effects of elatobium abietinum on nutrient fluxes in sitka spruce canopies receiving elevated...
TRANSCRIPT

Effects of Elatobium abietinum on nutrient ¯uxes in Sitka sprucecanopies receiving elevated nitrogen and sulphur deposition
Bernhard Stadler*³, Thomas MuÈ ller², Lucy Sheppard³ and Alan Crossley³
*Bayreuth Institute for Terrestrial Ecosystem Research, University of Bayreuth, 95440 Bayreuth, Germany, ²Centre for Agricultural Landscape
and Land Use Research, MuÈncheberg, and Institute of Microbial Ecology and Soil Biology, Gutshof 7, 14641 Paulinenaue, Germany and
³Centre for Ecology and Hydrology, Bush Estate, Penicuik, Midlothian EH26 0QB, UK
Abstract 1 In a ®eld acid mist simulation experiment, Sitka spruce (Picea sitchensis) was
sprayed with different pollutant treatments: N, NH4NO3; S, Na2SO4; NS Acid,
NH4NO3 + H2SO4 and control, no spray. Treatment effects on the abundance of
the green spruce aphid Elatobium abietinum and honeydew production were
assessed. In addition, needles were sampled for phyllosphere micro-organisms. In
a manipulative experiment, shoots were established and maintained as with or
without E. abietinum infestation in order to determine the effects of infestation
on needle loss and throughfall nutrient ¯uxes.
2 Aphid numbers were highest during the end of May and early June, with almost
twice as many needles infested in the NS Acid treatment compared with the
other treatments. Honeydew production was not affected by the treatments.
3 On infested shoots, increasing numbers of yellowing and dead needles were
recorded above the throughfall collectors as the season progressed. The numbers
of dead needles falling into the collectors were signi®cantly higher beneath
infested shoots. There were strong positive correlations between aphid numbers
above the throughfall collectors, the number of yellowing and dead needles on
the shoots and the number of needles in the funnels of the throughfall collectors.
Litter production was more affected by aphid number than by pollutant treat-
ment.
4 Bacteria, yeasts and ®lamentous fungi were more proli®c on infested needles and
treatment effects on colony forming units (CFUs) were most pronounced in the
NS Acid treatment.
5 Fluxes of inorganic nitrogen beneath infested shoots were generally lower than
beneath uninfested shoots. This effect was more pronounced in those treatments
that supplied N i.e. N, NS Acid. The combination of aphid infestation and N-
addition exerted the strongest in¯uence on nutrient ¯uxes. The ¯uxes of potas-
sium and of organic carbon (DOC) were higher beneath infested shoots in all
treatments, through most of the survey period.
Keywords Elatobium abietinum, epiphytic micro-organisms, forested ecosystems,
honeydew, nutrient cycling, pollutants, wet deposition.
Introduction
The response of forested ecosystems to the deposition of
pollutants has been of considerable interest during the last few
decades (Schulze et al., 1989; Field et al., 1992; MuÈller-
Dombois, 1992). Investigations on direct effects of pollutants
on plants have addressed changes in community structure
(Berendse & Elberse, 1990; Tilman, 1990) changes in
physiological processes such as gas exchange (Schulze, 1989;
Curtis & Wang, 1998), growth rates (Bloom et al., 1985;
Sheppard et al., 1999), allocation processes (Wilson, 1988;
Ingestad & AÊ gren, 1991) and effects on nutrient uptake and loss
(Lovett et al., 1985; Sayre & Fahey, 1999). Indirect effects of airCorrespondence: Bernhard Stadler. Tel.: + 49 (0)921 55 5622; fax:
+ 49 (0)921 55 5799; e-mail: [email protected]
ã 2001 Blackwell Science Ltd
Agricultural and Forest Entomology (2001) 3, 253±261Agricultural and Forest Entomology (2001) 3, 253±261

pollutants on plants may originate from increased growth rates of
forest insect populations, which may achieve pest status (Alstad
et al., 1982; Dohmen et al., 1984; Warrington & Whittaker,
1990; Brown et al., 1993; Fleming & Volney, 1995; Docherty
et al., 1997), resulting in detrimental effects on their host plants
(Schowalter et al., 1986; Holopainen et al., 1991). Less clear,
however, are the combined effects of these direct and indirect
stressors on ecosystem response, for example on trophic
interactions and nutrient cycling, resulting from plant±herbivore
interactions under changing environmental conditions.
The green spruce aphid, Elatobium abietinum (Walker), is a
major pest species of Sitka spruce, Picea sitchensis (Bong.)
Carr., and causes considerable economic damage to the forest
industry throughout the spruce growing regions of the UK
(Carter & Nichols, 1988; Straw et al., 1998). Feeding on the
needles results in needle yellowing within a week (Kloft &
Ehrhardt, 1959) and needle fall within 4±6 weeks (Parry, 1974)
eventually leading to a reduction in shoot and tree growth (Straw
et al., 1998). Litterfall studies have demonstrated a reduction in
needle N content on infested spruce (Pedersen & Bille-Hansen,
1999). Studies on trophic interactions induced by phytophagous
insects in the canopies of forest trees indicate a pronounced
effect of the biotic community on nutrient cycling. Herbivores
produce excreta (honeydew, frass) and physically damage the
foliage, improving conditions for microbial growth both in the
canopies and soil, leading to increased carbon availability and
decreased nitrogen ¯uxes (Lovett & Ruesink, 1995; Stadler
et al., 1998; Michalzik & Stadler, 2000; Stadler & MuÈller, 2000).
In this study we report on the effects of aphids on nutrient
¯uxes through the canopy of Sitka spruce under increased N and
S inputs to the forest ecosystem via wet deposition. In particular
we address the questions: (1) does wet deposition affect litter
production through changes in the infestation patterns of aphids;
(2) do pollutants affect trophic interactions between aphids and
epiphytic micro-organisms; (3) does the combination of
pollutant application and aphid infestation differentially affect
nutrient ¯uxes in throughfall of Sitka spruce.
Materials and methods
Experimental design
The experiment covers 1.5 ha, on a very acidic drained basin
peat (< pH 3.0 measured in CaCl2 1 : 5 SoilV : LiquidV),
290 m a.s.l., in the Scottish Borders. The trees, P. sitchensis,
were planted in 1986 at 2 m spacing on ridges of peat removed to
form the drainage ditches. Each plot contained 10 trees in two
lines, and was replicated four times for each treatment. When
treatment commenced in May 1996, the basal branches had
begun to interlock and the 10-year-old trees had a mean height of
3.8 m. The experimental design, site information and meth-
odologies are detailed in Sheppard et al. (1999). Treatments
were made up in rainwater, collected at the site, and applied to
the upper canopy as mist droplets (100±250 mm diameter) from
full cone sprayers (two per tree) supported by a framework of
galvanized steel poles. There were six treatments in the main
experiment, but only four were examined, for the period spring
and early summer 2000. The N treatment provided additional N
at 48 kg/ha/y, the S treatment provided additional S at 50 kg/ha/
y, and the NS Acid treatment at pH 2.5 provided the same
amounts of S and N, but together. The control treatment received
no spray. Treatments were supplied as NH4NO3 (N), Na2SO4
(S), H2SO4 + NH4NO3 (NS Acid). Ions were supplied at a
concentration of 1.6 mol/m3 (NH4+, NO3±, SO4
2±) and 3.2 mol/
m3 (H+, Na+). Background N-deposition at the site is
approximately 8 kg N/ha (wet + dry) and the treatment solution
provides the sprayed trees with an additional 10% of the annual
rainfall. The mean pH of the rainwater is about 4.5 (Sheppard
et al., 1999).
Aphid abundance and honeydew production
The overall abundance of E. abietinum in each treatment plot
was surveyed every 2 weeks from April until July on one-year-
old needles by turning them upside down and counting the
number of infested needles (needles with at least one aphid) per
10 cm of shoot length. These aphids attack one-year-old and
older needles but not growing needles, probably because of a
volatile substance present in the epicuticular wax (Jackson &
Dixon, 1996). Therefore, this method does not give the absolute
number of aphids on a tree during any point in time, especially if
aphids change their feeding places during the growing period or
in response to prevailing weather conditions (Dixon, 1998), but
it provides a relative measurement by which to compare aphid
abundance on trees subjected to different treatments. However,
because these aphids do not cluster in dense colonies, but tend to
spread over a shoot, the number of infested needles should give a
reasonable indication of the overall infestation pattern at the
study site. The survey was restricted to shoots growing at breast
height (c. 1.3 m) and at least 25 shoots were monitored in each
plot on each date. Shoots were chosen at random each time and
no distinction was made between main and side shoots.
The amount of honeydew produced by E. abietinum on trees
subjected to different treatments was measured on 16 June and 4
July. The honeydew was collected by mounting a pre-weighed
aluminium foil (diameter 8 cm) laid in a Petri dish underneath an
aphid colony feeding on one-year-old shoots. For each sampling
date, ®ve replicate foils were set up in each treatment replicate
plot. No effort was made to control the exposure of a branch
where the measurement was made. After 24 h the foils were
removed and the number of aphids above the foils counted. The
honeydew was dried at 30 °C for 2 days and the foil re-weighed
on a Mettler microbalance. The amount of honeydew produced
was calculated as the average of both sampling dates for a single
aphid irrespective of its developmental state. Mean temperatures
on the 2 days were 13.0 °C and 11.3 °C, respectively, with
temperatures recorded every 30 min.
Analysis of micro-organisms in the phyllosphere
On 19 June, current and one-year-old shoots were cut off using
sterile scissors and each shoot was placed immediately into
sterile stomacher polyethylene bags for transport to the
laboratory. Five shoots infested with aphids and ®ve uninfested
shoots were chosen randomly from trees in each treatment (N, S,
NS Acid, Control), but only one shoot per tree was collected. In
addition, the shoots came from trees growing in different
replicate plots. Shoots were sampled mainly from second order
ã 2001 Blackwell Science Ltd, Agricultural and Forest Entomology, 3, 253±261
254 Bernhard Stadler et al.254 Bernhard Stadler et al.
peripheral branches similar to those above the throughfall
collectors. Because of the density of the growing trees no effort
was made to collect shoots facing the same aspect. No rainfall
was recorded for 4 days prior to the sampling of the shoots.
For the microbial analyses, 5 g of needles from one-year-old
shoots were cut off with sterile scissors and blended with
145 mL water for 2 min. These sample solutions were then
logarithmically diluted with quarter-strength Ringer solution
and analysed by spread plating. A Standard II nutrient agar
(Merck, Darmstadt, Germany) medium (pH 7.0) was used to
grow aerobic heterotrophic bacteria, supplemented with 0.4 g/L
cycloheximide (Merck) to inhibit the growth of fungi. The yeasts
and ®lamentous fungi were grown on a Sabouraud-1% dextrose-
1% maltose agar (Merck) (pH 5.5) medium, to which 0.4 g/L
chloramphenicol (Berlin-Chemie, Berlin, Germany) was added
to suppress bacterial growth. All plates were incubated at 25 °C
for 5 days before the number of colony forming units (CFU) was
counted.
Manipulation experiment and throughfall collection
In each treatment (N, S, NS Acid, Control) three polyethylene
collectors (sample volume 5 L) were placed beneath shoots that
were arti®cially infested with 20 adult aphids, and another set of
collectors beneath uninfested shoots. This de®ned a common
infestation pattern above all the throughfall collectors at the start
of the study. Subsequently, aphid numbers above each through-
fall collector were only counted on the needles de®ned by a
vertical projection into the canopy of the collector sampling area
(diameter 20 cm). Again, shoots were turned upside down,
examined for aphids and their numbers on the needles were
counted between throughfall collection dates. On uninfested
control shoots, any aphids that happened to colonize the shoots
above the collectors were manually removed with a pair of ®ne
tweezers. No insecticides were used in case they interfered with
throughfall chemistry. In addition, the numbers of yellowing and
dead needles due to current or earlier aphid infestations on the
shoots of the previous year were counted. This was done by
selecting three or four shoots in the canopy above the collectors
(as previously de®ned) and counting the needles in a particular
category (yellowing, those which showed green and yellow
bands, or dead needles that were completely brown) out of a total
of 100 needles.
Each sampler was located beneath a different tree. The trees
were chosen at random in different replicate chambers and the
collectors were placed towards the periphery of the canopy to
collect throughfall from mainly the one-year-old shoots. No
more than ®ve or six shoots were located above each sampler.
Throughfall samples were collected every two weeks from 3
May to 5 July. Only the last sampling period was longer
(25 days), due to small amounts of rainfall. Previous experi-
ments have shown that the concentrations of organic and mineral
nitrogen are not signi®cantly in¯uenced by microbes washed
into the throughfall collectors, even when much longer sampling
intervals are used (Michalzik et al., 1997).
On each sampling date, the numbers of needles landing in the
funnel of the throughfall collectors were counted to determine
litter fall due to aphid infestation and treatment. Throughfall
collectors were cleaned with deionized water after each
sampling interval and a new coarse ®lter was inserted in the
funnel to prevent debris being washed into the collectors.
After transfer to the laboratory, samples were immediately
vacuum ®ltered (cellulose-acetate membrane, 0.45 mm) and
stored at 4 °C until the next day when the chemical analyses
were done.
Chemical analyses
Dissolved organic carbon (DOC) was measured by infrared
detection of CO2 after persulphate UV digestion using a DOC
analyser (Foss Heraeus, Liqui TOC, Hanau, Germany).
Ammonium-N (NH4-N), nitrate-N (NO3-N), Cl±, Mg2+, K+
and Ca2+ were measured by ion chromatography (Metrohn Ion
Analyser, Metrohn Ltd, U.K). Dissolved organic nitrogen
(DON) was calculated by difference, subtracting the sum of
NH4-N and NO3-N from the total dissolved nitrogen (Ntotal).
Total dissolved nitrogen was measured as NOx after thermo-
oxidation at 700 °C (Abimed: TN-05, Langenfeld, Germany).
Statistical analyses
Differences in the number of CFUs between aphid treatments
were tested with the Mann±Whitney U-test. Because of the high
variability in rainfall amounts during the experiment, through-
fall ¯uxes are presented as differences between infested and
uninfested trees from the different treatments (N, S, NS Acid,
Control). This approach precludes statistical treatment, but is
better suited to highlighting the effect of aphids and microbes on
throughfall chemistry during the course of the season. Fluxes
were expressed as either mg/m2/14 days (or per 25 days),
depending on the sampling frequency as determined by rainfall.
Multivariate analysis of variance (GLM) was used to determine
the effect of treatment and infestation level on ions not included
in the treatment spray. This analysis was restricted to the ®rst
three sampling dates when aphid numbers were increasing.
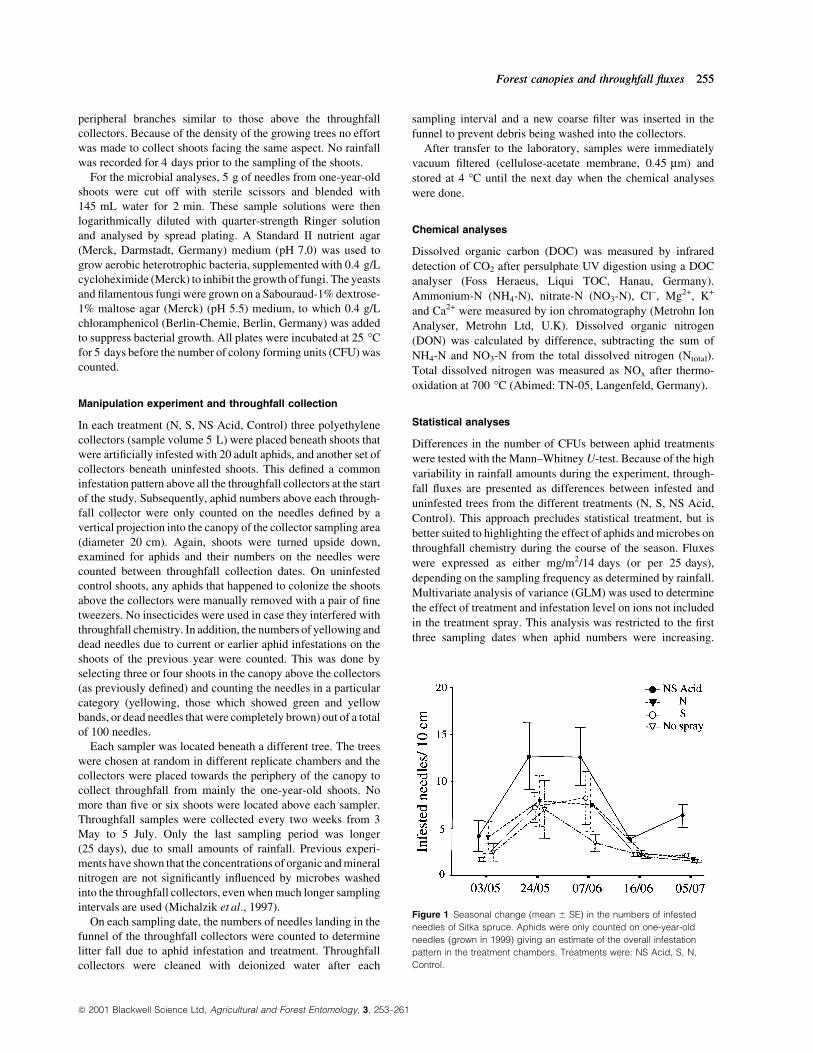
Figure 1 Seasonal change (mean 6 SE) in the numbers of infested
needles of Sitka spruce. Aphids were only counted on one-year-old
needles (grown in 1999) giving an estimate of the overall infestation
pattern in the treatment chambers. Treatments were: NS Acid, S, N,
Control.
ã 2001 Blackwell Science Ltd, Agricultural and Forest Entomology, 3, 253±261
Forest canopies and throughfall ¯uxes 255Forest canopies and throughfall ¯uxes 255
Omitting the period when aphid numbers were declining helps to
exclude possible artefacts associated with delayed effects. For
example, needles turn yellow and die even if the aphids have fed
for only a short period of time (Parry, 1974). Homogeneity of
variances was tested using Levine's test and normality using the
Shapiro±Wilk test. Except for the Cl-¯ux, all other ion ¯uxes
were log- and square-root-transformed to satisfy the assumption
of homogeneity of variances. When treatment effects were not
signi®cant, the data were pooled for correlation analyses with
aphid infestation. All data were analysed with the SPSS
statistical package (SPSS, 1999).
Results
Aphid infestation
On one-year-old shoots, spruce aphid numbers started to
increase in early May and peaked during the end of May/early
June (Fig. 1). Subsequently, the number of infested needles
declined to levels similar to those in early spring. During this
period, trees subjected to the NS Acid treatment had signi®cantly
more needles infested with aphids than those from the other
treatments (repeated measure ANOVA: F4,93 = 18.643,
P < 0.001). On a one-year-old shoot (10 cm in length), an
average of 12.7 needles were infested in the NS Acid treatment
compared to 6.9 in the N, S and control treatments.
Honeydew production was not affected by treatment (ANOVA:
F3,39 = 0.595, P = 0.622). On average a single aphid produced
0.0157 mg honeydew over 24 h.
Phyllosphere micro-organisms
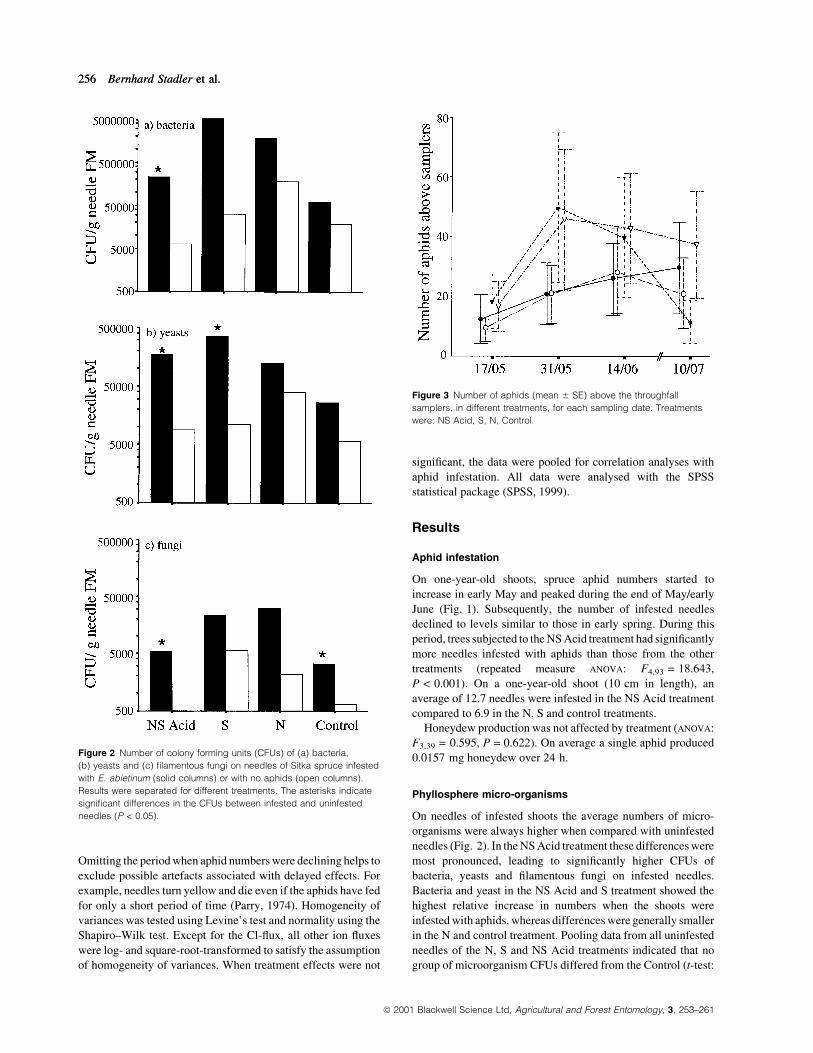
On needles of infested shoots the average numbers of micro-
organisms were always higher when compared with uninfested
needles (Fig. 2). In the NS Acid treatment these differences were
most pronounced, leading to signi®cantly higher CFUs of
bacteria, yeasts and ®lamentous fungi on infested needles.
Bacteria and yeast in the NS Acid and S treatment showed the
highest relative increase in numbers when the shoots were
infested with aphids, whereas differences were generally smaller
in the N and control treatment. Pooling data from all uninfested
needles of the N, S and NS Acid treatments indicated that no
group of microorganism CFUs differed from the Control (t-test:
Figure 2 Number of colony forming units (CFUs) of (a) bacteria,
(b) yeasts and (c) ®lamentous fungi on needles of Sitka spruce infested
with E. abietinum (solid columns) or with no aphids (open columns).
Results were separated for different treatments. The asterisks indicate
signi®cant differences in the CFUs between infested and uninfested
needles (P < 0.05).
Figure 3 Number of aphids (mean 6 SE) above the throughfall
samplers, in different treatments, for each sampling date. Treatments
were: NS Acid, S, N, Control.
ã 2001 Blackwell Science Ltd, Agricultural and Forest Entomology, 3, 253±261
256 Bernhard Stadler et al.256 Bernhard Stadler et al.
bacteria: t = 0.904, d.f. = 18, P(2-tailed) = 0.378; yeasts:
t = 1.564, d.f. = 17, P(2-tailed) = 0.136; fungi: t = 0.800,
d.f. = 18, P(2-tailed) = 0.434). This suggests the 10% increase
in water applied with the treatment solution had no effect on
epiphytic micro-organisms. In total, numbers of CFUs on
infested needles increased 13.9-fold for bacteria, 6.3-fold for
yeasts and 5.9-fold for fungi compared to those on uninfested
needles.
Manipulation experiment
The number of aphids above the throughfall collectors at the ®rst
sampling date after the manipulation was similar in the different
treatments, with an average of 14.2 aphids per shoot (Fig. 3).
Aphid densities increased until the end of May and tended to
decline from mid-June. The variability in aphid number above
the collectors was always rather high, which might re¯ect the
mobility of these aphids. Note that the overall infestation pattern
given in Fig. 1 does not exactly mirror the aphid densities above
the throughfall collectors (Fig. 3). This was because the aphids
kill the needles they feed on, forcing them to move to adjacent
needles/branches. As the throughfall collectors remained in the
same place throughout the experiment, the number of aphids
above a sampler might change quite substantially, which is
indicated by the large standard errors.
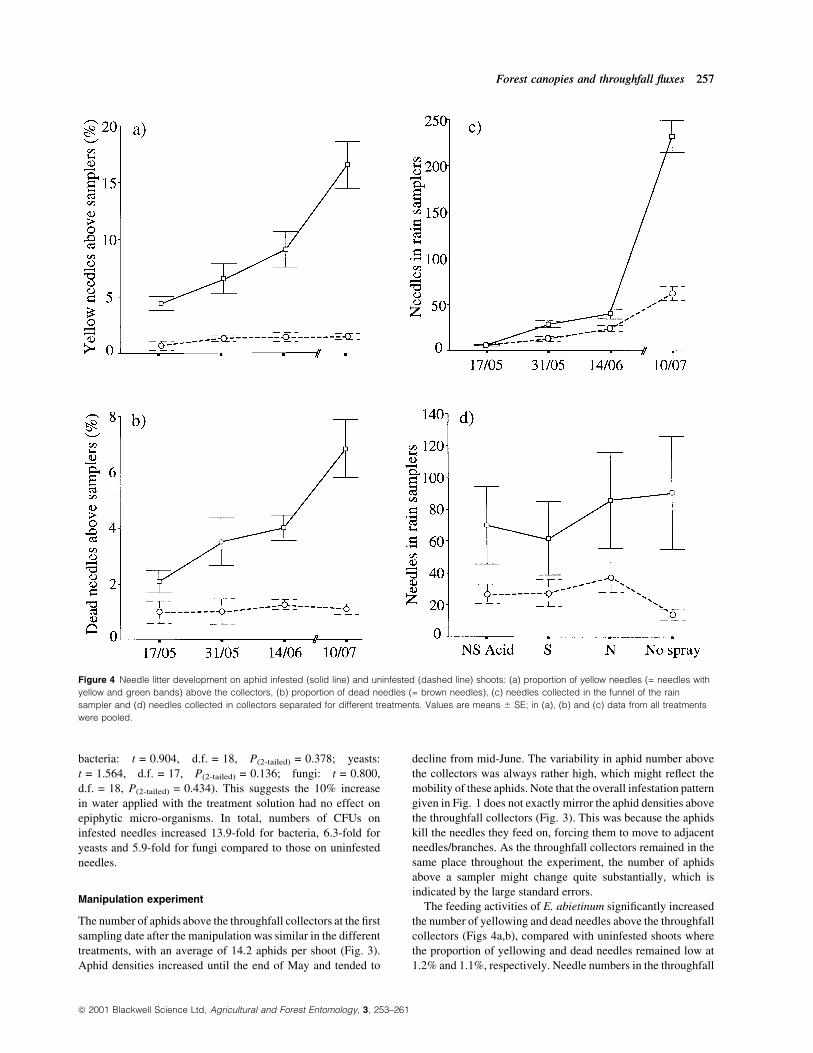
The feeding activities of E. abietinum signi®cantly increased
the number of yellowing and dead needles above the throughfall
collectors (Figs 4a,b), compared with uninfested shoots where
the proportion of yellowing and dead needles remained low at
1.2% and 1.1%, respectively. Needle numbers in the throughfall
Figure 4 Needle litter development on aphid infested (solid line) and uninfested (dashed line) shoots: (a) proportion of yellow needles (= needles with
yellow and green bands) above the collectors, (b) proportion of dead needles (= brown needles), (c) needles collected in the funnel of the rain
sampler and (d) needles collected in collectors separated for different treatments. Values are means 6 SE; in (a), (b) and (c) data from all treatments
were pooled.
ã 2001 Blackwell Science Ltd, Agricultural and Forest Entomology, 3, 253±261
Forest canopies and throughfall ¯uxes 257Forest canopies and throughfall ¯uxes 257
collectors increased exponentially with the duration of the
infestation (Fig. 4c). Signi®cantly more needles were present in
the throughfall collectors beneath infested shoots (t-test:
t = 3.487, d.f. = 94, P < 0.001) leading to a 66% increase in
needle litter (Fig. 4d). Despite the short delay in the develop-
ment of yellowing of needles after the aphids start feeding, there
was a positive correlation between the number of aphids and
yellow needles above the collectors (rp = 0.809, P < 0.0001,
n = 71). A similar correlation was obtained between the number
of aphids and dead needles above the collectors (rp = 0.508, P
< 0.0001, n = 71).
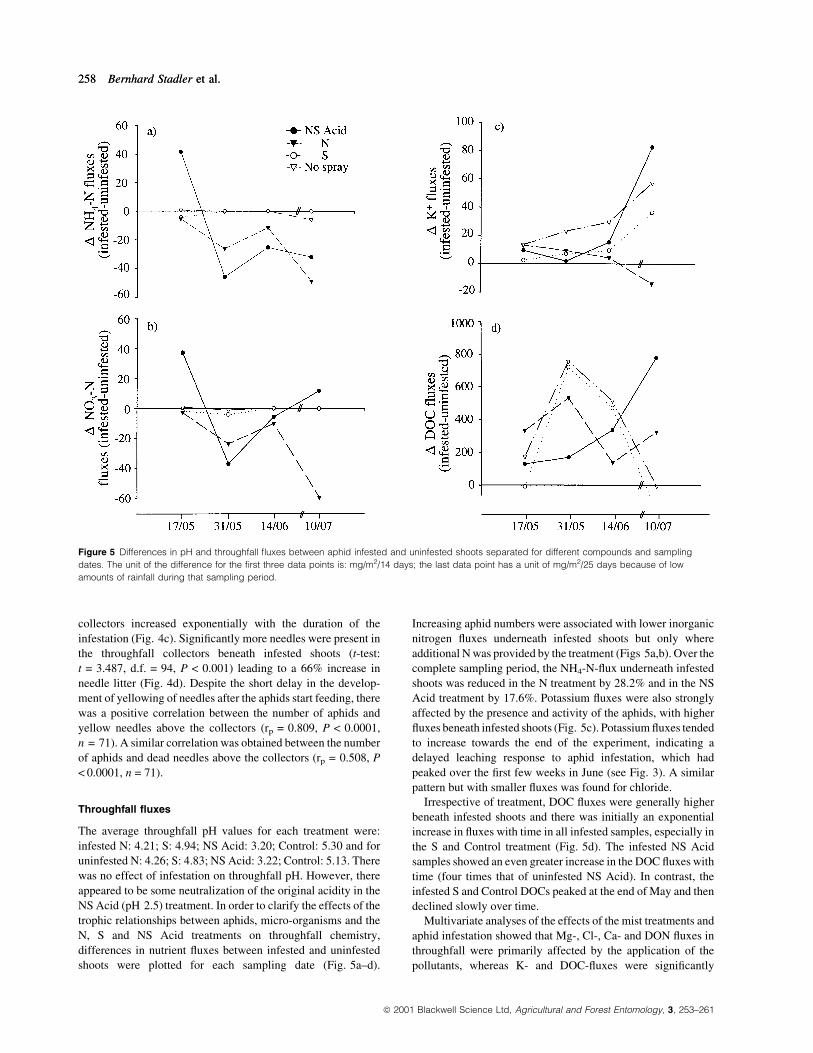
Throughfall ¯uxes
The average throughfall pH values for each treatment were:
infested N: 4.21; S: 4.94; NS Acid: 3.20; Control: 5.30 and for
uninfested N: 4.26; S: 4.83; NS Acid: 3.22; Control: 5.13. There
was no effect of infestation on throughfall pH. However, there
appeared to be some neutralization of the original acidity in the
NS Acid (pH 2.5) treatment. In order to clarify the effects of the
trophic relationships between aphids, micro-organisms and the
N, S and NS Acid treatments on throughfall chemistry,
differences in nutrient ¯uxes between infested and uninfested
shoots were plotted for each sampling date (Fig. 5a±d).
Increasing aphid numbers were associated with lower inorganic
nitrogen ¯uxes underneath infested shoots but only where
additional N was provided by the treatment (Figs 5a,b). Over the
complete sampling period, the NH4-N-¯ux underneath infested
shoots was reduced in the N treatment by 28.2% and in the NS
Acid treatment by 17.6%. Potassium ¯uxes were also strongly
affected by the presence and activity of the aphids, with higher
¯uxes beneath infested shoots (Fig. 5c). Potassium ¯uxes tended
to increase towards the end of the experiment, indicating a
delayed leaching response to aphid infestation, which had
peaked over the ®rst few weeks in June (see Fig. 3). A similar
pattern but with smaller ¯uxes was found for chloride.
Irrespective of treatment, DOC ¯uxes were generally higher
beneath infested shoots and there was initially an exponential
increase in ¯uxes with time in all infested samples, especially in
the S and Control treatment (Fig. 5d). The infested NS Acid
samples showed an even greater increase in the DOC ¯uxes with
time (four times that of uninfested NS Acid). In contrast, the
infested S and Control DOCs peaked at the end of May and then
declined slowly over time.
Multivariate analyses of the effects of the mist treatments and
aphid infestation showed that Mg-, Cl-, Ca- and DON ¯uxes in
throughfall were primarily affected by the application of the
pollutants, whereas K- and DOC-¯uxes were signi®cantly
Figure 5 Differences in pH and throughfall ¯uxes between aphid infested and uninfested shoots separated for different compounds and sampling
dates. The unit of the difference for the ®rst three data points is: mg/m2/14 days; the last data point has a unit of mg/m2/25 days because of low
amounts of rainfall during that sampling period.
ã 2001 Blackwell Science Ltd, Agricultural and Forest Entomology, 3, 253±261
258 Bernhard Stadler et al.258 Bernhard Stadler et al.

affected by the aphids (Table 1). No signi®cant interaction
effects were identi®ed, indicating the speci®city of both effects.
Discussion
Understanding to what extent insects contribute to the
disturbance and destabilization of forest ecosystems under
changing environmental conditions is dependent on our ability to
link life-history attributes, population and community dynamics
and their regulating mechanisms with biochemical ¯uxes. Forest
canopies provide a useful subsystem in which to study the
integration of some of these factors (Schowalter, 2000).
Early indicators of damage to Sitka spruce needles by the
green spruce aphid, E. abietinum, are yellow chlorotic bands that
develop at the sites of stylet insertion that can lead to needles
dying and turning brown. Infestations rarely kill the tree but can
result in reduced growth (Carter, 1977; Seaby & Mowat, 1993;
Straw et al., 1998). Aphids grown on trees exposed to a variety of
pollutants, including SO2 have been shown to exhibit enhanced
growth rates (Holopainen et al., 1991; Brown et al., 1993;
Docherty et al., 1997; Watt et al., 1998). For example, in an SO2
fumigation experiment aphid numbers on Sitka spruce increased
threefold but only on well watered trees (Warrington &
Whittaker, 1990). The overall aphid population densities in
this study were comparable with those recorded in other ®eld
studies with moderate to severe degrees of infestations (Day,
1984), with the highest abundance being observed in the NS
Acid. However, growth analyses showed no reduction in NS
Acid tree growth compared to the other treatments. On the
contrary, stem growth was enhanced by 15±20% (Sheppard
et al., 1999). The absence of enhanced aphid numbers when N
and S were supplied separately suggests that the aphids were
responding to the combined input of N and S and possibly
acidity.
There were positive correlations between aphid numbers
above the throughfall collectors and the number of yellowing or
dead needles. In addition, the numbers of needles that fell into
the collectors were signi®cantly higher beneath infested shoots,
as previously observed by Parry (1974), Day & McClean (1991)
and Watt et al. (1998). About 66% of the needles shed during the
experimental period could be attributed to aphids, rather than to
direct treatment effects or to the growth conditions. This
suggests that the impacts of aphids may mask pollutant effects
on litterfall. Interestingly, the highest litterfall has consistently,
for over 2 years, been recorded in the NS Acid treatments
(Sheppard & Crossley, 2000). There were also considerable
differences in precipitation and periods with strong winds during
the study period, which could affect the number of dead needles
dropping from the shoots and thus affect the strength of the
correlation with aphid abundance. Similarly, all biotic processes
in the canopy are likely to be dominated by the prevailing
weather conditions. Despite these confounding factors, trophic
links between aphids and micro-organisms were detectable in all
treatment chambers. For example, nitrogen ¯uxes declined more
underneath infested trees, whereas DOC-¯uxes increased
underneath infested trees. The decline of inorganic nitrogen
¯uxes underneath infested trees was more pronounced in the N
addition chambers (N, NS Acid). Therefore, both compounds
(honeydew, N) need to be available to signi®cantly affect
nutrient cycling in the canopy. The ¯ux of potassium in
throughfall is a good indicator of the feeding pressure, showing
a strong relationship with aphid abundance and thus, yellowing
and dead needles. Therefore, the results for E. abietinum were
similar to those for folivores in deciduous forests (Seastedt et al.,
1983) where the rate of cycling of K is enhanced in a similar way.
The leaching of potassium was not affected by the pollutants per
se, showing its sensitivity to herbivore stressors.
Under ®eld conditions, no differences in the amount of
honeydew produced by E. abietinum on trees subjected to
different treatments could be found. Although these aphids
produced only small quantities of honeydew, the mechanism
determining how the droplets are ¯icked off the anus are more
important than the absolute amount of honeydew produced.
Stadler & MuÈller (2000) showed that epiphytic micro-organisms
grew better when honeydew was scattered in tiny droplets across
leaves than when concentrated in a few spots. As a consequence,
the overall distribution of aphids within a tree might play an
important role in trophic cascades and the variability in
throughfall chemistry. Elatobium abietinum had strong positive
effects on the growth of epiphytic micro-organisms and these
were especially pronounced in the NS Acid treatment (Fig. 2),
which might explain the strong initial decline in NH4-N and
NO3-N-¯uxes underneath infested spruce (Figs 5a.b). This was a
surprising response, because at pH 2.5 we expected less
microbial activity on the needle surface. It is conceivable that
an acid-tolerant micro¯ora, which also uses honeydew as energy
source developed on the needle surface in the NS Acid treatment.
Currently, our data only re¯ect the situation in the peripheral
part of the canopy and it remains to be shown how biotic
interactions and throughfall dynamics are linked and vary
Table 1 Multivariate (Wilks' lambda) and univariate F-statistics from GLM-MANOVA of the effects of spraying treatment and the presence of aphids on
throughfall ¯uxes (mg/m2/14 days) during the ®rst 6 weeks of the experiment. Signi®cant effects are in italics
Wilks' lambda K-¯ux Mg-¯ux Cl-¯ux Ca-¯ux DOC-¯ux DON-¯ux
dfa F P dfb F P F P F P F P F P F P
Spraying treatment 3, 71 9.34 < 0.001 3, 63 0.90 0.444 44.55 < 0.001 7.14 < 0.001 37.55 < 0.001 0.70 0.556 10.24 < 0.001
Aphid Infestation 1, 71 14.28 < 0.001 1, 63 17.54 < 0.001 0.81 0.373 0.27 < 0.603 0.34 0.563 49.64 < 0.001 2.11 0.151
Spraying ¢ Infestation 3, 71 1.530 0.085 3, 63 0.81 0.490 0.62 0.602 0.48 0.701 0.92 0.439 1.58 0.203 1.47 0.231
aDegrees of freedom (treatment, error) for multivariate analysis.bDegrees of freedom (treatment, error) for univariate analysis.
ã 2001 Blackwell Science Ltd, Agricultural and Forest Entomology, 3, 253±261
Forest canopies and throughfall ¯uxes 259Forest canopies and throughfall ¯uxes 259

horizontally and vertically within the canopy. Nevertheless,
even in the periphery of the Sitka spruce canopy E. abietinum
exerted a strong effect on litterfall, microbial communities and
throughfall chemistry. These effects were quite robust and in
terms of throughfall chemistry, were most pronounced in the
treatments that contained nitrogen (N, NS Acid). Therefore, if
aphid numbers are positively affected via pollutants (directly or
indirectly), signi®cantly higher rates of nutrient cycling might be
expected in the canopies of forest trees receiving elevated N
deposition.
Acknowledgements
We would like to thank Masaaki Chiwa for his generous help
with the ion chromatography. Petra Dietrich and Gunter Ilgen
helped with the DOC and DON analysis. Neil Cape and David
Fowler have contributed to the discussion. The European
Science foundation provided a travel grant to B.S. Financial
support came from the German Ministry for Research and
Technology (FoÈrdernummer: BMBF No. PT BEO 51-
0339476B). The UK Department of the Environment,
Transport and the Regions fund the experimental facility at
Deepsyke forest (Contract No. EPG1/3/52).
References
Alstad, D.N., Edmunds, G.F. & Weinstein, L.H. (1982) Effects of air
pollutants on insect populations. Annual Review of Entomology, 27,
369±384.
Berendse, F. & Elberse, W.T. (1990) Competition and nutrient
availability in heathland and grassland ecosystems. Perspectives on
Plant Competition (ed. by J. D. Grace and D. Tilman), pp. 93±116.
Academic Press, San Diego.
Bloom, A.J., Chapin, F.J.S. & Mooney, H.A. (1985) Resource
limitation in plants ± an economic analogy. Annual Review of
Ecology and Systematics, 16, 363±392.
Brown, V.C., Ashmore, M.R. & McNeill, S. (1993) Experimental
investigation of the effects of air pollutants on aphids on coniferous
trees. Forstwissenschaftliches Centralblatt, 112, 128±132.
Carter, C.I. (1977) Impact of the green spruce aphid on growth: can a
tree forget its past? Forestry Commission Research and
Development, 116, 1±8.
Carter, C.I. & Nichols, J.F.A. (1988) The Green Spruce Aphid and
Sitka Spruce Provenances in Britain. Forestry Commission,
Edinburgh.
Curtis, P.S. & Wang, X. (1998) A meta-analysis of elevated CO2
effects on woody plant mass, form, and physiology. Oecologia, 113,
299±313.
Day, K.R. (1984) The growth and decline of a population of spruce
aphid (Elatobium abietinum) during a three year study, and changing
pattern of fecundity, recruitment and alary polymorphism in a
Northern Ireland Forest. Oecologia, 64, 118±124.
Day, K.R. & McClean, S.I. (1991) In¯uence of the green spruce aphid
on defoliation and radial stem growth of Sitka spruce. Annals of
Applied Biology, 119, 451±423.
Dixon, A.F.G. (1998) Aphid Ecology. Chapman & Hall, London.
Docherty, M., Salt, D.T. & Holopainen, J.K. (1997) The impacts of
climate change and pollution on forest insect pests. Forests and
Insects (ed. by A. D. Watt, N. E. Stork and M. D. Hunter), pp.
229±247. Chapman & Hall, London.
Dohmen, G.P., McNeill, S. & Bell, J.N.B. (1984) Air pollution
increases Aphid fabae pest potential. Nature, 307, 52±53.
Field, C.B., Chappin, F.S., Matson, P.A. & Mooney, H.A. (1992)
Responses of terrestrial ecosystems to changing atmosphere. Annual
Review of Ecology and Systematics, 23, 201±235.
Fleming, R.A. & Volney, W.J.A. (1995) Effects of climate change on
insect defoliator population processes in Canada's boreal forest:
some plausible scenarios. Water Air and Soil Pollution, 82,
445±454.
Holopainen, J.K., Kainulainen, E., Oksanen, J., Wulff, A. &
KaÈrenlampi, L. (1991) Effect of exposure to ¯uoride, nitrogen
compounds and SO2 on the numbers of spruce shoot aphids on
Norway spruce seedlings. Oecologia, 86, 51±56.
Ingestad, T. & AÊ gren, G.I. (1991) The in¯uence of plant nutrition on
biomass allocation. Ecological Application, 2, 168±174.
Jackson, D.L. & Dixon, A.F.G. (1996) Factors determining the
distribution of the green spruce aphid, Elatobium abietinum, on
young and mature needles of spruce. Ecological Entomology, 21,
358±364.
Kloft, W. & Ehrhardt, P. (1959) Untersuchungen uÈber die SaugtaÈtigkeit
und Schadwirkung der Sitka®chtenlaus Liomyzaphis abietina
(Walk.) (Neomyzaphis abietina Walk.). Phytopathologische
Zeitschrift, 35, 401±410.
Lovett, G.M., Lundberg, S.E., Richter, D.D. & Thomson, D.W. (1985)
The effects of acidic deposition on cation leaching from three
deciduous forest canopies. Canadian Journal of Forest Research,
15, 1055±1060.
Lovett, G.M. & Ruesink, A.E. (1995) Carbon and nitrogen miner-
alization from decomposing gypsy moth frass. Oecologia, 104,
133±138.
Michalzik, B., Dorsch, T. & Matzner, M. (1997) Stability of dissolved
organic nitrogen (DON) and mineral nitrogen in bulk precipitation
and throughfall. Zeitschrift fuÈr P¯anzenernaÈhrung und Bodenkunde,
160, 433±434.
Michalzik, B. & Stadler, B. (2000) Effects of phytophagous insects on
soil solution chemistry: herbivores as switches for the nutrient
dynamics in the soil. Basic and Applied Ecology, 2, 117±123.
MuÈller-Dombois, D. (1992) A global perspective on forest decline.
Environmental Toxicology and Chemistry, 11, 1069±1076.
Parry, W.H. (1974) Damage caused by the green spruce aphid to
Norway and Sitka spruce needles. Annals of Applied Biology, 77,
113±120.
Pedersen, L.B. & Bille-Hansen, J. (1999) A comparison of litterfall and
element ¯uxes in even aged Norway spruce, Sitka spruce and beech
stands in Denmark. Forest Ecology and Management, 114, 55±70.
Sayre, R.G. & Fahey, T.J. (1999) Effects of rainfall acidity and ozone
on foliar leaching in red spruce (Picea rubens). Canadian Journal of
Forest Research, 29, 487±496.
Schowalter, T.D. (2000) Insect Ecology: an Ecosystem Approach.
Academic Press, San Diego.
Schowalter, T.D., Hargrove, W.W. & Crossley, D.A. (1986) Herbivory
in forested ecosystems. Annual Review of Entomology, 31, 177±196.
Schulze, E.-D. (1989) Air pollution and forest decline in a spruce
(Picea abies) forest. Science, 244, 776±783.
Schulze, E.-D., Lange, O. & Oren, R. (1989) Forest Decline and Air
Pollution. A Study of Spruce (Picea abies) on Acid Soils. Ecological
Studies, Vol. 77 Springer, Berlin.
Seaby, D.A. & Mowat, D.J. (1993) Growth changes in 20-year old
Sitka spruce Picea sitchensis after attack by the green spruce aphid
Elatobium abietinum. Forestry, 66, 371±379.
Seastedt, T.R., Crossley, D.A. & Hargrove, W.W. (1983) The effects
of low-level consumption by canopy arthropods on the growth and
nutrient dynamics of black locust and red maple trees in the
Southern Appalachians. Ecology, 64, 1040±1048.
Sheppard, L.J. & Crossley, A. (2000) Responses of a Sitka spruce
ã 2001 Blackwell Science Ltd, Agricultural and Forest Entomology, 3, 253±261
260 Bernhard Stadler et al.260 Bernhard Stadler et al.

ecosystem after 4 years of simulated wet N deposition: effects of
NH4NO3 supplied with and without acidity (H2SO4 pH 2.5).
Sheppard, L.J., Crossley, A., Cape, J.N., Harvey, F., Parrington, J. &
White, C. (1999) Early effects of acid mist on Sitka spruce planted
on acid peat. Phyton, 39, 1±25.
SPSS (1999) SPSS for Windows, Release 9.0.1. SPSS, Chicago.
Stadler, B., Michalzik, B. & MuÈller, T. (1998) Linking aphid ecology
with nutrient ¯uxes in a coniferous forest. Ecology, 79, 1514±1525.
Stadler, B. & MuÈller, T. (2000) Effects of herbivores on epiphytic
micro-organisms in canopies of forest trees. Canadian Journal of
Forest Research, 30, 631±638.
Straw, N.A., Fielding, N.J., Green, G. & Coggan, A. (1998) The impact
of green spruce aphid, Elatobium abietinum (Walker), on the growth
of young Sitka spruce in Hafren Forest, Wales: pattern of defoliation
and effect on shoot growth. Forest Ecology and Management, 104,
209±225.
Tilman, D. (1990) Constzraints and trade-offs: toward a predictive
theory of competition and succession. Oikos, 58, 3±15.
Warrington, S. & Whittaker, J.B. (1990) Interactions between Sitka
spruce, the green spruce aphid, sulphur dioxide pollution and
drought. Environmental Pollution, 65, 363±370.
Watt, A., Fluckinger, W., Leith, I.D. & Lindsay, E. (1998)
Atmospheric pollution, elevated CO2 and spruce aphids. The Green
Spruce Aphid in Western Europe: Ecology, Status, Impacts and
Prospects for Management (ed. by K. R. Day, G. Halldorsson, S.
Harding and N. Straw), pp. 86±96. Forestry Commission Technical
Paper, 24. HMSO, London.
Wilson, J.B. (1988) A review of evidence on the control of shoot: root
ratio, in relation to models. Annals of Botany, 61, 433±449.
Accepted 1 August 2001
ã 2001 Blackwell Science Ltd, Agricultural and Forest Entomology, 3, 253±261
Forest canopies and throughfall ¯uxes 261Forest canopies and throughfall ¯uxes 261