effects of acid rain on growth and nutrient concentrations in scots pine and norway spruce seedlings...
TRANSCRIPT

ELSEVIER 0 2 6 9 - 7 4 9 1 ( 9 4 ) 0 0 0 5 4 - 9
Environmental Pollution, Vol. 89, No. 2, pp. 177-187, 1995 Copyright © 1995 Elsevier Science Limited
Printed in Great Britain. All rights reserved 0269-7491/95 $09.50+0.00
EFFECTS OF ACID RAIN ON GROWTH A N D NUTRIENT CONCENTRATIONS IN SCOTS PINE A N D N O R W A Y SPRUCE
SEEDLINGS GROWN IN A NUTRIENT-RICH SOIL
Jaana Back, a Satu Hut tunen , a Minna Tu runen b & Jukka L a m p p u ~
~Department of Botany, University of Oulu, FIN-90570 Oulu, Finland bArctic Centre, University of Lapland, PO Box 122, FIN-96101 Rovaniemi, Finland
(Received 1 March 1994; accepted 23 June 1994)
Abstract The effects of artificially applied acid precipitation on growth and nutrient concentrations of Scots pine (Pinus sylvestris L.) and Norway spruce (Picea abies [L.] Karst.) seedlings were investigated in a long-term acid irrigation experiment in field conditions. Seedlings of northern and southern origin were planted in boxes con- taining peat and composted soil rich in nutrients, and sprinkler irrigated with water acidified with nitric and sulphuric acids to pH 3 or pH 4 for periods varying from two to three and a half growing seasons during 1986-1989. Water irrigated (pH 5.4-7.6) and non-irri- gated groups of seedlings were also included in the exper- iment. At the end of the experiment needles, main and lateral shoots and roots were collected from the seedlings for the determination of height growth and biomass par- titioning, and for the analysis of S, N, Mg, P, K, Ca, Mn and Fe concentrations. The treatment effects com- pared to the irrigated control were studied using multi- variate analyses of variance and covariance.
In the pine seedlings the total dry matter production increased by 25-70% compared with the irrigated controls when the total wet deposition to the seedlings exceeded 67 kg S ha 1 and 36 kg N h~ 1 (e.g. after two growing sea- sons' exposure of the pH 3 treatment). The increase was mainly due to an increase in needle dry weight (54-72% greater at pH 3) and root weight (20-65% greater at pH 3), whereas the height growth or shoot weight growth were less affected. The northern provenance pine seedlings responded more clearly to the pH 3 irrigation than the southern ones. The treatments had no consistent effects on any of the growth variables studied in the spruce seedlings, however. The pines had higher root and foliage Ca concen- trations as a result of the acid irrigation, whereas in spruce, acid rain decreased the Ca concentration in needles and shoots. Root Mn and Fe concentrations were higher in both species as a result of the pH 3 treatment. A higher soil con- ductivity and Ca concentration resulted from the prolonged pH 3 treatment. The results strongly support the hypothe- sis that the long-term growth and nutrient allocation response of conifers to acid precipitation is dependent both on the tree species and on the nutritional status of the soil.
Keywords: Acid rain, Scots pine, Norway spruce, seedling growth, nutrient status.
INTRODUCTION
177
Tree growth is a result of multiple, interacting, physio- logical processes influenced by an inherited genetic con- stitution and the ambient environment. The growth of coniferous trees is closely connected to the amount of nitrogen and other nutrients available for growth (Agren, 1985; Kachi & Rorison, 1989). The enhanced forest growth in Scandinavia and throughout northern Europe has recently been attributed to the increases in deposition of nitrogenous compounds (Kauppi et al., 1992). Air pollutants that limit the carbon gain or nutrient availability may, however, suppress the growth rate and total biomass production, and thus affect the allocation pattern (McLaughlin & Shriner, 1980; Troiano et al., 1983).
An increase in precipitation acidity predisposes tree seedlings to a number of environmental stresses, which are reflected in seedling germination, growth and sur- vival (e.g. Percy, 1986; Jacobson et al., 1990a; Shep- pard et al., 1993a,b). Increases in growth and the needle nitrogen concentration of conifer seedlings have been reported after exposure to acidic deposition con- taining nitrate or ammonium ions (Billen et al., 1990; Jacobson et al., 1990b; Dean & Johnson, 1992; Shel- burne et al., 1993; Sheppard et al., 1993a), whereas decreases in growth parameters have been seen in response to sulphate-containing precipitation (Leith et al., 1989; Jacobson et al., 1990a, b; McLaughlin et al., 1993; Sheppard et al., 1993a). However, considerable differences have been reported in the response of indi- vidual conifer species to acid rain (Lee & Weber, 1979; Percy, 1986; Abouguendia & Baschak, 1987; BiUen et al., 1990). Such variability may result from genetic diff- erences in susceptibility, nutrient imbalances in the soil or foliage, or from the direct influence of acid precipi- tation on the foliage. Interactions between acid rain and the chemical and biological characteristics of the soil have been found to be connected to the growth response of conifer seedlings in several experiments (e.g. Ashenden & Bell, 1988; Reich et al., 1988; Payer et al., 1990; Shipley et al., 1992). A complex interaction between soil nutrients, acid precipitation and climatic stresses has also been suggested as one likely cause of the forest decline phenomenon expressed as needle
178 J. Bdck et al.
discolouration and loss of photosynthetically active foliage (e.g. Payer et al., 1990; Htittl & Fink, 1991).
Research on the long-term responses of plants to environmental changes can reveal the dynamics of adaptation of the ecosystem or its components, and lead to more realistic conclusions of the direction and magnitude of the observed phenomena. Until recently the growth responses of conifers exposed to acid rain for several successive growing seasons were seldom reported (Billen et al., 1990; Dean & Johnson, 1992: Shelburne et al., 1993: Temple et al., 1993; B~,ck et al., 1994). The aim of this study was to follow the growth, nutrient status and viability of Scots pine (Pinus o'lvestris L.) and Norway spruce (Picea abies [L.] Karst.) seedlings of northern and southern provenance, growing in a nutrient-rich artificial soil and exposed to an elevated nitrate and sulphate load applied in the form of acid rain during several growing seasons.
MATERIAL AND M E T H O D S
Plant material An acid rain experiment was established in a natural pine forest about 5 km north of the center of Oulu, (65°00'N, 25°30'E) (Huttunen et al., 1990). Scots pine (Pinus syh,estris L.) and Norway spruce (Picea abies [L.] Karst,) seedlings were planted in June 1986 in plas- tic boxes (volume: 11 or 22 litres), with a perforated bottom, containing a mixture of composted and steam- sterilized soil, unfertilized peat and sand (1:2-5:0-5) . The seedlings, representing 29 provenances, were ob- tained from nurseries of the National Board of Forestry and the Finnish Forest Research Institute. At the time of planting the seedlings were one to two- years-old, and 10-20 cm in height. To avoid planting shock, the seedlings were planted in the boxes in June, and irrigation was begun in late August. The acid irri- gation experiment was begun in August 1986 and lasted until September 1989. Due to seedling mortality and harvesting during the experiment, additional seedlings from the same provenances were planted in spring 1987 and 1988. There were thus three different exposure duration groups (i.e. sulphate and nitrate doses), the exposure extending for 2, 3 or 3.5 growing seasons. The seedlings were divided into northern and southern provenance groups according to the geo- graphical recommendations for planting, the demarca- tion line running through Oulu, 65°N (provenances are listed in Huttunen et al. (1990)).
Table 1. The total (ambient + artificial) wet deposition of sulphur and nitrogen in kg ha -1
Duration pH 3 pH 4
S N S N
2 66.83 36.09 20.38 6-21 3 122-19 54.50 29.80 8.68 3-5 140-11 58.97 38-76 10.38
Duration = exposure duration (growing seasons). Numbers are means of four replicate blocks.
Table 2. Ambient air and precipitation quality, and climatic data during the course of the experiment in Ouln (Anonymous
1991; Finnish Meteorological Institute, 1986-1989)
1986 1987 1988 1989
SO 2 (/xg m -~)" 7 5 5 5 NO x (/,g m 3), 9 18 22 14 pH" 6.1 5.6 6.9 6.6
Precipitation (mm) b 237 350 255 295 Temperature sum (dd) ~ 1066 920 1300 1212 Frost sum (dd) c -1724 1217 -885
~Mean values from May-September. bSum between May and September. 'Sum of daily mean temperatures <O°C between October and April.
gated block received 5 mm of artificial precipitation for 5-10 min three times a week in addition to the ambient rainfall amounting to 60 mm month ~ during summer months in Oulu (long-term mean, Finnish Meteorologi- cal Institute 1986-1989). The acidic rain was allowed to pass through both the foliage and the soil. The irriga- tion water was prepared by adjusting the pH of tap water (1986-87), and later on, distilled water, to 3 or 4 by adding H2SO 4 and HNO3 (w:w, 2 :1) . The expo- sures were performed between 25 August to 13 October in 1986 (14 exposures), 11 May to 28 October in 1987 (64 exposures), 3 June to 17 October in 1988 (54 expo- sures) and 17 May to 23 August in 1989 (34 exposures). The total wet depositions of sulphur and nitrogen (am- bient+artificial) are given in Table 1, and the ambient air quality and weather conditions during the experi- ment in Table 2. The maximum snow depth in the experimental area was around 600 mm, and the duration of the snow cover was from mid-November to mid-April, approximately.
Irrigation treatments The experiment was arranged as a randomised block design with four treatments, each replicated in four blocks consisting of 80-120 seedlings planted in 20-30 boxes. The boxes were placed at a distance of ca 1.5 m from a whirling sprinkler (Uniflex Violetta), which dis- tributed the precipitation evenly over a radius of 1.2-2 m from the center of the circle of boxes. The treatments were: pH 3, pH 4, water irrigated control (IC; pH 5.4-7.6) and unwatered dry control (DC). Each irri-
Measurements Seedling height and viability were assessed prior to the start of the treatments, and only the visibly healthy seedlings were planted. The viability of the seedlings was followed by the same persons bi-annually, in spring before the treatment began and in autumn after the exposure had ended, and classified in five viability classes as follows: (i) healthy needles, good annual growth, normal bud set/burst: (ii) colour changes in <30% of needles, fairly good annual growth, normal

Effects of acid rain on Scots pine and Norway spruce seedlings 179
bud set/burst; (iii) colour changes in 30-50% of needles, annual growth slightly below average, delay in bud set/burst; (iv) colour changes in >50% of needles, poor annual growth, bud set/burst delayed; (v) dead seedlings. The dead seedlings were removed and replaced with seedlings from the same provenances to maintain the number of seedlings at a sufficient level.
At the end of the exposures in August 1989, 433 pine and 339 spruce seedlings were harvested for measure- ment of the length of the main root and above-ground growth, and as well for determination of the dry weight of shoots, remaining needles and roots. The roots were carefully rinsed with tap water. The above-ground parts of the seedlings were separated from the roots, and divided into current (1989), previous-year (1988) and older shoots (including the leader and lateral shoots). The lengths of the leader shoot and main root were measured before separation.
After division, the plant components were oven-dried at +60°C for 48 h. The needles were then removed from the shoots and both components weighed. The root/shoot and current year shoot/needle ratios were calculated from the dry weights. Nutrient concentra- tions (Mg, P, S, K, Ca, Mn, Fe) in the finely milled needles, shoots and roots were analysed on a sequential X-ray spectrometer (Siemens SRS 303AS, Germany), and the nitrogen concentration in the needles was analysed spectrophotometrically according to a modi- fied micro-Kjeldahl method. Minimum sample size for the element analyses was 0.05 g dry wt (N), 0.11 g dry wt (P, S, K, Ca, Mn and Fe) and 0-4 g dry wt (Mg), and therefore not all the elements could be analysed on all the samples. Owing to the small amounts of shoot material per seedling, and to the variable amounts of bark included in the smaller and larger shoots, the matrix of the milled samples used in the analysis of shoot sulphur concentrations varied, and a correction factor (0,54 × S, estimated from the arithmetic means of the pooled data) was used to correct the analysis results for the coarser-milled spruce shoot material (including less bark) with those of the finely milled powder (including larger proportion of bark). Because the results for the pine shoot matrix could not be corrected using any sin- gle factor, the element concentrations of the pine shoots are not included in the results.
The extractable concentrations of K, Ca, Mg and A1 in the growth substrate in the boxes were analysed on 15 ml fresh soil samples taken in August 1986 (before the experiment) and in August 1989 (after the 3.5 growing seasons' exposure period) by atomic absorp- tion spectrophotometry after extraction with 100 ml of 1 M ammonium acetate (pH 4.7; K, Ca, Mg) or 1 M KC1 (A1). Conductivity and pH (H20) was determined on soil mixed with 100 ml of water. Soil NH4+-N was analysed only before the experiment and amounted to 13.9/zg g-i fresh wt.
Statistical analyses For statistical purposes the data were separated accord- ing to species, provenance group, treatment, needle or
shoot year class and exposure duration group (i.e. N and S dose). Each exposure duration group was treated separately, and the results are presented for seedlings that have spent two or three growing seasons in the treatments. Only very few seedlings were treated for the whole period (3.5 growing seasons), and therefore this group is not included in the statistical analyses. The number of classifying variables was too high to justify the use of block-specific means for treatments, and therefore the growth, biomass accumulation, element concentrations and viability of the seedlings were com- pared for statistically significant differences using the box-specific arithmetic means of four individual seedlings placed in the same box. The number of boxes per treatment, dose, species and provenance group var- ied from 3 to 12. To reveal differences in seedling growth and element concentrations, multivariate analy- ses of variance and covariance (using the initial plant height as a covariate) were performed using a SAS sta- tistical package (SAS, 1988). Paired differences of treat- ments were studied a priori with Dunnett's t-test (comparison with IC) at a significance level of 0.05. The changes in seedling viability classes between obser- vation dates were tested for statistical significance with McNemar's test of symmetry (BMDP). Before the sta- tistical analyses, ln(y+l) transformation was applied to decrease, when necessary, the heterogeneity of the vari- ances. Changes in soil chemical characteristics were analysed with one-factor or two-factor repeated analy- sis of variance (StatView512+, Macintosh; n = 30-54 15 ml soil cores per treatment). Pearson correlation coefficients describing the inter-relationships between the variables were also calculated.
RESULTS
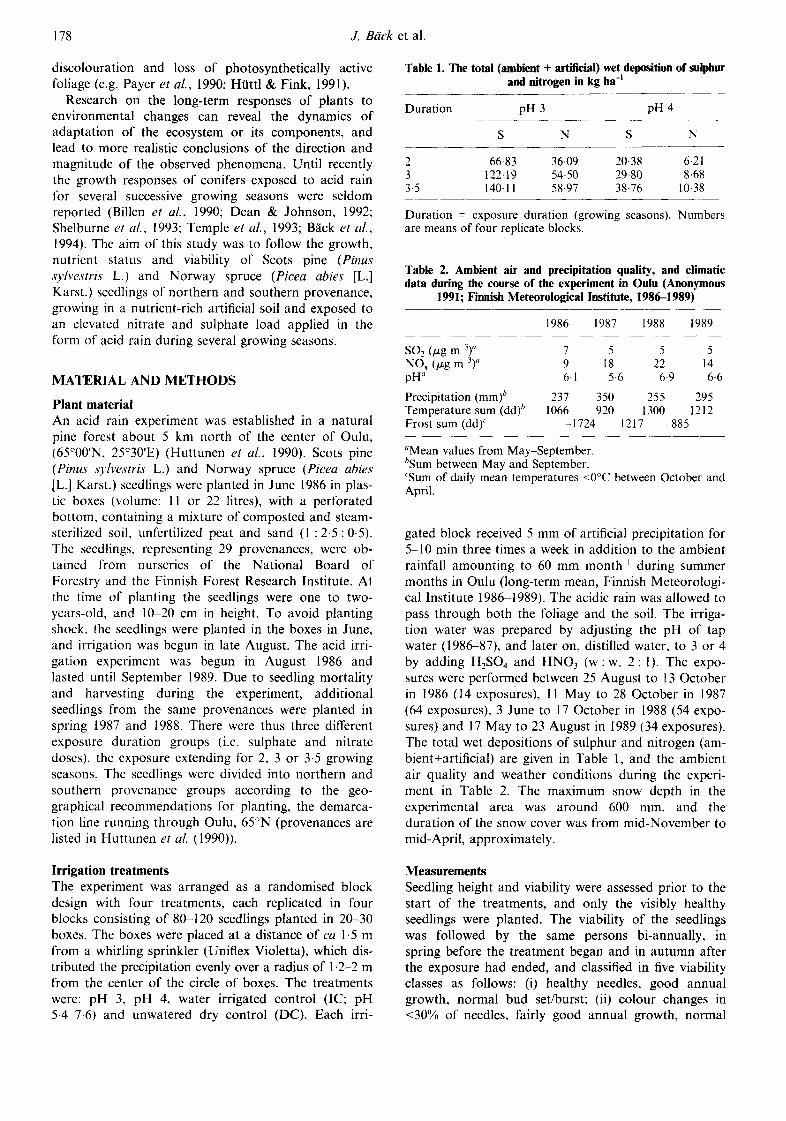
Growth and viability of the seedlings The species and the duration or level of the acid irriga- tion treatment significantly affected many of the seedling growth parameters. The needle dry matter pro- duction of the pine seedlings responded positively to the pH 3 treatment, but the annual height growth was less affected (Fig. 1, Table 3). After two growing sea- sons' irrigation at pH 3 (the wet deposition of S to- talling 67 kg ha -1, and the wet deposition of N totalling 36 kg ha 1; Table 1) the annual total shoot and needle dry weight of the pine seedlings was 25--45% greater than that in the non-irrigated control (DC) or irrigated control (IC) (Fig. 1). Dry weights of the current needles of southern provenance pine seedlings increased by 54% and 64% as a result of two growing seasons treat- ment at pH 3 compared to IC and DC, and those of northern provenance seedlings by 72% and 64%, respectively (Fig. 1). The dry weight of the previous year needles was also greatest in pH 3 seedlings of the northern pine provenance. The shoot/needle ratio of current year growth was smallest in the pH 3 treated pine seedlings in both provenance groups and in all the exposure duration groups due to the minor changes in shoot weight (data not shown).
180 J. Back et al.
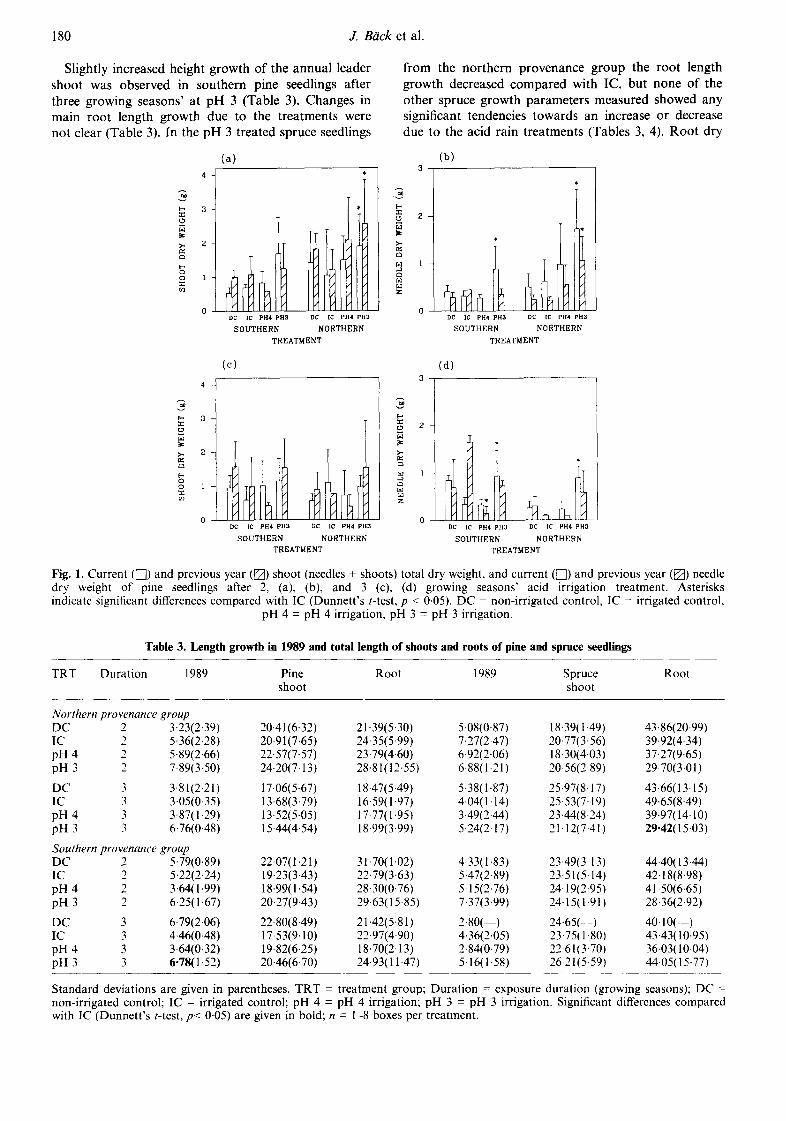
Slightly increased height growth of the annual leader shoot was observed in southern pine seedlings after three growing seasons' at pH 3 (Table 3). Changes in main root length growth due to the treatments were not clear (Table 3). In the pH 3 treated spruce seedlings
from the northern provenance group the root length growth decreased compared with IC, but none of the other spruce growth parameters measured showed any significant tendencies towards an increase or decrease due to the acid rain treatments (Tables 3, 4). Root dry
(a) (b) 3
4
"v
>.. 2 ~ '*
~ M
DC IC PH4 pH3 DC [C PH4 PH3 DC IC PH4 PH3 DC IC PH4 PH3
SOUTHERN NORTHERN SOUTHERN NORTHERN
T R E A T M E N T T R E A T M E N T
v
>. 2
¢71
o 1
(c) (d) 3
>-
DC [C PH4 PH3
S O U T H E R N
DC ]C PH4 PH3
NORTHERN
0 0 DC IC PH4 PH3 DC lC PH4 PH3
SOUTHERN NORTHERN T R E A T M E N T T R E A T M E N T
Fig. 1. Current (D) and previous year (V-3) shoot (needles + shoots) total dry weight, and current (D) and previous year (D) needle dry weight of pine seedlings after 2, (a), (b), and 3 (c), (d) growing seasons' acid irrigation treatment. Asterisks indicate significant differences compared with IC (Dunnett's t-test, p < 0.05). DC -- non-irrigated control, IC -- irrigated control,
pH 4 -- pH 4 irrigation, pH 3 = pH 3 irrigation.
Table 3. Length growth in 1989 and total length of shoots and roots of pine and spruce seedlings
TRT Duration 1989 Pine Root 1989 Spruce Root shoot shoot
Northern provenance group DC 2 3.23(2.39) 20.41(6.32) 21-39(5.30) 5.08(0.87) 1 8 . 3 9 ( 1 - 4 9 ) 43.86(20-99) IC 2 5.36(2.28) 20.91(7.65) 2 4 - 3 5 ( 5 - 9 9 ) 7.27(2.47) 20.77(3-56) 39.92(4.34) pH 4 2 5.89(2.66) 22.57(7.57) 23.79(4-60) 6.92(2.06) 1 8 - 3 0 ( 4 . 0 3 ) 37.27(9.65) pH 3 2 7.89(3.50) 24.20(7-13) 28.81(12.55) 6.88(1.21) 20.56(2.89) 29-70(3.01)
DC 3 3.81(2.21) 1 7 . 0 6 ( 5 . 6 7 ) 1 8 . 4 7 ( 5 . 4 9 ) 5.38(1.87) 25-97(8.17) 43.66(13.15) IC 3 3 - 0 5 ( 0 - 3 5 ) 1 3 . 6 8 ( 3 . 7 9 ) 1 6 . 5 9 ( 1 . 9 7 ) 4.04(1.14) 25.53(7-19) 49.65(8.49) pH 4 3 3 - 8 7 ( 1 - 2 9 ) 1 3 . 5 2 ( 5 . 0 5 ) 1 7 . 7 7 ( 1 . 9 5 ) 3 - 4 9 ( 2 . 4 4 ) 23.44(8.24) 39.97(14-I0) pH 3 3 6 76(0.48) 1 5 . 4 4 ( 4 . 5 4 ) 1 8 . 9 9 ( 3 . 9 9 ) 5.24(2-17) 21.12(7.41 ) 29"42(15.03)
Southern provenance group DC 2 5.79(0.89) 22.07(1.21) 31-70(1.02) 4.33(1.83) 2 3 - 4 9 ( 3 - 1 3 ) 44.40(13-44) IC 2 5.22(2.24) 19 -23 (3 -43 ) 22.79(3.63) 5.47(2.89) 23.51(5.14) 42.18(8.98) pH 4 2 3,64(1.99) 1 8 . 9 9 ( 1 . 5 4 ) 28.30(0.76) 5.15(2.76) 24.19(2.95) 41.50(6.65) pH 3 2 6.25(1-67) 20.27(9.43) 29-63(15.85) 7-37(3-99) 24-15( 1.91 ) 28.36(2-92)
DC 3 6.79(2.06) 22.80(8.49) 21.42(5.81) 2.80(--) 24-65(--) 40.10(--) IC 3 4-46(0.48) 1 7 - 5 3 ( 9 . 1 0 ) 22.97(4.90) 4.36(2.05) 23.75(1.80) 43-43(10.95) pH 4 3 3.64(0.32) 1 9 . 8 2 ( 6 . 2 5 ) 1 8 . 7 0 ( 2 - 1 3 ) 2.84(0-79) 22.61(3.70) 36.03(10.04) pH 3 3 6 " 7 8 ( 1 . 5 2 ) 20.46(6.70) 24-93(11.47) 5-16(1.58) 26.21(5.59) 44.05(15-77)
Standard deviations are given in parentheses. TRT = treatment group; Duration = exposure duration (growing seasons): DC = non-irrigated control; IC = irrigated control; pH 4 = pH 4 irrigation; pH 3 = pH 3 irrigation. Significant differences compared with IC (Dunnett's t-test, p< 0.05) are given in bold; n = 1-8 boxes per treatment.
Effects of acid rain on Scots pine and Norway spruce seedlings
Table 4. Total dry weight of roots and ratios of root to shoot of the pine and spruce seedlings
181
TRT Duration Pine Spruce
Root Root: shoot Root Root: shoot dry wt (g) dry wt (g)
Northern provenance group DC 2 0.722(0.34) 0.405(0.13) 1-651(0.61) 0.765(0.44) I f 2 0.785(0.40) 0.901(1.28) 1.616(0-38) 0.593(0.08) pH 4 2 0.945(0.61) 0.476(0.17) 1.722(0.45) 0.914(0.66) pH 3 2 2"026(1.75) 0.501(0.19) 2.126(0.73) 0.683(0.08) DC 3 0.470(0.23) 0.570(0.13) 2.461 (1.33) 0.526(0.13) I f 3 0.392(0.17) 0.488(0.05) 3.145(1.23) 0.998(0.94) pH 4 3 0.434(0.17) 0.636(0.19) 2.391(1.59) 0.661(0-20) pH 3 3 0.593(0.30) 0.549(0.13) 1.887(1.28) 0.950(0.65)
Southern provenance group DC 2 0.701 (0.07) 0.500(0.04) 2-831 (0.83) 0.578(0.71 ) IC 2 0.401 (0.15) 0.430(0-16) 2.298(0.99) 0.880(0.52) pH 4 2 0.439(0.06) 0.506(0-10) 3.824(0.22) 0.751(0.01) pH 3 2 1-072(0.70) 0.443(0.14) 2.553(1.17) 0.705(0.13) DC 3 0.810(0.47) 0.578(0.45) 2.573(--) 0.527(--) IC 3 0.769(0.54) 0.849(0.65) 1.414(0.44) 0.485(0.12) pH 4 3 0.713(0.41) 0.571 (0.15) 1.643(0.69) 0.577(0.09) pH 3 3 1.002(0.59) 0.573(0.14) 3.598(3.11) 0.602(0.05)
Standard deviations are given in parentheses. Symbols as in Table 3. Significant differences compared with IC (Dunnett's t-test, p < 0.05) are given in bold; n -- 1-8 boxes per treatment.
weight in the pine seedlings of both provenance groups was greater after two growing seasons at pH 3 than in the other treatment groups (Table 4). The root/shoot ratio varied in the pine seedlings between 0-41 and 1.39 and in the spruce seedlings between 0.49 and 1.00, but no effects of acid irrigation were observed.
No needle necrosis or chlorosis clearly attributable to acid irrigation was observed during the course of the experiment (data not shown). Seedling viability, in gen- eral, decreased during the treatment, the greatest decrease occurring in the seedlings treated for the longest period. Each year the viability of the seedlings improved during
8 re
,-e
1.5
1.0
0.5
O.0
(a)
m It@
L~
8
L~
DC IC PH 4 PH3 DC IC PH4 PH3
SOUTHERN NORTHERN TREATMENT
(b)
20
15
10
5
0 DC ]C PH4 PH3 DC IC PH4 PH3
SOUTHERN NORTHERN
TREATMENT
o u
re
5
(c)
1.5
1.0
0 . 5
0.0 ili IIii DC IC PH4 PH3 DC [C PH4 PH3
SOUTHERN NORTHERN TREATMENT
(d)
2 O
L 1ii 8 lo
~ 5
0 DC IC PH4 PH3 DC [C PH4 PH3
SOUTHERN NORTHERN TREATMENT
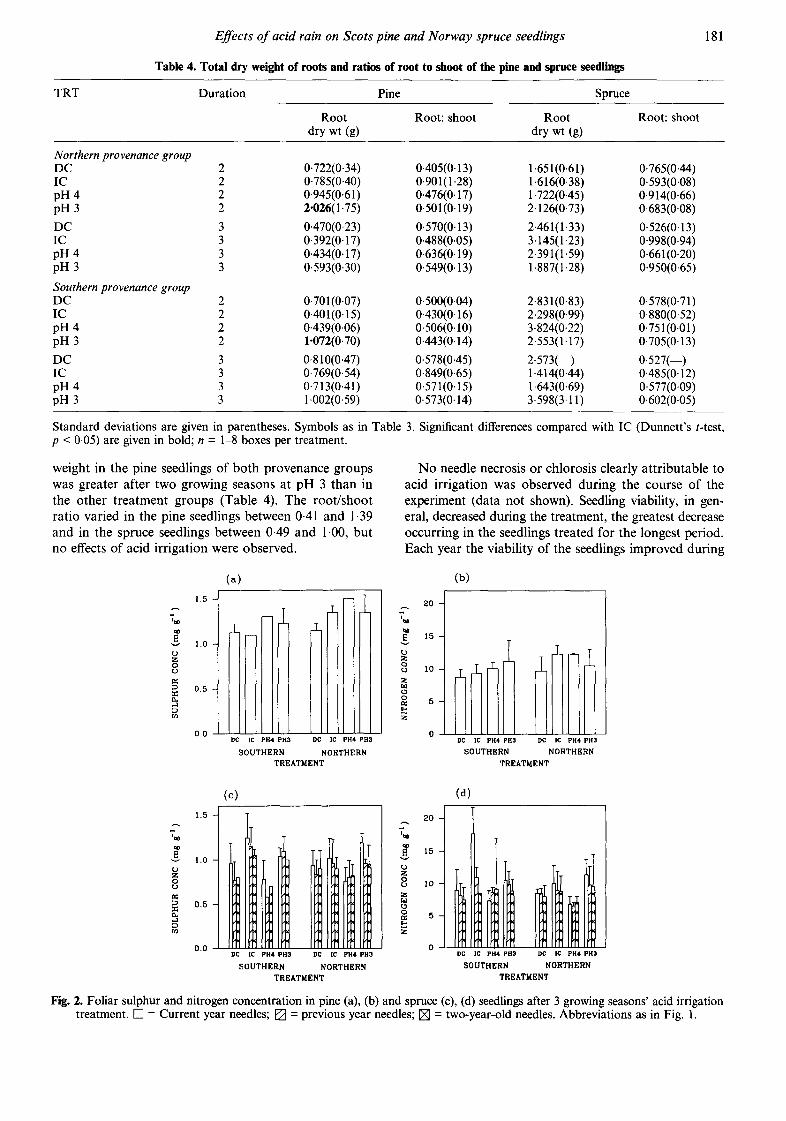
Fig. 2. Foliar sulphur and nitrogen concentration in pine (a), (b) and spruce (c), (d) seedlings after 3 growing seasons' acid irrigation treatment. [] -- Current year needles; [ ] = previous year needles; [ ] -- two-year-old needles. Abbreviations as in Fig. 1.

182 J. Brick et al.
~ . ~
..=N_~
|
II
6 6 6 6
6 6 6 6 6
d d d d ~
_ . ~
~.=- ~
K~ , - ~ . . ~ ' ~ ' ~
o ~
~ v
~ - =
~ .~.N
0
~
~ ~
6 o o 6
o o 6 0
.o
~: u = =
oo
[
: 0
I C'l
Z
I ,,,C
Z
oc
©
©
Effects of acid rain on Scots pine and Norway spruce seedlings 183
summer, and decreased during winter, but clear differ- ences were not found between treatment or provenance groups. The occurrence of winter injury was lowest in those provenances originating from the district of Oulu. Extensive fungal infections of spruce seedlings by Chrysomyxa ledi occurred during 1988-1989. The infec- tion pattern was similar in all provenance groups. The pine seedlings were more seriously infected by Ascoca- lyx abietina, Melampsora pinitorqua and Lophodermium sp., leading to the death of 50-100 seedlings per year.
Nutrient concentrations of needles, shoots and roots Marked differences in the nutrient concentrations between acid rain treatments occurred only in the current year needles after three growing seasons' treat- ment. The total foliar nitrogen concentration of current needles was 13% lower in the southern than in the northern pine provenance group, and 6% lower in the southern than in the northern spruce provenance group. When the data were analysed by treatment, the current year spruce foliage treated with clean water had the greatest needle nitrogen concentration (Fig. 2). Sul- phur accumulation in the needles and roots of irrigated pine (Fn~dles 1"93; Frootfi'40** ) and spruce (Fne~dJes6"05**; Froot~9'71**) was greater after three growing season's treatment compared with DC (Fig. 2), but no difference between the acid treatments and IC was seen.
Differences between the provenance groups as regards other nutrient concentrations were negligible, and therefore the data for provenance groups are hereafter pooled (Table 5). Calcium concentrations in the needles and roots of pine seedlings were 20-25% greater at pH 3 than in IC, although the difference was statistically significant only in the case of the needles. Needle and shoot Ca concentrations were 13-18% lower in the spruce seedlings from acid irrigation groups than from the IC group. Phosphorus concentration in the acid irrigated spruce needles was significantly lower than in
the needles of IC seedlings, and a slight decrease was also seen in the P concentration in shoots of the spruce seedlings treated with acid rain. Current needle K con- centrations were lowest in the pH 3 seedlings of both species. The N/P ratio was lowest in the DC and pH 4 treatment in the case of all spruce needle year classes (data shown only for current needles) and in the cur- rent year pine needles. The lowest Mn concentrations were found in the roots, shoots and needles of pine and spruce seedlings from the IC treatment. Correlations between the element concentrations and seedling growth were generally weak (r < 0.05).
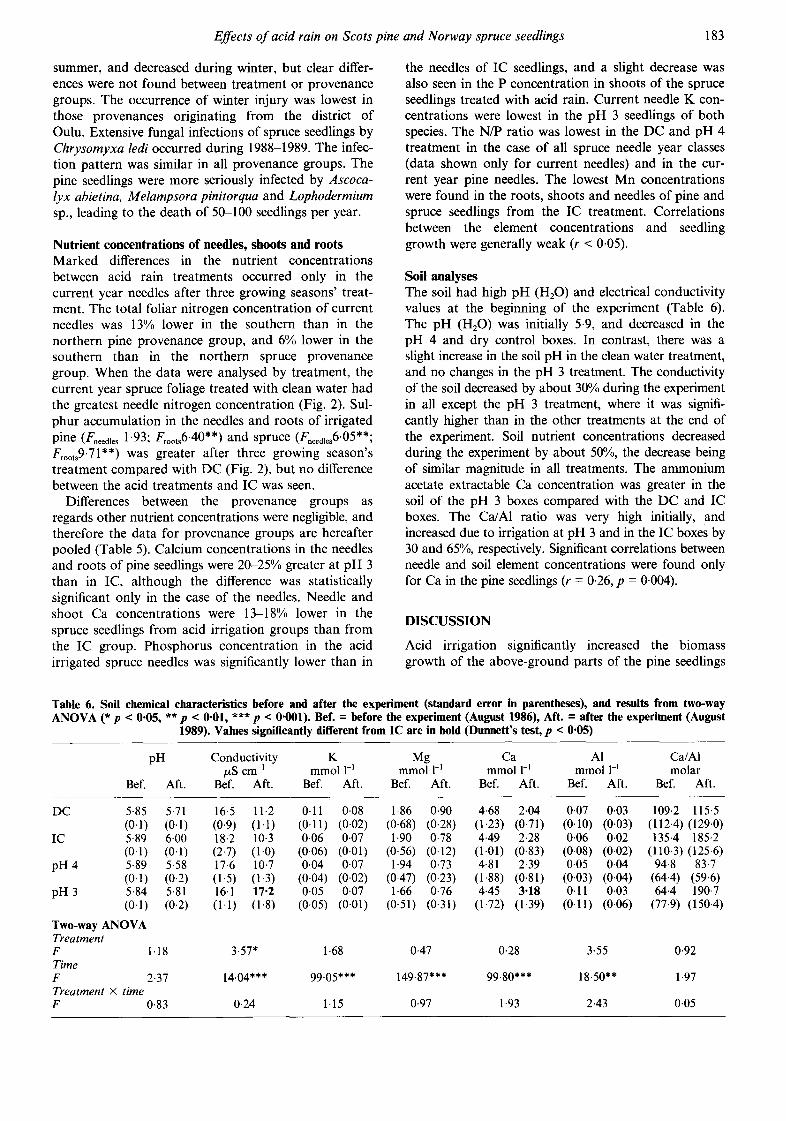
Soil analyses The soil had high pH (H20) and electrical conductivity values at the beginning of the experiment (Table 6). The pH (H20) was initially 5.9, and decreased in the pH 4 and dry control boxes. In contrast, there was a slight increase in the soil pH in the clean water treatment, and no changes in the pH 3 treatment. The conductivity of the soil decreased by about 30% during the experiment in all except the pH 3 treatment, where it was signifi- cantly higher than in the other treatments at the end of the experiment. Soil nutrient concentrations decreased during the experiment by about 50%, the decrease being of similar magnitude in all treatments. The ammonium acetate extractable Ca concentration was greater in the soil of the pH 3 boxes compared with the DC and IC boxes. The Ca/A1 ratio was very high initially, and increased due to irrigation at pH 3 and in the IC boxes by 30 and 65%, respectively. Significant correlations between needle and soil element concentrations were found only for Ca in the pine seedlings (r = 0-26, p = 0.004).
DISCUSSION
Acid irrigation significantly increased the biomass growth of the above-ground parts of the pine seedlings
Table 6. Soil chemical characteristics before and after the experiment (standard error in parentheses), and results from two-way ANOVA (* p < 0.05, ** p < 0"01, *** p < 0.001). Bef. = before the experiment (August 1986), Aft. = after the experiment (August
1989). Values significantly different from IC are in bold (Dunnett's test, p < 0"05)
pH Conductivity K p.S cm 1 mmol 1 i
Bef. Aft. Bef. Aft. Bef. Aft.
Mg Ca A1 Ca/A1 mmol 1-1 mmol 1 i mmol 1 1 molar
Bef. Aft. Bef. Aft. Bef. Aft. Bef. Aft.
DC 5-85 5.71 16.5 11.2 0.11 0.08 1.86 0.90 4.68 2.04 0.07 0.03 109.2 115.5 (0.1) (0.1) (0.9) (1 .1 ) (0.11) (0.02) (0.68) (0.28) (1.23) (0.71) (0.10) (0.03) (112.4)(129.0)
IC 5.89 6.00 18.2 10-3 0.06 0.07 1.90 0.78 4.49 2.28 0.06 0.02 135.4 185.2 (0.1) (0.1) (2.7) (1 .0 ) (0.06) (0.01) (0-56) (0.12) (1.01) (0.83) (0-08) (0.02) (110.3)(125-6)
pH 4 5.89 5.58 17.6 10.7 0.04 0.07 1.94 0.73 4.81 2.39 0-05 0.04 94-8 83.7 (0.1) (0.2) (1.5) (1 .3 ) (0.04) (0.02) (0.47) (0.23) (1-88) (0.81) (0-03) (0.04) (64.4) (59-6)
pH 3 5.84 5.81 16.1 17.2 0.05 0.07 1-66 0.76 4-45 3.18 0.11 0.03 64.4 190-7 (0.1) (0.2) (1-1) (1 .8 ) (0.05) (0.01) (0-51) (0-31) (1.72) (1.39) (0-11) (0.06) (77.9) (150-4)
Two-way ANOVA Treatment F I. 18 3.57* 1-68 0.47 0.28 3-55 0.92 Time F 2-37 14.04"** 99.05*** 149.87"** 99.80*** 18.50** 1.97 Treatment × time F 0.83 0.24 1.15 0.97 1.93 2.43 0.05

184 J. Bgick et al.
(a) 1.5
1 .0
0~ 0
o
5
(b)
0 . 5
0 . 0
1.5
1 .0
0 . 5
0 . 0 DC IC PH4 PH3 DC IC PH4 PH3 DC IC PH4 PH3 DC IC PH4 PH3
S O U T H E R N N O R T H E R N S O U T H E R N N O R T H E R N
TREATMENT TREATMENT
Fig. 3. Sulphur concentration in pine (a) and spruce (b) seedlings after 3 growing seasons' acid irrigation treatment. • -- Roots; [] = current year shoots. Asterisks indicate significant differences compared with IC (Dunnett's t-test, p < 0.05). Abbreviations as
in Fig. 1.
especially, the growth increase being clearest in the northern provenance pines. The response of Scots pine seedlings differed markedly from that of the Norway spruce seedlings. Under high acidic deposition levels, the Ca and Mn concentrations in the roots and needles of the pines increased together with increasing biomass and slightly increasing height growth, but in spruce there were only small changes. This is a novel finding, since the growth responses have in most cases been studied separately for each individual coniferous species (e.g. Jacobson et al., 1990a; Payer et al., 1990; Edwards et al., 1990, 1991; Dean & Johnson, 1992; McLaughlin & Tjoelker, 1992). The tendency of pines for such a growth increase may result from their adap- tation to nutrient poor soils, which amplifies the response of pine seedlings to increased nitrogen avail- ability. Enhanced nitrogen and sulphur inputs, coupled in this case with a fairly good nutrient supply from the soil, thus lead to greater growth stimulation in pines than in spruces.
The growth rate of conifer seedlings is affected for instance by seedling age, provenance, soil temperature and fertility, and the availability of moisture in the growth substrate (McLaughlin & Tjoelker, 1992; Shel- burne et al., 1993; Temple et al., 1993). Great variabil- ity has been found in the response of conifer seedling growth to acidic precipitation, the response primarily depending on the pH and composition of the artificial rain, soil nutrient availability and interacting air pollu- tant (mainly ozone) concentrations. Short-term expo- sures to acid rain or mist have resulted in neglible effects on the biomass or height growth of red spruce (Picea rubens Sarg.), Sitka spruce (Picea sitchensis [Bong.] Carribre), loblolly pine (Pinus taeda L.) and Ponderosa pine (Pinus ponderosa Dougl. ex Laws.) (Ashenden & Bell, 1988; Laurence et al., 1989; Kohut et al., 1990; Edwards et al., 1990, 1991; Jacobson et al., 1990a; Temple et al., 1993). In contrast, increased root and shoot biomass, or increased leaf area of red spruce, Norway spruce, Scots pine, Jeffrey pine (Pinus jeffreyi Murr.), slash pine (Pinus elliottii Engelm.), shortleaf pine (Pinus echinata Mill.) and silver fir (Abies alba Mill.) as a response to intermediate acidity containing
nitrate or ammonium ions are reported by Abrahamsen (1980), Taylor et al. (1986), Reich et al. (1987), Temple (1988), Billen et al. (1990), Jacobson et al. (1990a), Dean and Johnson (1992) and Shelburne et al. (1993). When the acidity is derived from sulphuric acid alone or together with nitric acid, decreases in growth are often seen (Jacobson et al., 1990a, b; Sheppard et al., 1993a; B~ick et al., 1994). Here the improved growth of Scots pine seedlings resulted from the exposure to rain containing both nitrate and sulphate, however.
Regulation of the seedling compartments seems to be a measure employed by the seedling to alleviate the effects of a restricted nutrient supply (Reich et al., 1987: Payer et al., 1990). Nitrogen is the dominant fac- tor determining the growth allocation processes, and the shoot-to-root ratio is highest in the case of suffi- cient or luxury nitrogen concentrations (Agren, 1985; Kachi & Rorison, 1989). In this study both the above- ground parts and the roots of pine seedlings increased along with increasing nitrogen deposition, maintaining a balance between biomass allocation to above-ground and below-ground organs.
The needle nitrogen concentrations were, in this experiment, slightly below the optimum for conifer seedlings (18 mg g ~ dry wt, according to Ingestad (1962)), especially when projected against the high concentra- tions of P, Ca and K (see e.g. Z6ttl et al. (1989), Raitio (1990)). The susceptibility of conifers to damage due to low temperatures increases with both enhanced and depressed levels of foliar nitrogen, the maximum hard- ening status being achieved at 13-18 mg N g~ dry wt (Aronsson, 1980; Sheppard et al., 1993a). Furthermore, because the ratio between N and other elements is also important for growth and survival (Simmons & Kelly, 1989; Sheppard et al., 1993a), it is possible that the observed nutrient imbalance promoted the winter dieback of the seedlings. The decreased viability may also indicate overall unsuitability of the rooting sub- strate for conifer seedlings, due to the rather high pH.
Calcium concentrations in the roots and needles in this study were comparable with the level of Ca in needles from adult spruce trees in the calcareous Alps (Fink, 1991), but D0 to 40-fold higher than in seedling studies

Effects o f acid rain on Scots pine and Norway spruce seedlings 185
carried out by Raitio (1990) or Tiirk et al. (1993), and were increased by the acid-irrigation treatment in Scots pine roots especially. Increasing acidity in precipitation enhances the transfer of cations from the root system to the above-ground plant compartments (Klemm, 1989). Calcium, Fe, Mn and Zn are not readily redis- tributed in the phloem, but rather tend to remain at the site where they are initially incorporated into the cellu- lar structures (Helmisaari, 1990). In addition, calcium is rather slowly transported from the roots to the above-ground organs, and the Ca concentrations in roots respond to Ca fertilization more rapidly than those in needles or stems (Ttirk et al., 1993). In accor- dance with our results, Ca in the needles of white pine seedlings and in the roots of loblolly pine seedlings also increased due to treatment with acid rain (Reich et al., 1988; Edwards et aL, 1991). In the spruce seedlings the IC treatment resulted in the highest Ca concentration in the needles and roots, possibly due to stimulated Ca uptake and decreased leaching by the clean water of neutral pH compared with the acid rain treatment (Abrahamsen et al., 1976). The water used for irriga- tion was initially tap water with a rather high Ca con- centration, but during the last two years the irrigation was performed with distilled water. The possible effect of the irrigation water on seedling element concentra- tions should have been visible in all irrigated seedlings compared to the non-irrigated ones, but only a decrease in the K concentration of the spruce shoots was observed due to irrigation.
The concentrations of Fe and Mn increased in both the roots and needles of the acid rain treated seedlings, possibly due to acid-induced increased solubility of the cations in the soil (Bergkvist, 1986). The solubility of A1, Fe and Mn in the soil is dependent on the soil pH, and in neutral soils A1 is precipitated as organic com- pounds which are not easily extractable (Duquette & Hendershot, 1987). Probably for this reason the soil in the boxes had a much higher Ca/A1 ratio than that in undisturbed forest soils. These soil and foliar element concentrations should therefore be compared with those from peat soils rather than mineral soils. The observed soil chemistry corresponds with that of similar pot studies, but field studies exhibit a large variation in the base cation concentrations (e.g. Z6ttl et al., 1989; Raitio, 1990). It is obvious that the buffer range of the soil in the boxes with high soil pH values had not yet been driven from base cation buffering into the A1 buffer range, which is usual in acidified soil types (Bergkvist & Folkeson, 1992). Plant uptake of cations was responsible for the observed 50% decrease in cation concentrations in the growth substrate, and if the exposure had been continued, signs of nutrient defi- ciencies and thereafter growth decreases might have been expected to occur. However, signs of increased Mn mobilisation or at least enhanced Mn uptake were to some extent already observable.
Correlation analysis of the element concentrations and seedling growth suggested that growth is influenced by many factors (potentially including temperature,
frost sums and precipitation) other than the nutrient concentrations in foliage or roots. In addition, weak dependences between growth and foliar and soil ele- ment concentrations are natural since the growth re- sponses are a result of a long-term development, whereas the element concentrations can change rather rapidly depending on the time during the growing sea- son (Helmisaari, 1990). The only slightly significant correlation was that with Ca, which increased in both the soil and the pine roots and foliage as a result of the pH 3 treatment in which growth was also the greatest. The lack of correlations can also be due to the fact that the seedlings should actually be classified as slightly N- deficient in relation to other nutrients, although the input of nitrate-N from the acid rain amounted to approximately 10 mg m 2 per irrigation episode at pH 3. It may be concluded also from this experiment, that an increased N supply can be utilized as growth if other nutrients like S, K, Ca or P are not limiting (Abraham- sen, 1980; Ashenden & Bell, 1988; Oren et al., 1988). Increasing nitrogen deposition would therefore be expected to lead to even greater growth increases, pro- viding that a decrease in the nutrient uptake rate due to leaching of cations from the soil or changes in the dynamics of soil organisms do not occur (Bergkvist, 1986; Oren et al., 1988).
The growth responses recorded in this study are in line with earlier observed changes in the cellular struc- tures of acid-rain treated needles of spruce and pine seedlings; increases in chloroplast growth and starch grain size were observed in the pH 3 treated needles (B~ick & Huttunen, 1992). The responses of pine and spruce seedlings to acid precipitation differ probably as a result of species-specific differences in nutrient uptake ability. The soil nutrient status greatly affects the growth responses, but cellular alterations seem to be rather similar, independent of the soil nutritional'status (B~ick & Huttunen, 1992; B~ick et al., 1994). It can be concluded on the basis of these results and those pre- sent in the literature that the magnitude and direction of the effects of acidic deposition on plant growth is strongly dependent on the plant species and on soil chemical and biological characteristics (Ashenden & Bell, 1988; Payer et al., 1990; KytOviita et al., 1990; B~ick et al., 1994).
ACKNOWLEDGEMENTS
The experiment was carried out as a part of the Finnish Acidification Project (HAPRO) financed by the Min- istry of Environment and Ministry of Agriculture and Forestry in Finland. The Academy of Finland and the Maj and Tor Nessling Foundation have also provided financial support for the work. The element analyses were carried out at the Institute of Electron Optics at the University of Oulu, and the assistance of Dr S. Sivonen and Mr O. Taikina-aho, Lic. Phil., especially is appreciated. The critical comments by Dr H. Raitio are greatly acknowledged. Revision of the language was performed by Mr J. Derome. The field work and

186 J. Back et al.
seedling measurements were assisted by several students and labora tory technicians f rom the Depar tmen t o f Botany and Botanical Gardens o f the Universi ty o f Oulu.
REFERENCES
Abouguendia, Z. M. & Baschak, L. A. (1987). Responses of two western Canadian conifers to simulated acid precipita- tion. Water, Air, Soil Pollut., 33, 15-22.
Abrahamsen, G. (1980). Impact of atmospheric sulphur deposition on forest ecosystems. In Atmospheric Sulfur De- position. Environmental Impact and Health Effects, ed. D. S. Shriner, C. R. Richmond & S. E. Lindberg. Ann Arbor Science Publishers, Michigan, USA, pp. 397-415.
Abrahamsen, G., Bjor, K., Horntvedt, R. & Tveite, B. (1976). Effects of acid precipitation on coniferous forest. In Impact o f Acid Precipitation on Forest and Freshwater Ecosystems in Norway. Research Report SNSF, Project FR6/76, ed. F. H Braekke. Norwegian Forest Research Institute, As, pp. 37-63.
Agren, G. I. (1985). Theory for growth of plants derived from the nitrogen productivity concept. Physiol. Plant., 64, 17-28.
Anonymous (1991). Oulun ilmanlaadun kehitys 1979-1990. Oulun kaupunki, ymp~irist6nsuojelutoimisto 6/1991 (in Finnish).
Aronsson, A. (1980). Frost hardiness in Scots pine (Pinus sil- vestris L.). II. Hardiness during winter and spring in young trees of different mineral nutrient status. Stud For. Suec., 155, 1 27.
Ashenden, T. W. & Bell, S. A. (1988). Growth responses of birch and sitka spruce exposed to acidified rain. Era,iron. Pollut., 51, 153-162.
B~ick, J. & Huttunen, S. (1992). Structural responses of nee- dles of conifer seedlings to acid rain treatment. New Phy- tol., 120, 77-88.
B~ick, J., Neuvonen, S. & Huttunen, S. (1994). Pine needle growth and fine structure after prolonged acid rain treat- ment in the subarctic. Plant. Cell Environ., 17, 1009-21.
Bergkvist, B. (1986). Leaching of metals from a spruce forest soil as influenced by experimental acidification. Water, Air, Soil Pollut., 31,901-1'6.
Bergkvist, B. & Folkeson, L. (1992). Soil acidification and el- ement fluxes of a Fagus sylvatica forest as influenced by simulated nitrogen deposition. Water, Air, Soil Pollut., 65, 111-33.
Billen, N., Sch~itzle, H., Seufert, G. & Arndt, U. (1990). Per- formance of some growth variables. Environ. Pollut., 68, 419-34.
Dean, T. J. & Johnson, J. D. (1992). Growth response of young slash pine trees to simulated acid rain and ozone stress. Can. J. For. Res., 22, 839-48.
Duquette, M. & Hendershot, W. H. (1987). Contribution of exchangeable aluminum to cation exchange capacity at low pH. Can. J. Soil Sci., 67, 175-85.
Edwards, N. T., Taylor, G. E. Jr, Adams, M. B., Simmons, G. L. & Kelly, J. M. (1990). Ozone, acidic rain and soil magnesium effects on growth and foliar pigments of Pinus taeda L. Tree Physiol., 6, 95-104.
Edwards, G. S., Edwards, N. T., Kelly, J. M. & Mays, P. A. (1991). Ozone, acidic precipitation, and soil Mg effects on growth and nutrition of loblolly pine seedlings. Environ. Exp. Bot., 31, 67-78.
Fink, S. (1991). The micromorphological distribution of bound calcium in needles of Norway spruce [Picea abies (L.) Karst.]. New Phytol., 119, 3340.
Finnish Meteorological Institute (1986-1989). Monthly Weather Reports. Helsinki, Finland.
Helmisaari, H.-S. (1990). Temporal variation in nutrient con-
centrations of Pinus sylvestris needles. Scand. J. For. Res., 5, 177-93.
Huttunen, S., Reinikainen, J. & Turunen, M. (1990). Winter- ing response of conifers to acid rain treatment under north- ern conditions. In Acidification in Finland, ed. P. Kauppi, P. Anttila & K. Kenttamies. Springer Verlag, Berlin, pp. 607-33.
Htittl, R. F. & Fink, S. (1991). Pollution, nutrition and plant function. In Plant Growth. Interactions with Nutrition and Environment, ed. J. R. Porter & D. W. Lawlor. Society for Experimental Biology, Seminar Series, 43, pp. 207-22.
Ingestad, T. (1962). Macro element nutrition of pine, spruce and birch seedlings in nutrient solutions. Medd. Stat. Skogsforskningsinst., 51, 1-131.
Jacobson, J. S., Heller, L. I., Yamada, K. E., Osmeloski, J. F., Bethard, T. & Lassoie, J. P. (1990a). Foliar injury and growth response of red spruce to sulfate and nitrate acidic mist. Can. J. For. Res., 20, 58-65.
Jacobson, J. S., Bethard, T., Heller, L. 1. & Lassoie, J. P. (1990b). Response of Picea rubens seedlings to intermittent mist varying in acidity, and in concentrations of sulfur, and nitrogen containing pollutants. Physiol. Plant., 78, 595-601.
Kachi, N. & Rorison, I. H. (1989). Optimal partitioning be- tween root and shoot in plants with contrasted growth pat- terns in response to nitrogen availability and temperature. Functional Ecol., 3, 549-59.
Kauppi, P. E., Mielik~iinen, K. & Kuusela, K. (1992). Biomass and carbon budget of European forests, 1971 to 1990. Science, 256, 1-74.
Klemm, O. (1989). Leaching and uptake of ions through above-ground Norway spruce tree parts. In Ecological Studies Vol, 77. Forest Decline and Air Pollution. A study of Spruce (Picea abies) on Acid Soils, ed. E.-D. Schulze, O. L. Lange & R. Oren. Springer Verlag, Berlin, pp. 210-37.
Kohut, R. J., Laurence, J. A., Amundson, R. G., Raba, R. M. & Melkonian, J. J. (1990). Effects of ozone and acidic precipitation on the growth and photosynthesis of red spruce after two years of exposure. Water. Air, Soil, Pollut., 51,277 86.
KytOviita, M.-M., Fritze, H. & Neuvonen, S. (1990). The effects of acid irrigation on soil microorganisms at Kevo, northern Finland. Environ. Pollut., 66, 21-31.
Laurence, J. A., Kohut, R. J. & Amundson, R. G. (1989). Response of red spruce seedlings exposed to ozone and simulated acidic precipitation in the field. Arch. Environ. Contam. Toxicol., 18, 285 90.
Lee, J. J. & Weber, D. E. (1979), The effect of simulated acid rain on seedling emergence and growth of eleven woody species. For. Sci., 25, 393 8.
Leinonen, L. & Juntto, S. (1992). llmanlaatumittauksia Air Quality Measurements 1991. Finnish Meteorological Insti- tute, Department of Air Quality, Helsinki, Finland.
Leith, I. D., Murray, M. B., Sheppard, L. J., Cape, J. N., Deans, J. D., Smith, R. I. & Fowler, D. (1989). Visible fo- liar injury of red spruce seedlings subjected to simulated acid mist. New Phytol., 113, 313-20.
McLaughlin, S. B. & Shriner, D. S. (1980).-Allocation of re- sources to defence and repair. In Plant Diseases, ed. J. B. Horsfall & E. B. Cowling., Vol 5. Academic Press, New York, pp. 407-31.
McLaughlin, S. B. & Tjoelker, M. J. (1992). Growth and physiological changes in red spruce saplings associated with acidic deposition at high elevations in the southern Ap- palachians, USA. For. Ecol. Manage., 51, 43 51.
McLaughlin, S. B., Tjoelker, M. G. & Roy, W. K. (1993). Acid deposition alters red spruce physiology: Laboratory studies support field observations. Can. J. For. Res., 23, 380-6.
Oren, R., Werk, K. S., Schulze, E.-D., Meyer, J., Schneider, B. U. & Schramel, P. (1988). Performance of two Picea

Effects o f acid rain on Scots pine and Norway spruce seedlings 187
abies (L.) Karst. stands at different stages of decline. VI. Nutrient concentration. Oceologia (Berl.), 77, 151-62.
Patton, R. L., Jensen, K. F. & Schier, G. A. (1991). Responses of red spruce seedlings to ozone and acid depo- sition. Can. J. For. Res., 21, 1354-9.
Payer, H. D., Pfirrmann, T., Kloos, M. & Blank, L. W. (1990). Clone and soil effects on the growth of young Nor- way spruce during 14 months exposure to ozone plus acid mist. Environ. Pollut., 64, 209-27.
Percy, K. (1986). The effects of simulated acid rain on germi- native capacity, growth and morphology of forest tree seedlings. New Phytol., 104, 473-84.
Raitio, H. (1990). Decline of young Scots pines in a dry heath forest. Acta Universitatis Ouluensis Ser. A, 216.
Reich, P. B., Schoettle, A. W., Stroo, H. F., Troiano, J. & Amundson, R. G. (1987). Effects of ozone and acid rain on white pine (Pinus strobus) seedlings grown in five soils. I. Net photosynthesis and growth. Can. J. Bot., 65, 977-87.
Reich, P. B., Schoettle, A. W., Stroo, H. F. & Amundson, R. G. (1988). Effects of ozone and acid rain on white pine (Pinus strobus) seedlings grown in five soils. III. Nutrient rela- tions. Can. J. Bot., 66, 1517-31.
SAS (1988). SAS Procedures Guide. Release 6.03 edition. SAS Institute, Cary, North Carolina, USA.
Shelburne, V. B., Reardon, J. C. & Paynter, V. A. (1993). The effects of acid rain and ozone on biomass and leaf area of shortleaf pine (Pinus echinata Mill.). Tree Physiol., 12, 163-72.
Sheppard, L. J., Cape, J. N. & Leith, I. D. (1993a). Influence of acidic mist on frost hardiness and nutrient concentra- tions in red spruce seedlings. 1. Exposure of the foliage and the rooting environment. New Phytol., 124, 595-605.
Sheppard, L. J., Cape, J. N. & Leith, I. D. (1993b). Influence of acidic mist on frost hardiness and nutrient concentra-
tions in red spruce seedlings. 2. Effects of misting frequency and rainfall exclusion. New Phytol., 124, 607-15.
Shipley, B., Lechowicz, M., Dumont, S. & Hendershot, W. H. (1992). Interacting effects of nutrients, pH-A1 and ele- vated CO 2 on the growth of red spruce (Picea rubens Sarg.) seedlings. Water, Air, Soil Pollut., 64, 585-600.
Simmons, G. L. & Kelly, J. M. (1989). Effects of acidic pre- cipitation, 03, and soil Mg status on throughfall, soil, and seedling loblolly pine nutrient concentrations. Water, Air, Soil Pollut., 43, 199-210.
Taylor, G. E. Jr, Norby, R. J., McLaughlin, S. B., Johnson, A. H. & Turner, R. S. (1986). Carbon dioxide assimilation and growth of red spruce (Picea rubens Sarg.) seedlings in response to ozone, precipitation acidity, and soil type. Oceologia (Berl.), 70, 163-71.
Temple, P. J. (1988). Injury and growth of jeffrey pine and giant sequoia in response to ozone and acidic mist. Envi- ron. Exp. Bot., 28, 323-33.
Temple, P. J., Reichers, G. H., Miller, P. R. & Lennox, R. W. (1993). Growth responses of ponderosa pine to long-term exposure to ozone, wet and dry acidic deposition, and drought. Can. J. For. Res., 23, 59~i6.
Troiano, J., Colavito, L., Heller, L., McCune, D. C. & Jacob- son, J. S. (1983). Effects of acidity of simulated rain and its joint action with ambient ozone on measures of biomass and yield in soybean. Environ. Exp. Bot., 23, 113-9.
Tiark, S., Giilpen, M. & Fink, S. (1993). Aufnahme, Trans- port und Verbleib yon Calcium und Magnesium in Fichten (Picea abies [L.] Karst.) und Kiefern (Pinus silvestris L.) bei unterschiedlicher Ern~ihrung und Schadstoffbelastung. Forstw. Cbl., 112, 191-208.
Zrttl, H. W., Hi~ttl, R. F., Fink, S., Tomlinson, G. H. & Wisniewski, J. (1989). Nutritional disturbances and histo- logical changes in declining forests. Water, Air, Soil Pollut.