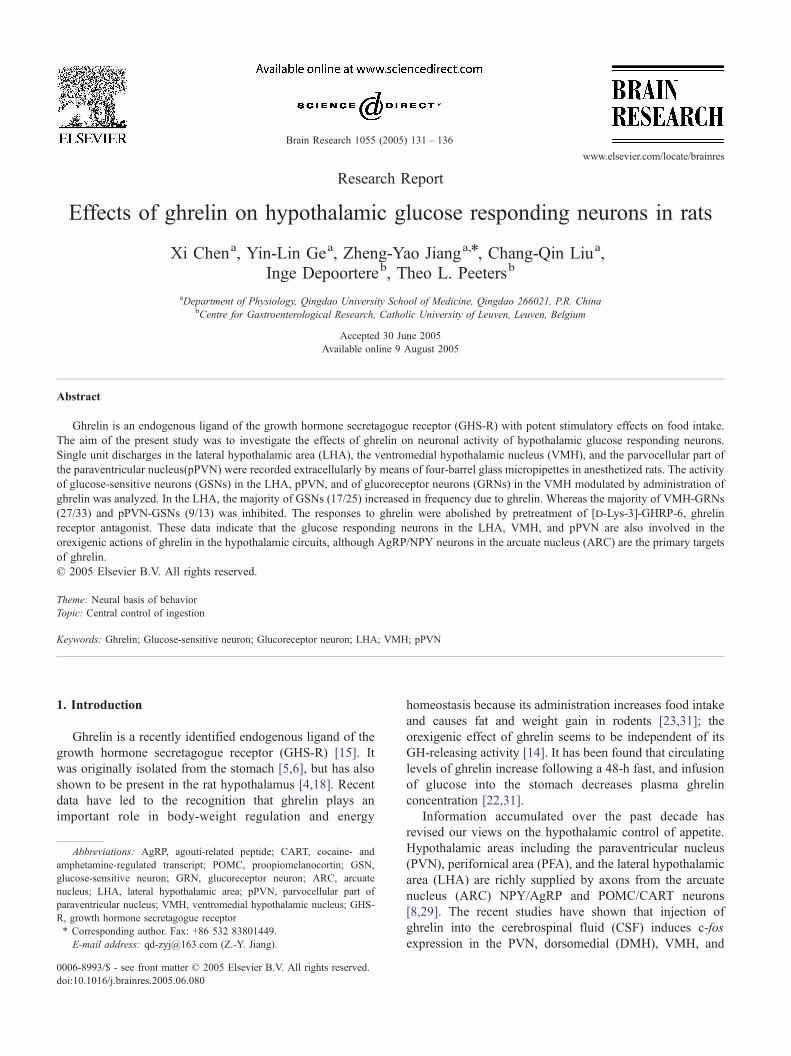
trated in Fig. 1. Administration of ghrelin increased the
firing rate of LHA-GSNs by 62.0 T 12.9%. This increase in
neuronal activity was statistically significant compared with
control level (P < 0.05). The ghrelin-induced response
lasted for 148.2 T 29.3 s. In contrast, ghrelin had no effect
on 20% (5/25) GSNs and 78.7% (37/47) non-GSNs. These
data show that ghrelin had an excitatory effect on a large
proportion of GSNs in the LHA. In addition, the effective-
ness of ghrelin receptor antagonist was tested by the
administration of [d-Lys-3]-GHRP-6 on the 10 LHA-GSNs.
After treatment with [d-Lys-3]-GHRP-6, the ghrelin-
induced response was abolished.
Fig. 1. Effects of ghrelin, [d-Lys-3]-GHRP-6 on firing rate of GSNs in LHA. (A)
NaCl, and ghrelin. Application of 2 M glucose and ghrelin caused a significant dec
0.5 M NaCl had no effect. (B) Similar responses were observed after treatment w
induced excitatory response was abolished.
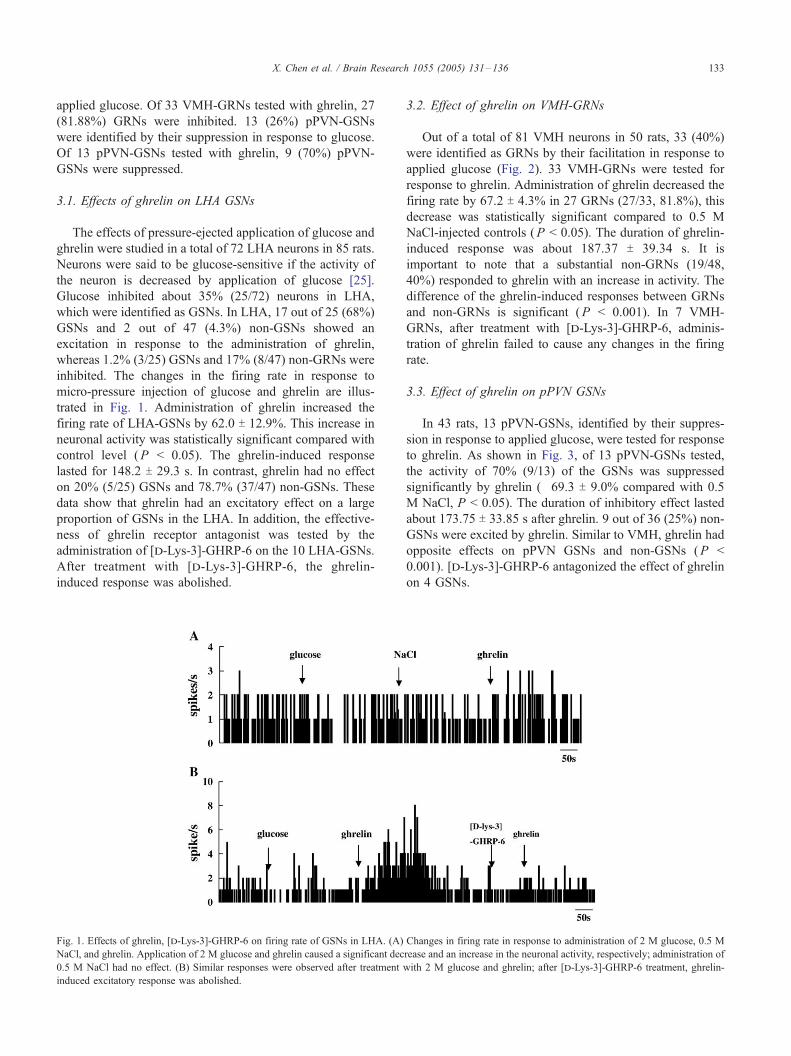
3.2. Effect of ghrelin on VMH-GRNs
Out of a total of 81 VMH neurons in 50 rats, 33 (40%)
were identified as GRNs by their facilitation in response to
applied glucose (Fig. 2). 33 VMH-GRNs were tested for
response to ghrelin. Administration of ghrelin decreased the
firing rate by 67.2 T 4.3% in 27 GRNs (27/33, 81.8%), this
decrease was statistically significant compared to 0.5 M
NaCl-injected controls (P < 0.05). The duration of ghrelin-
induced response was about 187.37 T 39.34 s. It is
important to note that a substantial non-GRNs (19/48,
40%) responded to ghrelin with an increase in activity. The
difference of the ghrelin-induced responses between GRNs
and non-GRNs is significant (P < 0.001). In 7 VMH-
GRNs, after treatment with [d-Lys-3]-GHRP-6, adminis-
tration of ghrelin failed to cause any changes in the firing
rate.
3.3. Effect of ghrelin on pPVN GSNs
In 43 rats, 13 pPVN-GSNs, identified by their suppres-
sion in response to applied glucose, were tested for response
to ghrelin. As shown in Fig. 3, of 13 pPVN-GSNs tested,
the activity of 70% (9/13) of the GSNs was suppressed
significantly by ghrelin (�69.3 T 9.0% compared with 0.5
M NaCl, P < 0.05). The duration of inhibitory effect lasted
about 173.75 T 33.85 s after ghrelin. 9 out of 36 (25%) non-
GSNs were excited by ghrelin. Similar to VMH, ghrelin had
opposite effects on pPVN GSNs and non-GSNs (P <
0.001). [d-Lys-3]-GHRP-6 antagonized the effect of ghrelin
on 4 GSNs.
Changes in firing rate in response to administration of 2 M glucose, 0.5 M
rease and an increase in the neuronal activity, respectively; administration of
ith 2 M glucose and ghrelin; after [d-Lys-3]-GHRP-6 treatment, ghrelin-
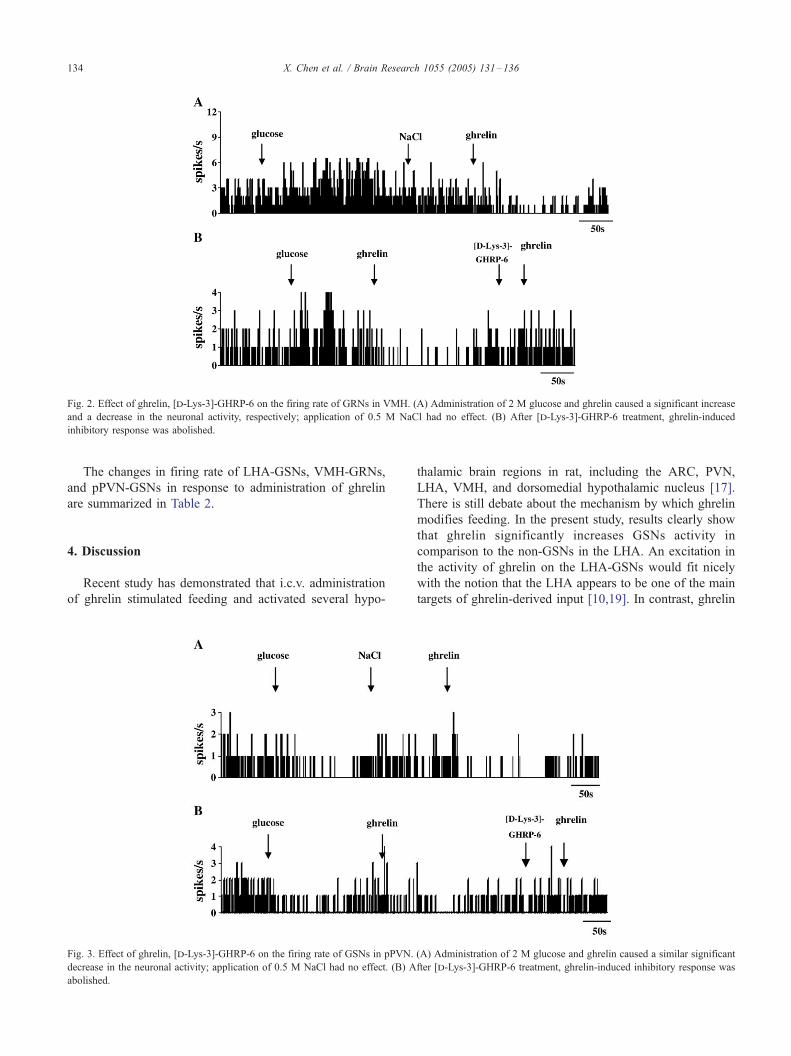
Fig. 2. Effect of ghrelin, [d-Lys-3]-GHRP-6 on the firing rate of GRNs in VMH. (A) Administration of 2 M glucose and ghrelin caused a significant increase
and a decrease in the neuronal activity, respectively; application of 0.5 M NaCl had no effect. (B) After [d-Lys-3]-GHRP-6 treatment, ghrelin-induced
inhibitory response was abolished.
X. Chen et al. / Brain Research 1055 (2005) 131–136134
The changes in firing rate of LHA-GSNs, VMH-GRNs,
and pPVN-GSNs in response to administration of ghrelin
are summarized in Table 2.
4. Discussion
Recent study has demonstrated that i.c.v. administration
of ghrelin stimulated feeding and activated several hypo-
Fig. 3. Effect of ghrelin, [d-Lys-3]-GHRP-6 on the firing rate of GSNs in pPVN.
decrease in the neuronal activity; application of 0.5 M NaCl had no effect. (B) A
abolished.
thalamic brain regions in rat, including the ARC, PVN,
LHA, VMH, and dorsomedial hypothalamic nucleus [17].
There is still debate about the mechanism by which ghrelin
modifies feeding. In the present study, results clearly show
that ghrelin significantly increases GSNs activity in
comparison to the non-GSNs in the LHA. An excitation in
the activity of ghrelin on the LHA-GSNs would fit nicely
with the notion that the LHA appears to be one of the main
targets of ghrelin-derived input [10,19]. In contrast, ghrelin
(A) Administration of 2 M glucose and ghrelin caused a similar significant
fter [d-Lys-3]-GHRP-6 treatment, ghrelin-induced inhibitory response was

Table 2
Changes in firing rate of glucose responding neurons after ghrelin
n Changes in firing rate (%)
LHA-GSNs 17 +62.0 T 12.9
VMH-GRNs 27 �67.2 T 4.3
pPVN-GSNs 9 �69.3 T 9.0
P < 0.05 Compared with 0.5 M NaCl group.
X. Chen et al. / Brain Research 1055 (2005) 131–136 135
inhibits a large proportion of the GRNs in the VMH. This
finding is in agreement with the traditional notion that the
VMH mediates the cessation of eating. It has been
postulated that the PVN plays a pivotal role in the putative
brain network involved in the control of satiety and energy
balance [12]. Information of both anorexigenic systems and
orexigenic peptidergic systems seems to converge in the
PVN [2,11]. We were able to show that about 70% of GSNs
in pPVN were inhibited by administration of ghrelin. A
suppression in the activity of ghrelin on the pPVN GSNs fits
well into the concept that the pPVN GSNs might be the
second-order catabolic effectors located downstream of the
ARC which play a role in the satiety regulation [29]. It is
also interesting to note that leptin has opposite effects. It
inhibits GSNs in the LHA and enhances activity of the
glucose responsive neurons in the VMH and the pPVN [30].
There is convincing evidence that the notion of specific
Fcenter_ of the brain that controls food intake and body
weight has been replaced by the distinct hypothalamic
neuropeptide-containing pathways [29,32]. There is also
new hypothesis that populations of first-order NPY-AgRP
neurons and POMC/CART neurons in the ARC are
regulated by leptin and project to the pPVN, LHA, VMH,
and perifornical area (PFA), which are locations of second-
order hypothalamic neuropeptide neurons involved in the
regulation of food intake and energy homeostasis [29].
Moreover, the LHA GSNs containing orexin project directly
to the glucose-sensitive NPY neurons in the medial ARC
(mARC) where both NPY and AgRP are released [7,27].
Furthermore, it has been shown that leptin suppresses the
activity of the GSNs in the mARC [9,28], whereas it
enhances the activity of GRNs in the lateral ARC (lARC).
The GSNs in mARC contain NPY [21], and the GRNs in
lARC are POMC/CART neurons [16,20].
It has been suggested that ghrelin exerts its effect on
energy balance primarily by binding to the growth hormone
secretagogue(GHS)-receptor located on ARC neurons [1].
In this study, we found that the effects of ghrelin on the
hypothalamic glucose responding neurons in the LHA,
VMH, and pPVN are completely abolished by pretreatment
with [d-Lys-3]-GHRP-6, ghrelin receptor antagonist. It is
well known that ghrelin producing neurons are located
mainly in the ARC, whereas GHS-Rs are distributed in
various regions of the rat brain, including the ARC, VMH,
PVN, LHA, infundibular nucleus, periventricular nucleus,
lateral mammillary nucleus, and the hippocampus [19].
Olzewski et al. [24] recently demonstrated that intra-LH
injection of ghrelin at a dose inducing food intake resulted
in a significant increase of activation of orexin-A-containing
neurons. In a recent paper, Chen et al. [3] demonstrated that
the NPY/AgRP neurons in the ARC were the mediator of
ghrelin-stimulated feeding. With the new evidence provided
by Cowley et al. [4], ghrelin was expressed in a previously
uncharacterized group of neurons in the hypothalamus.
These neurons lay in the space between the lateral, arcuate,
ventromedial, dorsomedial, and paraventricular nuclei, and
they sent projections to several of these nuclei as well as
outside the hypothalamus.
In summary, we conclude that the glucose responding
neurons in the LHA, VMH, and pPVN are also involved in
the orexigenic actions of ghrelin in the hypothalamic
circuits, although the AgRP/NPY neurons in the ARC are
the primary targets of ghrelin-stimulated feeding.
Acknowledgments
This work was supported by the National Natural Science
Foundation of China (No. 30370467) and by the grant from
the Bilateral Scientific and Technological Cooperation
between Flanders and China (Contract BIL01/13).
References
[1] M. Bagnasco, G. Tulipano, M.R. Melis, A. Argiolas, D. Cocchi, E.E.
Muller, Endogenous ghrelin is an orexigenic peptide acting in the arcuate
nucleus in response to fasting, Regul. Pept. 111 (2003) 161–167.
[2] C. Broberger, J. Johansson, M.S. Challing, T. Hokfelt, The neuro-
peptide Y/agouti-related protein(AGRP) brain circuitry in normal,
anorectic, and monosodium glutamate-treated mice, Proc. Natl. Acad.
Sci. U. S. A. 95 (1998) 15043–15048.
[3] H.Y. Chen, M.E. Trumbauer, A.S. Chen, D.T. Weingarth, J.R. Adams,
E.G. Frazier, Z. Shen, D.J. Marsh, S.D. Feighner, X.M. Guan, Z. Ye,
R.P. Nargund, R.G. Smith, L.H. Van der Ploeg, A.D. Howard, D.J.
MacNeil, S. Qian, Orexigenic action of peripheral ghrelin is mediated
by neuropeptide Y and agouti-related protein, Endocrinology 145
(2004) 2607–2612.
[4] M.A. Cowley, R.G. Smith, S. Diano, M. Tschop, N. Pronchuk, K.L.
Grove, C.J. Strasburger, M. Bidlingmaier, M. Esterman, M.L. Heiman,
L.M. Garcia-Segura, E.A. Nillni, P. Mendez, M.J. Low, P. Sotonyi,
J.M. Friedman, H. Liu, S. Pinto, W.F. Colmers, R.D. Cone, T.L.
Horvath, The distribution and mechanism of action of ghrelin in the
CNS demonstrates a novel hypothalamic circuit regulating energy
homeostasis, Neuron 37 (2003) 649–661.
[5] Y. Date, M. Kojima, H. Hosoda, A. Sawaguchi, M.S. Mondal, T.
Suganuma, S. Matsukura, K. Kangawa, M. Nakazato, Ghrelin, a novel
growth hormone-releasing acylated peptide, is synthesized in a distinct
endocrine cell type in the gastrointestinal tracts of rats and humans,
Endocrinology 141 (2000) 4255–4261.
[6] D. Dornonvile, M. Bjorkqvist, A.K. Sandvik, I. Bakke, C.M. Zhao,
D. Chen, R. Hankanson, A-like cells in the rat stomach contain
ghrelin and do not operate under gastrin control, Regul. Pept. 99
(2001) 141–150.
[7] C.F. Elias, C.B. Saper, E. Maratos-Flier, N.A. Tritos, C. Lee, J. Kelly,
M. Yanaigawa, J.K. Elmquist, Chemically defined projections linking
the mediobasal hypothalamus and the lateral hypothalamic area,
J. Comp. Neurol. 402 (1998) 442–459.
[8] J. Elmquist, C. Elias, C. Saper, From lesions to leptin: hypothalamic
control of food intake and body weight, Neuron 22 (1999) 221–232.

X. Chen et al. / Brain Research 1055 (2005) 131–136136
[9] H. Funahashi, T. Yada, S. Muroya, M. Tagigawa, T. Ryushi, S. Horie,
Y. Nakai, S. Shioda, The effect of leptin on feeding-relating neurons in
the rat hypothalamus, Neurosci. Lett. 264 (1999) 117–120.
[10] X.M. Guan, H. Yu, O.C. Palyha, K.K. McKee, S.D. Feighner, D.J.
Sirinathsinghiji, R.G. Smith, L.H. Van der Ploeg, A.D. Howard,
Distribution of mRNA encoding the growth hormone secretagogue
receptor in the brain and peripheral tissue, Brain Res. Mol. Brain Res.
48 (1997) 23–29.
[11] C. Haskell-Luevano, P. Chen, C. Li, K. Chang, M.S. Smith, J.L.
Cameron, R.D. Cone, Characterization of the neuroanatomical
distribution of agouti-related protein immunoreactivity in the rhesus
monkey and the rat, Endocrinology 140 (1999) 1408–1415.
[12] T.L. Horvath, S. Diano, P. Sotonyi, M. Heiman, M. Tschop, Minire-
view: ghrelin and the regulation of energy balance—A hypothalamic
perspective, Endocrinology 142 (2001) 4163–4169.
[13] J.H. Jhamandas, R.W. Lind, L.P. Renau, Angiotensin II may mediate
excitatory neurotransmission from the subfornical organ to the
hypothalamic supraoptic nucleus: an anatomical and electrophysio-
logical study in the rat, Brain Res. 487 (1989) 52–56.
[14] J. Kamegai, H. Tamura, T. Shimizu, S. Ishii, H. Sugihara, I.
Wakabayashi, Central effect of ghrelin, an endogenous growth
hormone secretagogue, on hypothalamic peptide gene expression,
Endocrinology 141 (2000) 4797–4800.
[15] M. Kojima, H. Hosoda, Y. Date, M. Nakazato, H. Matsuo, K.
Kangawa, Ghrelin is a growth-hormone releasing acylated peptide
from stomach, Nature 402 (1999) 656–660.
[16] P. Kristensen, M.E. Judge, L. Thim, U. Rubel, KN. Christjansen, B.S.
Wulff, J.T. Clausen, P.B. Jensen, O.D. Madsen, N. Brang, P.J. Larsen,
S. Hastrup, Hypothalamic CART is a new anorectic peptide regulated
by leptin, Nature 393 (1998) 72–76.
[17] C.B. Lawrence, A.C. Snape, F.M. Baudoin, S.M. Luckman, Acute
central ghrelin and GH secretagogues induce feeding and activate
brain appetite centers, Endocrinology 143 (2002) 155–162.
[18] S. Lu, J.L. Guan, Q.P. Wang, K. Uehara, S. Yamada, N. Goto, Y. Date,
M. Nakazato, M. Kojima, K. Kangawa, S. Shioda, Immunocytochem-
ical observation of ghrein-containing neurons in the rat arcuate
nucleus, Neurosci. Lett. 321 (2002) 157–160.
[19] V. Mitchell, S. Bouret, J.C. Beauvillain, A. Schilling, M. Perent, C.
Kordon, Comparative distribution of mRNA encoding the growth
hormone secretagogue-receptor (GHS-R) in Mirocebus murinus
(primate, lemurian) and rat forebrain and pituitary, J. Comp. Neurol.
429 (2001) 469–489.
[20] T.M. Mizuno, H. Bergen, C. Priest, J. Robersts, Gold-thioglucose
(GTG) reduces hypothalamic proopiomelanocortin (POMC) mRNA
correlated body weight gain and impaired sensitivity, Abstr. - Soc
Neurosci. 24 (1998) 705.
[21] S. Muroya, T. Yada, S. Shioda, M. Tagigawa, K. Goto, Glucose-
sensitive neurons in the rat arcuate nucleus contain neuropeptide Y,
Neurosci. Lett. 264 (1999) 113–116.
[22] E. Nakagawa, N. Nagaya, H. Okumura, et al., Hyperglycaemia
suppresses the secretion of ghrelin, a novel growth-hormone-releasing
peptide: responses to the intravenous and oral administration of
glucose, Clin. Sci. (Lond) 103 (2002) 325–328.
[23] M. Nakazato, N. Murakami, Y. Date, M. Kojima, H. Matsuo, K.
Kangawa, S. Matsukura, A role for ghrelin in the central regulation of
feeding, Nature 409 (2001) 194–198.
[24] P.K. Olzewski, D.H. Li, M.K. Billington, C.M. Kotz, A.S. Levine,
Neural basis of orexicgenic effects of ghrelin acting within lateral
hypothalamus, Peptides 24 (2003) 597–602.
[25] Y. Oomura, Endogenous chemical sense in control of feeding, in: D.
Ottoson, E.R. Perl, R.F. Schmidt, H. Shimazu, W.D. Willis (Eds.),
Progress in Sensory Physiology, Springer-Verlag, Heidelberg, 1989,
pp. 171–191.
[26] G.T. Paxinos, C. Waston, The Rat Brain in Stereotaxic Coordinates,
4th edR, Academic Press, New York, 1998.
[27] C. Peyron, D.K. Tighe, A.N. van den Pol, L. de Lecea, H.C. Heller, J.G.
Sutecliffe, T.S. Kilduff, Neurons containing hypocrtin (orexin) project
to multiple neuronal systems, J. Neurosci. 18 (1998) 9996–10015.
[28] K. Sakai, K. Nagamori, T. Shiraishi, K. Muramoto, Y. Oomura, Effects
of leptin and neuropeptide Y (NPY) on rat arcuate neurons, Abstr. -
Soc. Neurosci. 24 (1998) 449.
[29] M. Schwartz, S. Woods, D.J. Porte, R. Seeley, D. Baskin, Central
nervous system control of food intake, Nature 404 (2000) 661–671.
[30] T. Shiraishi, Y. Oomura, K. Sasaki, M.J. Wayner, Effects of leptin and
orexin-A on food intake and feeding related hypothalamic neurons,
Physiol. Behav. 71 (2000) 251–261.
[31] M. Tschop, D.L. Smiley, M.L. Heiman, Ghrelin induces adiposity in
rodents, Nature 407 (2000) 908–913.
[32] S. Woods, R. Seeley, D.J. Porte, M. Schwartz, Signals that regulate
food intake and energy homeostasis, Science 280 (1998) 1378–1385.