effect of metals on β-actin and total protein synthesis in cultured human intestinal epithelial...
TRANSCRIPT

Journal of Pharmacological and Toxicological Methods 63 (2011) 47–58
Contents lists available at ScienceDirect
Journal of Pharmacological and Toxicological Methods
j ourna l homepage: www.e lsev ie r.com/ locate / jpharmtox
Original article
Effect of metals on β-actin and total protein synthesis in cultured human intestinalepithelial cells
Anthony R. Calabro, Dmitry I. Gazarian, Frank A. Barile ⁎St. John's University, College of Pharmacy and Allied Health Professions, Department of Pharmaceutical Sciences, Toxicology Division, Queens, New York 11439, USA
Abbreviations: 3H-D-M, [3H]-D-mannitol; As, arsenicmercury; Mn, manganese; Ni, nickel; PP, paracellular pelial electrical resistance; TJ, tight junction; TMER, trans-m⁎ Corresponding author. St. John's University Colleg
Parkway, Queens, NY 11439, USA. Tel.: +1 718 990 264E-mail address: [email protected] (F.A. Barile).
1056-8719/$ – see front matter © 2010 Elsevier Inc. Aldoi:10.1016/j.vascn.2010.04.012
a b s t r a c t
a r t i c l e i n f oArticle history:
Received 16 June 2009Accepted 27 April 2010Keywords:Caco-2 cellsCell cultureIn vitro cytotoxicityParacellular permeabilityTrans-epithelial electrical resistanceTrans-membrane electrical resistanceβ-actin
Introduction: As an important structural protein, β-actin is associated with anchoring of tight junctions(TJs) to the cell scaffold. Caco-2 cells, an immortal intestinal epithelial cell line, rely on β-actin to form intactmonolayers with high transepithelial electrical resistance in cell culture inserts. Methods: We examined theeffect of six metals on expression of β-actin mRNA and β-actin synthesis, on total and net production of newlysynthesized proteins, onparacellular transport of TJmarkers, andon cell viability in confluentmonolayers. [3H]-glycineand [3H]-tyrosine were used as indicators of newly synthesized proteins in the absence or presence of increasingconcentrations of arsenic, cadmium, copper, manganese, mercury and nickel. The monolayers were exposed to 24-hsingle exposures as well as continuous daily repeated doses of metals for 48-h and 96-h. Results: Results suggest thatdecreases in newly synthesized proteins, in which β-actin represents about 10%, correlated with 2- to 5-fold higherexpressionofβ-actinmRNA for thehigher concentrations ofmetals. Interestingly, IC50s calculated for each chemical for24-h acute and 48- and 96-h repeated dosing experiments, using theMTT viability assay and paracellular permeability
markers, decreased newly synthesized and total proteins to 10% and 40% of control, respectively. Discussion: Overall,the results indicate that, at equivalent concentrations, themetals affect β-actinmRNA and newly synthesized proteinsbefore cell viability and paracellular permeability are compromised. Consequently the results help in elucidatingmechanisms of metal cytotoxicity that lead to understanding the relationship between tight junction integrity,paracellular transport, and cell viability.© 2010 Elsevier Inc. All rights reserved.
1. Introduction
Widespread exposure to metals in the environmental or occupa-tional setting is associated with long term in vivo toxicity. In addition,studies of metal toxicity demonstrate highly selective activation ofstress signaling pathways leading to the development of humancancers (Kasprzak, Sunderman, & Salnikow, 2003). Alternatively,despite well-recognized carcinogenic potentials of such toxic metalsas chromium, nickel andmetalloid arsenic, themolecular mechanismsunderlying their cell-transforming ability remain poorly understood.Carcinogenic metals are typically weak mutagens and with theexception of chromium, they do not form DNA adducts, whichrepresent a key initiating event in the cancer-inducing activity oforganic carcinogens. As described below, the view that elevated
; Cd, cadmium; Cu, copper; Hg,rmeability; TEER, transepithe-embrane electrical resistance.e of Pharmacy, 8000 Utopia0; fax: +1 718 990 1877.
l rights reserved.
production of reactive oxygen species as the main pathway in metalcarcinogenicity is not supported with the current understanding ofthe mutagenic capacity of most metals.
For instance, occupational exposure to nickel (Ni) and arsenic (As)has been shown to increase the incidence of human cancers.Epidemiological studies have implicated Ni(II) compounds as humancarcinogens based upon an increased mortality from respiratory tractmalignancies in refinery workers chronically exposed to nickel-containing dusts and fumes (Salnikow & Zhitovich, 2008). Theproposed mechanism of the Ni-induced tumors centers aroundchanges in DNA methylation and histone acetylation, as well asinactivation or suppression of various DNA repair transcription factors(Karaczyn, Golebiowski, & Kasprzak, 2005; Kasprzak et al., 2003; Leeet al., 1995).
Human bioaccumulation of mercury (Hg) due to acute and chronicoccupational and environmental exposure has been implicated inneurological and psychological pathologies, as well as renal, immu-nological, and cardiovascular diseases (Jarup, 2003). In fact, a highdietary intake of mercury from consumption of fish is hypothesized toincrease the risk of coronary heart disease (Jarup, 2003). Interestingly,the toxicity associated with consumption of large amounts of methylmercury in seafood, with mercury burdens in the range of 9 to 24 µg,is well documented (Balshaw, Edwards, Daughtry, & Ross, 2007).

48 A.R. Calabro et al. / Journal of Pharmacological and Toxicological Methods 63 (2011) 47–58
Groundwater contamination with high concentrations of As (10 to100 µg/L) is associated with significantly higher mortality rates forcancers of the bladder, kidney, skin, liver, and colon (Tchounwou,Centeno, & Patlolla, 2004). A major mechanism by which As exerts itstoxic effect is through impairment of cellular respiration anduncoupling of oxidative metabolism. Additionally, in vitro studieshave linked the carcinogenic effect to the ability of As to induce geneamplification, arrest cells in mitosis, inhibit DNA repair, and stimulateexpression of c-fos gene in mammalian cells (Huang, Huang, & Lee,2000). Although As carcinogenesis is influenced by cellular metabo-lism and duration of exposure, the risks for human cancer transpire atvarious concentrations and environmental levels (Salnikow & Zhitovich,2008).
Occupational and environmental exposure to cadmium (Cd), as ahuman carcinogen, is linked to lung, bladder, liver, kidney, stomach,and prostate cancers (Waalkes, 2000). Because it is poorly mutagenicat low doses, explanations for Cd carcinogenicity relies on itsepigenetic mechanism, namely interference with apoptotic geneexpression, inhibition of DNA repair enzymes, and up-regulation ofmitogenic signaling (Waalkes, 2000).
In vivo, exposure to manganese (Mn)-containing compoundsresults in neurological and developmental toxicity at even moderatedoses, while in mammalian cells, higher doses of Mn cause DNAdamage and chromosome mutations. Interestingly, however, it is notregarded as a human carcinogen (Gerber, Léonard, & Hantson, 2002).
Finally, interference with normal physiological roles of copper(Cu) has been correlated with human cancers. For instance, increasedexpression of copper-transporting ATPase genes (Cu-ATPases), whichare essential for human cell growth and development, has beenobserved in some human cancer specimens, and may be associatedwith tumorigenesis and chemotherapy resistance (Zhang, Li, Yao, &Chen, 2009). Recently, Turski and Thiele (2009) have demonstratedthat a family of copper-dependent intracellular lysyl oxidase-likeproteins (LOXL1-4) interferes with Snail repressor proteins, the latterof which normally regulate the transcription of E-cadherin tomaintain cell–cell adhesion. The authors suggest that hyperactivityof these copper-bound cofactors (LOXL2 in particular) diminishesglycogen synthase kinase 3β-mediated phosphorylation, therebyinhibiting E-cadherin expression, and compromising tight junctions.Clinical human studies have also confirmed the role of elevated LOXL2expression in highly invasive forms of metastatic breast, colon, andesophageal cancers (Turski & Thiele, 2009).
In vivo and in vitro, epithelial cells regulate paracellular perme-ability by forming junctional complexes between the cells. Thejunctional complexes consist of desmosomes, adherens junctionsand tight junctions, the latter of which contribute to the barrierfunction of the epithelial membrane. Specifically, as a “house-keeping” protein in stationary mammalian cells, β-actin is an integralcomponent in maintaining the integrity of tight junctions (TJs). TJsfasten adjacent epithelial cells and regulate the passage of ions andwater between cells (Gonzalez-Mariscal, Betanzos, Nava, & Jaramillo,2003). As TJs are particularly important in the intestinal lumen andproximal renal tubule, chemicals that interfere with β-actin synthesiscan alter TJ integrity and selectively impair absorption and intercellulartransport (Fanning, Jameson, Jesaitis, & Anderson, 1998).
Several in vitro studies have shown reduced paracellular function-ality in the presence of metal exposure. For instance, Rossi et al. (1996)demonstrated that Cd, Cu and zinc (Zn) alter Caco-2 cells' ability totransport specific markers across the cell monolayer. Exposure to themetals also resulted in morphological alterations of TJ integrity asdemonstrated by changes in the transepithelial electrical resistance(TEER) across themembrane. Similarly, Duizer, Gilde, Versantvourt, andGroton (1999) showed that the decreased epithelial barrier function ofintestinal epithelial cells is accompanied by tight junction disruption.They concluded that Cd+2 causes increasedparacellularpermeability bydisruption of junctional function and structure. Ferruzza, Scacchi,
Scarino, and Sambuy (2002) present data that suggest differentmechanisms are responsible for the changes in TJ permeabilityproduced by Cu and iron exposure in Caco-2 cells. They report that Cu(II) led to an overall reduction in F-actin staining with extensivedepolymerization in areas of the monolayer formation on semi-permeable membranes in the absence of cellular loss, whereas Fe(II)treatment produceddisorganization of F-actin and decreased stainingofthe perijunctional actin filaments. Interestingly, no changes in localiza-tion of the junctional proteins ZO-1, occludin and E-cadherin wereobserved after treatment with 100 mm FeSO4. Natoli et al. (2009)showed that acute exposure to 50 µM of Fe(II)ascorbate in Caco-2 cellsresulted in damage to tight junctions. The toxicity was representedby an increase in cell monolayer permeability and by partialdelocalization of the tight junction protein, claudin, from the plasmamembrane to an intracellular compartment. And, Navarro-Moreno et al.(2009) show that subacute lead intoxication alters renal tubularepithelial cell structure through loss of apical microvilli, decrement ofthe luminal space and the respective apposition and proximity of apicalmembranes, and formation of atypical intercellular contacts andadhesion structures. Alternatively, in vivo studies show that manganesedisplays a unique behavior with regard to its toxicity. Althoughmanganese does not represent a significant carcinogenic risk to thepopulation through occupational exposure, themetal causes Parkinson-like symptoms when inhaled even at moderate amounts over longerperiods of time (Gerber et al., 2002). However, the metal precipitatesDNA damage and chromosome aberrations and large amounts oforganic manganese derivatives are embryotoxic in mammals. Interest-ingly, Prozialeck, Edwards, Lamar, and Smith (2006) argue that severalproximal tubule-derived cell lines, commonly used as model systemsfor in vitro toxicity studies, express varied levels of cell adhesionmolecules (E-cadherin, N-cadherin, ZO-1, occludin, and claudins-2and -7), while some cell lines have weak claudin-7 and occludinexpression. They conclude that proximal tubule-derived cell linesexhibit characteristics that most closely resemble those of the proximaltubule in vivo, and that these cell lines are appropriate models forstudying the effects of toxicants on cell–cell junctions and cell adhesionmolecules. In a separate review, Prozialeck and Edwards (2007)summarize the effects of a variety of toxic substances, including metalssuch as Hg and Cd, on gap junctions and cell adhesion molecules inepithelial cell models, both in vitro and in vivo. Furthermore, theyelucidate the role of integrins and connexins in maintaining junctionalintegrity in the presence of toxicant and metal exposure. Overall, it iswell understood, although not frequently justified in explanations oftoxicological intercellular mechanisms, that preserving the integrity ofgap junctional intercellular communication (GJIC) is essential inmaintaining normal cellular development. Thus, the carcinogenic effectof chemical teratogens and tumor-promoting chemicals, inflammatoryfactors, hormones and growth factors, may be explained in part byinterfering with GJIC. In progenitor cells, the chemicals alter prolifer-ation, apoptosis or senescence, while in terminally differentiated cells,these chemicals alter gene expression or apoptosis and possibly de-dedifferentiation. This epigenetic pathway may explain why an“initiated” stem cell would be prevented from terminally differentiatingor from dividing asymmetrically in the presence of chemical carcino-gens, teratogens, reproductive toxicants and neurotoxicants. That is,the chemicals alter gene expression, in either stem cells, progenitorcells or terminally differentiated cells, by transcriptional, translationalor post-translational modifications, thus interfering with intercellularcommunication (Trosko, 2007).
We have studied the human intestinal epithelial cell line, Caco-2,and mortal rat intestinal cells, as in vitromodels for acute cytotoxicityand paracellular permeability for a variety of cytotoxic compounds(Konsoula & Barile, 2005, 2007). In cell culture, Caco-2 cells formintact monolayers on semi-permeable inserts that yield an organiza-tional arrangement suitable for examining the effects of metalexposure on tight junction integrity. Based on the effects of metals

49A.R. Calabro et al. / Journal of Pharmacological and Toxicological Methods 63 (2011) 47–58
on cell adhesion molecules as noted above, we postulate thatmonitoring tight junction formation and stability provides a methodfor detecting subtle changes in membrane integrity as a result ofmetal exposure. Since β-actin serves as one of the main structuralanchors for tight junctions, any disruption in β-actin gene expressionor protein synthesis reflects interference with paracellular transportand compromise of the epithelial barrier. To this end, we comparedthe effects of 24-h single exposures with 48- and 96-h repeated,continuous dosing of cultured cells with six metals, while monitoringproduction of newly synthesized proteins, β-actin synthesis and geneexpression, paracellular transport and cell viability.
2. Materials and methods
2.1. Cell culture
Cell culture supplies were obtained from LIFE technologies(Carlsbad, CA, USA), and from VWR (Bridgeport, NJ, USA). Chemicals(N99.9% purity) were obtained from Sigma Chemical Co., (St. Louis,MO, USA) and from Alfa Products (Ward Hill, MA, USA). Immortalhuman colon epithelial cells (Caco-2, American Type CultureCollection; Rockville, MD, USA), passage numbers 22–40, were sub-cultured and seeded at 104 cells/cm2 in 96-well plates or onto 6-wellor 12-well plates fitted with BD Falcon polyethylene terepthalate(PET) culture plate inserts (5×104 cells/insert). Cultures were grownin Dulbecco's modified Eagle's medium supplemented with 10% fetalbovine serum (DMEM-10), 1% non-essential amino acids (NEAA), 1%glutamine, 50 U/ml penicillin and 50 μg/ml streptomycin in 7.5% CO2
in air. All metal exposure experiments were performed on confluentmonolayers.
2.2. Incorporation of radio-labeled precursors into newly synthesizedproteins
Confluent monolayers of Caco-2 cells were exposed to increasingconcentrations of metals for 24-h and incubated with [2-3H]-glycine(21 Ci/mmol) or L-[3,5-3H]-tyrosine (51 Ci/mmol)(Amersham Bio-Chemicals, GE Healthcare Bio-Sciences Corp., Piscataway, NJ, USA) inMEM without FBS, to a final concentration of 1.0 µCi/ml. Cells wereharvested and radioactivity was measured as previously described(Barile & Cardona, 1998; Barnes, Houser, & Barile, 1990).
2.3. Quantification of mRNA expression for β-actin
Total mRNAwas purified using Invitrogenmicro-to-midi total RNApurification kit (Carlsbad, CA, USA). Primers for β-actin were obtainedfrom Applied BioSystems (Foster City, CA, USA). Multiplex RT-PCR forβ-actin fragment (295 bp) was performed in the same tube as theinternal standard (IS; 498 bp) using Invitrogen SuperScript IIITM one-step system with Platinum®Taq DNA Polymerase, as previouslydescribed (18 S rRNA was amplified with a primer–competimercombination using QuantumRNA™ Classic IS Kit [Ambion Inc, Austin,TX, USA]) (Gupta et al., 2005; Kebache, Zuo, Chevet, & Larose, 2002;Washer, Law, Boland, Horon, & Feinberg, 1996). Electrophoresis ofPCR products was performed on 1% agarose gels for 1-h at constant100 v and stained with ethidium bromide. Band intensity wasmeasured relative to IS with the UVP Biospectrum 600 ImagingSystem® (UVP, Upland, CA, USA). Ratio of the band intensities for β-actin relative to IS was calculated as a semi-quantitativemeasurementof mRNA synthesized according to mRNA extracted per sample.
2.4. Quantification of β-actin synthesis and total protein
Synthesis of total β-actin protein was performed using thePathscan® Total β-actin Sandwich ELISA kit (Cell Signaling Technol-ogy, Inc.; www.cellsignal.com). The method is based on a solid phase
sandwich enzyme-linked immunosorbent assay (ELISA) that detectsendogenous levels of β-actin. Reactions are measured by readingabsorbance at 450 nm on the BioTek FL600® fluorescence/absorbanceplate reader (BioTek Instruments Inc., Winooski, VT, USA).
Total protein was determined using the Pierce BCA (bicinchoninicacid) protein assay reagent kit (Thermo Scientific, Rockford, IL, USA).Absorbance of final protein product was read at 562 nm.
2.5. MTT cell viability assay
Median effect analysis of cell viability for metal cytotoxicity wasperformed as previously described (Konsoula & Barile, 2005). Briefly,cells were grown to confluence in 96-well plates and exposed in theabsence or presence of increasing concentrations of metals, at 37 °C inan atmosphere of 7.5% CO2 in air. Viability was evaluatedwith theMTT(3-(4,5-dimethylthiazolyl-2)-2,5-diphenyl-tetrazolium bromide)assay (cell counts were simultaneously performed using a hemocy-tometer). Absorbance was read at 550 nm using the BioTek FL600®
fluorescence/absorbance plate reader and viability was expressed as apercentage of the control group. Determinations from each group-concentration represent the mean of six separate wells (wells withmedia only served as blanks) and each experiment was repeated atleast three times. For continuous dosing assessment, confluentmonolayers of Caco-2 cells were incubatedwith increasing concentra-tions of each chemical for 48- and96-h. Freshmedium (with chemical)was replaced in each well daily until the end of the experiment.
2.6. Trans-membrane electrical resistance (TMER) measurements
Caco-2 cells were seeded onto 6- or 12-well plates fitted with PETculture plate inserts. DMEM, supplemented as above, was added to theapical and basolateral chambers and replenished three times a week.Cultures were confluent at 7-days, but maximum resistance values(between 750 and 1000 Ω-cm2) were achieved in intact monolayersafter 15-days. Trans-membrane specific electrical resistance (TMER)was measured as previously described (Calabro, Konsoula, & Barile,2008) using the Millicell-ERS® resistance system (Millipore) beforeand after 24-, 48- and 96-h incubation with test chemicals. Blanks(inserts without cells containing media and chemical) were used todetermine baseline values, and are expressed as percent TMER ofuntreated control groups.
2.7. Paracellular permeability (PP) assay
Cells were seeded onto 6- or 12-well plates fitted with PET cultureplate inserts, as described above for TMER experiments, and incubatedwith test chemicals for 48- and 96-h. Newmediumwith test chemicalswas introduced each day until the end of the assay. Low- and high-molecular weight indicatorswere added to thewells in the last 90 minof incubation. D-[1-3H]-mannitol (3H-D-M; 20 Ci/mmol; 9:1 inethanol; mw=182) has low lipophilicity and lucifer yellow (LY,mw=450) is more hydrophobic; however, both permeate via theparacellular route. To each apical chamber of an insert, 150 µl ofDMEM-10was added plus HEPES, mannitol (0.1% w/v), and [3H]-D-M,to a final concentration of 1.0 µCi/ml (ethanol solvent was evaporatedbefore application; Konsoula & Barile, 2005). Simultaneously, DMEMwithout radioactivity was dispensed into the basolateral chamber. Atthe end of the incubation period, passage of [3H]-D-M through themembrane was determined by dissolving an aliquot of the basolateralmedium in Budget-solve® and measuring the cpm by liquid scintil-lation (MicroBeta TriLux scintillation counter, Perkin-Elmer AnalyticalSciences, Shelton CT, USA). Inserts without cells (blanks) and controlcultures (minus chemical) were monitored simultaneously. Sampleswere also taken from the apical solutions to measure the initial donorconcentrations. Background radioactivity was determined using

Table 1IC50 (mmol/L) for Caco-2 cells using MTT assay: 24-h single dosing and 48- and96-h repeated dosing exposures.
Metals MTT 24-h MTT 48-h MTT 96-h
Arsenic trioxide 0.068 0.0131 0.0096Cadmium chloride 0.109 0.041 0.034Cupric sulfate 1.566 0.670 0.439Manganese chloride 9.6 8.186 0.356Mercury chloride 0.019 0.333 0.242Nickel chloride 0.968 0.820 0.778
50 A.R. Calabro et al. / Journal of Pharmacological and Toxicological Methods 63 (2011) 47–58
DMEM, and dpm were calculated based on the instrument's countingefficiency for [3H] (∼45%).
Lucifer yellow was used at concentrations of 1 mg/ml in DMEMand applied to the apical chamber. At the end of the exposure/incubation period, the basolateral medium was collected and theconcentration of LY was determined by measuring fluorescenceintensity (BioTek FL600 fluorescence/absorbance microplate reader).Inserts without cells (blanks) were also monitored simultaneously.Excitation and emission wavelengths for LY are 430 nm and 540 nm,
Fig. 1. Concentration-effect graphs of confluent monolayers of Caco-2 cells exposed toincreasing concentrations of: A. cadmium, B. copper, C. nickel, D.manganese, E.mercury,F. arsenic, for 24-h single dose exposures and 48- and 96-h repeated dose exposure. Cellswere processed for the MTT assay as described in Materials and methods. Results areexpressed as percent of control (nometal exposure). Each bar represents themean±SEof six replicates from one of at least 3 representative experiments.
Slope (m) and regression (R2) calculations for treatment groups are noted below.All plots were analyzed for one-way ANOVA (pb0.001); SEM of percent values areshown in Table 3; *Significant at pb0.001.
IC50 data R2 m
24-h (y) vs. 48-h (x) 0.99* 1.224-h (y) vs. 96-h (x) 0.97* 2.748-h (y) vs. 96-h (x) 0.98* 2.3
respectively. As with the PP assay, fluorescence values for treatmentgroups were compared to control cell cultures. Relative cell perme-ability is expressed as a percent of untreated control groups.
2.8. Statistical analysis
IC50s were extrapolated from concentration-effect curves usinglinear regression analysis (Fig. 1A–F). Calculation of IC50 value for eachchemical is based on a minimum of 3 experiments performed atseparate times, 6 to 8 groups (concentrations) per chemical (includingcontrol and blank), and 6 separate measurements (samples) pergroup, (N=3; sample number of measurements per experiment=144).When the IC50 was not bracketed in the initial dosage range usedfor the chemical, the experiments were repeated and the concentra-tions were adjusted as necessary.
For statistical significance (Pb0.05), the coefficient of determina-tion (R2), slope (m), and t-statistic (two-tailed paired Students' t-testwith the more stringent equal variances assumption) were calculated(Microsoft Excel®). All experimentswere repeated at least three times.Since the data generated from the β-actin synthesis experimentsweredetermined to be of high precision and cost, these repetitions wereperformed only twice (Table 3). Values in all figures and tables areexpressed as percentage of untreated controls, unless otherwise noted.
3. Results
Table 1 shows MTT IC50 data generated from exposure of Caco-2cells to 24-h single dosing and 48- and 96-h repeated dosing with
Table 2IC50 (mmol/L) for Caco-2 cells using TMER measurements: 24-h single dosing and 48-and 96-h repeated dosing exposures.
Metals TMER 24-h TMER 48-h TMER 96-h
Arsenic trioxide 0.13 0.0051 0.0045Cadmium chloride 0.049 0.030 0.022Cupric sulfate 1.8 0.578 0.473Manganese chloride 3.082 8.740 5.204Mercury chloride 0.09 0.181 1.192Nickel chloride 8.4 0.673 0.757
Slope (m) and regression (R2) calculations for treatment groups are noted below.All plots were analyzed for one-way ANOVA (pb0.001); SEM of percent values areshown in Table 3; * Significant at pb0.001.
IC50 data R2 m
24-h (y) vs. 48-h (x) 0.04 0.1824-h (y) vs. 96-h (x) 0.04 0.3148-h (y) vs. 96-h (x) 0.96* 1.7

51A.R. Calabro et al. / Journal of Pharmacological and Toxicological Methods 63 (2011) 47–58
metals. IC50s were calculated based on concentration-effect curves(Fig. 1A–F). All experiments were performed until the regression (R2)for the percent of control vs. log of concentration was greater than0.90 for at least 6 concentrations including control. Comparison of 24-,48-, and 96-h IC50 values were not significantly different from eachother (paired Students' t-test). However, comparisons using one-wayANOVA, slopes (m) and regressions for the lines were significantlydifferent (*). Using these statistical indicators, IC50 values for 48- and96-h repeat dosing were lower than 24-h data, suggesting that cellswere more sensitive to 48- and 96-h exposures. It is important to notethat in all assays, the steeper slopes of the lines suggest that the 24-
Fig. 2. Incorporation of [2-3H]-glycine by confluent monolayers of Caco-2 cells exposed to inharvested and radioactivity was measured as described in Materials and methods. Logaritexpressed as a percent of the control cells (nometal exposure). Each bar represents themeandifferent from control group, Pb0.05.
h assays (y-values), in general, require higher doses than 48- or 96-h assays (x-values) for determination of cell viability [in thecalculation of slope on log scale, when m is greater than 1.0, as inTable 1, the line is shifted to the left and the y-values (24-h) are higherthan corresponding x-values (48- and 96-h) (Konsoula & Barile,2005)]. It is not unreasonable to conclude therefore that, at equivalentdoses, cell viability is affected to a greater extent with prolonged,repeated exposures.
Table 2 summarizes the IC50 data generated from TMER studiesfrom exposure of Caco-2 cells to 24-h single dosing and 48- and96-h repeated dosing with metals. In contrast to the MTT assay
creasing concentrations of: A. Cd, B. Ni, C. Cu, D. Mn, E. Hg, and F. As, for 24-h. Cells werehmic concentrations are based on IC50s determined with the TMER assay. Results are±SE of six samples. Each experiment was repeated at least three times. *=significantly

52 A.R. Calabro et al. / Journal of Pharmacological and Toxicological Methods 63 (2011) 47–58
results, there were no significant differences between acute andrepeated dosing experiments. Only the slopes and regression valuesfor 48- vs. 96-h plots (m=1.7, R2=0.96)were significantly different.Consequently, transmembrane electrical resistance is not signifi-cantly affected over a 24- to 96-h dosage range. As noted above,therefore, although there were no significant differences amongTMER assays (with one exception), the flatter slopes of the lines onlog scale reflect a shift to the right (Table 2:m=0.18 and 0.31, for 24-vs. 48-h and 24- vs. 96-h, respectively), suggesting that the 24-h single concentrations (y-values) are lower than corresponding 48-
Fig. 3. Incorporation of L-[3,5-3H]-tyrosine by confluent monolayers of Caco-2 cells exposedwere harvested and radioactivity was measured as described in Materials and methods. LogResults are expressed as a percent of the control (no metal exposure). Each bar represen*=significantly different from control group, pb0.05.
or 96-h repeated concentrations (x-values) [whenm, orΔy/Δx, is lessthan 1.0, the data line is shifted to the right, thus x-values are higherthan corresponding y-values (Konsoula & Barile, 2005)]. Thus thedata indicate that repeated 48- or 96 h dosing does not affect TMER assignificantly as cell viability, and confirms our previous conclusionsthat cell viability is disrupted before PP is compromised (Konsoula &Barile, 2005; Calabro et al., 2008).
Fig. 2A through F summarize the results of [3H]-glycine incorpo-ration into net production of newly synthesized proteins in theabsence or presence of six metals. A significant reduction (N50%) of
to increasing concentrations of: A. Cd, B. Ni, C. Cu, D. Mn, E. Hg, and F. As, for 24-h. Cellsarithmic concentrations are based on IC50s determined with the TMER assay (Table 2).ts the mean±SE of six samples. Each experiment was repeated at least three times.

53A.R. Calabro et al. / Journal of Pharmacological and Toxicological Methods 63 (2011) 47–58
radioactivity was detected with all metals at concentrations of 10−5-to 100-times their mean IC50 TMER concentrations. It is important tonote that 5 metals and arsenic demonstrated greater than 95% or 70%reduction of glycine incorporation, respectively, at the IC50 valuesdetermined with the TMER assay for the corresponding metal,although significant differences from control were detected at evenlower concentrations.
Fig. 4. Expression of gene transcripts for internal standard (IS) and β-actin for confluent monE. Hg, and F. As, for 24-h. Cells were harvested, mRNA was extracted, and RT-PCR amplificaconcentrations are based on IC50s determined with the TMER assay (Table 2). Bar graphs repractin is 295 bp and for the 18S rRNA IS is 498 bp, relative to the DNA ladder (not shown). Amoto attain a final ratio of approximately 1.0 (dotted line). Each bar represents the mean±SE
Fig. 3A through F summarizes the results of [3H]-tyrosineincorporation into net production of newly synthesized proteins inthe presence of six metals. As with [3H]-glycine incorporation, asignificant reduction (N50%) of radioactivity was determined for [3H]-tyrosine with all metals at concentrations 10−5- to 100-times theirmean IC50 TMER concentrations. In general, the decreases parallelthose seen with [3H]-glycine; 5 metals and arsenic demonstrated
olayers of Caco-2 cells exposed to increasing concentrations of: A. Cd, B. Ni, C. Cu, D. Mn,tion of transcripts was performed as described in Materials and methods. Logarithmicesent the ratio of β-actin/IS as relative band intensities; the expected PCR product for β-unt of PCR product applied to eachwell was determined empirically for the control cellsof three to six samples and each experiment was repeated at least three times.

54 A.R. Calabro et al. / Journal of Pharmacological and Toxicological Methods 63 (2011) 47–58
greater than 95% or 18% reduction of tyrosine incorporation,respectively, at the IC50 value determined with the TMER assay forthe corresponding metal.
Fig. 4A through F illustrates the amplification of β-actin genetranscripts from extracted mRNA expressed from cell cultures during24-h exposures to increasing concentrations ofmetals. Image analysis ofband intensities was used to calculate semi-quantitative determinationsof β-actin/IS ratios (control ratios were empirically adjusted so that β-actin/IS ratio would approximate 1.0 upon application to gels, andsample aliquots that were applied to each well were identical). Moreimportantly β-actin expression, relative to IS, is based on the samequantity of mRNA starting material extracted from treated cultures,rather than on the number of cells—i.e. although total amount of mRNAextracteddecreases in proportion to cell numbers in the treated cultures,the expression of β-actin/IS increases per unit of mRNA isolated.
All metals demonstrated a 1.5 to 3.0-fold increase in β-actin mRNAexpression (relative to IS) at concentrations comparable to thoseshowing decreases in newly synthesized proteins. Almost none of thedoses used demonstrated gene expression significantly below controlvalues (*; only Fig. 4C, copper, is below 1.0 and significantly differentfrom control). Thus, as cell numbers and newly synthesized proteinsdecrease in the presence of trace metals, synthesis of mRNA for β-actin rises, (standardized to control mRNA synthesis).
Figs. 5, 6 and 7 illustrate the representative data obtained fromexposure of Caco-2 cells to metals using passive paracellularpermeability (PP) markers. The concentrations are based on themost effective IC50s determined experimentally for either the MTT(Table 1) or TMER (Table 2) studies as well as the upper and lowerlimits (data not shown). Analysis of the data indicates thatparacellular transport of [3H]-D-mannitol, as a tight junction marker,
Fig. 5. Effect of: A. Cd 48-h, B. Cd 96-h, C. Cu 48-h, and D. Cu 96-h, on paracellular transport instudies. Scale for Percent of Control Response for PP markers (bars) are on left axis; for TMER100% (bars) or based on 1000 Ω-cm2 (TMER). LY = lucifer yellow, [3H]-D-M = [3H]-D-man
more closely parallels the fall in TMER than LY, indicating thattransport of [3H]-D-mannitol to the basolateral surface correlatesbetter with a decrease in resistance. This correlation, and the con-centrations used, confirms our previous acute cytotoxicity resultswith these metals and other chemicals using a variety of PP markers(Konsoula & Barile, 2005). In only a few instances, such as withcadmium (Fig. 5A) and manganese (Fig. 6A,B), is LY transported at agreater rate than [3H]-D-mannitol, as TMER decreases. These resultsare probably associated with effect of Cd or Mn on non-lineartransport of low molecular weight molecules, both of which areknown to interfere with passage through Ca+2 channels.
Table 3 summarizes and compares the results of experimentsperformed at 24-h exposures with six metals at single concentrationsrepresented by the 24-h IC50 for TMER experiments. Values aresystematically compared by percent of control for: cell viability, (MTTassay), newly synthesized proteins (3H-glycine and 3H-tyrosineincorporation), mRNA β-actin gene expression and total β-actinsynthesis (PCR data and ELISA assay, respectively), cell proliferation(cell counts), total protein synthesis, and amount of proteinsynthesized per unit cell (per 104 cells). Each data point representsthe mean of at least 3 experiments incorporating 6 concentrations permetal and involving 6 replications per concentration (wells pergroup-concentration) [experiments for β-actin, cell counts, and totalprotein were performed twice]. Close analysis reveals that: 1. exceptfor Cd, MTT values are lower than the concentrations necessary toreduce TMER by 50% (TMER IC50 concentrations), suggesting that cellviability decreases before PP is compromised. This occurs mostdramatically for nickel—i.e. 8.4 mM lowers TMER by 50% butdecreases cell viability to 16% of control (Table 3). This data confirmsour previous results for several diverse compounds with both Caco-2
Caco-2 cells. Concentrations correspond to IC50s generated with either theMTT or TMERmeasurements (dashed–dotted line), scale is on the right axis. Control values are set atnitol, TMER = transmonolayer electrical resistance.
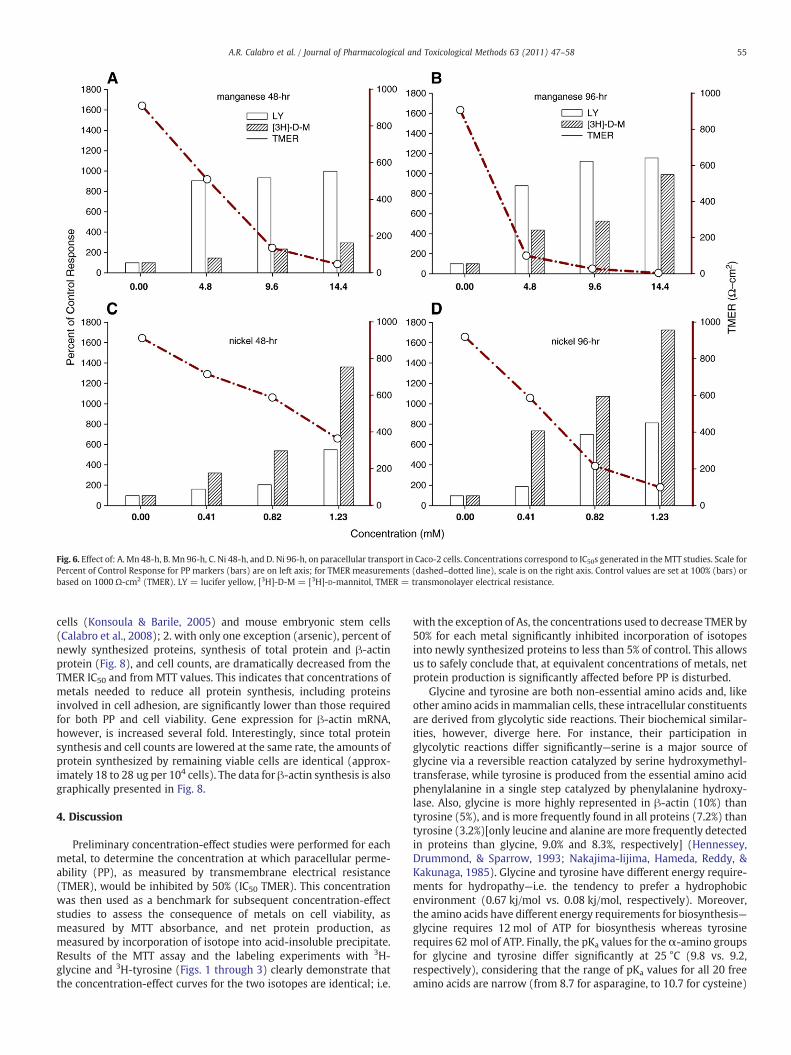
Fig. 6. Effect of: A. Mn 48-h, B. Mn 96-h, C. Ni 48-h, and D. Ni 96-h, on paracellular transport in Caco-2 cells. Concentrations correspond to IC50s generated in theMTT studies. Scale forPercent of Control Response for PP markers (bars) are on left axis; for TMER measurements (dashed–dotted line), scale is on the right axis. Control values are set at 100% (bars) orbased on 1000 Ω-cm2 (TMER). LY = lucifer yellow, [3H]-D-M = [3H]-D-mannitol, TMER = transmonolayer electrical resistance.
55A.R. Calabro et al. / Journal of Pharmacological and Toxicological Methods 63 (2011) 47–58
cells (Konsoula & Barile, 2005) and mouse embryonic stem cells(Calabro et al., 2008); 2. with only one exception (arsenic), percent ofnewly synthesized proteins, synthesis of total protein and β-actinprotein (Fig. 8), and cell counts, are dramatically decreased from theTMER IC50 and from MTT values. This indicates that concentrations ofmetals needed to reduce all protein synthesis, including proteinsinvolved in cell adhesion, are significantly lower than those requiredfor both PP and cell viability. Gene expression for β-actin mRNA,however, is increased several fold. Interestingly, since total proteinsynthesis and cell counts are lowered at the same rate, the amounts ofprotein synthesized by remaining viable cells are identical (approx-imately 18 to 28 ug per 104 cells). The data for β-actin synthesis is alsographically presented in Fig. 8.
4. Discussion
Preliminary concentration-effect studies were performed for eachmetal, to determine the concentration at which paracellular perme-ability (PP), as measured by transmembrane electrical resistance(TMER), would be inhibited by 50% (IC50 TMER). This concentrationwas then used as a benchmark for subsequent concentration-effectstudies to assess the consequence of metals on cell viability, asmeasured by MTT absorbance, and net protein production, asmeasured by incorporation of isotope into acid-insoluble precipitate.Results of the MTT assay and the labeling experiments with 3H-glycine and 3H-tyrosine (Figs. 1 through 3) clearly demonstrate thatthe concentration-effect curves for the two isotopes are identical; i.e.
with the exception of As, the concentrations used to decrease TMER by50% for each metal significantly inhibited incorporation of isotopesinto newly synthesized proteins to less than 5% of control. This allowsus to safely conclude that, at equivalent concentrations of metals, netprotein production is significantly affected before PP is disturbed.
Glycine and tyrosine are both non-essential amino acids and, likeother amino acids in mammalian cells, these intracellular constituentsare derived from glycolytic side reactions. Their biochemical similar-ities, however, diverge here. For instance, their participation inglycolytic reactions differ significantly—serine is a major source ofglycine via a reversible reaction catalyzed by serine hydroxymethyl-transferase, while tyrosine is produced from the essential amino acidphenylalanine in a single step catalyzed by phenylalanine hydroxy-lase. Also, glycine is more highly represented in β-actin (10%) thantyrosine (5%), and is more frequently found in all proteins (7.2%) thantyrosine (3.2%)[only leucine and alanine are more frequently detectedin proteins than glycine, 9.0% and 8.3%, respectively] (Hennessey,Drummond, & Sparrow, 1993; Nakajima-Iijima, Hameda, Reddy, &Kakunaga, 1985). Glycine and tyrosine have different energy require-ments for hydropathy—i.e. the tendency to prefer a hydrophobicenvironment (0.67 kj/mol vs. 0.08 kj/mol, respectively). Moreover,the amino acids have different energy requirements for biosynthesis—glycine requires 12 mol of ATP for biosynthesis whereas tyrosinerequires 62 mol of ATP. Finally, the pKa values for the α-amino groupsfor glycine and tyrosine differ significantly at 25 °C (9.8 vs. 9.2,respectively), considering that the range of pKa values for all 20 freeamino acids are narrow (from 8.7 for asparagine, to 10.7 for cysteine)

Fig. 7. Effect of: A. As 48-h, B. As 96-h, C. Hg 48-h, and D. Hg 96-h, on paracellular transport in Caco-2 cells. Concentrations correspond to IC50s generated with either theMTT or TMERstudies. Scale for Percent of Control Response for PP markers (bars) are on left axis; for TMER measurements (dashed–dotted line), scale is on the right axis. Control values are set at100% (bars) or based on 1000 Ω-cm2 (for TMER). LY = lucifer yellow, [3H]-D-M = [3H]-D-mannitol, TMER = transmonolayer electrical resistance.
56 A.R. Calabro et al. / Journal of Pharmacological and Toxicological Methods 63 (2011) 47–58
(Horton, Moran, Ochs, Rawn, and Scrimgeour, 2005). Thus, suchdifferences in biochemical features between the amino acids wouldpredict different results for any set of parallel studies, particularlyconcerning incorporation into nascent proteins. The close agreementbetween the results with radioactive glycine and tyrosine, however,
Table 3Summary of effects of 24-h single dose exposures to 6 metals on Caco-2 cells.
Groups±metals IC50 TMER(mM)
MTT GLY TYR β-amR
Control group 0 100±1.2 100±2.3 100±2.0 1
Cadmium chloride 0.05 55.6±4.2 2.4±1.1 3.0±9.5 107
Nickel chloride 8.4 16.0±6.5 1.4±3.7 2.5±2.7 159
Cupric sulfate 1.8 44.0±5.0 1.8±3.1 0.2±1.5 110
Manganese chloride 3.1 43.7±1.6 1.8±2.3 1.9±1.1 250
Mercury chloride 0.09 45.0±3.4 2.7±5.1 1.3±2.4 307
Arsenic trioxide 0.13 47.8±8.9 31.3 ±5.1 85.1±0.8 177
All values are expressed as percent of control group in the absence or presence of metal, excone concentration, i.e. the IC50 concentration determined for TMER in paracellular permeabil(paired 2-sample for means assuming equal variances). Statistical analysis reveals that all vaper 104 cells.Abbreviations: GLY = [3H]glycine incorporation into acid-insoluble precipitate; IS = intern
a Represents ratio of β-actin mRNA expression in the presence or absence of metal, as cob Each value represents mean of separate experiments, 6 replications per data point/expc From BCA assay for total proteins.
with the exception of arsenic, indicates that the decreases in amountsof label incorporated into acid-insoluble material are most likely dueto decreases in net protein production, rather than to changes in poolsize or availability (uptake). Moreover, monitoring the quick turnoverof proteins in the presence of metals is best determined by the
ctinNAa
β-actin proteinproductionb
Cellcountb
Total proteinsynthesisb,c
µg totalprotein/104 cellsb
00±5.2 100±2.8100±1.6
100±3.8100±2.0
100±3.0100±2.1
23.5±3.221.6±2.1
.8±1.0 11.5±0.24.3±0.2
40.1±0.840.6±2.7
40.0±1.642.2±1.3
22.4 ±1.025.2±2.1
.8±1.7 5.4±0.32.8±0.2
33.9±2.130.4±0.9
37.9±0.526.6±1.1
22.4±1.223.7±1.0
.9±2.0 10.4±0.315.7±0.2
37.2±1.835.5±1.3
49.5±0.547.6±0.4
28.1±1.324.7±0.9
.0±3.9 6.5±0.213.9±0.3
26.1±1.328.4±1.8
23.8±2.834.9±0.3
21.7±2.018.4±1.1
.3±4.1 8.5±0.28.9±0.2
42.2±2.046.9±1.9
34.1±1.030.1±2.3
20.3±1.522.1±2.1
.4±3.3 25.9±0.923.1±0.5
56.3±1.962.3±1.4
59.8±4.565.8±1.1
25.9±3.223.3±1.3
ept for IC50 TMER concentrations (mM, column 2). Values represent effect measured atity assays. All groups were analyzed for one-way ANOVA (pb0.001) and Student's t-testlues are significantly different from control groups (pb0.001) except for µg total protein
al standard; TYR = [3H]tyrosine incorporation into acid-insoluble precipitate.mpared to IS.eriment.

Fig. 8. Effect of metals on confluent monolayers of Caco-2 cells exposed to 2concentrations for 24-h. Cells were harvested and β-actin synthesis was quantified asdescribed in Materials and methods. Concentrations are based on IC50 and 10−2×IC50
for each metal as previously determined with the TMER assays (Tables 2 and 3). Resultsare expressed as percent of the control cells (no metal exposure). Each bar representsthe mean±SE of six samples; experiments were repeated twice. *=significantlydifferent from control group, Pb0.05.
57A.R. Calabro et al. / Journal of Pharmacological and Toxicological Methods 63 (2011) 47–58
percentage change between experimental and control groups, butmay also involve the minor contribution of protein degradation overthe 24-h time span, the data for which is not pertinent for the scope ofthis study.
In our studies, incorporation of glycine and tyrosine in cell culturesreflects a decrease of newly synthesized proteins at concentrations ofmetals significantly lower than those affecting TMER and cell viability.Amino acid incorporation into acid-insoluble material was reduced bygreater than 90% from controls at concentrations equivalent to thecalculated TMER IC50 concentrations. Total protein synthesis, usingthe BCA assay (Table 3), did not decrease as dramatically as with theradio-labeling experiments, but in fact, the former paralleled the cellcount data. Consequently, micrograms of total protein produced perunit cell did not change from control at the IC50 concentrations. Totalsynthesis for β-actin protein, however, decreased significantly, at arate lower than total protein production, and more closely mimics theradioactive experiments. Interestingly, gene expression for β-actinmRNA, however, is increased several fold. We conclude, therefore,that the metals at concentrations that affect PP are not specific fortotal proteins but appear to target TJ proteins such as β-actin, andsimultaneously trigger mRNA synthesis for the latter.
This data confirms our previous results with Caco-2 cells(Konsoula & Barile, 2005) and mouse embryonic stem cells (Calabroet al., 2008) that cell viability decreases before PP is compromised.More importantly, nascent protein production, total protein produc-tion at confluence, and specifically, β-actin protein, are affected bymetal exposure before TJ integrity or cell viability are compromised.This also implies that concentrations of metals needed to reduce allprotein synthesis, including proteins involved in cell adhesion, aresignificantly lower than those required for both PP and cell viability.Subsequent data is necessary to prove that there is a disconnectionbetween transcription and post-translational modification of TJproteins, where in fact there may be an upstream signaling pathwaystimulated by the alteration in cell adhesion molecules.
Related studies have demonstrated enhanced cellular defensemechanisms in the presence of metal exposure. For instance, Bae,Gennings, Carter, Yang and Campain (2001) reported that cytotoxicityswitches from synergistic to antagonistic when keratinocytes areexposed to increasing concentrations of metal mixtures, particularlywith expression of metallothioneins. In an attempt to understand thecomplex biological effects of cadmium, Koizumi and Yamada (2003)show that cellular protection and anti-oxidant genes, such as thoseassociated with metallothioneins and heat shock proteins, wereinduced in the presence of non-lethal Cd toxicity. Bertin and Averbeck(2006) review the suppression of protein production and cellularproliferation in mammalian cells with Cd, while modulatingcorresponding gene expression and signal transduction. Recentstudies indicate that cell adhesion molecules associated with TJs,such as occludin and claudins, are primary targets for metal-inducedrenal injury (Prozialeck & Edwards, 2007). Furthermore, Pb and Hgdisrupt heme synthesis and microtubules, respectively, the latter ofwhich interfereswith developmental processes (Clarkson, 1987). And,metals, such as As, Cd, Hg, and Pb, induce oxidative stress conditions,triggering cellular anti-oxidant mechanisms in an attempt to abatemetal toxicity (Ercal, Gurer-Orhan, & Aykin-Burns, 2001).
Although the Caco-2 cell model has historically been instrumentalfor absorption studies, limited reports have investigated the relation-ship between in vitro paracellular permeability and cytotoxicity. Forinstance, Laparra, Velez, Barbera, Montoro, and Farre (2005) haveshown that inorganic As (As[III]) alters TJ permeability in Caco-2 cells.Chao, Taylor, Daddona, Broughall, and Fix (1998) reported that long-chain acylcarnitines increase intestinal absorption of small hydrophiliccompounds through the paracellular pathway by dilating paracellularspaces in Caco-2monolayers. By calculating the ratio between IC50 andEC50 values, Liu, LeCluyse, and Thakker (1999) developed a potencyindex for novel absorption enhancers, which can be used to predict theeffective concentration at which transepithelial electrical resistance(TEER) drops to 50%. Duizer et al. (1999) detected a decrease in TEERwith 5- to 30-µM CdCl2 exposure in Caco-2 cells, with a correspondingdisruption of the paracellular barrier, yet without a significant loss ofviability. Boveri et al., (2004) report that CdCl2 toxicity is detected inCaco-2 cells at very low concentrations using LDH release and HSP70as toxicological endpoints. Okada, Narai, Matsunaga, Fusetani, andShimizu (2000) compared changes in TEER in Caco-2 cell monolayerswith cell viability for marine toxins using the LDH release assay. Theydetected that a decrease in TEER, associated with increased transe-pithelial permeability, preceded LDH release.
Thus, the current study follows our initial findings that investigatedthe relationship between PP and cytotoxicity in Caco-2 and othermammalian cells (Konsoula & Barile, 2005; Schmidt, Cheng, Marino,Konsoula, & Barile, 2004). We report that the decreased epithelialbarrier function of intestinal epithelial cells is accompanied by TJdisruption due, in part, to specific alterations in β-actin synthesis, andthat passive paracellular transport of TJ markers correlates well withIC50s determined with the TMER measurements. Since the concentra-tions ofmetals required to decrease protein synthesiswere significantlylower than IC50s for MTT and PP measurements, and the β-actin genewas up-regulated for the same concentrations, we conclude thatsynthesis andmRNAexpression forβ-actin is altered before cell viabilityand TJ integrity are compromised. The mechanism suggests that metaltoxicity independently alters transcriptional and post-translationalpathways over short periods of time and at relatively low concentra-tions, and may account for initiation or promotion of the carcinogenicpathway. Thus the study furthers our understanding of how metalscontribute to the carcinogenic process by interfering with TJ integrity.
Acknowledgements
This work was supported in part by grants from the InternationalFoundation for Ethical Research (IFER, Chicago, IL, USA) and the

58 A.R. Calabro et al. / Journal of Pharmacological and Toxicological Methods 63 (2011) 47–58
Alternatives Research & Development Foundation (ARDF, Jenkintown,PA, USA). Parts of this study were presented at the annual meetings ofthe Society for In Vitro Biology, June 2007, and the U.S. Society ofToxicology, March 2008 and 2009. The authors wish to thank Mr.Sushil Pai for technical assistance with the MTT assays.
References
Bae, D. S., Gennings, C., Carter, W., Yang, R. H. S., & Campain, J. A. (2001). Toxicologicalinteractions among arsenic, cadmium, chromium, and lead in human keratinocytes.Toxicological Sciences, 63, 132−142.
Balshaw, S., Edwards, J., Daughtry, B., & Ross, K. (2007). Mercury in seafood:Mechanisms of accumulation and consequences for consumer health. Reviews onEnvironmental Health, 22, 91−113.
Barile, F. A., & Cardona, M. (1998). Acute cytotoxicity testing with cultured human lungand dermal cells. In Vitro Cellular & Developmental Biology, 34, 631−635.
Barnes, Y., Houser, S., & Barile, F. A. (1990). Temporal effects of ethanol on growth,thymidine uptake, protein and collagen production in human foetal lungfibroblasts.Toxicology in Vitro, 4, 1−7.
Bertin, G., & Averbeck, D. (2006). Cadmium: Cellular effects, modifications ofbiomolecules, modulation of DNA repair and genotoxic consequences. Biochimie,88, 1549−1559.
Boveri, M., Pazos, P., Gennari, A., Casado, J., Hartung, T., & Prieto, P. (2004). Comparisonof the sensitivity of different toxicological endpoints in Caco-2 cells after cadmiumchloride treatment. Archives of Toxicology, 78, 201−206.
Calabro, A. R., Konsoula, R., & Barile, F. A. (2008). Evaluation of in vitro cytotoxicity andparacellular permeability of intact monolayers with mouse embryonic stem cells.Toxicology in Vitro, 22, 1273−1284.
Chao, A. C., Taylor,M. T., Daddona, P. E., Broughall,M., & Fix, J. A. (1998).Molecularweight-dependent paracellular transport of fluorescent model compounds induced bypalmitoylcarnitine chloride across the human intestinal epithelial cell line Caco-2.Journal of Drug Targeting, 6, 37−43.
Clarkson, T. W. (1987). Metal toxicity in the central nervous system. EnvironmentalHealth Perspectives, 75, 59−64.
Duizer, E., Gilde, A. J., Versantvourt, C. H., & Groton, J. P. (1999). Effects of cadmiumchloride on the paracellular barrier function of intestinal epithelial cell lines.Toxicology and Applied Pharmacology, 155, 117−126.
Ercal, N., Gurer-Orhan, H., & Aykin-Burns, N. (2001). Toxic metals and oxidative stresspart I: Mechanisms involved in metal-induced oxidative damage. Current Topics inMedicinal Chemistry, 1, 529−539.
Fanning, A. S., Jameson, B. J., Jesaitis, A., & Anderson, J. M. (1998). The tight junctionprotein ZO-1 establishes a link between the transmembrane protein occludin andthe actin cytoskeleton. The Journal of Biological Chemistry, 45, 29745−29753.
Ferruzza, S., Scacchi, M., Scarino, M. L., & Sambuy, Y. (2002). Iron and copper alter tightjunction permeability in human intestinal Caco-2 cells by distinct mechanisms.Toxicology in Vitro, 16, 399−404.
Gerber, G. B., Léonard, A., & Hantson, P. (2002). Carcinogenicity, mutagenicity andteratogenicity of manganese compounds. Critical Reviews in Oncology/Hematology,42, 25−34.
Gonzalez-Mariscal, L., Betanzos, P., Nava, P., & Jaramillo, B. E. (2003). Tight junctionproteins. Progress in Biophysics and Molecular Biology, 81, 1−44.
Gupta, N., Miyauchi, S., Martindale, R. G., Herdman, A. V., Podolsky, R., Miyake, K., et al.(2005). Up-regulation of the amino acid transporter ATB0,+(SLC6A14) in colorectalcancer and metastasis in humans. Biochimica et Biophysica Acta, 1741, 215−223.
Hennessey, E., Drummond, D., & Sparrow, J. (1993). Molecular genetics of actinfunction. The Biochemical Journal, 282, 657−671.
Horton, H. R., Moran, L. A., Ochs, R. S., Rawn, J. D., & Scrimgeour, K. G. (2005). Chapter 17,Amino acidmetabolism, in: Principles of biochemistry, 3rd editionNJ, USA: Prentice Hall.
Huang, S. C., Huang, C. -Y. F., & Lee, T. C. (2000). Induction ofmitosis-mediated apoptosis bysodium arsenite in HeLa S3 cells. Biochemical Pharmacology, 60, 771−780.
Jarup, L. (2003). Hazards of heavymetal contamination. BritishMedical Bulletin, 68, 167−182.Karaczyn, A. A., Golebiowski, F., & Kasprzak, K. S. (2005). Truncation, deamidation, and
oxidation of histone H2B in cells cultured with nickel (II). Chemical Research inToxicology, 18, 1934−1942.
Kasprzak, K. S., Sunderman, F. W., & Salnikow, K. (2003). Nickel carcinogenesis.Mutation Research//Fund Mol. Mech. Muta., 533, 67−97.
Kebache, S., Zuo, D., Chevet, E., & Larose, L. (2002). Modulation of protein translation byNck-1. Proceedings of the National Academy of Sciences of the United States ofAmerica, 99, 5406−5411.
Konsoula, R., & Barile, F. A. (2005). Correlation of in vitro cytotoxicity with paracellularpermeability in Caco-2 cells. Toxicology in Vitro, 19, 675−684.
Konsoula, R., & Barile, F. A. (2007). Correlation of in vitro cytotoxicity with paracellularpermeability in mortal rat intestinal cells. Journal of Pharmacological andToxicological Methods, 55, 176−183.
Koizumi, S., & Yamada, H. (2003). DNA microarray analysis of altered gene expressionin cadmium exposed human cells. Journal of Occupational Health, 45, 331−334.
Laparra, J. M., Velez, D., Barbera, R., Montoro, R., & Farre, R. (2005). An approach to As(III)andAs(V) bioavailability studieswithCaco-2 cells. Toxicology inVitro, 19, 1071−1078.
Lee, Y. W., Klein, C. B., Kargacin, B., Salnikow, K., Kitahara, J., Dowjat, K., et al. (1995).Carcinogenic nickel silences gene expression by chromatin condensation and DNAmethylation: A newmodel for epigenetic carcinogens.Molecular and Cellular Biology,15, 2547−2557.
Liu, D. Z., LeCluyse, E. L., & Thakker, D. R. (1999). Dodecylphosphocholine-mediatedenhancement of paracellular permeability and cytotoxicity in Caco-2 cell mono-layers. Journal of Pharmaceutical Sciences, 88, 1161−1168.
Nakajima-Iijima, S., Hamada, H., Reddy, P., & Kakunaga, T. (1985). Molecular structure ofthe human cytoplasmic β-actin gene: Interspecies homology of sequences in theintrons. Proceedings of the National Academy of Sciences of the United States of America,82, 6133−6137.
Natoli, M., Felsani, A., Ferruzza, S., Sambuy, Y., Canali, R., & Scarino, M. L. (2009).Mechanisms of defence from Fe(II) toxicity in human intestinal Caco-2 cells.Toxicology in Vitro, 23, 1510−1515.
Navarro-Moreno, L. G., Quintanar-Escorza, M. A., González, S., Mondragón, R., Cerbón-Solorzáno, J., Valdés, J., & Calderón-Salinas, J. V. (2009). Effects of lead intoxicationon intercellular junctions and biochemical alterations of the renal proximal tubulecells. Toxicology in Vitro, 23, 1298−1304.
Okada, T., Narai, A., Matsunaga, S., Fusetani, N., & Shimizu, M. (2000). Assessment of themarine toxins by monitoring the integrity of human intestinal Caco-2 cellmonolayers. Toxicology in Vitro, 14, 219−226.
Prozialeck, W. C., & Edwards, J. R. (2007). Cell adhesion molecules in chemically-induced renal injury. Pharmacology & Therapeutics, 114, 74−93.
Prozialeck, W. C., Edwards, J. R., Lamar, P. C., & Smith, C. S. (2006). Epithelial barriercharacteristics and expression of cell adhesion molecules in proximal tubule-derived cell lines commonly used for in vitro toxicity studies. Toxicology in Vitro, 20,942−953.
Rossi, A., Poverini, R., Di Lullo, G., Modesti, A., Modica, A., & Scarino, M. L. (1996). Heavymetal toxicity following apical and basolateral exposure in the human intestinalcell line caco-2. Toxicology in Vitro, 10, 27−36.
Salnikow, K., & Zhitovich, A. (2008). Genetic and epigenetic mechanisms in metalcarcinogenesis and cocarcinogenesis: Nickel, arsenic and chromium. ChemicalResearch in Toxicology, 21, 28−44.
Schmidt, C. M., Cheng, C. N., Marino, A., Konsoula, R., & Barile, F. A. (2004). Hormesiseffect of trace metals on cultured normal and immortal human mammary cells.Toxicology and Industrial Health, 20, 1070−1082.
Tchounwou, P. B., Centeno, J. A., & Patlolla, A. K. (2004). Arsenic toxicity, mutagenesis,and carcinogenesis—A health risk assessment and management approach.Molecular and Cellular Biochemistry, 255, 47−55.
Trosko, J. E. (2007). Gap junctional intercellular communication as a biological “RosettaStone” in understanding, in a systems biological manner, stem cell behavior,mechanisms of epigenetic toxicology, chemoprevention and chemotherapy. TheJournal of Membrane Biology, 218, 93−100.
Turski, M. L., & Thiele, D. J. (2009). New roles for coppermetabolism in cell proliferation,signaling, and disease. The Journal of Biological Chemistry, 284, 717−721.
Waalkes, M. (2000). Cadmium carcinogenesis in review. Journal of Inorganic Biochemistry,79, 241−244.
Washer, L. P., Law, D., Boland, C. R., Horon, I., & Feinberg, A. (1996). Limited up-regulation of DNA methyltransferase in human colon cancer reflecting increasedcell proliferation. Proceedings of the National Academy of Sciences of the United Statesof America, 93, 10366−10370.
Zhang, Y., Li, M., Yao, Q., & Chen, C. (2009). Roles and mechanisms of coppertransporting ATPases in cancer pathogenesis. Medical Science Monitor, 15, 1−5.