effects of combined entorhinal cortex-hippocampal lesions on locomotor behavior, spontaneous...
TRANSCRIPT

Physiology & Behavior, Vol. 21, pp. 177-187. Pergamon Press and Brain Research Publ., 1978. Printed in the U.S.A.
Effects of Combined Entorhinal Cortex-Hippocampal Lesions on Locomotor
Behavior, Spontaneous Alternation and Spatial Maze Learning in the Rat I
D A N I E L P O R T E R K I M B L E
Universi ty o f Oregon, Eugene , OR 97403
(Rece ived 29 D e c e m b e r 1977)
KIMBLE, D. P. Effects of combined entorhinal cortex-hippocampal lesions on locomotor behavior, spontaneous alterna- tion and spatial maze learning in the rat. PHYSIOL. BEHAV. 21(2) 177-187, 1978.--Male Sprague-Dawley rats with combined lesions to the dorsal and lateral aspects of the entorhinal cortex in one hemisphere and of the contralateral dorsolateral hippocampal formation were compared with both operated and unoperated controls on three different behav- iors, monitored across a 53 day postoperative period. The rats with the combined entorhinal cortex-hippocampal lesions (EH) showed transient hyperactivity in the open field, transient reduction in spontaneous alternation levels in an unbaited T-maze and persistent deficits in learning spatial maze problems. The results of the present experiment are discussed in comparison with those from experiments on rats with bilateral hippocampal lesions and those from experiments on rats with bilateral entorhinal cortex lesions. Although some similarities among these findings suggest that these two brain regions probably function in a coordinated fashion with respect to these behaviors, differences in the various syndromes are also discussed.
Hippocampus Entorhinal cortex Locomotor activity Spontaneous alternation Spatial maze learning
IT HAS been known since the time of Cajal that the entorhinal cortex and the hippocampai formation in the rat were closely related neuroanatomically [2]. More recent anatomical investigations have shown that both the hip- pocampus proper and the dentate gyrus receive major affe- rents from the lateral and medial entorhinal cortex [5, 14, 16, 17, 27, 28]. Efferents from the hippocampus to entorhinal cortex have also been demonstrated [4]. Electrophysiologi- cal evidence indicates that the afferent system from the entorhinal cortex to the ipsilateral dentate gyrus is monosynaptic.
Recently, a great deal of interest has been generated in these entorhinal-hippocampal connections by the dis- coveries of Cotman, Lynch, Steward and others that follow- ing lesions to the entorhinal cortex and subsequent denerva- tion of the dentate granule cells, an impressive amount of collateral sprouting or "reactive synaptogenesis" occurs in the dentate gyrus and hippocampus [14, 16, 17, 27, 28, 30]. Much of this growth process is due to the formation of new synapses: "about one-half to two-thirds of the reacquired synaptic connections are probably formed de novo" ([17] p. 38). This reactive synaptogenesis is produced by intact affe- rents from a variety of sources, including the septal nuclei, hippocampal commissural and associational afferents, and a normally sparse contralaterai path from the entorhinal cortex
[27,28]. In addition, some contribution of fibers from outside the brain may be involved. Loy and Moore have reported that following anterior hippocampal lesions, there is an anomalous innervation of hippocampal tissue via collateral sprouting from terminal axons of neurons originating in the superior cervical ganglion of the sympathetic nervous sys- tem [13].
While a considerable amount of anatomical research suggests that the hippocampal formation and the entorhinal cortex might form a functional unit, until recently there has been very little relevant behavioral evidence. Many reports have appeared concerning the behavioral role of the hip- pocampus, but there has not been a similar effort with regard to the entorhinal cortex. Recent reports, however, suggest that a similarity may exist between the effects of hippocam- pal lesions and those of the entorhinal cortex. Both hip- pocampal and entorhinal cortex lesions produce deficits in the acquisition of spatial maze problems [6, 7, 10, 15, 19, 22], both produce hyperactivity and both reduce levels of spon- taneous and learned alternation [8, 9, 11, 12, 18, 26, 29]. There are conflicting reports, however, as to whether all of these behavioral changes are permanent or transient.
If the hippocampal formation and the entorhinal cortex do form a functional unit with respect to the performance of these behaviors, then severing the neural connections be-
q would like to thank Ruth BreMiller and Susan Anderson for the histology.
Copyr igh t © 1978 Brain Research Pub l i ca t ions Inc.--0031-9384/78/0801-0177502.00/0

178 KIMBLE
tween them should produce a behavioral syndrome similar to or indistinguishable from those produced by bilateral hip- pocampal or bilateral entorhinal cortex lesions, even if the animal still possesses one intact hippocampus and one intact entorhinal cortex. The present report presents data gathered across a 53 day postoperative period on the effects of such a disconnection, in which lesions were produced in the hip- pocampal formation of one hemisphere and the entorhinal cortex in the other hemisphere.
METHOD
Animals
Animals were 24 male Sprague-Dawley albino rats (CD, randombred) obtained from the Charles River Co., Wil- mington, MA. They were maintained in individual cages on a 12-hr diurnal light cycle. All rats were about 120 days old and weighed about 400 g at the time of surgery. Except for times specified in the procedure section, food and water were available on an ad lib basis.
Apparatus
Activity was measured in a wooden box measuring 78.7 x 78.7 cm with 11.4 cm high walls. The floor was marked off into 36 equal sized squares. The field was closed with a sheet of clear Plexiglas. The same apparatus, when combined with wooden inserts, a start box, and a goal box, served as the Hebb-Wilfiams maze apparatus [3]. Nine different maze pat- terns selected from the Rabinovitch-Rosvold series were used [23]. The start box and goal box were identical in size, each measuring 30 cm in length, 13.5 cm in width, and 15 cm in height. Clear Plexiglas served as the top for both the start and goal boxes. Spontaneous alternation was evaluated in a wooden T-maze made up of a 56.3 × 11.4 cm stem and two 31.7 x 24.1 cm goal arms, set at right angles to the stem. The goal arms were equipped with Plexiglas guillotine-type doors. The walls of the maze were 18 cm high. Both the T-maze and the Hebb-Williams apparatus were painted grey.
Procedure: Preoperative Training
All animals were gentled for several days after their arri- val until they could be handled freely without gloves. Prior to surgery, all rats were observed during a five min period in the open field, and the number of squares entered in this time recorded. An entry was counted if a rat placed its two front paws in a square. This activity measure was the basis for the construction of two groups equivalent in preoperative activ- ity. The day following the activity measurements, all rats were placed on a 24 hr water deprivation schedule. Begin- ning one week later they were given six consecutive days of practice problems in the Hebb-Williams apparatus, using practice problems A-F as outlined in the Rabinovitch- Rosvold protocol. At the end of this pretraining, all rats were running five consecutive trials in less than 60 sec.
Surgery
Anesthesia was induced by intraperitoneal injections of sodium pentobarbital (Nembutal) in doses of 50 mg/kg. At- ropine sulphate, in doses of approximately 0.04 mg/rat was also administered to reduce mouth and throat secretions. Twelve rats received aspiration lesions to the entorhinal cor- tex in one cerebral hemisphere (the right hemisphere in six animals, the left hemisphere in the other six) and were then
given aspiration lesions to the dorsolateral hippocampal for- mation in the contralateral hemisphere, using a technique described previously [7]. The remaining twelve animals were divided into unoperated controls (UC, N--6) and operated controls. The operated controls (OC, N=6) received either unilateral entorhinal cortex lesions or a unilateral dorsolat- eral hippocampal lesion. All operations were performed in one stage using low-power magnification. One rat from the combined entorhinal-hippocampal (EH) group showed ab- normal posture and movements postoperatively and was dis- carded. The other 23 animals survived the surgery well, showed no sensory or motor abnormalities, and completed all phases of the experiment.
Postoperative Testing
Postoperative testing began on postoperative Day 2 and continued through Day 53. On Day 2 rats were placed, one at a time, in the center of the open field and the number of squares entered within 5 rain was recorded. The floor was wiped with damp paper toweling after each test. This activity measurement was taken for each rat on postoperative Days 2, 4, 9 and 14. Since the main focus of the experiment was on maze learning ability, it was necessary to place the animals on a water deprivation schedule at various times throughout the testing period. All rats were placed on a 24 hr water deprivation schedule (10 rain access to water each day) on Postoperative Day 3, kept on this schedule through the test- ing on Day 13, replaced on the deprivation schedule on Day 47, and removed following testing on Day 51. At all other times they had free access to water in their home cages. Food was constantly available on an ad lib basis.
Each rat was tested for its tendency to spontaneously alternate its choices in the T-maze. Each rat was released from the start box into the main stem and allowed up to 3 rain to enter one of the two goal arms. A guillotine-type door was then gently closed behind the rat, 20 sec was allowed to elapse, and the rat was replaced in the start box for the next trials. Three such trials were run on Postoperative Days 3, 5, 10, 15 and 53. The tendency to alternate the previous goal arm choice on the second and third trials of each day was recorded. Thus, for a given day, a rat could score 0, 50 or 100% alternation.
Spatial maze learning capacity was tested using nine different maze problems selected from the Rabinovitch- Rosvold series. Three maze "sets" were constructed, each set composed of one "easy" , one "moderately difficult" and one "difficult" maze problem, as defined by the perform- ance of normal rats used by Rabinovitch and Rosvold [23]. Set 1 consisted of maze problems 1, 5 and 11; Set 2 consisted of maze problems 2, 6 and 10; Set 3 consisted of problems 4, 7 and 12. Set 1 was administered to all animals on Postoperative Days 6, 7 and 8. Set 2 was administered on Postoperative Days 11, 12 and 13. Set 2 was administered on Postoperative Days 49, 50 and 51. These times were chosen to correspond to periods before, during and after the peak periods of synaptic reorganization in the dentate gyrus fol- lowing entorhinal lesions [27]. Each rat was run seven trials on each problem, with one maze problem being presented each day as schedule. A water reward was available in the goal box. Rats were placed into the start box and released into the maze. The rat 's path through the maze was recorded on a data sheet with the floor plan of that maze problem, and the number of error lines crossed later counted from the protocol. An error was counted if a rat's two front paws (or,

COMBINED CORTEX-HIPPOCAMPAL LESIONS 179
more rarely, one front and one rear paw on the same side) crossed an " e r ro r " line.
In summary, the postoperative schedule for each rat was: Day 2 Open field activity Day 3 Spontaneous alternation. Placed on 24 hr water
deprivation schedule following testing Day 4 Open field activity Day 5 Spontaneous alternation Day 6 Maze problem 1 Day 7 Maze problem 5 Day 8 Maze problem 11 Day 9 Open field activity Day 10 Spontaneous alternation Day 11 Maze problem 2 Day 12 Maze problem 6 Day 13 Maze problem 10. Removed from water
deprivation following testing Day 14 Open field activity Day 15 Spontaneous alternation
Day 47 Placed on 24 hr water deprivation schedule Day 49 Maze problem 4 Day 50 Maze problem 7 Day 51 Maze problem 12. Removed from water
deprivation following testing Day 52 Spontaneous alternation
RESULTS
Open Field Activity
A transient but significant increase in activity was ob- served in the EH animals. No significant postoperative change was seen in either the operated or unoperated con- trols. Table 1 shows these results. Group differences were compared using the Mann-Whitney U-test [25].
TABLE 1 OPEN FIELD ACTIVITY. SQUARES ENTERED/SESSION
Postoperative Day Hippocampal- Controls Entorhinal
2 170 132t 4 195 163t 9 292 1955
14 221 180"
*n.s. ,p =0.02 ,p =o.oo2
The EH rats were clearly more active on all the tests, but this difference failed to reach statistical significance on Day 14. Rats from all groups showed slight increases in activity on the two testing days in the water deprivation period (Days 4 and 9), but there was no significant differential effect, as the EH rats increased their activity approximately 25% dur- ing deprivation, compared with a 17% increase for control animals. The largest activity difference was seen on Postoperative Day 9. There is, of course, a possible con- founding of the changing deprivation conditioning and activ- ity levels, although certain results seem clear. The hyperac- tivity observed in the EH rats on Postoperative Day 2 oc-
curred under ad lib conditions. This relative hyperactivity continued on Days 4 and 9 (under water deprivation). The absence of statistically significant differences on Day 14 may, however, be related to the return to ad lib conditions 24 hr previously. Previous research [9] has shown that a similar open field hyperactivity following bilateral hippocampal le- sions appears around Day 5 and returns to normal at about Day 11-14.
Spontaneous Alternation
Due to the design of the experiment, spontaneous alter- nation, like the open field observations, was evaluated under both ad lib and water deprivation conditions. Observations on Postoperative Days 3, 15 and 53 were under ad lib condi- tions, while observations on Days 5 and 10 were taken while the animals were on a 24 hr water deprivation schedule. Differences in percent alternation were evaluated using a Mann-Whitney U-test. The controls showed significantly higher rates of alternation on Postoperative Days 3, 10 and 15 (p =0.05), while the difference between controls and the EH rats narrowly missed the 0.05 significance level on Day 5. On Day 53, however, the EH rats alternated at a 77% level, slightly higher (although statistically insignificant) than the 71% rate shown by the control rats. It is, of course, highly possible that the results on this day represent a spuri- ously low level of alternation by the control rats. Examina- tion of the data show that of 11 EH rats alternated at a 100% level, as compared with 7 of 12 control rats. Thus, it would appear that the deficit of EH rats in spontaneous al- ternation was not present on this day. Whether or not this represents a permanent " recovery" of normal spontaneous alternation tendencies in these rats cannot be reliably de- termined on the basis of present data. However , earlier work [9] also suggests that some recovery of normal alternation behavior may occur in rats with bilateral hippocampal le- sions. Further data on this question will be necessary before firm conclusions regarding " r ecove ry" can be made.
TABLE 2 SPONTANEOUS ALTERNATION. MEAN PERCENT ALTERNATION
(PERCENT FIRST-TRIAL ALTERNATION)
Postoperative Day Hippocampal- Controls Entorhinal
3 20 (20) 59 (45)t 5 55 (55) 71 (83)*
10 45 (45) 83 (83)t 15 55 (64) 87 (83)t 53 77 (72) 71 (67)*
tp =0.05 "*=n.s.
Spatial Maze Learning
The EH rats displayed a significant and persistent deficit in learning the spatial maze problems at all three testing ses- sions. See Table 3 and Figs. 1-9. The EH rats made about twice as many errors as the control animals on each set of maze problems. The total number of errors on each set was significantly higher for the EH rats as evaluated by Mann- Whitney U-tests (p=0.002, two-tailed). While both groups
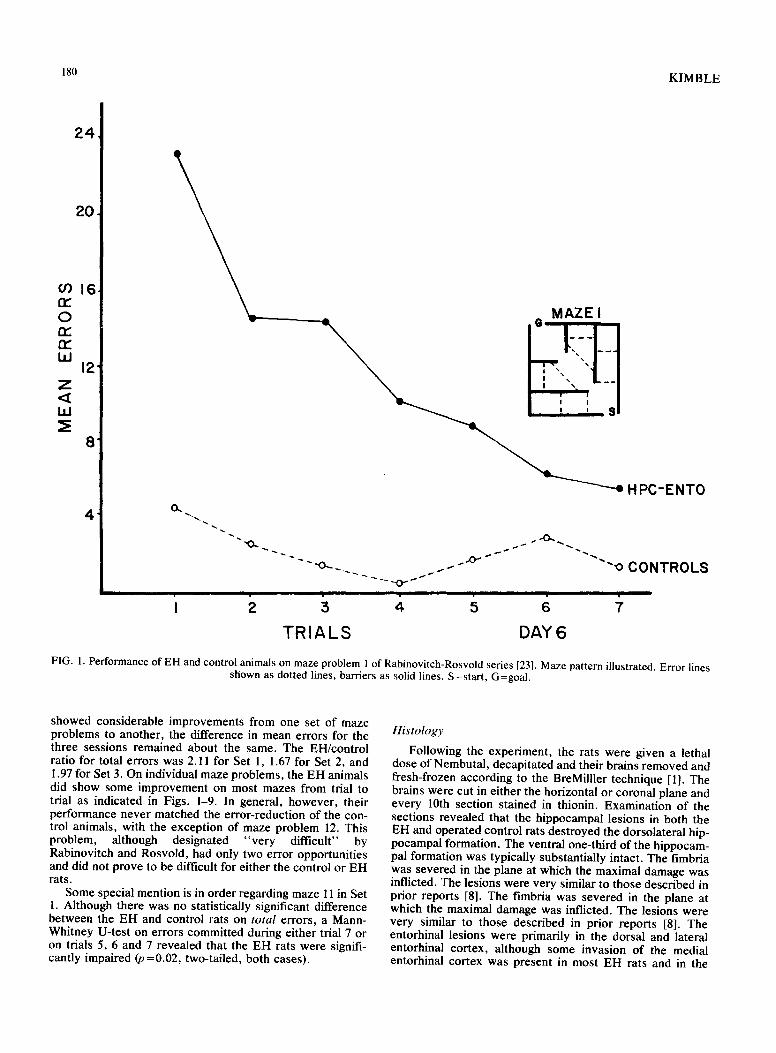
180 KIMBLE
24
20
o 0 1 6 fr-
O n,"
I , I 12
Z < LI.I
8
4 O~
~ . . . , 0 ~ i i i
HPC-ENTO
""°"" "~"oCONTROLS
T R I A L S DAY6
FIG. 1. Performance of EH and control animals on maze problem 1 of Rabinovitch-Rosvold series [23]. Maze pattern illustrated. Error lines shown as dotted lines, barriers as solid lines. S=start, G=goal.
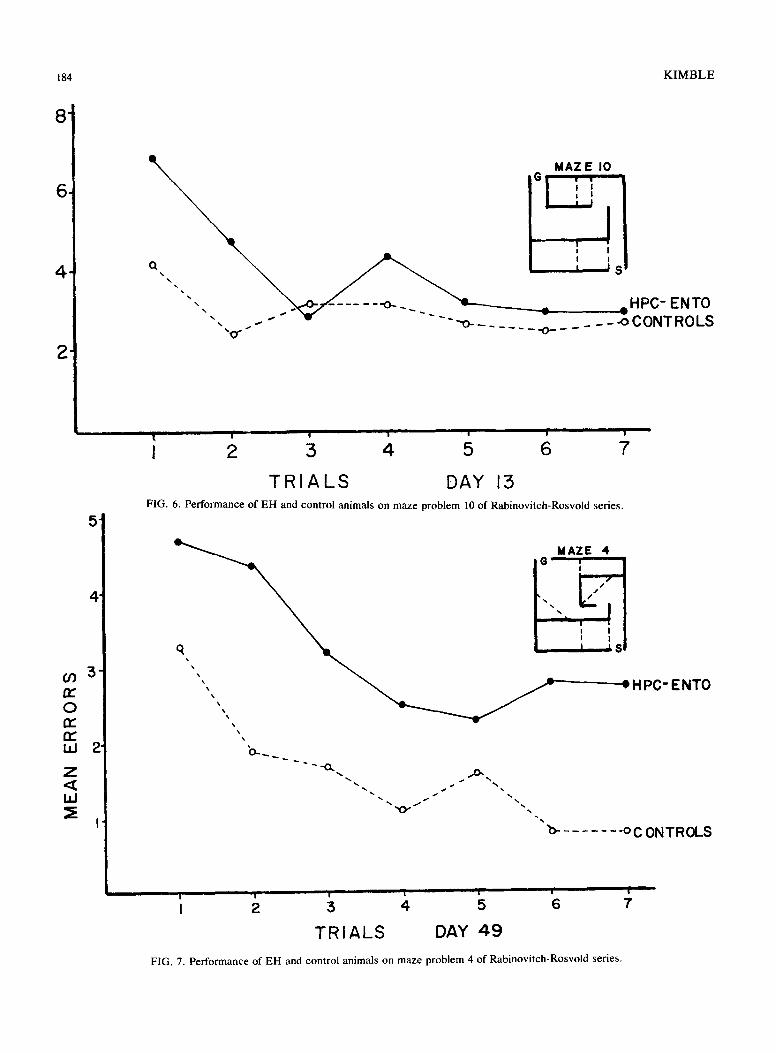
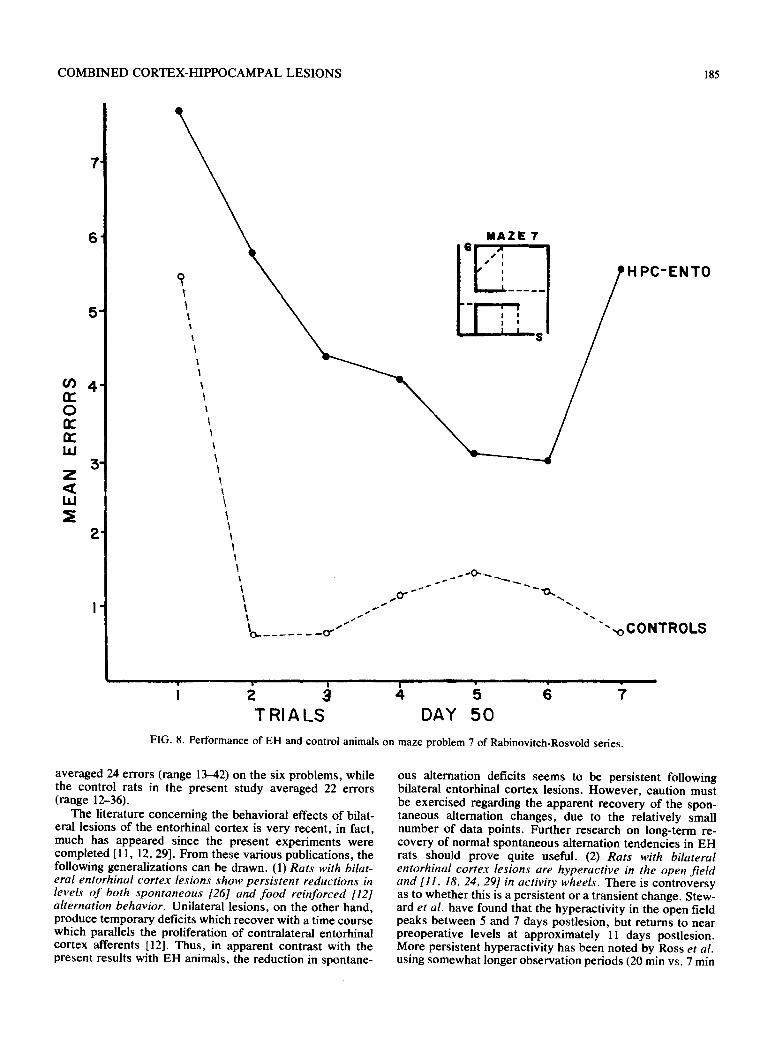
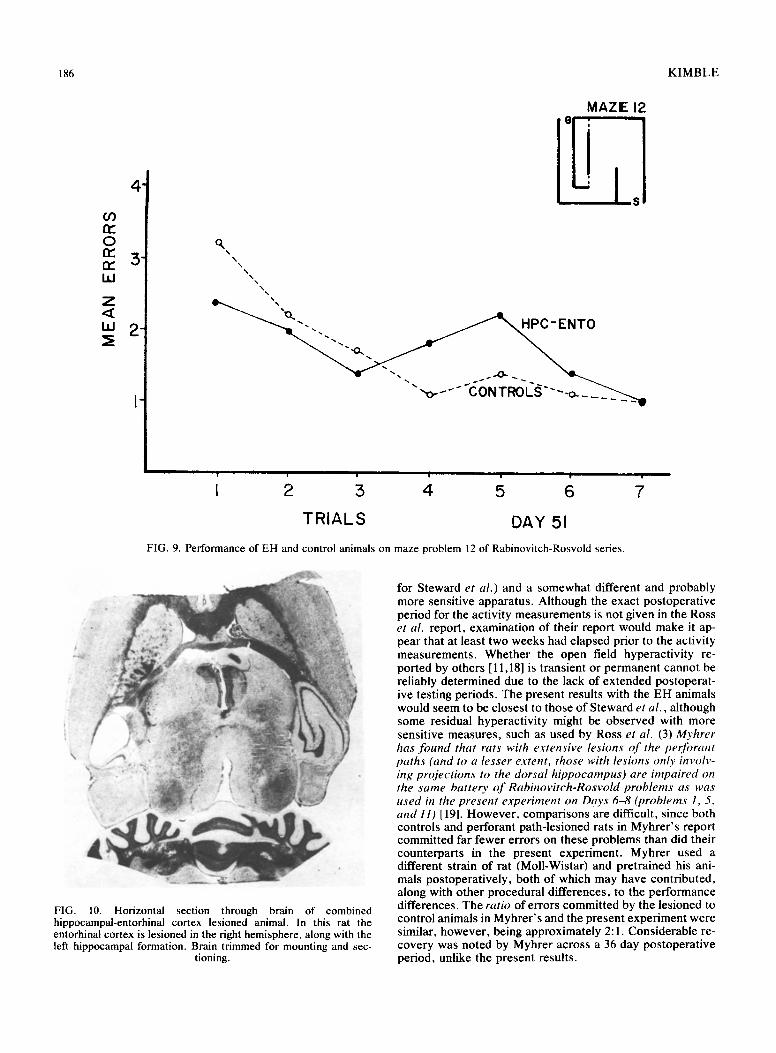
showed considerable improvements from one set of maze problems to another, the difference in mean errors for the three sessions remained about the same. The EH/control ratio for total errors was 2.11 for Set 1, 1.67 for Set 2, and 1.97 for Set 3. On individual maze problems, the EH animals did show some improvement on most mazes from trial to trial as indicated in Figs. 1-9. In general, however, their performance never matched the error-reduction of the con- trol animals, with the exception of maze problem 12. This problem, although designated "very difficult" by Rabinovitch and Rosvold, had only two error opportunities and did not prove to be difficult for either the control or EH rats.
Some special mention is in order regarding maze 11 in Set 1. Although there was no statistically significant difference between the EH and control rats on total errors, a Mann- Whitney U-test on errors committed during either trial 7 or on trials 5, 6 and 7 revealed that the EH rats were signifi- cantly impaired (p =0.02, two-tailed, both cases).
Histology
Following the experiment, the rats were given a lethal dose of Nembutal, decapitated and their brains removed and fresh-frozen according to the BreMilller technique [1]. The brains were cut in either the horizontal or coronal plane and every 10th section stained in thionin. Examination of the sections revealed that the hippocampal lesions in both the EH and operated control rats destroyed the dorsolateral hip- pocampal formation. The ventral one-third of the hippocam- pal formation was typically substantially intact. The fimbria was severed in the plane at which the maximal damage was inflicted. The lesions were very similar to those described in prior reports [8]. The fimbria was severed in the plane at which the maximal damage was inflicted. The lesions were very similar to those described in prior reports [8]. The entorhinal lesions were primarily in the dorsal and lateral entorhinal cortex, although some invasion of the medial entorhinal cortex was present in most EH rats and in the
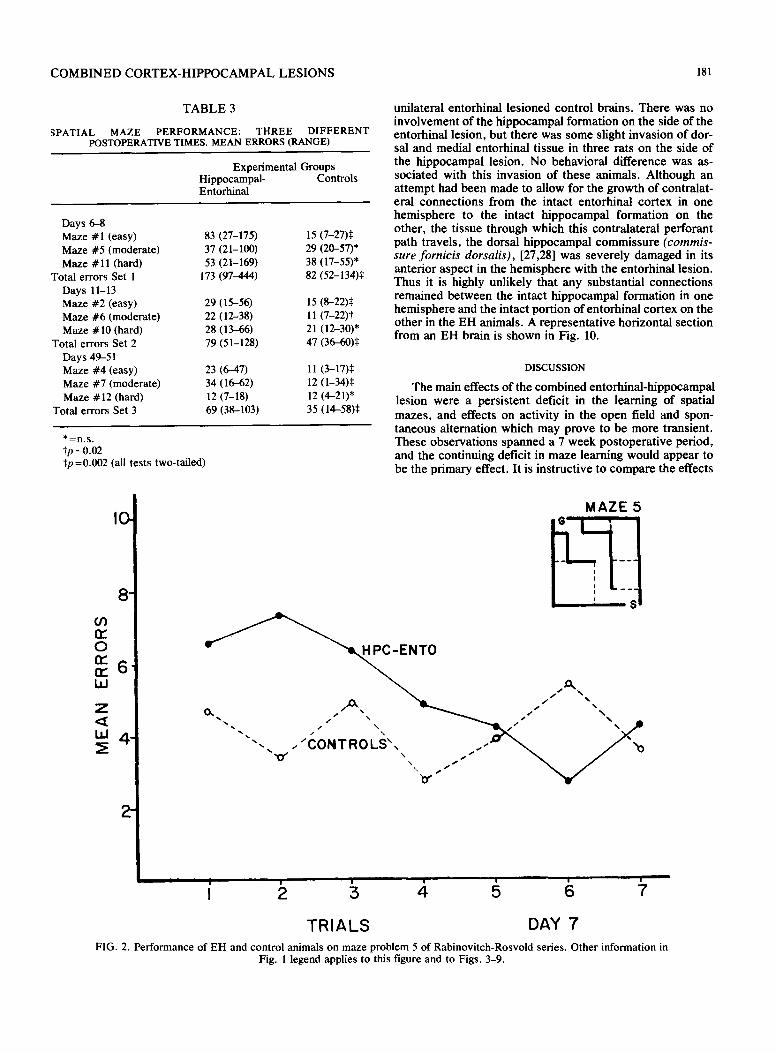
COMBINED CORTEX-HIPPOCAMPAL LESIONS 181
TABLE 3
SPATIAL MAZE PERFORMANCE: THREE DIFFERENT POSTOPERATIVE TIMES. MEAN ERRORS (RANGE)
Experimental Groups Hippocampal- Controls Entorhinal
Days 6-8 Maze #1 (easy) 83 (27-175) 15 (7-27)$ Maze #5 (moderate) 37 (21-100) 29 (20-57)* Maze #11 (hard) 53 (21-169) 38 (17-55)*
Total errors Set I 173 (97 ~aa) 82 (52-134):~ Days 11-13 Maze #2 (easy) 29 (15-56) 15 (8-22):~ Maze #6 (moderate) 22 (12-38) 11 (7-22)1" Maze #10 (hard) 28 (13-66) 21 (12-30)*
Total errors Set 2 79 (51-128) 47 (36-60)~: Days 49-51 Maze #4 (easy) 23 (6-47) 11 (3-17):~ Maze #7 (moderate) 34 (16-62) 12 (1-34):~ Maze #12 (hard) 12 (7-18) 12 (4-21)*
Total errors Set 3 69 (38-103) 35 (14-58)¢
* : n . s . tp =0.02 Sp =0.002 (all tests two-tailed)
unilateral entorhinal lesioned control brains. There was no involvement of the hippocampal formation on the side of the entorhinal lesion, but there was some slight invasion of dor- sal and medial entorhinal tissue in three rats on the side of the hippocampal lesion. No behavioral difference was as- sociated with this invasion of these animals. Although an attempt had been made to allow for the growth of contralat- eral connections from the intact entorhinal cortex in one hemisphere to the intact hippocampal formation on the other, the tissue through which this contralateral perforant path travels, the dorsal hippocampal commissure (commis- sure fornicis dorsalis), [27,28] was severely damaged in its anterior aspect in the hemisphere with the entorhinal lesion. Thus it is highly unlikely that any substantial connections remained between the intact hippocampal formation in one hemisphere and the intact portion of entorhinal cortex on the other in the EH animals. A representative horizontal section from an EH brain is shown in Fig. 10.
DISCUSSION
The main effects of the combined entorhinal-hippocampal lesion were a persistent deficit in the learning of spatial mazes, and effects on activity in the open field and spon- taneous alternation which may prove to be more transient. These observations spanned a 7 week postoperative period, and the continuing deficit in maze learning would appear to be the primary effect. It is instructive to compare the effects
03
O 13E t'r- IM
Z
LU
I0-
.
6
4
2"
M A Z E 5
e ~ ~ ~ - E N T O
TRIALS DAY 7' FIG. 2. Performance of EH and control animals on maze problem 5 of Rabinovitch-Rosvold series. Other information in
Fig. l legend applies to this figure and to Figs. 3-9.
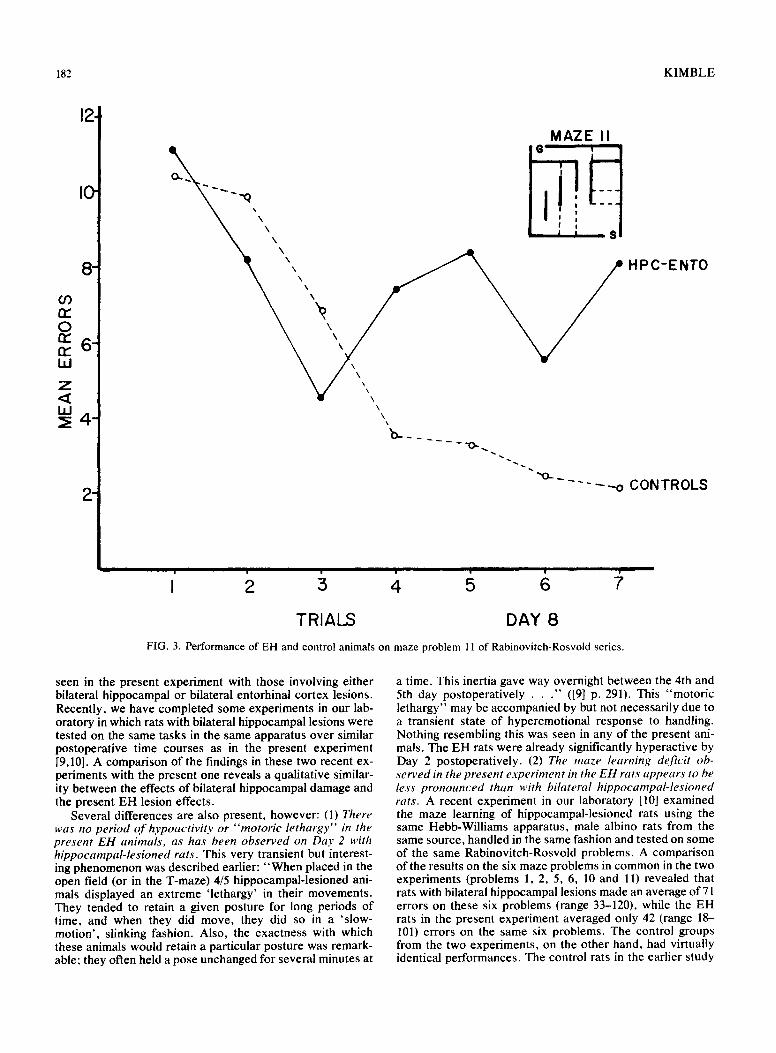
182 KIMBLE
0 re" n-
Z < LLI
12.
I0"
8 -
.
.
2 -
M A Z E I I . I G , '
'~N I l
W \ \
\ k
H P C - E N T O
- - o C O N T R O L S
i i i i i i i i i i i i i i
I z i 4 5 6 7
TRIALS DAY 8 FIG. 3. Performance of EH and control animals on maze problem 11 of Rabinovitch-Rosvold series.
seen in the present experiment with those involving either bilateral hippocampal or bilateral entorhinal cortex lesions. Recently, we have completed some experiments in our lab- oratory in which rats with bilateral hippocampal lesions were tested on the same tasks in the same apparatus over similar postoperative time courses as in the present experiment [9,10]. A comparison of the findings in these two recent ex- periments with the present one reveals a qualitative similar- ity between the effects of bilateral hippocampal damage and the present EH lesion effects.
Several differences are also present, however: (1) There was no period o f hypoactivity or "motoric lethargy" in the present EH animals, as has been observed on Day 2 with hippocampal-lesioned rats. This very transient but interest- ing phenomenon was described earlier: "When placed in the open field (or in the T-maze) 4/5 hippocampal-lesioned ani- mals displayed an extreme 'lethargy' in their movements. They tended to retain a given posture for long periods of time, and when they did move, they did so in a 'slow- motion', slinking fashion. Also, the exactness with which these animals would retain a particular posture was remark- able; they often held a pose unchanged for several minutes at
a time. This inertia gave way overnight between the 4th and 5th day postoperatively . . ." ([9] p. 291). This "motoric lethargy" may be accompanied by but not necessarily due to a transient state of hyperemotional response to handling. Nothing resembling this was seen in any of the present ani- mals. The EH rats were already significantly hyperactive by Day 2 postoperatively. (2) The maze learning deficit ob- served in the present experiment in the EH rats appears to be less pronounced than with bilateral hippocampal-lesioned rats. A recent experiment in our laboratory [10] examined the maze learning of hippocampal-lesioned rats using the same Hebb-Williams apparatus, male albino rats from the same source, handled in the same fashion and tested on some of the same Rabinovitch-Rosvold problems. A comparison of the results on the six maze problems in common in the two experiments (problems 1, 2, 5, 6, 10 and l l ) revealed that rats with bilateral hippocampal lesions made an average of 71 errors on these six problems (range 33-120), while the EH rats in the present experiment averaged only 42 (range 18- 101) errors on the same six problems. The control groups from the two experiments, on the other hand, had virtually identical performances. The control rats in the earlier study
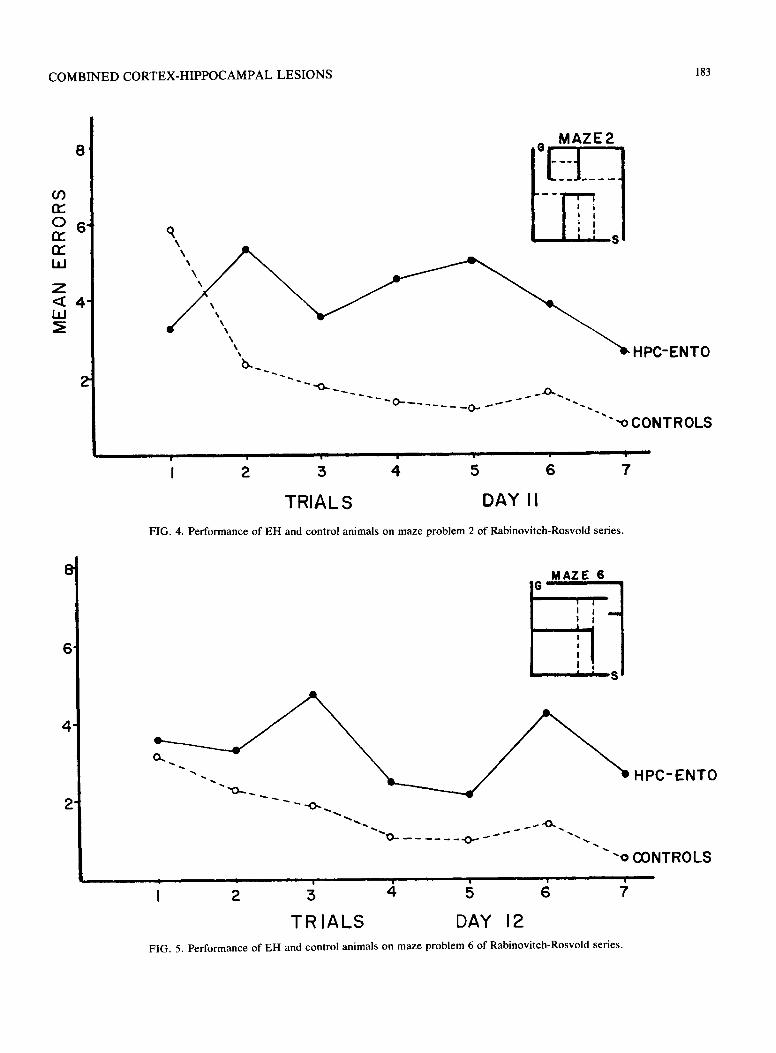
C O M B I N E D C O R T E X - H I P P O C A M P A L L E S I O N S 183
,
03 n." 0 6" r r r r IM
Z < 4 " LLI
2
M A Z E 2
q
~", HPC-ENTO
" ' " ' " o C O N T R O L S
i i i • • i
I 2 3 4 5 6 7
TRIAL S DAY II
FIG. 4. Performance of E H and control animals on maze problem 2 of Rabinovi tch-Rosvold series.
8
,
.
.
MAZE ,,6,
J I !
' i !
! I I
. . . . . .O .~ ~ ~ * . ~ .~.
-o CONTROLS
HPC-ENTO
| i | i
TRIALS DAY 12 FIG. 5. Performance of EH and control animals on maze problem 6 of Rabinovi tch-Rosvold series.
184 K I M B L E
.
.
.
.
03 n," 0 nr' tY i,I
Z
ILl
.
.
M A Z E I0,,
1 _ HPC- ENTO
--~. . . . . . . . -o CONTROLS
I III I i I I I I
I 2 3 4 5 6 7
T R I A L S D A Y 1 3
FIG. 6. Performance of EH and control animals on maze problem 10 of Rabinovitch-Rosvold series.
M A Z E 4
q, ~ L I • Sl
' " ' , ,, ~ ~r HPC-ENTO
,w
'o-_ ~ ",O..
"~ ' '~" ~ . , . s S ~ " ~ '% "~
% /
'b oCONTROLS
i i I i I I I
TRIALS DAY 49 FIG. 7. Performance of EH and control animals on maze problem 4 of Rabinovitch-Rosvold series.
COMBINED CORTEX-HIPPOCAMPAL LESIONS 185
.
M A Z E 7
i .....
6
4"
o
3" Z
bJ
2'
% 1
. . 0 " " " 0 . . ,
ENTO
"",,oCONTROLS
- ~ e '- ' ~ " '~ i TRIALS DAY 50
FIG. 8. Performance of EH and control animals on maze problem 7 of Rabinovitch-Rosvold series.
averaged 24 errors (range 13-42) on the six problems, while the control rats in the present study averaged 22 errors (range 12-36).
The literature concerning the behavioral effects of bilat- eral lesions of the entorhinal cortex is very recent, in fact, much has appeared since the present experiments were completed [11, 12, 29]. From these various publications, the following generalizations can be drawn. (1) Rats with bilat- eral entorhinal cortex lesions show persistent reductions in levels o f both spontaneous [261 and food reinforced [12] alternation behavior. Unilateral lesions, on the other hand, produce temporary deficits which recover with a time course which parallels the proliferation of contralateral entorhinal cortex afferents [12]. Thus, in apparent contrast with the present results with EH animals, the reduction in spontane-
ous alternation deficits seems to be persistent following bilateral entorhinal cortex lesions. However , caution must be exercised regarding the apparent recovery of the spon- taneous alternation changes, due to the relatively small number of data points. Further research on long-term re- covery of normal spontaneous alternation tendencies in EH rats should prove quite useful. (2) Rats with bilateral entorhinal cortex lesions are hyperactive in the open f ield and [11, 18, 24, 29] in activity wheels. There is controversy as to whether this is a persistent or a transient change. Stew- ard et al. have found that the hyperactivity in the open field peaks between 5 and 7 days postlesion, but returns to near preoperative levels at approximately 11 days postlesion. More persistent hyperactivity has been noted by Ross et al. using somewhat longer observation periods (20 min vs. 7 min
186 KIMBLE
03 nr" 0 n," n,- W
Z <IZ W ~E
.
.
.
i.
MAZE 12
o, N
\ \
%\
" o - - " CONTROLS "--o-___
HI , , , I , ' I i
I 2 5 4 5 6 7
TRIALS DAY 51
FIG. 9. Performance of EH and control animals on maze problem 12 of Rabinovitch-Rosvold series.
\
FIG. 10. Horizontal section through brain of combined hippocampal-entorhinal cortex lesioned animal. In this rat the entorhinal cortex is lesioned in the right hemisphere, along with the left hippocampal formation. Brain trimmed for mounting and sec-
tioning.
for Steward et al.) and a somewhat different and probably more sensitive apparatus. Although the exact postoperative period for the activity measurements is not given in the Ross et al. report, examination of their report would make it ap- pear that at least two weeks had elapsed prior to the activity measurements. Whether the open field hyperactivity re- ported by others [11,18] is transient or permanent cannot be reliably determined due to the lack of extended postoperat- ive testing periods. The present results with the EH animals would seem to be closest to those of Steward et al., although some residual hyperactivity might be observed with more sensitive measures, such as used by Ross et al. (3) Myhrer has found that rats with extensive lesions o f the perforant paths (and to a lesser extent, those with lesions only involv- ing projections to the dorsal hippocampus) are impaired on the same battery o f Rabinovitch-Rosvold problems as was used in the present experiment on Days 6--8 (problems 1, 5, and 11) [19]. However, comparisons are difficult, since both controls and perforant path-lesioned rats in Myhrer's report committed far fewer errors on these problems than did their counterparts in the present experiment. Myhrer used a different strain of rat (Moll-Wistar) and pretrained his ani- mals postoperatively, both of which may have contributed, along with other procedural differences, to the performance differences. The ratio of errors committed by the lesioned to control animals in Myhrer's and the present experiment were similar, however, being approximately 2:1. Considerable re- covery was noted by Myhrer across a 36 day postoperative period, unlike the present results.

C O M B I N E D C O R T E X - H I P P O C A M P A L L E S I O N S 187
Thus , the E H rats in the present exper iment seem to be more like rats with bilateral h ippocampal lesions with re- spect to the effect on spontaneous al ternat ion and maze- learning difficulty and more like entorhinal- les ioned rats with respect to open field activity. The similarity o f these var ious syndromes supports the neuroana tomica l data linking the entorhinal cor tex with the h ippocampal formation. Exis t ing differences would argue, however , that these two brain re- gions, while funct ional ly linked, are not perfect ly redundant . In part icular , the more p ronounced and apparent ly more pers is tent deficit in learning spatial maze problems fol lowing bilateral h ippocampal lesions [6, 7, 10] as contras ted with
e i ther bilateral entorhinal cor tex lesions [19] or in the present E H animals, suggests that the h ippocampal formation may in fact play a part icularly significant role in some aspect of spatial maze learning, such as spatial or ientat ion and/or spa- tial memory , as had been proposed by several o ther inves- t igators [20, 21, 22]. The integrity of some substantial amount of entorhinal cor tex may be necessary , on the o ther hand, for normal spontaneous al ternat ion tendencies , since both rats with bilateral h ippocampal lesions [9] and perhaps the E H rats in the present exper iment , but not rats with bilateral entorhinal cor tex lesions [12] r e cove r their t endency to spontaneous ly al ternate choices in a normal fashion.
REFERENCES
1. BreMiiler, R. A. A rapid technique of preparing frozen sections of small brains. Physiol. Behav. 6: 463-464, 1971.
2. Cajal, S. Ramon Y. Histologie du systeme nerveux de I'homme et des vertebres. (Vol. 2)Paris: Maloine, 1911.
3. Hebb, D. O. and K. Williams. A method of rating animal intelli- gence. J. Genet. Psychol. 34: 59--65, 1946.
4. Hjorth-Simonsen, A. Hippocampal efferents to the ipsilateral entorhinal area: An experimental study in the rat. J. comp. Neurol. 142: 417-428, 1971.
5. Hjorth-Simonsen, A. Projection of the lateral part of the entorhinal area to the hippocampus and fascia dentata. J. comp. Neurol. 146: 219-231, 1972.
6. Kaada, B. R., E. W. Rasmussen and O. Kviem. Effects of hip- pocampal lesions on maze learning and retention in rats. Expl Neurol. 3: 333-355, 1961.
7. Kimble, D. P. The effects of bilateral hippocampal lesions in rats. J. cornp, physiol. Psychol. 56: 273-283, 1963.
8. Kimble, D. P. Choice behavior in rats with hippocampal le- sions. In: The Hippocampus, a Comprehensive Treatise, edited by R. L. Isaacson and K. H. Pribram. Plenum Press, 1975.
9. Kimble, D. P. Changes in behavior of hippocampal-lesioned rats across a 6-week postoperative period. Physiol. Psychol. 4: 289--293, 1976.
10. Kimble, D. P. and E. Dannen. Persistent spatial maze-learning deficits in hippocampal-lesioned rats across a 7-week postoperative period. Physiol. Psychol. 5: 409-413, 1977.
11. Kohler, C. and H. Sundberg. Locomotor activity and explora- tory behavior after medial entorhinal cortex lesions in the albino rat. Behav. Biol. 20: 419--432, 1977.
12. Loesche, J. and O. Steward. Behavioral correlates of denerva- tion and reinnervation of the hippocampal formation of the rat: Discovery of alternation performance following unilateral entorhinal lesions. Brain Res. Bull. 2: 31-39, 1977.
13. Loy, R. and R. Y. Moore. Anomalous innervation of the hip- pocampal formation by peripheral sympathetic axons following mechanical injury. Soc. Neurosci. Abst. 3: 428, 1977.
14. Lynch, G. and C. W. Cotman. The hippocampus as a model for studying anatomical plasticity in the adult brain. In: The Hip- pocampus, edited by R. L. Isaacson and K. H. Pribram. New York: Plenum, 1975, pp. 123-154.
15. Madsen, M. C. and D. P. Kimble. The maze behavior of hip- pocampectomized rats under massed and distributed traits. Psychon. Sci. 3: 193-194, 1965.
16. Matthews, D., C. Cotman and G. Lynch. An electron micro- scopic study of lesion-induced synaptogenesis in the dentate gyrus of the adult rat. I. Magnitude and time course of degener- ation. Brain Res. 115: 1-21, 1976.
17. Matthews, D., C. Cotman and G. Lynch. An electron micro- scopic study of lesion-induced synaptogenesis in the dentate gyrus of the adult rat. II. Reappearance of morphologically normal synaptic contacts. Brain Res. 115: 23--41, 1976.
18. Myhrer, T. Locomotor and avoidance behavior in rats with par- tial or total hippocampal perforant paths sections. Physiol. Be- hay. 15: 217-224, 1975.
19. Myhrer, T. Maze performance in rats with hippocampal perfor- ant path lesions: Some aspects of functional recovery. Physiol. Behav. 15" 433--437, 1975.
20. O'Keefe, J. and J. Dostrovsky. The hippocampus as a spatial map. Preliminary evidence from unit activity in the freely- moving rat. Brain Res. 34: 171-175, 1971.
21. O'Keefe, J. and L Nadel. Maps in the brain. New Scientist. 62: 749-751, 1974.
22. Olton, D. S. Spatial memory. Sci. Am. 236: 82-98, 1977. 23. Rabinovitch, M. S. and H. E. Rosvold. A closed-field intelli-
gence test for rats. Can. J. Psychol. 5: 122-128, 1951. 24. Ross, J. F., L. L. Walsh and S. P. Grossman. Some behavioral
effects of entorhinal cortex lesions in the albino rat. J. comp. physiol. Psychol. 85: 70-81, 1973.
25. Siegel, S. Nonparametric statistics for the behavioral sciences. New York: McGraw-Hill, 1956.
26. Smith, R. L., O. Steward, C. Cotman and G. Lynch. Axon sprouting in the hippocampal formation and behavioral recovery following unilateral entorhinal cortex lesions. (Abs.) Third Ann. Meet. Soci. Neurosci. San Diego, CA, 1973.
27. Steward, O., C. W. Cotman and G. S. Lynch. Growth of a new fiber projection in the brain of adult rats: re-innervation of the dentate gyrus by the contralateral entorhinal cortex following ipsilateral entorhinal lesions. Expl Brain Res. 20: 45--66, 1974.
28. Steward, O., C. Cotman and G. Lynch. A quantitative au- toradiographic and electrophysiological study of the reinnerva- tion of the dentate gyrus by the contralateral entorhinal cortex following ipsilateral entorhinal lesions. Brain Res. 114:181-200, 1976.
29. Steward, O. J. Leosche and W. C. Horton. Behavioral corre- lates of denervation and reinnervation of the hippocampal for- mation of the rat: Open field activity and cue utilization follow- ing bilateral entorhinal cortex lesions. Brain Res. Bull. 2:41-48, 1977.
30. Zimmer, J. and A. Hjorth-Simonsen. Crossed pathways from the entorhinal area to the fascia dentata II. Provokable in rats. J. comp. Neurol. 161: 71-102, 1975.