effect of nucleotides on ribonucleic acid formation during development of the amphibian embryo
TRANSCRIPT

BIOCHIMICA ET BIOPHYSICA ACTA I19
BBA 8195
E F F E C T OF NUCLEOTIDES ON RIBONUCLEIC ACID FORMATION
DURING DEVELOPM ENT OF T H E AMPHIBIAN
EMBRYO
ELISABETH AMBELLAN" AND GEORGE WEBSTER'*
Department o] Biochemistry, The Ohio State University, Columbus, Ohio (U.S.A.)
(Received July 25th, 1962)
SUMMARY
i. Adenine nucleotide treatments that promote neural tube closure in Rana pipiens embryos also produce an increase in RNA synthesis.
2. The order of effectiveness of treatment materials on total RNA synthesis, on 14C nucleotide incorporation into RNA, and on the morphological advance is the same: greatest with ATP, next with ADP, least with adenosine 3'-monophosphate.
3. The differences in effectiveness between adenine nucleotides on morphogen- esis and on RNA synthesis is not related to penetration. Adenosine 3'monophosphate, the least effective material, penetrates IO times as fast as ATP or ADP.
4- Rapid penetration of adenosine 3'monophosphate is not simply related to its level of phosphorylation. Adenosine 3'monophosphate also penetrates io times as fast as adenosine 5'monophosphate.
5. Synthesis of RNA alone does not ensure advanced morphological develop- ment. ADP at pH 7.2 in R. pipiens does not promote neural tube closure, and adenosine 5'-monophosphate delays tube closure. In both cases excess RNA synthesis is promoted.
INTRODUCTION
The effectiveness of nucleotides in promoting neural tube closure in growing amphibian embryos has been reported previously1, *. The adenine nucleotides were more effective than other nucleotides, and the triphosphates more effective than the diphosphates 3. A-3-P, yeast RNA, nicotinamide, and DPN had slight effects, while A-5-P retarded tube closure 3. Adverse morphological side-effects of ATP treatment were largely reversible by addition of equimolar A-3-P to solutions ~. ADP treatment produced the most nolmal morphology during the precocious neurulation in the Rana pipiens species investigated. The effects were not simply related to penetration, because the
Abbreviations: A-3-P, adenosine 3'-monophosphate; A-5-P, adenosine 5"-monophosphate. * Present address: Laboratoire de Morphologie animale, Facult6 des Sciences de l 'Universit6
libre de Bruxelles (Belgique.) "" Present address: Insti tute for Enzyme Research, University of Wisconsin, Madison, VVis-
consin (U.S.A.).
Biochim. Biophys..4eta, 68 (1963) I i9-128

120 E. AMBELLAN, G. WEBSTER
advanced morphology could be observed many days after removal of embryos from treatment solutions, and because A-3-P, the least effective material, penetrates lO times as fast as ADP or ATP 3. This suggests that more complicated mechanisms must be involved.
Among the ways that nucleotides might promote differentiation, the most ob- vious is by promoting synthesis of new RNA molecules. The nucleotides, especially the triphosphates, are known precursors of RNA 4, and there is an increase in total RNA starting at neurulation in normally growing embryos 5. RNA is essential for pro- tein synthesis e, and new protein synthesis accompanies neural tube formation 7. The present experiments, therefore, were to determine the effects of nucleotide solution treatments on RNA synthesis as a function of the visible morphological acceleration.
MATERIALS AND METHODS
Embryos were prepared and handled as previously described 1, except when large num- bers of eggs were required, they we.re enzymically dejellied with cysteine-activated papain, according to the method of SPIEGAL t. Embryos were taken for biochemical comparisons when controls were at the flat neural plate stage (SHuMWAY, Stage I4), and treated animals of the same age, raised under the same c0r~ditions, were at a later stage of neural tube closure. The exact extent of morphological advance varied between experiments, because the time for ending an experiment was often arbitrary. In some experiments another set of controls, some hours older than the first two groups, were also analyzed in order to compare changes in normally neurulating embryos with changes in experimentally accelerated embryos.
RNA was extracted by. the acid hydrolysis method of OCOR AND ROSEN 9. Eggs were homogenized in io °/o Ringer solution, HC1Q added to a final concentration of 7 %, extracted for I h in the cold, and washed twice with 5 ~/o HC10~. After washing the precipitate with 95 % ethanol, lipids were removed with ethanol-ether (3 : i, v/v), and then extracted with io ~/o HCI04 for about 18 h in the cold, washed twice with 5 % HC10~, and the supernatants combined as the partially hydrolyzed RNA fraction. Further extraction of the precipitate with IO °/o HC104 at 9 °o for 15 rain constituted the DNA fraction. Quantitative detertninations of RNA were measured by ultraviolet absorption (absorbancy at 260-290 m/z), and calibrated against a standard curve of yeast RNA extracted in the same way. Ultraviolet analysis of RNA fractions gave typical nucleic acid spectra with maxima about 258 m/, and minima at 23 ° m# with- out major contamination from degraded protein. Paper chromatograms of RNA fractions, further hydrolyzed to free bases 1° showed the presence of all four RNA ba- ses without major contamination from other nucleic acid material (except for one spot also present in the yeast RNA standards) and no evidence of thymine. In some experiments orcino111 and cysteine-sulfuric acid lz color reactions for ribose were used. Orcinol tests of the DNA fraction were positive, and confirmed STEINERT'S finding 13 that RNA extraction by this method is, while relatively pure, only about 9 ° % com- plete in amphibian embryos.
The SCHMIDT-THANNHAUSER-ScHNEIDER 14 method of alkaline hydrolysis of the precipitate left after removal of acid-soluble and lipid materials was investigated. Spectral analysis of this acidified hydrolysate showed no resemblance to a typical nucleic acid curve, but rather high contamination with degraded protein. DAVlDSON
Biochim. Biophys. Acta, 68 (I963) 119-128

RNA FORMATION IN AMPHIBIAN EMBRYO 121
AND SMELLIE TM pointed out that this acidified fraction after alkaline hydrolysis, p.re- sumably containing RNA nucleotides, may contain as much as 25 % non-nucleic acid phosphorus. Thus, nucleic acid determinations based on phosphorus measure- ments of "RNA fractions", isolated by this method appear to be an unreliable measure of nucleic acids in amphibia, without exhaustive purification to remove contaminating proteins and non-nucleic phosphorus, and final determination of N/P ratios, as sug- gested by VOLI(IN TM.
After treatment with 1*C materials, eggs were washed several times with large volumes of IO % Ringer solution until no counts were detectable in 3-4 ml of wash, and then given an additional rinse. Washes were Checked after various extractions to prevent carry-over of labeled soluble materials. The HC10, extracts of acid-soluble and RNA fractions were neutralized with KOH, the precipitates removed by centri- fuging, and supernatant samples plated on glass planchets at infinite thinness (less than o.i rag/cruZ). Self-absorption of less than 2 % was neglected. Counting was done on a Model D-47 Nuclear Gas Flow Counter.
ATP, ADP and A-5-P were obtained as sodium salts from the Sigma Chemical Co. Purity was determined by spectral analysis, and by the correspondence between concentrations expected for a given weight and the molar extinction values. ADP con- sistently gave about 20 % less absorbancy value than expected for a given weight, but since the spectrum was typical for adenine, and chromatograms showed no contaminating nucleotides, the impurity was assumed to be water of hydration. A-3-P was obtained from Boehringer Co., and papain from Nutritional Biochemi- cals Co.
Radioactive adenine nucleotides, all labeled at C-8, were obtained from Schwartz Laboratories; sodium ATP (2.8/~C/mg); sodium A-5-P (o.7/~C/mg); sodium A-3-P (i.i/zC/mg); and lithium ADP (1.8/zC/mg). The lithium ADP was converted to so- dium ADP via the barium salt. Chromatography of these materials showed that [I*CIATP contained about IO % ADP, and that the [liCJADP contained about 15 % ATP. The monophosphates showed no contamination of other nucleotides.
RESULTS
Increase in total RNA
The total RNA content of control animals and of animals of the same age that were treated with ADP solutions are shown in Fig. I: Experimental animals with closed neural tubes contained, in this experiment, about 20 % more total RNA than control animals, raised in Io % Ringer solution, that were still at the flat neural plate stage. Triplicate samples showed differences of 3 %. Another group of untreated controls, about 8 h older than the first two series, allowed to grow to closed neural tube stage, also showed about a 2o % increase in total RNA over younger controls, confirming STEINERT'S data ~ that there is a normal increase in RNA at neurulation. More often, older untreated controls showed intermediate values of RNA between the younger control and experimental groups. Absolute amounts of RNA increases in experimental animals and older controls varied widely between different experiments, because the time for ending an experiment was arbitrary, requiring only some visible morphological advance in treated animals.
Increases in total RNA as measured by ultraviolet absorption were confirmed in
Biochim. Biophys. Acta, 68 (z963) II9--I28

122 E. AMBELLAN, G. WEBSTER
0.50-
0.40.
¢-
O "~ 0.30-
< 0.20- >; :D
o.10-
0 , , , , ,
C C 2 C 3 E 1 E 2 E 3 CC
Fig. I . T o t a l R N A in con t ro l a n d e x p e r i m e n t a l embryos . R N A was ex t r ac t ed b y t he m e t h o d of OGUR AND ROSEN, a n d m e a s u r e d b y u l t r av io le t ab so rp t i on ( abso rbancy a t 26o-290 m/z). Sample g rou l m con ta ined"4 ° eggs each ra ised in IO % Ringe r so lu t ion (pH 5.6).
C series: 3 se t s of con t ro l s a t f l a t neu ra l p la te s tage , age 6o h. E series: 3 se t s of e x p e r i m e n t a l e m b r y o s showing closed neura l t ubes , age 6o h, t r ea t ed for 24 h
w i t h 2 m M A D P (pH 5.6). CC: One g roup of u n t r e a t e d e m b r y o s w i t h closed neura l tubes , 8 b older t h a n the C and E series.
T A B L E I
T H E EFFECTS OF NUCLEOTIDE SOLUTION TREATMENT ON TOTAL R N A
R N A e x t r a c t e d as descr ibed in Fig. I ca l ib ra ted aga i n s t s t a n d a r d curve oI y e a s t RNA, Ul t rav io le t a lmorpt ion , a b s o r b a n c y 26o-29o m/z; orcinol, a b s o r b a n c y a t 66o-6oo m/z and cys t e ine - su l fu r i c ac id d e t e r m i n a t i o n s b y a b s o r b a n c y a t 39o-425 m # .
Descriptions: Controls A Controls B Experinteutals Age at z8°: 60 k 68 k 60 k Morphology: Neural plate Neural tube Neural tube
E x p t . I T r e a t m e n t None None 2 m M A D P R N A - U l t r a v i o l e t d e t e r m i n a t i o n s :
/~g R N A / e g g 4.07 4.84 5.28 Inc rease - - 19 % 29 %
Orcinol d e t e r m i n a t i o n s : /~g R N A / e g g 3.38 4.18 4.64 Inc rease -- 23 % 32 %
E x p t . 2 T r e a t m e n t None None 2 m M A D P R N A - U l t r a v i o l e t d e t e r m i n a t i o n s :
Inc rease -- 9 % 18 % Orcinol d e t e r m i n a t i o n s :
Inc rease - - 8 % 24 %
E x p t . 3 T r e a t m e n t None None 2 m M A T P R N A - U l t r a v i o l e t d e t e r m i n a t i o n s :
Inc rease -- -- 22 % Cys t e ine - su l fu r i c acid
d e t e r m i n a t i o n s : increase -- -- 22 %
some experiments by orcinol and cysteine-sulfuric acid color tests as is reported in Table I. The amount of RNA increase was approximately the same for any one ex- periment whether measured by ultraviolet absorption or color reactions. Treated ani- mals showed a 30 % increase in RNA in one experiment, and about a 20 % increase
Biochim. Biophys. Acta, 68 (1963) 119-128
RNA FORMATION IN AMPHIBIAN EMBRYO 123
in the two other experiments reported, Older untreated controls had intermediate values of RNA. Total RNA was about 5 #g/egg, about the same order of magnitude (2-15 ~ug/egg) as the European species investigated by STEINERT 5.
Comparative e//ects o/di//erent adenine nucleotides on total RNA levels Dependent upon which adenine nucleotides were present, differences in total
RNA content were found in experimental animals at the time of precocious neurula- tion. ATP produced an average increase in total RNA of 35 %, with a wide range of lO-8O % increase in over 30 experiments, and with occasional increases of 2-3 fold. ADP produced average RNA increases of 25 % with a range of lO-5O %. A-3-P had slightly less effect on total RNA synthesis. One comparative experiment is reported in Table II.
There seems little doubt from these results that total RNA increases markedly during neurulation, whether neurulation proceeds normally, or is promoted by addi- tion of nucleotides.
TABLE II ] E F F E C T O F A-3-P, A D P A N D A T P o N T O T A L R N A L E V E L S
Trea tmen t was for 2 4 h a t z8 °. Materials were added to xo % Ringer solution. R N A was extract - ed as described in Fig. I, and measured by ul traviolet absorp t ion (absorbancy at 260-29o m/~).
Samples Izg RNAIegg Percent increase
Controls 2.7 2 - - 4 mM ATP 5.11 88 4 mM A D P 3.52 29 4 mM A-3-P 3.08 22
Incorporation o~ [14C]nucleotides into RNA
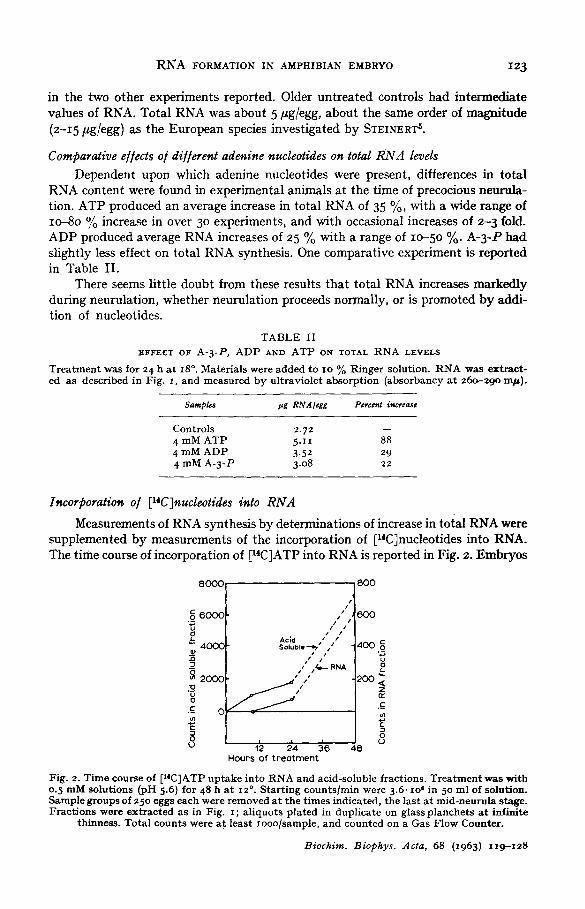
Measurements of RNA synthesis by determinations of increase in toial RNA were supplemented by measurements of the incorporation of [14C]nucleotides into RNA. The time course of incorporation of [14C]ATP into RNA is reported in Fig. 2. Embryos
8 0 0 0 : 8 0 0
.~ 6 0 0 0
'~" 4000 .ID 2 51 2ooo "0
-~ 0
0
/ / / i /s//
s I Acid i t / S °luble "-~/ /
/ i i / / t / ~ _ RNA
Hours of t r e a t m e n t
6 0 0
4oo g
L 2oo
Z rr ._¢
2 D O (9
48
Fig. 2. Time course of [14C]ATP up take in to RNA and acid-soluble fractions. T rea tmen t was with o.5 mM solut ions (pH 5.6) for 48 h at 12 °. S tar t ing counts /min were 3.6. lO 6 in 50 ml of solution. Sample groups of 25o eggs each were removed a t the t imes indicated, the last a t mid-neurula stage. Frac t ions were extracted as in Fig. i ; a l iquots plated in duplicate on glass p lanchets a t infinite
thinness. Total counts were a t least Iooo/sample , and counted on a Gas Flow Counter.
B i o c h i m . B i o p h y s . A c l a , 68 (z963) I I 9 - - 1 2 8

124 E. AMBELLAN, G. WEBSTER
were treated for 48 h prior to neurulation and the experiment was ended at mid-neu- rula stage. After 9 h of treatment (early gastrula) there were no detectable counts in RNA, but over IOOO counts/rain in the acid-soluble fractions. Radioactivity in RNA was first detectable after 24 h of treatment (end of gastrula). At some undetermined time between the 24 and 48-h periods measured (mid-neurula) there was a Io-fold increase in uptake of [14C]ATP.
Previously reported morphological studies ~ demonstrated that the effects of nucleotides on neurulation could be produced in about 9 ° rain when treatment was applied at the flat neural plate stage. Embryos were ~herefore treated with [14C]ADP at this stage (estimating a period of about 30 min before the visible effects of treat- ment might be expected to appear). Sample embryos were removed after 15, 3o and 60 rain. The results on 14C incorporation into RNA are shown in Table III. There was a 4-fold increase between the 15 and 6o-min periods:
TABLE II!
UPTAKE OF [14C]ADP INTO RNA AT START OF NEURULATION
Trea tmen t solut ion approx imated 0.5 mM ADP; wi th an initial act ivi ty of 1.2. lO 7 eounts /min/4o ml. R N A was extracted as described in Fig. i.
Sample Total counAslmin in RNA
I5 rain 32o 3 ° min 72o 6o rain lOO8
The experiments suggest that the rate of nucleotide incorporation into RNA depends to a large extent upon the stage of development and the synthetic activities, known and unknown, that occur at these stages. The contrast between these last two experiments (Fig. 2 and Table III), and the fact that both morphological effects and excess RNA synthesis can result long after removal of embryos from treatment, are compatible with the idea that the nucleotides remain in the free nucleotide pool (acid soluble) until the ond of gastrulation when RNA synthesis begins to increase rapidly, and only then are used for excess RNA synthesis.
Co,nmatiVe uptake o/ adenine nucleotides
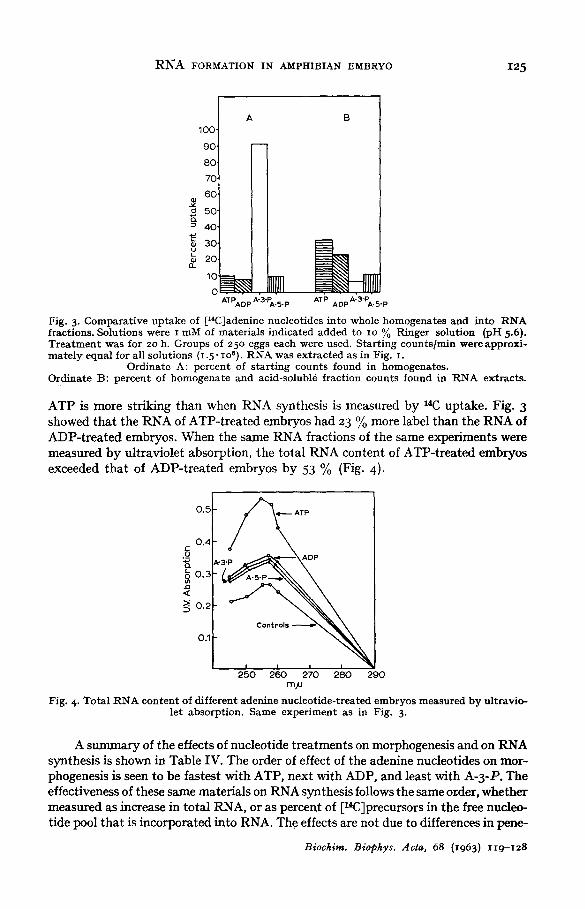
A comparison of incorporation of t14C]ATP, ADP, A-3-P and A-5-P into RNA is shown in Fig. 3. By far the largest percent of total counts taken up by the embryos was incorporated into RNA when ATP was used, the next largest incorporation was with [a4C]ADP, and the least with the [14C]monophosphates. That the uptake into RNA is not related to the available pool size is shown by the contrast of uptake into acid-soluble fractions, which also confirms previously reported data 8 that A-3-P penetrates IO times as fast as the other materials used. The contrast with A-5-P in Fig. 3 also shows that the rapid penetration of A-3-P is not simply related to the level of phosphorylation. I t is apparent that the failure of A-5-P to advance neurulation2, 3 is not due to its failure to be incorporated into RNA, for this slightly exceeds incor- potation of A-3-P into RNA.
When the RNA increase is measured by ultraviolet absorption, the influence of
Biochim. Biophys. Acta, 68 (I963) 119-128
RN'A FORMATION IN AMPHIBIAN EMBRYO I 2 5
A B 100'
9 0
80
70
6O
o 50
40
EL ~ 2o n
lO
(3 ATPAop m-pA.5.p ATP A~pA.3'~.5.p
gig. 3. Comparat ive uptake of [14C]adenine nucleotides into whole homogenates and into RNA fractions. Solutions were i mM of materials indicated added to IO % Ringer solution {pH 5.6). T rea tmen t was for 20 h. Groups of 250 eggs each were used. Star t ing counts/rain were approxi- mate ly equal for all solutions (i .5" Ioa) • I~NA was extracted as in Fig. I.
Ordinate A: percent of s tar t ing counts found in homogenates. Ordinate B: percent of homogenate and acid-soluble fraction counts found in RNA extracts.
ATP is more striking than when RNA synthesis is measured by 14C uptake. Fig. 3 showed that the RNA of ATP-treated embryos had 23 % more label than the RNA of ADP-treated embryos. When the same RNA fractions of the same experiments were measured by ultraviolet absorption, the total RNA content of ATP-treated embryos exceeded that of ADP-treated embryos by 53 % (Fig. 4).
O.~ ~ATP 0.4
8
~ 0.2
0.1 ~
250 260 270 280 290 m~u
Fig. 4. Total RNA content of different adenine nucleotide-treated embryos measured by ultravio- let absorption. Same experiment as in Fig. 3.
A summary of the effects of nucleotide treatments on morphogenesis and on RNA synthesis is shown in Table IV. The order of effect of the adenine nucleotides on mor- phogenesis is seen to be fastest with ATP, next with ADP, and least with A-3-P. The effectiveness of these same materials on RNA synthesis follows the same order, whether measured as increase in total RNA, or as percent of [l~C]precursors in the free nucleo- tide pool that is incorporated into RNA. The effects are not due to differences in pene-
Biochim. Biophys. Acta, 68 (r963) zz9--I28

I 2 0 E. AMBELLAN, G. WEBSTER
tration. I t can also be seen from Table IV that the effect of A-5-P treatment, which delays rather than advances neurulation, is not due to its failure to penetrate, or to induce RNA synthesis.
T A B L E IV
COMPARATIVE EFFECTS OF A D E N I N E NUCLEOTIDES ON MORPHOGENESIS AND ON R N ~ k SYNTHESIS
Percent increa,~e over controls Method of after treatment with
EHeds on measurement A T P A D P A-3-P A-5-P
Rate of morphogenes is Hours to close neural tubes 80 5 ° 25 --
Total R N & synthesis (average) Ultraviolet 35* 25 22 23
Incorpora t ion of 14C mater ia ls Percent of free into to ta l RNA nucleotide pool inc. 31 23 7 I I
Pene t ra t ion Percent of 1'C t r ea tmen t solutions in whole homogena tes X lO -3 IO 8 9 0 9
* Occasional increases in to ta l R N A after ATP t r e a t m e n t were 2-3-fold.
DISCUSSION
The present experiments clearly demonstrate the penetration of the [14C]- adenine moiety of added nucleotides into embryo cells. Ribose measurements that correspond quite precisely to ultraviolet measurements of increase in total RNA after treatment, indicate that the ribose penetrates to the same extent as the adenine part of the molecules. Phosphorus was not measured because of the difficulty in ob- taining reliable RNA measurements, using phosphorus determinations, after standard extraction methods. Labeled phosphorus is also difficult to follow in growing embryos during the period under study when numerous phosphate transfer enzymes are known to function. However, penetration of the entire nucleotide is assumed from the follow- ing aspects of the experimental results: the clear difference in morphological effects and in total RNA synthesis of ATP, ADP, A-3-P and A-5-P treatment; the reversi- bility of specific adverse morphological effects of ATP treatment with addition of equimolar A-3-P to solutions (embryos so treated resemble those treated with ADP alone ~) ; the lack of morphological effects of adenine or adenosine; and the pH depend- ence of the results 3. The possibility that the phosphate end groups of the nucleotides are eliminated at the membrane surface is not compatible with these results. While it has been reported that phosphorylated compounds do not pass cell membranes in some systems, it appears from the present data, and from the more complete list of effec- tive materials previously published 3, that the structural integrity of the base-ribose- phosphate bond is required for acceleration of morphology and for RNA synthesis in the in vivo amphibian system.
It was previously reported1, z that under optimum conditions morphological effects occur only at the start of neurulation, regardless of time of treatment, and can be observed many days after removal of embryos from treatment solutions. The pres- ent experiments indicate that treatment materials remain in the free nucleotide pool
Biochim. Biophys. Acta, 68 (i963) 119-128

RNA FORMATION IN AMPHIBIAN EMBRYO I27
until the end of gastrulation when increased RNA synthesis normally begins in untreat- ed controls. These combined results suggest that the initiation of RNA synthesis (and the enzymes essential for such synthesis) remain largely under genetic or DNA control, and are not affected by the added nucleotides. However, once the genetically controlled initiation of RNA synthesis has begun, added nucleotides can be used for excess RNA synthesis, as the present data show, and this in turn might induce pro- tein changes and syntheses known to occur in untreated embryos during neural tube closure 7.
If one way in which the nucleotides act to promote morphogenesis is b y an increase in net RNA synthesis, as when ATP, ADP or A-3-P is used, the converse is not true; that is, increased RNA synthesis alone is not sufficient to promote morphogenesis. This was noted with A-5-P which blocks neural tube closure, and with ADP at pH 7.2 which, in this species, had no visible effect 3, but in both cases increased total RNA. The resolution of these data could be that the specific nature of the excess RNA formed varies with the different nucleotides, and is a relevant factor in morphogenesis.
The morphological effectiveness of A-3-P, but not of A-5-P treatment has a parallel in data reported by WEINFELD, ROLL AND BROWN 17 who found that [14C]- A-3-P was twice as effective as [I*C]A-5-P as sources of the adenine and guanine moieties of both RNA and DNA molecules. These authors suggest that the major pathway of utilization of A-3-P involves conversion to some other derivative of adenosine, and not transformation into A-5-P, presumably the structural unit of RNA.
ATP treatment acts faster on neural tube closure and on induced RNA synthesis than ADP, and the contrast is greater when measured by ultraviolet absorption rather than by uptake of 14C precursors into RNA. This suggests that some additional role for ATP (possibly an indirect pathway leading to RNA synthesis not available to the di- or monophosphates) is detectable by ultraviolet absorption, but not as 14C uptake that measures only the adenine moiety common to all of the nucleotides. This might involve use of the triphosphates in adding phosphate to other bases or nu- cleosides that are in turn used in RNA synthesis. In this case, measurement of 14C uptake could detect only the more direct pathway of utilization of the adenine moiety, but ultraviolet measurements could detect both this and the indirect con- tribution of added phosphate to total RNA synthesis.
ACKNOWLEDGEMENTS
The co-operation of Professor JOHN PRICE and Professor HENRY PLAINE, and the Zoology Department of the Ohio State University is gratefully acknowledged. This investigation was supported by a research grant (C-3725) from the National Cancer Institute, U.S. Public Health Service, and by a Fellowship to one of us (E.A.) from the Division of General Medical Sciences, U. S. Public Health Service (GF-9374).
R E F E R E N C E S
1 E. AMBELLAN, Proc. Natl. Acad. Sci. U.S., 41 (1955) 428. * E. AMBELLAN, J. Embryot. Exptl. Morphol., 6 (1958) 86. 8 E. AMBELLAN AND G. C. WEBSTER, Develop. Biol., 5 (1962) 4.52.
D. B. STRAUS AND E. GOLDWASSER, J . Biol. Chem., 236 (1961) 849. 5 M. STEINERT, Bull. $oc. chim. biol., 34 (I952) 923. • J. BRACHET, Arch. Biol., 53 (194 I) 207.
I. LESL1E AND J. N. DAVlDSON, Biochim. Biophys. Acta, 7 (1951) 413 •
Biochim. Biophys. Acta, 68 (1963) 119-128

I 2 8 E. AMBELLAN, G. WEBSTER
8 M. SPIEGAL, Anat. Record, I i i (1951) 544- i M. OGUR AND G. ROSE~, Arch. Biochem., 26 (195o) 209.
10 G. W. CROSBIE, R. •. S. SMELLIE AND J. N. DAVIDSON, Biochem. J. , 54 (1953) 287. 11 W. ]~EJBAUM, Z. physiol. Chem. Hoppe-Seyler's, 258 (1939) 117. 1t Z. DISCHE, in E. CHARGAFF AND J. X~. DAVIDSON, The Nucleic Acids, Vol. I, A c a d e m i c P r e s s ,
N e w York, 1955, p. 3 °2. 1t M. STEINERT, Bull. soc. chim. biol., 33 (1951) 549- t* W. G. SCHNEIDER, J. Biol. Chem., 164 (1946) 747. It j . •. DAVIDSON AND R. M. S. SMELLIE, Biochem. J., 52 (1952) 599. 16 E, VOLKIN AND W. E. COHN, in E. GLICK, Methods o] Biochemical Analysis, Vol. I, In t e r sc i ence
Pub l i she r s , New York, 1954, p. 287. 17 H. WEINFELD, P. M. I~OLL AND G. W. BROWN, J. Biol. Chem., 213 (1955) 523 .
Bioehim. Biophys. Acta, 68 (1963) 119-128