effect of shoot size on the gas exchange and growth of containerized picea mariana seedlings under...
TRANSCRIPT

New Forests 13: 207–221, 1996.c 1996 Kluwer Academic Publishers. Printed in the Netherlands.
Effect of shoot size on the gas exchange and growth ofcontainerized Picea mariana seedlings under differentwatering regimes
M.S. LAMHAMEDI1, P.Y. BERNIER� and C. HEBERTNatural Resources Canada, Canadian Forest Service – Quebec, 1055 du PEPS, P.O. Box3800, Sainte-Foy, Quebec G1V 4C7, Canada; 1 Present address: Departement de foresterieet des ressources naturelles, Institut agronomique et veterinaire Hassan II, B.P. 6202,Rabat-Instituts, Rabat, Morocco (�Author for correspondence)
Received 17 February 1995; accepted 12 September 1995
Key words: black spruce, relative growth rate, shoot:root ratio, water relations, water stress
Application. The planting of large seedlings is currently viewed as one of the possible alterna-tives to herbicide applications in Eastern Canada. The results of our tests show that a four-foldincrease in shoot size (1.68 g to 6.82 g) in containerized black spruce stock does not signifi-cantly increase its susceptibility to water stress, even with an increase in the shoot:root ratio(2.2 g g�1 to 4.5 g g�1). With their much greater absolute growth, large seedlings offer abiologically feasible alternative to herbicides even on sites where competition might reducesoil water availability.
Abstract. Containerized black spruce (Picea mariana [Mill.] B.S.P.) seedlings of three dif-ferent sizes (small, medium, and large) were planted in raised sand beds maintained underwet, moderately dry or dry watering regimes during the growing season. The small seedlingswere of a conventional stock type. The two larger sizes were novel stock types grown inlarger containers. Physiological measurements during the summer showed that the small andmedium seedlings maintained nearly similar levels of gas exchanges and water status, but thatthe large seedlings had reduced net photosynthesis and stomatal conductance under all water-ing regimes. Analysis of dry masses showed comparable relative growth rates in the smalland medium seedlings, but a small to null growth in the large seedlings. Examination of rootrelative growth rate under wet conditions revealed significant root growth in small and mediumseedlings, but negligible growth in the large seedlings. It was concluded that increasing theshoot size of containerized seedlings can be achieved without increasing the susceptibility ofthe seedlings to water stress, as long as the vigour of the root system is maintained.
Mots cles: epinette noire, stress hydrique, relations hydriques, taux relatif de croissance,ratio tige:racines.
Resume. Nous avons plante dans des bacs de sable des plants d’epinette noire (Picea mariana[Mill.] B.S.P.) de trois tailles differentes (petits, moyens et grands) issus de production enconteneurs. Les bacs de sable ont ete maintenus a des niveaux differents de teneur en eau dusol (humide, modere et sec) durant la saison de croissance. Les plants “petits” etaient d’un typecouramment utilise pour le reboisement. Les plants “moyens et gros” etaient de types nou-veaux, produits dans des conteneurs plus volumineux. Les mesures physiologiques effectueestout au long de la saison de croissance ont revele des niveaux quasi-similaires d’echangesgazeux et de statut hydrique chez les plants petits et moyens, mais des niveaux inferieurs dephotosynthese nette et de conductivite stomatique chez les gros plants sous les trois regimes
VICTORY PIPS 111926 BIO1KAPnefo031.tex; 14/03/1997; 3:27; v.5; p.1

208
hydriques du sol. L’analyse des masses seches a montre des taux relatifs de croissancesimilaires chez les plants petits et moyens, mais des taux de faible a nul chez les gros plants.L’analyse des taux relatifs de croissance racinaire en regime hydrique humide a demontre unecroissance racinaire dynamique chez les plants petits et moyens, mais negligeable chez lesgros plants. Nous pouvons donc conclure qu’il est possible d’accroitre la cime de semis enconteneurs sans accroitre leur susceptibilite au stress hydrique apres reboisement, en autantque la qualite du systeme racinaire soit bonne.
Introduction
Mounting concern over the use of herbicides in forestry has led to a searchfor alternatives when plantations are to be established in areas of densecompetition. One possible alternative is the use of large planting stock that cancompete more efficiently for light and reduce long-term maintenance needs.Research on Douglas-fir (Pseudotsuga menziesii [Mirb.] Franco) (Newton etal. 1993), loblolly pine (Pinus taeda L.) (South et al. 1985) and Sitka spruce(Picea sitchensis [Bong.] Carr.) (South and Mason 1993) has shown that largeplanting stock can overtop herbaceous vegetation faster than small plantingstock.
Increasing the size of seedlings, and of containerized seedlings in par-ticular, poses new challenges. The production of novel stock types requiresmodifications to tried-and-tested cultural regimes with unknown effects onthe vigour of the seedlings. The successful establishment of planted seedlingsunder a range of environmental conditions also requires a certain equilib-rium between the evaporative surface and the water absorption capacity ofthe roots, an equilibrium that may or may not be present in larger seedlings.Tests with Douglas-fir, for example, have shown a lowering of predawn waterpotentials in greenhouse trials (Rose et al. 1993) and reduced growth on drysites with increased stock size (Hahn and Smith 1983). Similar reductionsin growth of larger seedlings have been observed in lodgepole pine (Pinuscontorta Dougl.) seedlings planted on dry sites (Baer et al. 1977). New largerstock types therefore need to be assessed to determine both their physiologicaland morphological quality.
In the province of Quebec (Canada), tests are currently under way forproducing, handling and planting containerized seedlings of much greatersize than what is currently the norm. One species targeted for this programis black spruce (Picea mariana [Mill.] B.S.P.), a species more susceptible towater stress than other boreal conifers of Eastern Canada (Grossnickle andBlake 1986; Blake and Sutton 1988; Lamhamedi and Bernier 1994). Waterstress has been shown to reduce initial growth of black spruce seedlings forat least two years after planting on moist boreal sites (Bernier 1993). Any
nefo031.tex; 14/03/1997; 3:27; v.5; p.2

209
link between seedling size and susceptibility to water stress should thereforebe apparent in such a sensitive species.
The objective of this study was to determine if larger containerized blackspruce seedlings were more susceptible to post-planting water stress thanthe conventional smaller containerized seedlings. The hypothesis was that anincrease in the transpiring surface area, either in absolute terms, or relative tothe root mass, will increase the post-planting susceptibility to water stress.
Materials and methods
Plant material and growth conditions
Containerized black spruce seedlings were obtained in three sizes from theprovincial nurseries at East-Angus and Saint-Modeste, Quebec (Table 1).Seeds came from the Quebec Ministry of Natural Resources’ bank of uncon-trolled provenances. The smallest stock type was from a normal productionrun. The other two types were test productions for the large seedling pro-gram. Differences in shoot size were achieved primarily through differencesin cavity size and spacing, and in fertilization rates. During the first year,all seedlings received 8–12 mg N. During the second year, small, mediumand large seedlings received about 30 mg, 160 mg and 300 mg of N percavity, respectively. All seedlings were grown over a two-year productionschedule. Small and medium seedlings were sown in their containers in earlyJune 1991 in unheated tunnels, transferred outdoors in October of that year,and raised in outdoor beds until May 1993. Large seedlings were sown in21 cm3 mini-cells in a polyethylene greenhouse in June 1991, hardened out-doors in the fall of that year, and stored in cold rooms in November 1991.In mid-January 1992, they were transplanted into large 700 cm3 containersand moved into a polyethylene greenhouse where they were kept under warmtemperatures and 16-hour days. In July 1992, they were moved outside wherethey remained until delivery in May 1993. Upon arrival at the laboratory, totalshoot volume was determined on all seedlings from the three stock types andonly seedlings with shoot volumes within one standard deviation from themean were retained for the experiment. Morphological characteristics weremeasured on subsamples of 20 seedlings from the populations of selectedseedlings (Table 1).
In mid-May 1993, seedlings were transplanted outdoors into three rec-tangular raised beds (4 � 1.20 � 0.5 m) filled with a sandy loam (83.8%sand, 15.8% loam, and 1.4% clay). Each bed was divided into four blocks.In each block, seven groups of three seedlings, or clusters, were planted,with one seedling of each type per cluster. The upper 15 cm of each sand
nefo031.tex; 14/03/1997; 3:27; v.5; p.3

210
Table 1. Initial characteristics of three seedling sizes of Picea mariana (mean � SD, n =20).
Small Medium Large
Container type Rigipot 45–110 Rigipot 45–340 VentBlock 15–700
Cavity volume (cm3) 110 340 700
Cavity depth (cm) 12 12 13
Root-collar diameter (mm) 2.43 � 0.05 4.57 � 0.12 9.49 � 0.27
Height (cm) 22.08 � 0.28 40.60 � 0.65 72.21 � 1.81
Dry masses (g)Needles 1.03 � 0.03 3.58 � 0.21 13.83 � 0.59Stem and branches 0.65 � 0.01 3.24 � 0.17 14.67 � 0.65Roots 0.79 � 0.03 1.54 � 0.11 20.76 � 1.38Total 2.65 � 0.06 8.36 � 0.16 49.26 � 2.02
Shoot:root ratio 2.17 � 0.09 4.54 � 0.09 1.48 � 0.10
bed was maintained at a different soil water content during the experiment.Target volumetric soil water contents (�v, cm3 cm�3) and correspondingsoil water potentials (SOIL, MPa) were wet (�v = 0.1, SOIL = –0.04),moderately dry (�v = 0.05,SOIL = –0.1) and dry (�v = 0.025,SOIL = –0.5)moisture regimes. These moisture regimes were selected on the basis of anearlier growth chamber experiment (Stewart and Bernier 1995). The sand bedassigned to the wet regime was watered daily in the evening with an automaticsprinkler. The other two beds were watered as needed to maintain the watercontent of the upper 15 cm close to target levels. Daily measurements of �v ofthe upper 15 cm were performed using a TDR unit (Model 1502b, TektronixInc., Richmond, OR, USA) (Topp et al. 1982; Hook et al. 1993). Removabletransparent polyethylene roofs were used to shelter the sand beds from therain while providing natural temperature and light conditions. Seedlings werefertilized weekly, with each bed receiving 34 g NPK (20–20–20 Plant ProductsLtd, Brampton, Ont., Canada).
Environmental parameters were monitored continuously over the growingseason. Soil temperatures were measured at –10 cm in each bed with copper-constantan thermocouples. Air temperature and relative humidity were mon-itored at seedling height with a Campbell Scientific 207 probe (CampbellScientific, Logan, UT, USA). Total incoming solar radiation was measuredwith a solarimeter (Kipp & Zonen, Delft, Holland). Average values for eachof these parameters during the periods of gas exchange measurements arepresented in Table 2.
nefo031.tex; 14/03/1997; 3:27; v.5; p.4
211
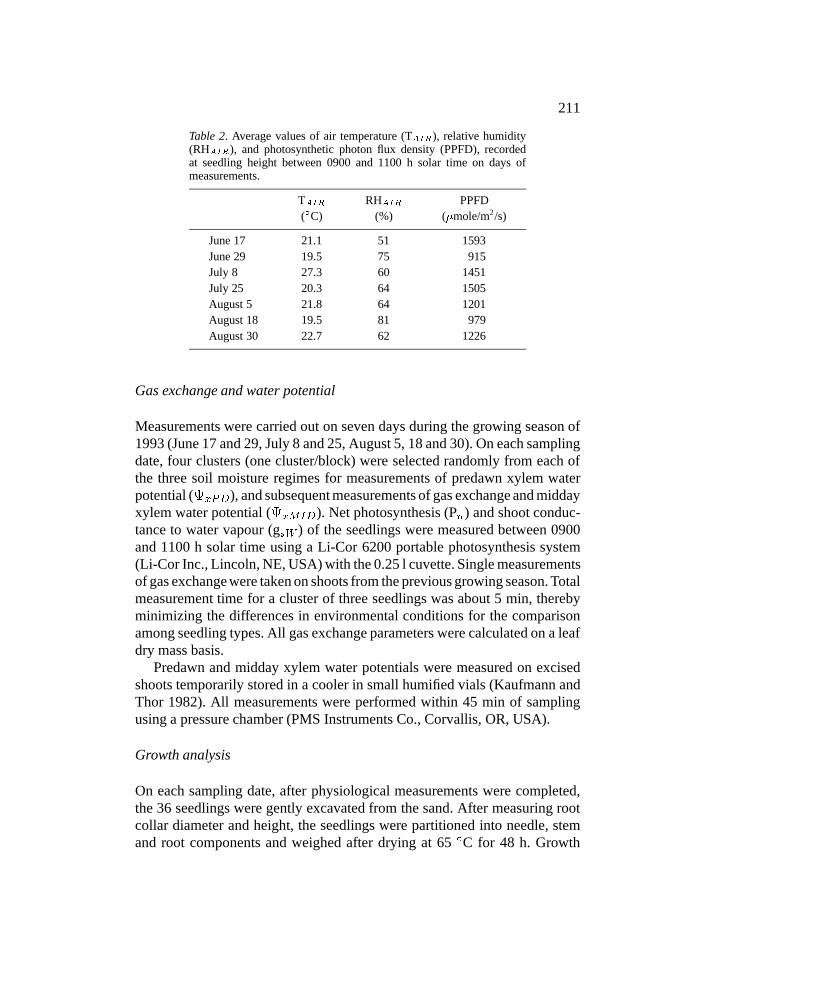
Table 2. Average values of air temperature (TAIR), relative humidity(RHAIR), and photosynthetic photon flux density (PPFD), recordedat seedling height between 0900 and 1100 h solar time on days ofmeasurements.
TAIR RHAIR PPFD(�C) (%) (�mole/m2/s)
June 17 21.1 51 1593June 29 19.5 75 915July 8 27.3 60 1451July 25 20.3 64 1505August 5 21.8 64 1201August 18 19.5 81 979August 30 22.7 62 1226
Gas exchange and water potential
Measurements were carried out on seven days during the growing season of1993 (June 17 and 29, July 8 and 25, August 5, 18 and 30). On each samplingdate, four clusters (one cluster/block) were selected randomly from each ofthe three soil moisture regimes for measurements of predawn xylem waterpotential (xPD), and subsequent measurements of gas exchange and middayxylem water potential (xMID). Net photosynthesis (Pn) and shoot conduc-tance to water vapour (gsW ) of the seedlings were measured between 0900and 1100 h solar time using a Li-Cor 6200 portable photosynthesis system(Li-Cor Inc., Lincoln, NE, USA) with the 0.25 l cuvette. Single measurementsof gas exchange were taken on shoots from the previous growing season. Totalmeasurement time for a cluster of three seedlings was about 5 min, therebyminimizing the differences in environmental conditions for the comparisonamong seedling types. All gas exchange parameters were calculated on a leafdry mass basis.
Predawn and midday xylem water potentials were measured on excisedshoots temporarily stored in a cooler in small humified vials (Kaufmann andThor 1982). All measurements were performed within 45 min of samplingusing a pressure chamber (PMS Instruments Co., Corvallis, OR, USA).
Growth analysis
On each sampling date, after physiological measurements were completed,the 36 seedlings were gently excavated from the sand. After measuring rootcollar diameter and height, the seedlings were partitioned into needle, stemand root components and weighed after drying at 65 �C for 48 h. Growth
nefo031.tex; 14/03/1997; 3:27; v.5; p.5

212
analysis was performed using an analysis of variance on the ln-transformeddry mass measurements of the seedlings collected on all seven sampling dates.In such an analysis, a significant interaction between date of measurementand treatment indicates a treatment-related difference in the rate of increaseof the log-transformed dry mass over time, and thus of relative growth rate(RGR) (Poorter and Lewis 1986; Poorter 1989). Time-courses of relativegrowth rates by stock type and watering regime are obtained first by fitting apolynomial of the form Ln(TDM) = a0 + a1D + � � � + anDn to the values oftotal seedling dry mass (TDM) and Julian date (D), and by differentiating theresulting function (Poorter and Lewis 1986; Poorter 1989).
Experimental design and statistical analysis
During the experiment, 252 seedlings (36 seedlings/date) were used forphysiological and growth measurements. The experiment was analyzed asa split-plot design within each watering regime with sampling date as themain plot and the three-seedling clusters as the subplots with four replicatesor blocks in each of the three sand beds. Comparisons of physiological andgrowth variables among the three sizes of seedlings were performed withthe mixed model procedure of the SAS 6.08 software using the restrictedmaximum likelihood (REML) approach (SAS Institute Inc. 1993). Measure-ments of xPD, of xMID and of dry masses were ln-transformed in orderto achieve normality and homogeneity of variances. Differences among thethree stock types regarding physiological and growth variables within eachwatering regime were determined by orthogonal a priori contrasts (Steel andTorrie 1980). Differences were considered significant at P < 0.05.
Results
Gas exchange and water relations
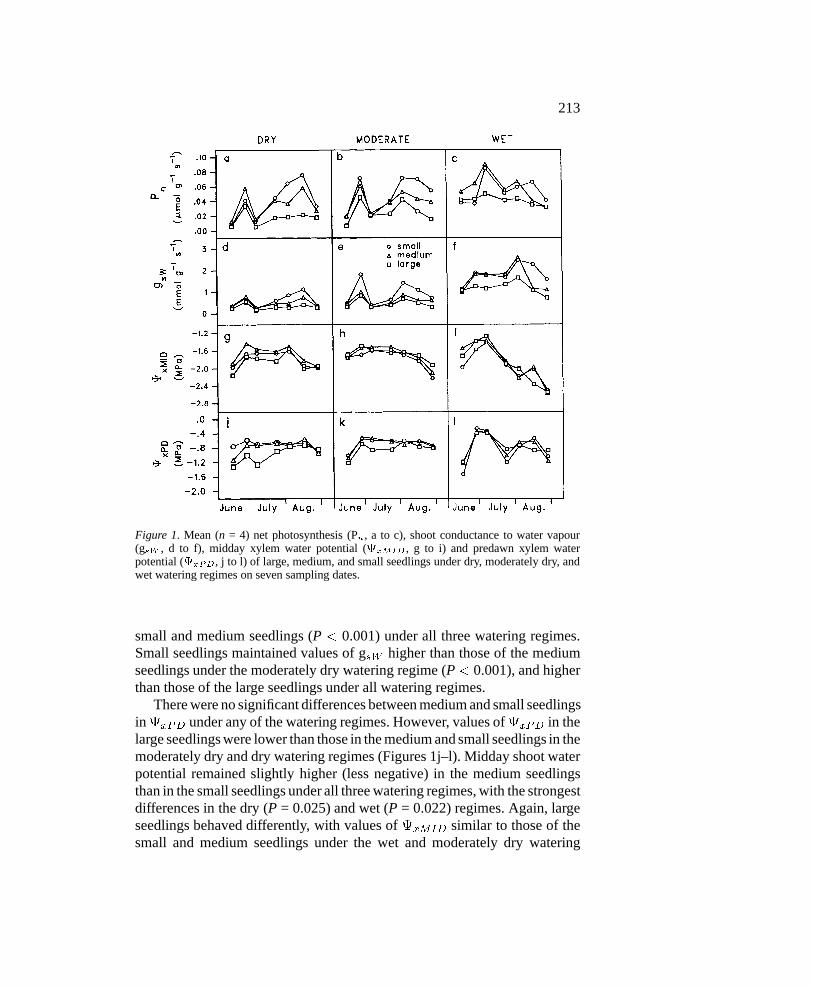
In general, reduced watering and increased seedling size both led to decreasesin gas exchange and shoot water potential (Figure 1, Table 3). However,significant two-way interactions between watering regime and seedling size(WR*S) for Pn,xMID andxPD, and between date and seedling size (D*S)for Pn and xPD, indicate that differences among seedling sizes varied withwatering regimes and over days.
Medium seedlings had values of Pn similar to those of the small seedlingsin both the wet and dry watering regimes, but weakly lower (P = 0.061) thanthose of the small seedlings in the moderately dry regime (Table 3, Figures1a–c). By contrast, large seedlings showed values of Pn lower than both
nefo031.tex; 14/03/1997; 3:27; v.5; p.6
213
Figure 1. Mean (n = 4) net photosynthesis (Pn, a to c), shoot conductance to water vapour(gsW , d to f), midday xylem water potential (xMID , g to i) and predawn xylem waterpotential (xPD, j to l) of large, medium, and small seedlings under dry, moderately dry, andwet watering regimes on seven sampling dates.
small and medium seedlings (P < 0.001) under all three watering regimes.Small seedlings maintained values of gsW higher than those of the mediumseedlings under the moderately dry watering regime (P < 0.001), and higherthan those of the large seedlings under all watering regimes.
There were no significant differences between medium and small seedlingsinxPD under any of the watering regimes. However, values ofxPD in thelarge seedlings were lower than those in the medium and small seedlings in themoderately dry and dry watering regimes (Figures 1j–l). Midday shoot waterpotential remained slightly higher (less negative) in the medium seedlingsthan in the small seedlings under all three watering regimes, with the strongestdifferences in the dry (P = 0.025) and wet (P = 0.022) regimes. Again, largeseedlings behaved differently, with values of xMID similar to those of thesmall and medium seedlings under the wet and moderately dry watering
nefo031.tex; 14/03/1997; 3:27; v.5; p.7
214
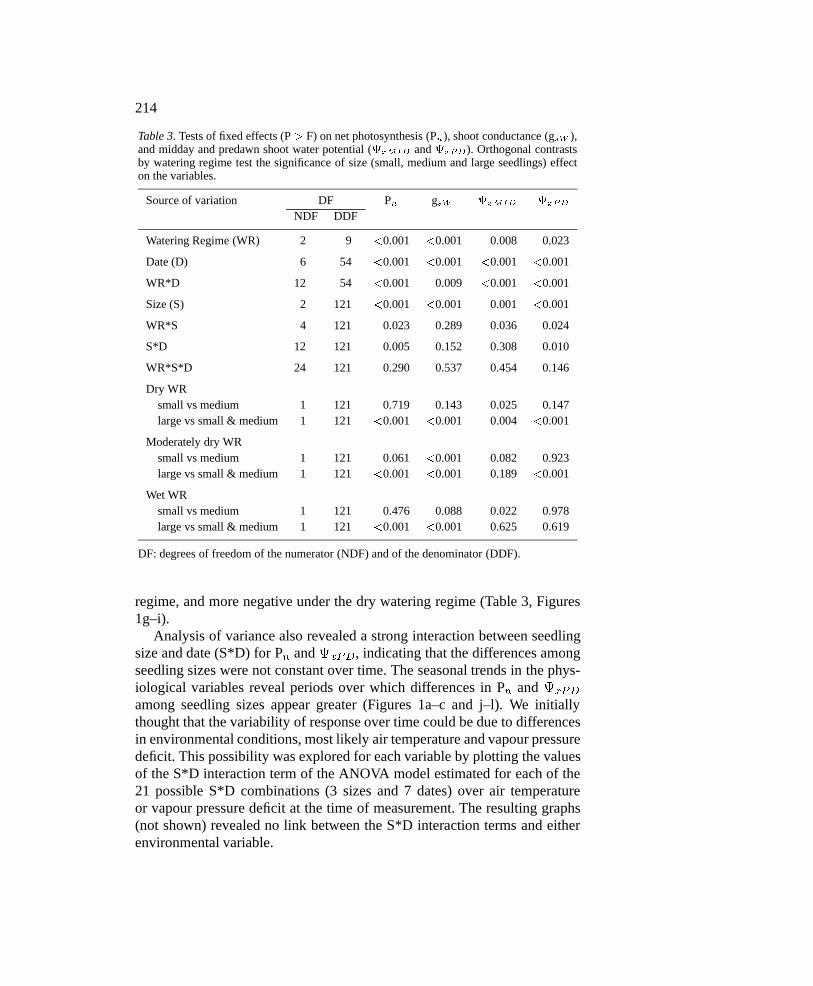
Table 3. Tests of fixed effects (P > F) on net photosynthesis (Pn), shoot conductance (gsW ),and midday and predawn shoot water potential (xMID and xPD). Orthogonal contrastsby watering regime test the significance of size (small, medium and large seedlings) effecton the variables.
Source of variation DF Pn gsW xMID xPD
NDF DDF
Watering Regime (WR) 2 9 <0.001 <0.001 0.008 0.023
Date (D) 6 54 <0.001 <0.001 <0.001 <0.001
WR*D 12 54 <0.001 0.009 <0.001 <0.001
Size (S) 2 121 <0.001 <0.001 0.001 <0.001
WR*S 4 121 0.023 0.289 0.036 0.024
S*D 12 121 0.005 0.152 0.308 0.010
WR*S*D 24 121 0.290 0.537 0.454 0.146
Dry WRsmall vs medium 1 121 0.719 0.143 0.025 0.147large vs small & medium 1 121 <0.001 <0.001 0.004 <0.001
Moderately dry WRsmall vs medium 1 121 0.061 <0.001 0.082 0.923large vs small & medium 1 121 <0.001 <0.001 0.189 <0.001
Wet WRsmall vs medium 1 121 0.476 0.088 0.022 0.978large vs small & medium 1 121 <0.001 <0.001 0.625 0.619
DF: degrees of freedom of the numerator (NDF) and of the denominator (DDF).
regime, and more negative under the dry watering regime (Table 3, Figures1g–i).
Analysis of variance also revealed a strong interaction between seedlingsize and date (S*D) for Pn and xPD, indicating that the differences amongseedling sizes were not constant over time. The seasonal trends in the phys-iological variables reveal periods over which differences in Pn and xPDamong seedling sizes appear greater (Figures 1a–c and j–l). We initiallythought that the variability of response over time could be due to differencesin environmental conditions, most likely air temperature and vapour pressuredeficit. This possibility was explored for each variable by plotting the valuesof the S*D interaction term of the ANOVA model estimated for each of the21 possible S*D combinations (3 sizes and 7 dates) over air temperatureor vapour pressure deficit at the time of measurement. The resulting graphs(not shown) revealed no link between the S*D interaction terms and eitherenvironmental variable.
nefo031.tex; 14/03/1997; 3:27; v.5; p.8

215
Growth analysis
Seedling size had a strong influence on the rates of dry mass (ln-transformed)accumulation, and of height and diameter growth, as shown by the significantS*D interactions for all measured morphological variables (Table 4). Becausethe ANOVA was performed on ln-transformed dry mass data, the significantS*D interactions for these variables can be interpreted as significant dif-ferences in RGR among seedling sizes. Watering regime also had a stronginfluence, as indicated by the significance of the S*WR interactions (Table4). A significant three-way WR*S*D interaction for many plant componentsalso reveals that the expression of the S*D interaction (differences in RGRamong seedling sizes) varied among watering regimes (Table 4). Contrastsperformed within each watering regime show a significant linear term of theS*D interaction (hence a difference in RGR) between the large seedlings andthe medium and small seedlings in nearly all growth variables (Table 4). Thesame contrasts reveal far fewer significant differences between the mediumand small seedlings, and only a weak trend (P = 0.095) for the total shoot drymass (ln-transformed) under the wet watering regime.
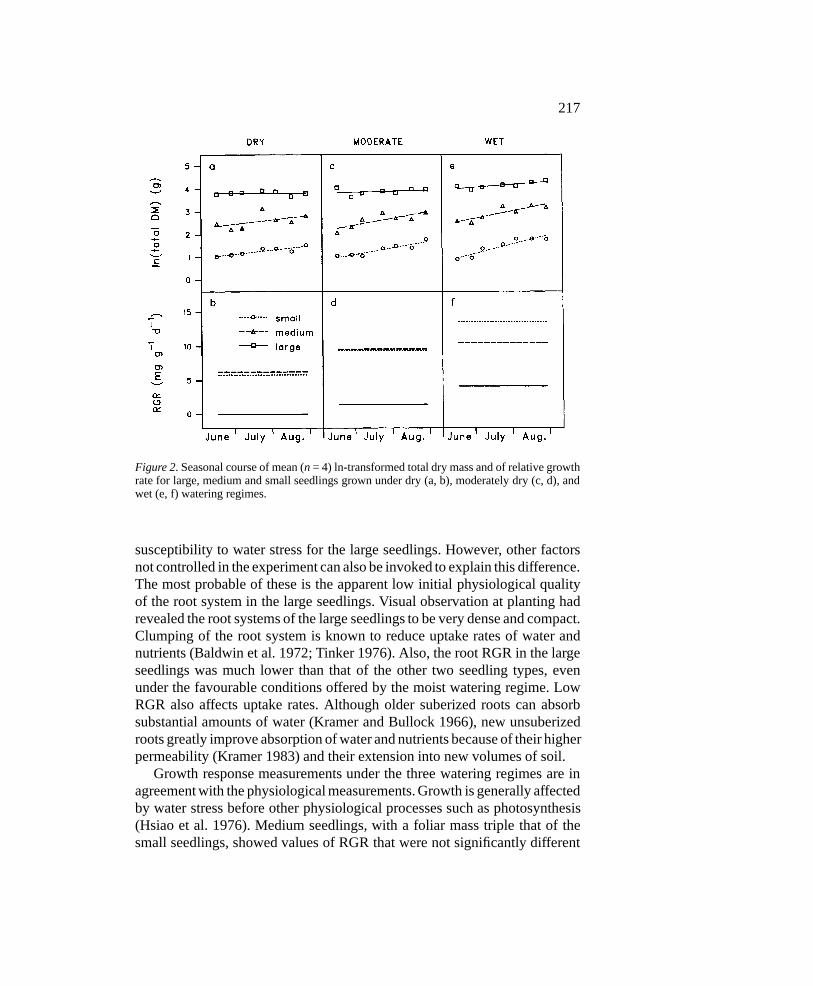
This last analysis is illustrated graphically in Figure 2, where linear func-tions have been fitted to the ln-transformed total shoot dry mass data plottedagainst Julian dates. First-order polynomials were used for the computationof RGR values since only the linear component was significant for the overallS*D interaction (analysis not shown). The first-order derivative of these func-tions, the RGR, clearly shows the pattern of decrease of RGR with decreasingsoil water availability, as well as the difference among seedling sizes withineach watering regime.
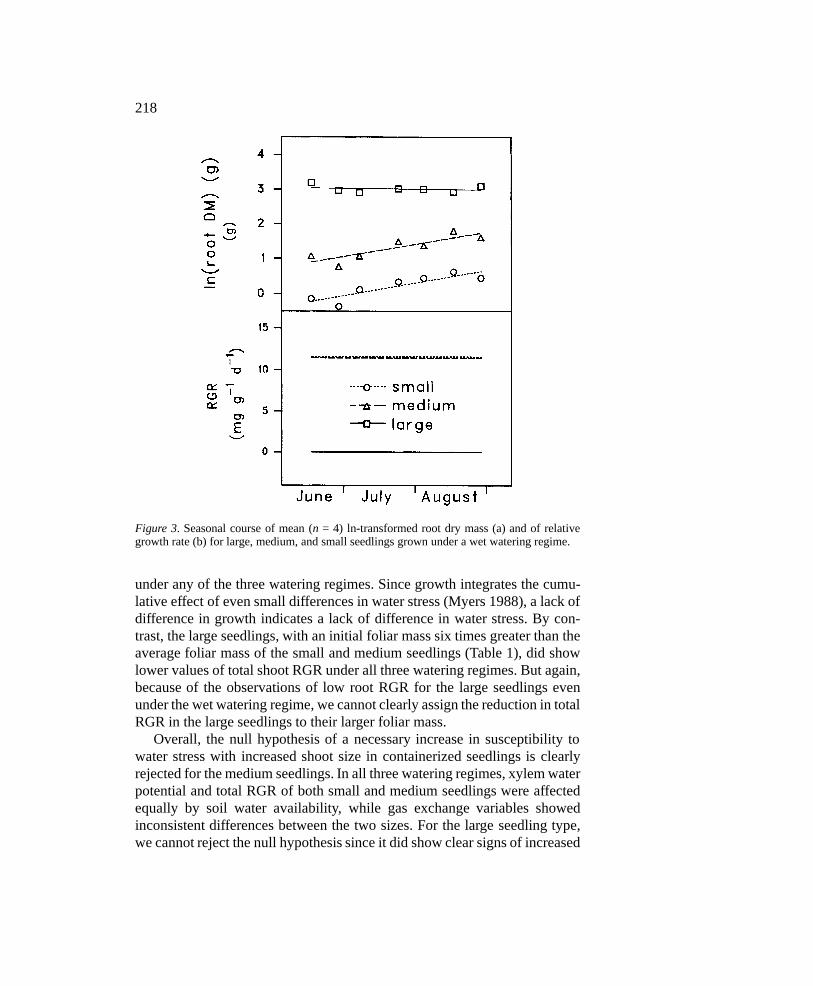
Growth analysis was also performed on the root dry mass of seedlingssubjected to the wet watering regime. The ANOVA (Table 4) had revealed asignificant S*D interaction for root mass, indicating significant differences inroot RGR among seedling sizes. Results of the RGR analysis on the seedlingsin the wet watering regime show identical RGRs for the medium and smallseedlings. By contrast, the root RGR of the large seedlings remained nearzero (Figure 3).
Discussion
The analysis of physiological measurements does not support a direct linkbetween increased foliar mass, either in absolute terms or relative to shootmass, and an increased susceptibility to water stress. A tripling of the foliarmass between the small and medium seedlings yielded no consistent differ-ences in either xPD or xMID within any of the three watering regimes.
nefo031.tex; 14/03/1997; 3:27; v.5; p.9
216
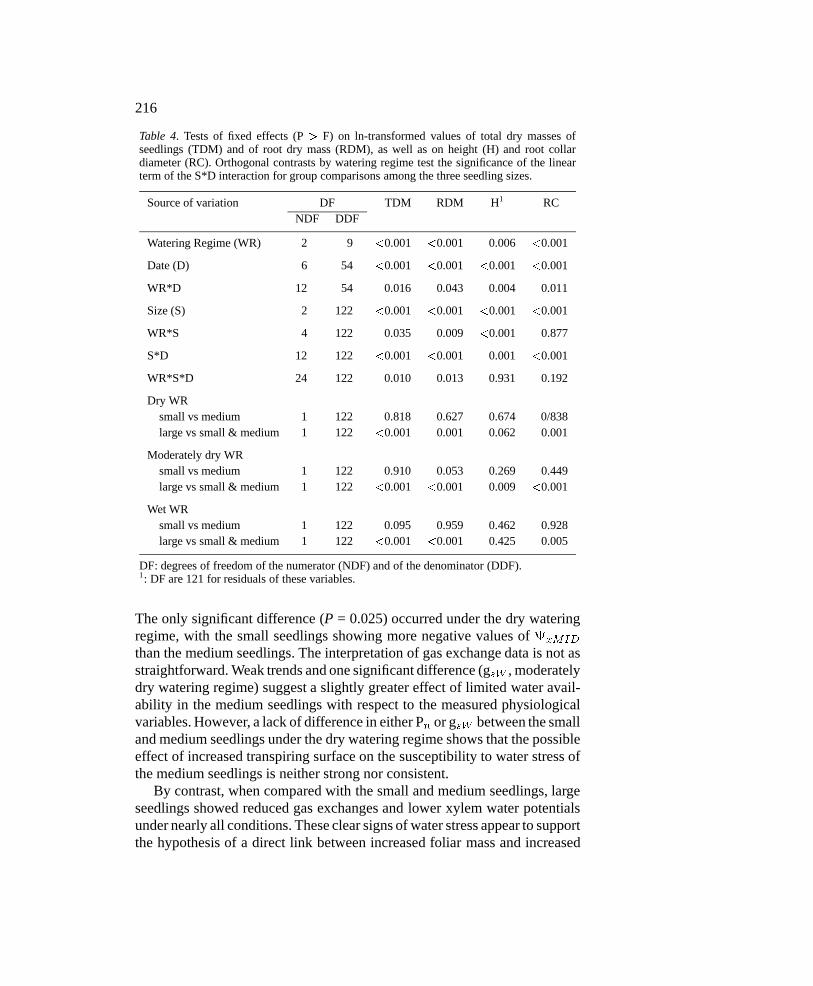
Table 4. Tests of fixed effects (P > F) on ln-transformed values of total dry masses ofseedlings (TDM) and of root dry mass (RDM), as well as on height (H) and root collardiameter (RC). Orthogonal contrasts by watering regime test the significance of the linearterm of the S*D interaction for group comparisons among the three seedling sizes.
Source of variation DF TDM RDM H1 RCNDF DDF
Watering Regime (WR) 2 9 <0.001 <0.001 0.006 <0.001
Date (D) 6 54 <0.001 <0.001 <0.001 <0.001
WR*D 12 54 0.016 0.043 0.004 0.011
Size (S) 2 122 <0.001 <0.001 <0.001 <0.001
WR*S 4 122 0.035 0.009 <0.001 0.877
S*D 12 122 <0.001 <0.001 0.001 <0.001
WR*S*D 24 122 0.010 0.013 0.931 0.192
Dry WRsmall vs medium 1 122 0.818 0.627 0.674 0/838large vs small & medium 1 122 <0.001 0.001 0.062 0.001
Moderately dry WRsmall vs medium 1 122 0.910 0.053 0.269 0.449large vs small & medium 1 122 <0.001 <0.001 0.009 <0.001
Wet WRsmall vs medium 1 122 0.095 0.959 0.462 0.928large vs small & medium 1 122 <0.001 <0.001 0.425 0.005
DF: degrees of freedom of the numerator (NDF) and of the denominator (DDF).1: DF are 121 for residuals of these variables.
The only significant difference (P = 0.025) occurred under the dry wateringregime, with the small seedlings showing more negative values of xMID
than the medium seedlings. The interpretation of gas exchange data is not asstraightforward. Weak trends and one significant difference (gsW , moderatelydry watering regime) suggest a slightly greater effect of limited water avail-ability in the medium seedlings with respect to the measured physiologicalvariables. However, a lack of difference in either Pn or gsW between the smalland medium seedlings under the dry watering regime shows that the possibleeffect of increased transpiring surface on the susceptibility to water stress ofthe medium seedlings is neither strong nor consistent.
By contrast, when compared with the small and medium seedlings, largeseedlings showed reduced gas exchanges and lower xylem water potentialsunder nearly all conditions. These clear signs of water stress appear to supportthe hypothesis of a direct link between increased foliar mass and increased
nefo031.tex; 14/03/1997; 3:27; v.5; p.10
217
Figure 2. Seasonal course of mean (n = 4) ln-transformed total dry mass and of relative growthrate for large, medium and small seedlings grown under dry (a, b), moderately dry (c, d), andwet (e, f) watering regimes.
susceptibility to water stress for the large seedlings. However, other factorsnot controlled in the experiment can also be invoked to explain this difference.The most probable of these is the apparent low initial physiological qualityof the root system in the large seedlings. Visual observation at planting hadrevealed the root systems of the large seedlings to be very dense and compact.Clumping of the root system is known to reduce uptake rates of water andnutrients (Baldwin et al. 1972; Tinker 1976). Also, the root RGR in the largeseedlings was much lower than that of the other two seedling types, evenunder the favourable conditions offered by the moist watering regime. LowRGR also affects uptake rates. Although older suberized roots can absorbsubstantial amounts of water (Kramer and Bullock 1966), new unsuberizedroots greatly improve absorption of water and nutrients because of their higherpermeability (Kramer 1983) and their extension into new volumes of soil.
Growth response measurements under the three watering regimes are inagreement with the physiological measurements. Growth is generally affectedby water stress before other physiological processes such as photosynthesis(Hsiao et al. 1976). Medium seedlings, with a foliar mass triple that of thesmall seedlings, showed values of RGR that were not significantly different
nefo031.tex; 14/03/1997; 3:27; v.5; p.11
218
Figure 3. Seasonal course of mean (n = 4) ln-transformed root dry mass (a) and of relativegrowth rate (b) for large, medium, and small seedlings grown under a wet watering regime.
under any of the three watering regimes. Since growth integrates the cumu-lative effect of even small differences in water stress (Myers 1988), a lack ofdifference in growth indicates a lack of difference in water stress. By con-trast, the large seedlings, with an initial foliar mass six times greater than theaverage foliar mass of the small and medium seedlings (Table 1), did showlower values of total shoot RGR under all three watering regimes. But again,because of the observations of low root RGR for the large seedlings evenunder the wet watering regime, we cannot clearly assign the reduction in totalRGR in the large seedlings to their larger foliar mass.
Overall, the null hypothesis of a necessary increase in susceptibility towater stress with increased shoot size in containerized seedlings is clearlyrejected for the medium seedlings. In all three watering regimes, xylem waterpotential and total RGR of both small and medium seedlings were affectedequally by soil water availability, while gas exchange variables showedinconsistent differences between the two sizes. For the large seedling type,we cannot reject the null hypothesis since it did show clear signs of increased
nefo031.tex; 14/03/1997; 3:27; v.5; p.12

219
water stress compared with the two smaller seedling types. However, evi-dence of low initial growth potential of the root system of the large seedlingssuggests that increased foliar mass may not be the actual cause of increasedwater stress in these seedlings.
The analysis of growth response to soil water availability reveals no linkbetween the shoot:root ratio and the post-planting susceptibility to waterstress. At planting time, the large and small seedlings had the most favourableshoot:root ratios (1.48 g g�1 and 2.17 g g�1 respectively) while the mediumseedlings had the least favourable one (4.54 g g�1, Table 2). However, theRGR of the medium seedlings was higher than that of the large seedlings, andequal to that of the small seedlings under all three watering regimes. In bare-root stock, investigators have reported better seedling field performance inseedlings with low initial shoot:root ratios (Haase and Rose 1993; Ritchie etal. 1993). In containerized stock, however, investigators have generally failedto find a consistent relationship between shoot:root ratio and post-plantationgrowth (Walker and Johnson 1980; Romero et al. 1986; Maass et al. 1989).The low density of the rooting medium and the ideal nursery conditionsencourage root growth inside containers beyond what might be necessary forgood field performance. Excessive root growth inside the root plug, as foundin the large seedlings used in this study, can even lead to low root qualityand therefore increased susceptibility to water stress in spite of an apparentfavourable shoot:root ratio.
In summary, this test indicates that increasing the shoot size of black sprucecontainerized seedlings to sizes at least as large as the medium seedlings testedin this study does not by itself significantly increase the susceptibility of theseedlings to water stress after planting. We think that this result should apply tomany other coniferous species, given the known susceptibility of black spruceto water stress. The results also suggest that shoot:root ratios in containerizedseedlings are not well related to their post-planting susceptibility to waterstress.
Acknowledgments
We would like to thank Benoit-M. Gingras and Corine Rioux (ministere desRessources naturelles du Quebec) for providing the plant material, MicheleBernier-Cardou for her advice in experimental design and statistical analysis,Pierre Davignon and Francois-Hugues Bernier for their technical assistance,and Pamela Cheers for her editorial comments. Thanks are also extended toDr. Steve Grossnickle for his critical review of the manuscript. Postdoctoralsupport for Dr. Lamhamedi was provided by the Natural Sciences and Engi-
nefo031.tex; 14/03/1997; 3:27; v.5; p.13

220
neering Research Council of Canada and by the Canadian Forest Service,Natural Resources Canada.
References
Baldwin, J. P., Tinker, P. B. and Nye, H. P. 1972. Uptake of solutes by multiple root systemsfrom soil. II. The theoretical effects of rooting density and pattern on uptake of nutrientsfrom soil. Plant Soil, 36: 693–708.
Baer, N., Ronco, F., Barney, C. W. and Baer, N. W. 1977. Effects of Watering, Shading and Sizeof Stock on Survival of Planted Lodgepole Pine. USDA For. Ser. Rocky Mount. Forestand Range Exp. Sta. Res. Note RM-347.
Bernier, P. Y. 1993. Comparing natural and planted black spruce seedlings. I. Water relationsand growth. Can. J. For. Res. 23: 2427–2434.
Blake, T. J. and Sutton, R. Y. 1988. Stomatal conductance; the key to adaptation in newlyplanted jack pine and black spruce. Plant Physiol. (Life Sci. Adv.) 7: 125–130.
Grossnickle, S. C. and Blake, T. J. 1986. Environmental and physiological control of needleconductance for bare-root black spruce, white spruce, and jack pine seedlings on borealcutover sites. Can. J. Bot. 64: 943–949.
Haase, D. L. and Rose, R. 1993. Soil moisture stress induces transplant shock in stored andunstored 2+0 Douglas-fir seedlings of varying root volumes. For. Sci. 39: 275–294.
Hahn, P. F. and Smith, A. J. 1983. Douglas-fir planting stock performance. Comparison afterthe third growing season. Tree Planters’ Notes, 34(1): 33–39.
Hook, W. R., Livingston, N. J., Sun, Z. J. and Hook, P. B. 1993. Remote diode shortingimproves measurements of soil water by time domain reflectometry. Soil Sci. Soc. Am. J.56: 1384–1391.
Hsiao, T. C., Acevedo, E. A., Fereres E. and Henderson, D. W. 1976. Stress metabolism: waterstress, growth and osmotic adjustment. Philos. Trans. R. Soc. Lond. B. 273: 479–500.
Kaufmann, M. R. and Thor, G. L. 1982. Measurement of water stress in subalpine trees: effectsof temporary tissue storage and needle age. Can. J. For. Res. 12: 969–972.
Kramer, P. J. 1983. Water Relations of Plants. Academic Press, New York. 489 p.Kramer, P. J. and Bullock, H. C. 1966. Seasonal variations in the proportions of suberized and
unsuberized roots of trees in relation to the absorption of water. Am. J. Bot. 53: 200–204.Lamhamedi, M. S. and Bernier, P. Y. 1994. Ecophysiology and field performance of black
spruce (Picea mariana): a review. Ann. Sci. For. 51: 529–551.Maass, D. I., Colgan, A. N., Cochran, N. L., Haag, C. L. and Hatch, J. A. 1989. Field
performance of five species in four different containers in Maine. North. J. Appl. For. 6:183–185.
Myers, B. J. 1988. Water stress integral – a link between short-term stress and long-termgrowth. Tree Physiol. 4: 315–323.
Newton, M., Cole, E. C. and White, D. E. 1993. Tall planting stock for enhanced growth anddomination of brush in the Douglas-fir region. New For. 7: 107–121.
Poorter, H. 1989. Plant growth analysis: towards a synthesis of the classical and the functionalapproach. Physiol. Plant. 75: 237–244.
Poorter, H. and Lewis, C. 1986. Testing differences in relative growth rate: A method avoidingcurve fitting and pairing. Physiol. Plant. 67: 223–226.
Ritchie, G. A., Tanaka, Y., Meade, R. and Duke, S. D. 1993. Field survival and early growthof Douglas-fir rooted cuttings: relationship to stem diameter and root system quality. For.Ecol. Manage. 60: 237–256.
Romero, A. E., Ryder, J., Fisher, J. T. and Mexal, J. G. 1986. Root system modification ofcontainer stock for arid land planting. For. Ecol. Manage. 16: 281–290.
nefo031.tex; 14/03/1997; 3:27; v.5; p.14

221
Rose, R., Gleason, J. F. and Atkinson, M. 1993. Morphological and water-stress characteristicsof three Douglas-fir stocktypes in relation to seedling performance under different soilmoisture conditions. New For. 7: 1–17.
SAS Institute Inc. 1993. SAS Software: Changes and Enhancements, Release 6.08. SASInstitute Inc., Cary, NC, USA.
South, D. B., Boyer, J. N. and Bosh, L. 1985. Survival and growth of lobolly pine as influencedby seedling grade: 13-year results. South. J. Appl. For. 9: 76–81.
South, D. B. and Mason, W. L. 1993. Influence of differences in planting stock size on earlyheight growth of Sitka spruce. Forestry, 66: 83–96.
Steel, R. G. D. and Torrie, J. H. 1980. Principles and Procedures of Statistics. A BiometricalApproach. McGraw-Hill Publishing Company, New York, 633 p.
Stewart, J. D. and Bernier, P. Y. 1995. Gas exchange and water relations of three sizes ofcontainerized Picea mariana seedlings subjected to atmospheric and edaphic water stressunder controlled conditions. Ann. Sci. For. 52: 1–9.
Tinker, P. B. 1976. Roots and water – transport of water to plant roots in soil. Philos. Trans. R.Soc. Lond. B. 273: 445–461.
Topp, G. C., Davis, J. L. and Annan, A. P. 1982. Electromagnetic determination of soil waterusing TDR. I. Application to wetting fronts and steep gradients. Soil Sci. Soc. Am. J. 46:672–678.
Walker, N. R. and Johnson, H. J. 1980. Containerized Seedling Field Performance in Albertaand the Northwest Territories. Environ. Can., Can. For. Serv., Northern For. Res. Cent.,Edmonton, Alta., Info. Rep. NOR-X-218, 32 p.
nefo031.tex; 14/03/1997; 3:27; v.5; p.15