ectoparasite fitness in auxiliary hosts: phylogenetic distance from a principal host matters
TRANSCRIPT

Ectoparasite fitness in auxiliary hosts: phylogenetic distancefrom a principal host matters
I . S . KHOKHLOVA* , L . J . F IELDEN† , A . A . DEGEN* & B. R. KRASNOV‡*Wyler Department of Dryland Agriculture, French Associates Institute for Agriculture and Biotechnology of Drylands, Jacob Blaustein Institutes for Desert Research,
Ben-Gurion University of the Negev, Midreshet Ben-Gurion, Israel
†School of Science and Math, Truman State University, Kirksville, MO, USA
‡Mitrani Department of Desert Ecology, Swiss Institute for Dryland Environmental and Energy Research, Jacob Blaustein Institutes for Desert Research, Ben-Gurion
University of the Negev, Midreshet Ben-Gurion, Israel
Keywords:
auxiliary hosts;
fitness;
fleas;
principal host;
rodents.
Abstract
We studied reproductive performance in two flea species (Parapulex chephre-
nis and Xenopsylla ramesis) exploiting either a principal or one of eight auxil-
iary host species. We predicted that fleas would produce more eggs and
adult offspring when exploiting (i) a principal host than an auxiliary host
and (ii) an auxiliary host phylogenetically close to a principal host than an
auxiliary host phylogenetically distant from a principal host. In both flea
species, egg production per female after one feeding and production of new
imago after a timed period of an uninterrupted stay on a host differed signif-
icantly between host species. In general, egg and/or new imago production
in fleas feeding on an auxiliary host was lower than in fleas feeding on the
principal host, except for the auxiliary host that was the closest relative of
the principal host. When all auxiliary host species were considered, we did
not find any significant relationship between either egg or new imago pro-
duction in fleas exploiting an auxiliary host and phylogenetic distance
between this host and the principal host. However, when the analyses were
restricted to auxiliary hosts belonging to the same family as the principal
host (Muridae), new imago production (for P. chephrenis) or both egg and
new imago production (for X. ramesis) in an auxiliary host decreased signifi-
cantly with an increase in phylogenetic distance between the auxiliary and
principal host. Our results demonstrated that a parasite achieves higher fit-
ness in auxiliary hosts that are either the most closely related to or the most
distant from its principal host. This may affect host associations of a parasite
invading new areas.
Introduction
One of the main mechanisms to maximize reproductive
success of an individual is selection of habitats that pro-
vide the greatest fitness output (Rosenzweig, 1981,
1987, 1991). Selection of a habitat with higher per cap-
ita resource abundance and/or that allows easier
resource acquisition presumably results in higher fitness
reward. One of the consequences of fitness increase is
an increased abundance, so that the abundance of a
consumer in a habitat is often considered a measure of
its performance and efficiency of resource exploitation
in this habitat (Morris, 1987).
Long before the theory of habitat selection was for-
mulated, scientists who studied parasitic species imple-
mented similar ideas for the explanation of unequal
distribution of conspecific parasites among different
host species. Traditionally, a host in which a parasite
attains the highest abundance and/or prevalence is con-
sidered to be its principal host, whereas other hosts
exploited by the parasite are considered to be auxiliary
hosts (Dogiel et al., 1961; Dogiel, 1964). It has been
Correspondence: Boris R. Krasnov, Mitrani Department of Desert Ecology,
Swiss Institute for Dryland Environmental and Energy Research, Jacob
Blaustein Institutes for Desert Research, Ben-Gurion University of the
Negev, Sede-Boqer Campus, 84990 Midreshet Ben-Gurion, Israel.
Tel.: +972 8 6596841; fax: +972 8 6596772 e-mail: [email protected]
ª 2 01 2 THE AUTHORS . J . E VOL . B I OL .
1JOURNAL OF EVOLUT IONARY B IO LOGY ª 20 1 2 EUROPEAN SOC I E TY FOR EVOLUT IONARY B IO LOGY
doi: 10.1111/j.1420-9101.2012.02577.x

shown that, in at least some parasites, exploitation of
the principal versus the auxiliary host has resulted in
different fitness rewards with the rewards being highest
with the principal host (Krasnov et al., 2002).
Nevertheless, abundance and/or prevalence of a para-
site varies substantially among auxiliary hosts. To
account for this variation, parasitologists classify an
auxiliary host as being a ‘normal’, ‘secondary’, ‘acci-
dental’ or ‘exceptional’ host for a parasite (Hopkins,
1949; Holland, 1964; Wenzel & Tipton, 1966; Marshall,
1981). Mechanisms for the variation in abundance of a
parasite among auxiliary host are poorly understood.
Poulin (2005) proposed a hypothesis that the ultimate
cause for the variation in parasite abundance among
auxiliary hosts is the difference among these hosts in
their phylogenetic relatedness to the principal host. The
rationale underlying this hypothesis is that phyloge-
netic relatedness among species should mirror their
physiological and ecological similarities (e.g. Harvey &
Pagel, 1991). If so, then an increase in phylogenetic dis-
tance between an auxiliary host and a principal host
should correlate negatively with parasite abundance in
the auxiliary host. Such a negative correlation was
reported for fleas (Insecta: Siphonaptera) parasitic on
small Holarctic mammals (Krasnov et al., 2004a); how-
ever, Poulin (2005) did not find any support for this
prediction for metazoan parasites of freshwater fish.
In an attempt to find proximate mechanisms for the
decrease in flea abundance in auxiliary hosts with an
increase in phylogenetic distance from the principal
hosts, Khokhlova et al. (2012a,b) investigated feeding
performance of two flea species (Xenopsylla ramesis and
Parapulex chephrenis) when they exploited their princi-
pal hosts and auxiliary hosts with varying phylogenetic
relatedness to the principal hosts. Feeding performance
of fleas was measured by bloodmeal size and energy
expenditure for blood digestion. Unexpectedly, it was
found that fleas did not always perform better on a
principal host than on an auxiliary host and that, in
some cases, fleas fed better on hosts that were phyloge-
netically distant from than close to their principal host.
One of the reasons behind the contradiction between
these results and the pattern of flea abundances in aux-
iliary hosts with different degrees of relatedness to the
principal host is that variation in feeding performance
(including energy cost of bloodmeal processing) among
hosts might not necessarily be a good proxy for fitness
achieved in these hosts. Nevertheless, haematological
properties vary between vertebrate species (Harrington
et al., 2001) and therefore could affect the fitness of
haematophagous insects via their effect on the nutritive
and/or energy value of blood (Harrington et al., 2001;
Lyimo & Ferguson, 2009; Lyimo et al., 2012). Conse-
quently, identifying the proximate mechanism that
explains variation in parasite abundance among auxil-
iary hosts as a function of phylogenetic distances
between these hosts and the principal host requires
experimental measurement of direct fitness-related
variables that would measure the ability of a parasite to
translate resources extracted from a host into the
parasite’s offspring.
Here, we addressed the question of the effect of phy-
logenetic distance between an auxiliary host and princi-
pal host on reproductive performance of fleas in this
auxiliary host. To answer this question, we measured
fitness via production of eggs and new adults in two
flea species (Parapulex chephrenis and Xenopsylla ramesis)
after they exploited either a principal or one of eight
auxiliary host species. We predicted that fleas would
produce more eggs and more new adults when exploit-
ing (i) a principal host than an auxiliary host and (ii)
an auxiliary host phylogenetically close to a principal
host than one phylogenetically distant from a principal
host. The two flea species used in this study are com-
mon in desert habitats of southern Israel. Parapulex che-
phrenis is parasitic mainly on the Egyptian spiny mouse
Acomys cahirinus and is found less often on the golden
spiny mouse Acomys russatus and the gerbils Meriones
crassus and Gerbillus dasyurus (Krasnov et al., 1997,
1999). Xenopsylla ramesis is found on a variety of gerbil-
line rodents, but attains the highest abundance and
prevalence on M. crassus (Krasnov et al., 1997, 1999).
Materials and methods
Fleas and rodents
We used fleas from our laboratory colonies started in
1999 from field-collected specimens. Parapulex chephre-
nis were collected from A. cahirinus, whereas X. ramesis
were collected from Psammomys obesus, M. crassus and
G. dasyurus. Details on breeding and maintenance of
fleas can be found elsewhere (e.g. Krasnov et al., 2002,
2003; Khokhlova et al., 2009a,b, 2010a). In brief, fleas
were maintained on their characteristic rodent hosts
(P. chephrenis on A. cahirinus and A. russatus and X. ra-
mesis on M. crassus and G. dasyurus) that were kept indi-
vidually in plastic cages with a wire mesh floor over a
pan with a mixture of sand and dried bovine blood (lar-
val nutrient medium). Air temperature was maintained
at 25 °C and photoperiod at 12:12 (L:D) h. Every
2 weeks, all substrate and bedding material from each
rodent cage (including nest box and pan) were col-
lected into plastic boxes with perforated lids and trans-
ferred to an incubator (FOC225E; Velp Scientifica srl,
Milano, Italy; 25 °C air temperature and 75% relative
humidity) where the fleas developed. All fleas used in
experiments were selected randomly from colonies.
We used nine rodent species, eight of them from the
family Muridae and one from the family Cricetidae
(Mesocricetus auratus). Among murids, there were five
gerbillines (M. crassus, G. dasyurus, Gerbillus andersoni,
Gerbillus pyramidum and Gerbillus nanus) and three
murines (A. cahirinus, A. russatus and Mus musculus).
ª 2 0 12 THE AUTHORS . J . E VOL . B IO L . do i : 1 0 . 1 1 1 1 / j . 1 4 20 - 9 10 1 . 2 01 2 . 0 25 7 7 . x
JOURNAL OF EVOLUT IONARY B IOLOGY ª 2012 EUROPEAN SOC I E TY FOR EVOLUT IONARY B IO LOGY
2 I . S . KHOKHLOVA ET AL.

Meriones crassus, G. dasyurus, G. nanus and both Acomys
spp. were from laboratory colonies started in 1997–1999 and 2009 (G. nanus). Gerbillus andersoni, G. pyrami-
dum and M. musculus (feral house mice) were captured
in the wild in desert habitats (gerbils) or settlements
(mice) in southern Israel. The rodents had all ectopara-
sites removed using a toothbrush and forceps and then
were maintained in a separate animal room for
2 months prior to experiments. Mesocricetus auratus
(golden hamsters) were available commercially.
Details on rodent maintenance in colonies can be
found elsewhere (Krasnov et al., 2002, 2003; Khokhl-
ova et al., 2009a,b, 2010a). In short, rodents were
maintained in plastic cages (60 9 50 cm and 40 cm
high) with sawdust bedding at 25 °C air temperature
and 12D:12L regime. Initially, each cage contained a
male and a female. Young individuals were moved to a
new cage at 2 months of age to prevent overpopulation
and inbreeding. Millet seeds and alfalfa (Medicago sp.)
were offered daily ad libitum. No drinking water was
available to the rodents as the alfalfa supplied enough
moisture for their needs. Spiny mice (Acomys) were also
offered commercial cat chow once a week. Only male
rodents aged 6–8 months were used in experiments.
Each rodent was used for feeding a single group of fleas
only once.
Experimental procedures
Fleas were randomly selected from the incubated devel-
opment boxes. An individual rodent was placed in a
plastic cage (60 cm by 50 cm by 40 cm) with a floor of
3–5 mm of clean sand covered by a wire mesh (5 mm
by 5 mm). According to a rodent’s size, 20–50 newly
emerged (24- to 48-h-old) female and 10–30 male fleas
(P. chephrenis or X. ramesis) were released into a cage
and allowed to feed for 3 days. Our preliminary observa-
tions demonstrated that fleas start to oviposit no sooner
than on the second day of the stay on a host under these
conditions. Each treatment with each flea species and
each host species was replicated five to nine times.
After 3 days of an uninterrupted stay in a rodent’s
cage, fleas were collected from both the rodent’s body
and cage substrate. To collect fleas from the rodent’s
body, we brushed them out over a white plastic pan
with a toothbrush. The hair of the rodent was brushed
several times until no flea was recovered. We examined
the fleas collected from the rodent’s body and cage sub-
strate under light microscopy (409 magnification) and
counted the number of fleas (males and females sepa-
rately) with and without blood in their midguts. After
collecting fleas, the substrate was removed from each
cage, added to about 0.5 g of larvae nutrient medium
(94% dry bovine blood, 5% millet flour and 1% grin-
ded excrements of the respective host species), placed
in a plastic box (20 cm by 10 cm by 10 cm) and trans-
ferred to an incubator at 25 °C air temperature and
92–95% RH. Relative humidity was regulated in 38 cm
by 23 cm by 13 cm acrylic humidity chambers using
saturated salt solutions. Temperature and humidity
were monitored using Fisherbrand Traceable Humidity/
Temperature Pen with Memory (Fisher Scientific Inter-
national, NJ, USA). Starting on the 35th or 24th day
after the onset of the experiments (minimal duration of
development of P. chephrenis and X. ramesis, respec-
tively, established in our earlier experiments; see Kras-
nov et al., 2001; Khokhlova et al., 2010b), we checked
the boxes daily until all new adults emerged or for
2 weeks after the emergence of the last adult. We
counted newly emerged fleas and calculated offspring
production in each group of fleas per parent female flea
that took a blood meal during the 3-day uninterrupted
stay on a host. Because some fleas were killed by a
rodent during this period, the number of parent females
that produced eggs was estimated as an average
between the number of female fleas placed on a rodent
(i.e. initial number of females) and collected from this
rodent and cage substrate (i.e. final number of females)
after 3 days. Prior to this, the number of unfed females
(i.e. those with empty midguts) was subtracted from
both initial and final numbers of females. Re-analyses
of the data using either initial or final number of
females (with or without subtraction of the number of
unfed fleas) for calculating the offspring production did
not affect the results.
To obtain flea eggs, fleas of each group (i.e. recovered
from the same rodent individual) that took a bloodmeal
were placed in Petri dishes with the bottom covered by
a thin layer of clean sand and small pieces of filter
paper and were then transferred to an incubator
(FOC225E; Velp Scientifica srl, Milano, Italy) at 25 °Cair temperature and 90% RH for 24 h. On the fourth
day of the experiment, we checked the Petri dishes and
filter paper and collected all eggs laid by all females in
a given group. These eggs were not taken into account
in the subsequent analyses. On the fifth day of the
experiment, we placed each individual rodent in a wire
mesh (5 mm by 5 mm) tube (15 cm length and 5 cm
diameter for M. crassus, spiny mice and hamsters or
10 cm in length and 2 cm in diameter for other gerbils
and house mice) that limited movement and did not
allow self-grooming. Tubes with rodents were placed in
individual white plastic baths. Fleas (P. chephrenis or
X. ramesis) previously collected from this same rodent
were then released into the hair of the rodent. After
feeding on a host for 2 (X. ramesis) or 6 (P. chephrenis)
h (time necessary for satiation estimated from our preli-
minary observations), fleas were collected and exam-
ined for blood in the midguts as described above. Fleas
of each group with blood in their midguts were
returned to Petri dishes that contained sand and pieces
of filter paper and transferred to an incubator for 24 h,
after which we again checked the Petri dishes and filter
paper, counted newly laid eggs and calculated the mean
ª 2 01 2 THE AUTHORS . J . E VOL . B I OL . do i : 1 0 . 1 11 1 / j . 1 4 2 0 - 9 1 0 1 . 2 0 1 2 . 0 2 5 77 . x
JOURNAL OF EVOLUT IONARY B IO LOGY ª 20 1 2 EUROPEAN SOC I E TY FOR EVOLUT IONARY B IO LOGY
Ectoparasite fitness in auxiliary hosts 3
number of eggs produced per fed female after one feed-
ing event.
Data analyses
Parapulex chephrenis and X. ramesis differ in their pat-
terns of host exploitation (time required for satiation,
frequency of feedings, time spend on and off the host
body; see Krasnov, 2008). Consequently, we analysed
data separately for each flea species. As mentioned
above, flea fitness was evaluated from the (a) mean
number of eggs produced by a fed female flea during a
day after one feeding event and (b) mean number of
new adults produced per parent female flea during
3 days of an uninterrupted stay on a host. Prior to
analyses, these dependent variables were log + 1-trans-
formed. Bar diagrams represent nontransformed data.
We used one-way ANOVAs to test for the effect of host
species on the fitness-related variables. To compare
mean egg production and mean production of new off-
spring between the principal host and each of the aux-
iliary hosts, we used univariate tests of significance for
planned comparisons. To test for the relationships
between the fitness-related variables of fleas when they
exploited an auxiliary host and the phylogenetic dis-
tance between this host and a principal host of a flea,
we calculated mean values of these variables across all
flea groups fed on the same auxiliary host species and
regressed them against the phylogenetic distance of an
auxiliary host from a principal host (A. cahirinus for
P. chephrenis and M. crassus for X. ramesis). Information
on topology and branch length of a phylogenetic tree
for the nine rodent species was obtained from various
sources (see details and the tree in Khokhlova et al.,
2012a). Phylogenetic distances were calculated from
branch length of a phylogenetic tree using the package
‘ape’ (Paradis et al., 2004) implemented in the R 2.13.0
software environment (R Development Core Team,
2011). As phylogenetic information was included in the
independent variable, no further phylogenetic correc-
tion was necessary.
Results
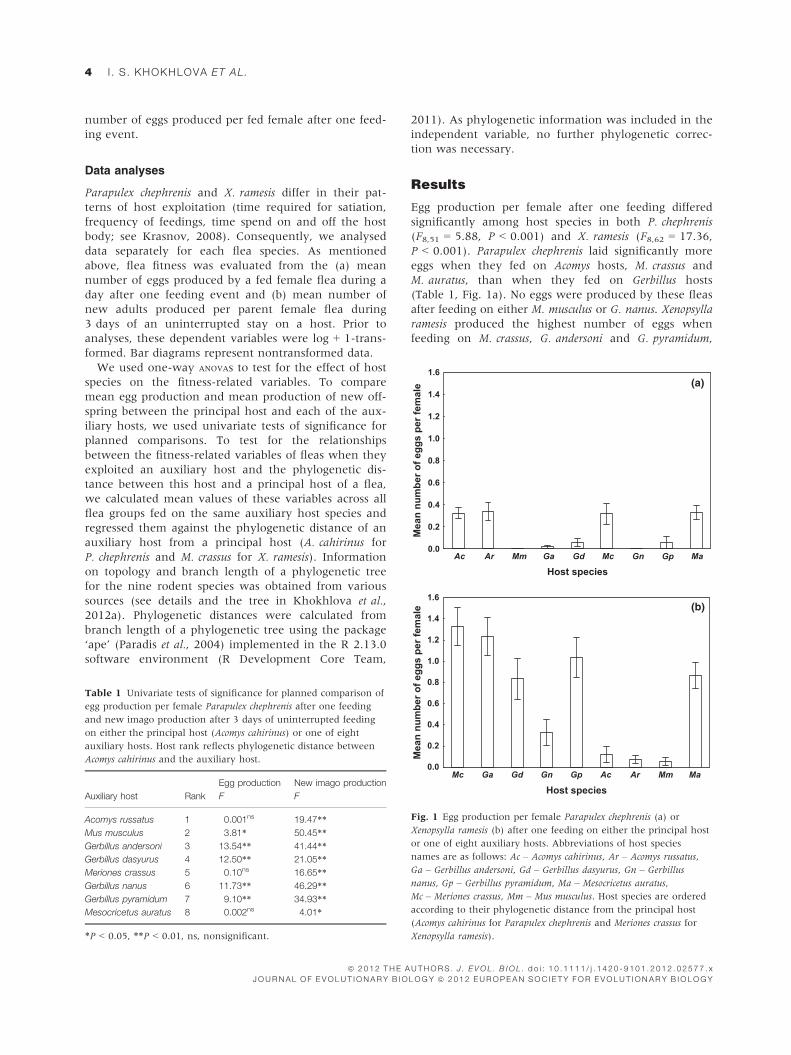
Egg production per female after one feeding differed
significantly among host species in both P. chephrenis
(F8,51 = 5.88, P < 0.001) and X. ramesis (F8,62 = 17.36,
P < 0.001). Parapulex chephrenis laid significantly more
eggs when they fed on Acomys hosts, M. crassus and
M. auratus, than when they fed on Gerbillus hosts
(Table 1, Fig. 1a). No eggs were produced by these fleas
after feeding on either M. musculus or G. nanus. Xenopsylla
ramesis produced the highest number of eggs when
feeding on M. crassus, G. andersoni and G. pyramidum,
Table 1 Univariate tests of significance for planned comparison of
egg production per female Parapulex chephrenis after one feeding
and new imago production after 3 days of uninterrupted feeding
on either the principal host (Acomys cahirinus) or one of eight
auxiliary hosts. Host rank reflects phylogenetic distance between
Acomys cahirinus and the auxiliary host.
Auxiliary host Rank
Egg production
F
New imago production
F
Acomys russatus 1 0.001ns 19.47**
Mus musculus 2 3.81* 50.45**
Gerbillus andersoni 3 13.54** 41.44**
Gerbillus dasyurus 4 12.50** 21.05**
Meriones crassus 5 0.10ns 16.65**
Gerbillus nanus 6 11.73** 46.29**
Gerbillus pyramidum 7 9.10** 34.93**
Mesocricetus auratus 8 0.002ns 4.01*
*P < 0.05, **P < 0.01, ns, nonsignificant.
Host species
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6
Mea
n nu
mbe
r of e
ggs
per f
emal
e (a)
Ac Ar Mm Ga Gd Mc Gn Gp Ma
Mc Ga Gd Gn Gp Ac Ar Mm Ma
Host species
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6
Mea
n nu
mbe
r of e
ggs
per f
emal
e (b)
Fig. 1 Egg production per female Parapulex chephrenis (a) or
Xenopsylla ramesis (b) after one feeding on either the principal host
or one of eight auxiliary hosts. Abbreviations of host species
names are as follows: Ac – Acomys cahirinus, Ar – Acomys russatus,
Ga – Gerbillus andersoni, Gd – Gerbillus dasyurus, Gn – Gerbillus
nanus, Gp – Gerbillus pyramidum, Ma – Mesocricetus auratus,
Mc – Meriones crassus, Mm – Mus musculus. Host species are ordered
according to their phylogenetic distance from the principal host
(Acomys cahirinus for Parapulex chephrenis and Meriones crassus for
Xenopsylla ramesis).
ª 2 0 12 THE AUTHORS . J . E VOL . B IO L . do i : 1 0 . 1 1 1 1 / j . 1 4 20 - 9 10 1 . 2 01 2 . 0 25 7 7 . x
JOURNAL OF EVOLUT IONARY B IOLOGY ª 2012 EUROPEAN SOC I E TY FOR EVOLUT IONARY B IO LOGY
4 I . S . KHOKHLOVA ET AL.
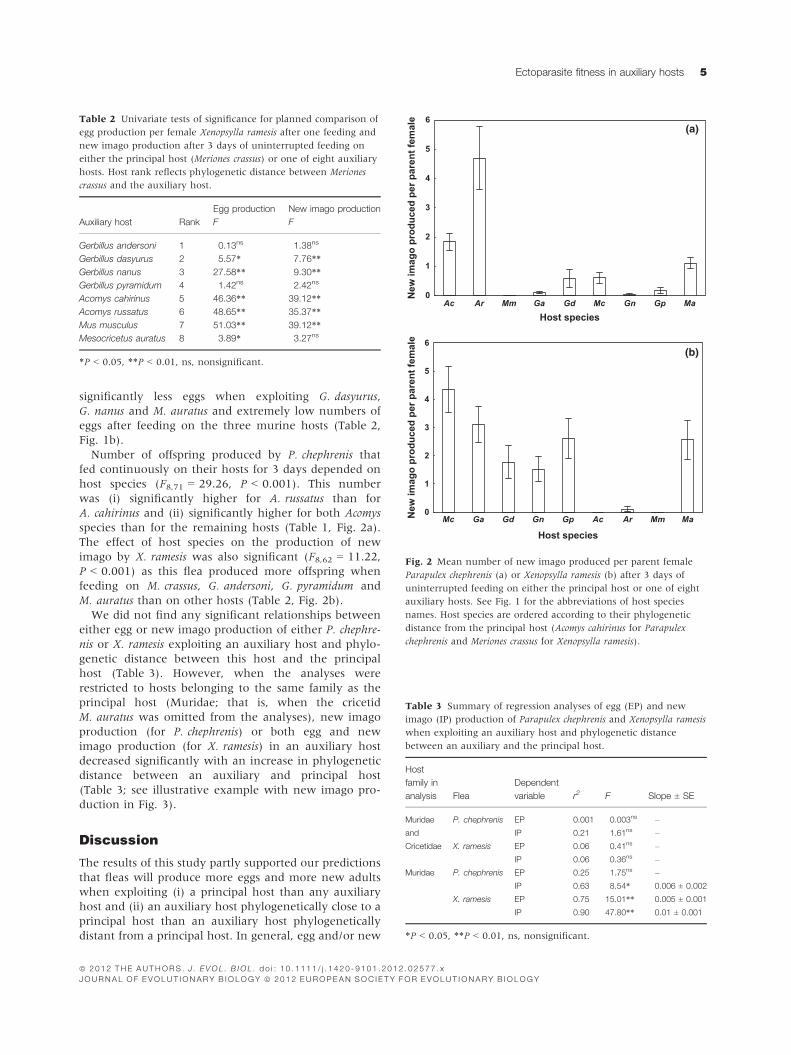
significantly less eggs when exploiting G. dasyurus,
G. nanus and M. auratus and extremely low numbers of
eggs after feeding on the three murine hosts (Table 2,
Fig. 1b).
Number of offspring produced by P. chephrenis that
fed continuously on their hosts for 3 days depended on
host species (F8,71 = 29.26, P < 0.001). This number
was (i) significantly higher for A. russatus than for
A. cahirinus and (ii) significantly higher for both Acomys
species than for the remaining hosts (Table 1, Fig. 2a).
The effect of host species on the production of new
imago by X. ramesis was also significant (F8,62 = 11.22,
P < 0.001) as this flea produced more offspring when
feeding on M. crassus, G. andersoni, G. pyramidum and
M. auratus than on other hosts (Table 2, Fig. 2b).
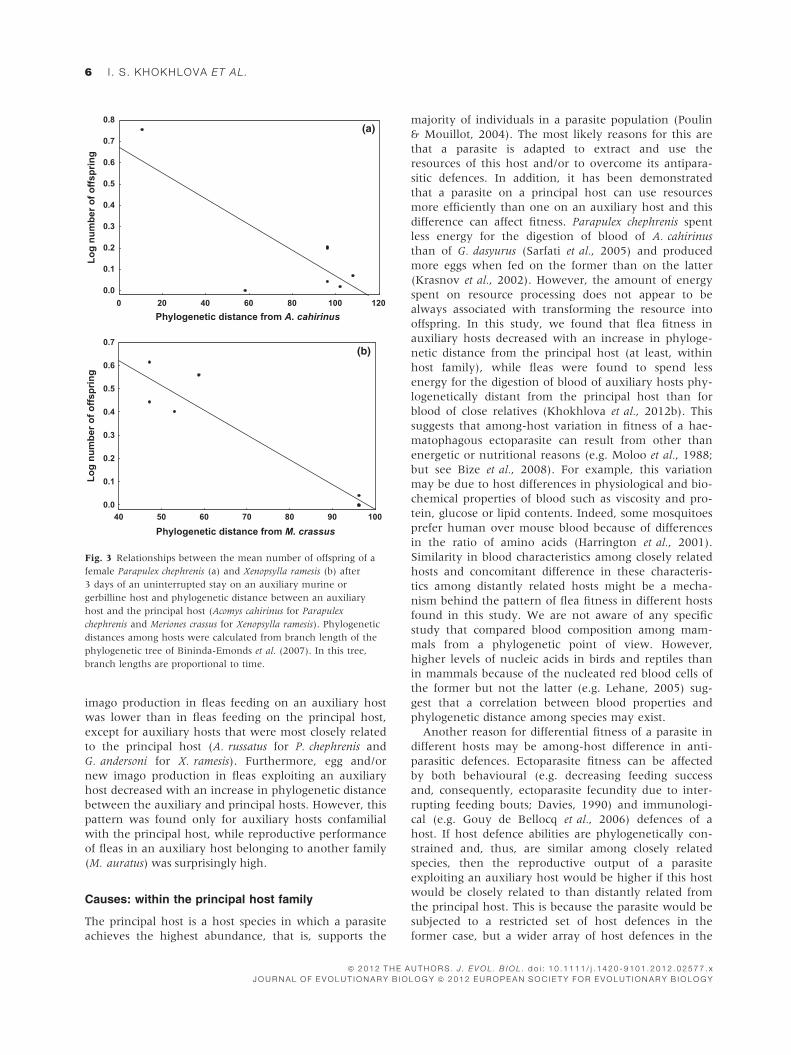
We did not find any significant relationships between
either egg or new imago production of either P. chephre-
nis or X. ramesis exploiting an auxiliary host and phylo-
genetic distance between this host and the principal
host (Table 3). However, when the analyses were
restricted to hosts belonging to the same family as the
principal host (Muridae; that is, when the cricetid
M. auratus was omitted from the analyses), new imago
production (for P. chephrenis) or both egg and new
imago production (for X. ramesis) in an auxiliary host
decreased significantly with an increase in phylogenetic
distance between an auxiliary and principal host
(Table 3; see illustrative example with new imago pro-
duction in Fig. 3).
Discussion
The results of this study partly supported our predictions
that fleas will produce more eggs and more new adults
when exploiting (i) a principal host than any auxiliary
host and (ii) an auxiliary host phylogenetically close to a
principal host than an auxiliary host phylogenetically
distant from a principal host. In general, egg and/or new
Table 2 Univariate tests of significance for planned comparison of
egg production per female Xenopsylla ramesis after one feeding and
new imago production after 3 days of uninterrupted feeding on
either the principal host (Meriones crassus) or one of eight auxiliary
hosts. Host rank reflects phylogenetic distance between Meriones
crassus and the auxiliary host.
Auxiliary host Rank
Egg production
F
New imago production
F
Gerbillus andersoni 1 0.13ns 1.38ns
Gerbillus dasyurus 2 5.57* 7.76**
Gerbillus nanus 3 27.58** 9.30**
Gerbillus pyramidum 4 1.42ns 2.42ns
Acomys cahirinus 5 46.36** 39.12**
Acomys russatus 6 48.65** 35.37**
Mus musculus 7 51.03** 39.12**
Mesocricetus auratus 8 3.89* 3.27ns
*P < 0.05, **P < 0.01, ns, nonsignificant.
Host species
0
1
2
3
4
5
6
New
imag
o pr
oduc
ed p
er p
aren
t fem
ale
(a)
Ac Ar Mm Ga Gd Mc Gn Gp Ma
Mc Ga Gd Gn Gp Ac Ar Mm Ma
Host species
0
1
2
3
4
5
6
New
imag
o pr
oduc
ed p
er p
aren
t fem
ale
(b)
Fig. 2 Mean number of new imago produced per parent female
Parapulex chephrenis (a) or Xenopsylla ramesis (b) after 3 days of
uninterrupted feeding on either the principal host or one of eight
auxiliary hosts. See Fig. 1 for the abbreviations of host species
names. Host species are ordered according to their phylogenetic
distance from the principal host (Acomys cahirinus for Parapulex
chephrenis and Meriones crassus for Xenopsylla ramesis).
Table 3 Summary of regression analyses of egg (EP) and new
imago (IP) production of Parapulex chephrenis and Xenopsylla ramesis
when exploiting an auxiliary host and phylogenetic distance
between an auxiliary and the principal host.
Host
family in
analysis Flea
Dependent
variable r2 F Slope ± SE
Muridae
and
Cricetidae
P. chephrenis EP 0.001 0.003ns –
IP 0.21 1.61ns –
X. ramesis EP 0.06 0.41ns –
IP 0.06 0.36ns –
Muridae P. chephrenis EP 0.25 1.75ns –
IP 0.63 8.54* �0.006 ± 0.002
X. ramesis EP 0.75 15.01** �0.005 ± 0.001
IP 0.90 47.80** �0.01 ± 0.001
*P < 0.05, **P < 0.01, ns, nonsignificant.
ª 2 01 2 THE AUTHORS . J . E VOL . B I OL . do i : 1 0 . 1 11 1 / j . 1 4 2 0 - 9 1 0 1 . 2 0 1 2 . 0 2 5 77 . x
JOURNAL OF EVOLUT IONARY B IO LOGY ª 20 1 2 EUROPEAN SOC I E TY FOR EVOLUT IONARY B IO LOGY
Ectoparasite fitness in auxiliary hosts 5
imago production in fleas feeding on an auxiliary host
was lower than in fleas feeding on the principal host,
except for auxiliary hosts that were most closely related
to the principal host (A. russatus for P. chephrenis and
G. andersoni for X. ramesis). Furthermore, egg and/or
new imago production in fleas exploiting an auxiliary
host decreased with an increase in phylogenetic distance
between the auxiliary and principal hosts. However, this
pattern was found only for auxiliary hosts confamilial
with the principal host, while reproductive performance
of fleas in an auxiliary host belonging to another family
(M. auratus) was surprisingly high.
Causes: within the principal host family
The principal host is a host species in which a parasite
achieves the highest abundance, that is, supports the
majority of individuals in a parasite population (Poulin
& Mouillot, 2004). The most likely reasons for this are
that a parasite is adapted to extract and use the
resources of this host and/or to overcome its antipara-
sitic defences. In addition, it has been demonstrated
that a parasite on a principal host can use resources
more efficiently than one on an auxiliary host and this
difference can affect fitness. Parapulex chephrenis spent
less energy for the digestion of blood of A. cahirinus
than of G. dasyurus (Sarfati et al., 2005) and produced
more eggs when fed on the former than on the latter
(Krasnov et al., 2002). However, the amount of energy
spent on resource processing does not appear to be
always associated with transforming the resource into
offspring. In this study, we found that flea fitness in
auxiliary hosts decreased with an increase in phyloge-
netic distance from the principal host (at least, within
host family), while fleas were found to spend less
energy for the digestion of blood of auxiliary hosts phy-
logenetically distant from the principal host than for
blood of close relatives (Khokhlova et al., 2012b). This
suggests that among-host variation in fitness of a hae-
matophagous ectoparasite can result from other than
energetic or nutritional reasons (e.g. Moloo et al., 1988;
but see Bize et al., 2008). For example, this variation
may be due to host differences in physiological and bio-
chemical properties of blood such as viscosity and pro-
tein, glucose or lipid contents. Indeed, some mosquitoes
prefer human over mouse blood because of differences
in the ratio of amino acids (Harrington et al., 2001).
Similarity in blood characteristics among closely related
hosts and concomitant difference in these characteris-
tics among distantly related hosts might be a mecha-
nism behind the pattern of flea fitness in different hosts
found in this study. We are not aware of any specific
study that compared blood composition among mam-
mals from a phylogenetic point of view. However,
higher levels of nucleic acids in birds and reptiles than
in mammals because of the nucleated red blood cells of
the former but not the latter (e.g. Lehane, 2005) sug-
gest that a correlation between blood properties and
phylogenetic distance among species may exist.
Another reason for differential fitness of a parasite in
different hosts may be among-host difference in anti-
parasitic defences. Ectoparasite fitness can be affected
by both behavioural (e.g. decreasing feeding success
and, consequently, ectoparasite fecundity due to inter-
rupting feeding bouts; Davies, 1990) and immunologi-
cal (e.g. Gouy de Bellocq et al., 2006) defences of a
host. If host defence abilities are phylogenetically con-
strained and, thus, are similar among closely related
species, then the reproductive output of a parasite
exploiting an auxiliary host would be higher if this host
would be closely related to than distantly related from
the principal host. This is because the parasite would be
subjected to a restricted set of host defences in the
former case, but a wider array of host defences in the
Phylogenetic distance from A. cahirinus
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
Log
num
ber o
f offs
prin
g
(a)
0 20 40 60 80 100 120
40 50 60 70 80 90 100
Phylogenetic distance from M. crassus
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
Log
num
ber o
f offs
prin
g
(b)
Fig. 3 Relationships between the mean number of offspring of a
female Parapulex chephrenis (a) and Xenopsylla ramesis (b) after
3 days of an uninterrupted stay on an auxiliary murine or
gerbilline host and phylogenetic distance between an auxiliary
host and the principal host (Acomys cahirinus for Parapulex
chephrenis and Meriones crassus for Xenopsylla ramesis). Phylogenetic
distances among hosts were calculated from branch length of the
phylogenetic tree of Bininda-Emonds et al. (2007). In this tree,
branch lengths are proportional to time.
ª 2 0 12 THE AUTHORS . J . E VOL . B IO L . do i : 1 0 . 1 1 1 1 / j . 1 4 20 - 9 10 1 . 2 01 2 . 0 25 7 7 . x
JOURNAL OF EVOLUT IONARY B IOLOGY ª 2012 EUROPEAN SOC I E TY FOR EVOLUT IONARY B IO LOGY
6 I . S . KHOKHLOVA ET AL.

latter case (Combes, 2001; Poulin & Mouillot, 2004).
However, the existence and strength of phylogenetic
signals (i.e. resemblance among phylogenetically related
species; Blomberg & Garland, 2002) in mammalian
antiparasitic defences remain to be studied.
Causes: outside the principal host family
Unexpectedly, egg and new imago production of both
fleas were high when they fed on a host heterofamilial
with the principal hosts (M. auratus). This could result
from the lack of defences against these fleas in this
host. Natural selection would not favour specific anti-
parasitic defences of a given host if it does not encoun-
ter this parasite or the frequency of parasite exposure is
low (Poulin et al., 1994; Shudo & Iwasa, 2001). Indeed,
results of experimental studies with rodent fleas forced
to feed on their preferred rodent host, a nonpreferred
rodent host or an ‘alien’ bat host, showed that fleas
performed better on a bat host than on the nonpre-
ferred rodent (Krasnov et al., 2007; Korine et al., 2012).
Furthermore, no cricetid host is naturally infested with
either P. chephrenis or X. ramesis. In addition, M. auratus
is a laboratory species, so it has not been attacked by
ectoparasites for many generations and, thus, its
defence tools could be weak. The latter explanation is,
however, weakened by the fact that some of rodents
used in this study were bred in our laboratory over
many generations, so their antiparasitic defences might
be affected similarly to M. auratus.
Evolutionary consequences: geographicaldistribution and host associations
Our study suggests that it may be advantageous for an
ectoparasite to select an auxiliary host which is either
the most closely or the most distantly (within some
realistic phylogenetic distance) related to the principal
host species. Evidence from distributional data of fleas
and their hosts provides support for both of such cases.
For example, among Palearctic fleas, the set of host spe-
cies used by a flea often appeared to be more taxonom-
ically clustered (i.e. the host species were more closely
related to each other) than in random subsets taken
from the regional pool of host species (Krasnov et al.,
2004b). Krasnov (2008) examined the change of principal
hosts across a geographical range in 177 flea species
from 49 geographical regions and evaluated between-
region change of the identity of the principal host spe-
cies and genus. It was found that the frequency distri-
bution of the mean number of the principal host
genera per region was much less left-skewed than the
mean number of the principal host species per region,
suggesting that the species that substitutes for the prin-
cipal host across the geographical range of a flea was
often phylogenetically (or at least taxonomically)
related (Krasnov, 2008). In other cases, a parasite
expanding its geographical range was found to switch
to a very distant relative of its principal host. For exam-
ple, Hoplopsyllus anomalus is a flea normally associated
with sciurid rodents (Spermophilus). However, in the
some areas of California, this flea is found mainly on a
heteromyid rodent (Dipodomys ingens; Tabor et al.,
1993).
Furthermore, the success of a parasite to invade a
new area has often been explained by ecological fitting
which occurs when parasites specialize on a resource
that is widespread among many host species, such as
blood, and track this resource rather than a host lineage
per se (Kethley & Johnston, 1975; Janzen, 1985; Brooks
et al., 2006a,b). Our results suggest that ecological fit-
ting may be phylogenetically restricted and depends on
phylogenetic relatedness of a potentially new host to an
original principal host. A parasite would colonize host
species that are either the most closely or the most dis-
tantly related to the principal host but not hosts with
intermediate phylogenetic relatedness to the latter.
A principal host: not only physiology
In both flea species, egg and/or new imago production
did not differ significantly between the principal host
and the most closely related auxiliary host (A. russatus
for P. chephrenis and G. andersoni for X. ramesis). More-
over, the new imago production in P. chephrenis was
significantly higher in the auxiliary host A. russatus
than in the principal host A. cahirinus. This suggests
that a principal host supports the majority of indivi-
duals in a parasite population not only due to physio-
logical reasons. For fleas and hosts in this study, these
additional reasons could be associated with habitat dis-
tribution and social behaviour of host species.
Although the reproductive performance of X. ramesis
on G. andersoni was as high as on M. crassus, this flea
does not co-occur naturally with the former host in
southern Israel (Krasnov et al., 1999). This is because
X. ramesis occupies mainly habitats with loess soils
(Krasnov et al., 1997), whereas G. andersoni is a strict
sand-dweller (Mendelssohn & Yom-Tov, 1999). Both
Acomys hosts of P. chephrenis occupy the same habitats.
The main ecological difference between these hosts is
that A. cahirinus nest communally, whereas A. russatus
is solitary (Mendelssohn & Yom-Tov, 1999; Shargal
et al., 2000). Host social behaviour may affect abun-
dance and distribution of contact-transmitted parasites
(Møller et al., 1993; Loehle, 1995; Ezenwa, 2004). In
particular, group leaving promotes contact between sus-
ceptible and infected individuals, so that increasing host
group size often results in increasing prevalence and/or
abundance of a parasite (Cote & Poulin, 1995). This
seems to be the case for P. chephrenis and A. cahirinus.
In conclusion, this study revealed proximate mecha-
nisms underlying the relationships between parasite
abundance in principal and auxiliary hosts and the
ª 2 01 2 THE AUTHORS . J . E VOL . B I OL . do i : 1 0 . 1 11 1 / j . 1 4 2 0 - 9 1 0 1 . 2 0 1 2 . 0 2 5 77 . x
JOURNAL OF EVOLUT IONARY B IO LOGY ª 20 1 2 EUROPEAN SOC I E TY FOR EVOLUT IONARY B IO LOGY
Ectoparasite fitness in auxiliary hosts 7

effect of phylogenetic positions of these hosts. Ecology,
geography and behavioural factors also play a role in
manifestation of these relationships.
Acknowledgments
This study was supported by the United States-Israel
Bi-National Science Foundation [grant number
2008142 to BRK, ISK and LJF]. The study was con-
ducted under permits from the Israel Nature and
National Parks Protection Authority (permits 2010/
37131 and 2011/38082) and the Ben-Gurion Univer-
sity Committee for the Ethical Care and Use of Ani-
mals in Experiments (authorization IL-52-07-2009).
This is publication no. 774 of the Mitrani Department
for Desert Ecology.
References
Bininda-Emonds, O.R.P., Cardillo, M., Jones, K.E., MacPhee,
R.D.E., Beck, R.M.D., Grenyer, R. et al. 2007. The delayed
rise of present-day mammals. Nature 446: 507–512.Bize, P., Jeanneret, C., Klopfenstein, A. & Roulin, A. 2008.
What makes a host profitable? Parasites balance host nutri-
tive resources against immunity Am. Nat. 171: 107–118.Blomberg, S.P. & Garland, T. 2002. Tempo and mode in evolu-
tion: phylogenetic inertia, adaptation and comparative meth-
ods. J. Evol. Biol. 15: 899–910.Brooks, D.R., Leon-Regagnon, V., McLennan, D.A. & Zelmer,
D. 2006a. Ecological fitting as a determinant of the commu-
nity structure of platyhelminth parasites of anurans. Ecology
87: S76–S85.Brooks, D.R., McLennan, D.A., Leon-Regagnon, V. & Hoberg,
E. 2006b. Phylogeny, ecological fitting and lung flukes: help-
ing solve the problem of emerging infectious diseases. Rev.
Mex. Biodivers. 77: 225–233.Combes, C. 2001. Parasitism. The Ecology and Evolution of Inti-
mate Interactions. Univ. Chicago Press, Chicago.
Cote, I. & Poulin, R. 1995. Parasitism and group size in social
animals: a meta-analysis. Behav. Ecol. 6: 159–165.Davies, C.R. 1990. Interrupted feeding of blood-sucking
insects: causes and effects. Parasitol. Today 6: 19–22.Dogiel, V.A. 1964. General Parasitology, English Translation of 3rd
edn. Oliver and Boyd, Edinburgh-London.
Dogiel, V.A., Petrushevski, G.K. & Polyanski, Y.I. 1961. Parasi-
tology of Fishes, English translation of 3rd edn. Oliver and Boyd,
Edinburgh-London.
Ezenwa, V. 2004. Host social behavior and parasitic infection:
a multifactorial approach. Behav. Ecol. 15: 446–454.Gouy de Bellocq, J., Krasnov, B.R., Khokhlova, I.S., Ghasar-
yan, L. & Pinshow, B. 2006. Immunocompetence and flea
parasitism in a desert rodent. Funct. Ecol. 20: 637–646.Harrington, L.C., Edman, J.D. & Scott, T.W. 2001. Why do
female Aedes aegypti (Diptera: Culicidae) feed preferentially
and frequently on human blood? J. Med. Entomol. 38: 411–422.
Harvey, P.H. & Pagel, M.D. 1991. The Comparative Method in
Evolutionary Biology. Oxford Univ. Press, Oxford.
Holland, G.P. 1964. Evolution, classification and host relation-
ships of Siphonaptera. Annu. Rev. Entomol. 9: 123–146.
Hopkins, G.H.E. 1949. The host-associations of the lice of
mammals. Proc. Zool. Soc. Lond. 119: 387–604.Janzen, D.H. 1985. On ecological fitting. Oikos 45: 308–310.Kethley, J.B. & Johnston, D.E. 1975. Resource tracking pat-
terns in bird and mammal ectoparasites. Misc. Publ. Entomol.
Soc. Am. 9: 231–236.Khokhlova, I.S., Serobyan, V., Krasnov, B.R. & Degen, A.A.
2009a. Effect of host gender on blood digestion in fleas:
mediating role of environment. Parasitol. Res. 105: 1667–1673.
Khokhlova, I.S., Serobyan, V., Krasnov, B.R. & Degen, A.A.
2009b. Is the feeding and reproductive performance of the
flea, Xenopsylla ramesis, affected by the gender of its rodent
host, Meriones crassus? J. Exp. Biol. 212: 1429–1435.Khokhlova, I.S., Serobyan, V., Degen, A.A. & Krasnov, B.R.
2010a. Host gender and offspring quality in a flea parasitic
on a rodent. J. Exp. Biol. 213: 3299–3304.Khokhlova, I.S., Ghazaryan, L., Degen, A.A. & Krasnov, B.R.
2010b. Infestation experience of a rodent host and offspring
viability of fleas: variation among host–parasite associations.
J. Exp. Zool. A Ecol. Genet. Physiol. 313: 680–689.Khokhlova, I.S., Fielden, L.J., Degen, A.A. & Krasnov, B.R.
2012a. Feeding performance of fleas on host species: is phy-
logenetic distance between hosts important? Parasitology 139:
60–68.Khokhlova, I.S., Fielden, L.J., Degen, A.A. & Krasnov, B.R.
2012b. Digesting blood of an auxiliary host in fleas: effect of
phylogenetic distance from a principal host. J. Exp. Biol. 215:
1259–1265.Korine, C., Krasnov, B.R., Khokhlova, I.S. & Pinshow, B.
2012. Effects of host diet and thermal state on feeding per-
formance of the flea Xenopsylla ramesis. J. Exp. Biol. 215:
1435–1441.Krasnov, B.R. 2008. Functional and Evolutionary Ecology of Fleas:
A Model for Ecological Parasitology. Cambridge Univ. Press,
Cambridge.
Krasnov, B.R., Shenbrot, G.I., Medvedev, S.G., Vatschenok,
V.S. & Khokhlova, I.S. 1997. Host-habitat relations as an
important determinant of spatial distribution of flea assem-
blages (Siphonaptera) on rodents in the Negev Desert. Para-
sitology 114: 159–173.Krasnov, B.R., Hastriter, M., Medvedev, S.G., Shenbrot, G.I.,
Khokhlova, I.S. & Vatshchenok, V.S. 1999. Additional
records of fleas (Siphonaptera) on wild rodents in the south-
ern part of Israel. Israel J. Zool. 45: 333–340.Krasnov, B.R., Khokhlova, I.S., Fielden, L.J. & Burdelova, N.V.
2001. Development rates of two Xenopsylla flea species in
relation to air temperature and humidity. Med. Vet. Entomol.
15: 249–258.Krasnov, B.R., Khokhlova, I.S., Oguzoglu, I. & Burdelova, N.V.
2002. Host discrimination by two desert fleas using an odour
cue. Anim. Behav. 64: 33–40.Krasnov, B.R., Sarfati, M., Arakelyan, M.S., Khokhlova, I.S.,
Burdelova, N.V. & Degen, A.A. 2003. Host-specificity and
foraging efficiency in blood-sucking parasite: feeding pat-
terns of a flea Parapulex chephrenis on two species of desert
rodents. Parasitol. Res. 90: 393–399.Krasnov, B.R., Shenbrot, G.I., Khokhlova, I.S. & Poulin, R.
2004a. Relationships between parasite abundance and the
taxonomic distance among a parasite’s host species: an
example with fleas parasitic on small mammals. Int. J. Parasi-
tol. 34: 1289–1297.
ª 2 0 12 THE AUTHORS . J . E VOL . B IO L . do i : 1 0 . 1 1 1 1 / j . 1 4 20 - 9 10 1 . 2 01 2 . 0 25 7 7 . x
JOURNAL OF EVOLUT IONARY B IOLOGY ª 2012 EUROPEAN SOC I E TY FOR EVOLUT IONARY B IO LOGY
8 I . S . KHOKHLOVA ET AL.

Krasnov, B.R., Mouillot, D., Shenbrot, G.I., Khokhlova, I.S. &
Poulin, R. 2004b. Geographical variation in host specificity
of fleas (Siphonaptera): the influence of phylogeny and local
environmental conditions. Ecography 27: 787–797.Krasnov, B.R., Korine, C., Burdelova, N.V., Khokhlova, I.S. &
Pinshow, B. 2007. Between-host phylogenetic distance and
feeding efficiency in haematophagous ectoparasites: Rodent
fleas and a bat host. Parasitol. Res. 101: 365–371.Lehane, M.J. 2005. The Biology of Blood-Sucking in Insects, 2nd
edn. Cambridge Univ. Press, Cambridge.
Loehle, C. 1995. Social barriers to pathogen transmission in
wild animal populations. Ecology 76: 326–335.Lyimo, I.N. & Ferguson, H.M. 2009. Ecological and evolution-
ary determinants of host species choice in mosquito vectors.
Trends Parasitol. 25: 189–196.Lyimo, I.N., Keegan, S.P., Ranford-Cartwright, L.C. & Fergu-
son, H.M. 2012. The impact of uniform and mixed species
blood meals on the fitness of the mosquito vector Anopheles
gambiae s.s.: does a specialist pay for diversifying its host spe-
cies diet? J. Evol. Biol., 25: 452–460.Marshall, A.G. 1981. The Ecology of Ectoparasitic Insects. Acad.
Press, London.
Mendelssohn, H. & Yom-Tov, Y. 1999. Mammalia of Israel.
Fauna Palaestina. Israel Acad. Sci. Hum., Jerusalem.
Møller, A., Dufva, R. & Allander, K. 1993. Parasites and the
evolution of host social behavior. Adv. Stud. Behav. 22: 65–102.
Moloo, S.K., Grootenhuis, J.G., Kar, S.K. & Karstad, L. 1988.
Survival and reproductive performance of female Glossina
morsitans when maintained on the blood of different species
of wild mammals. Med. Vet. Entomol. 2: 347–350.Morris, D.W. 1987. Ecological scale and habitat use. Ecology 68:
362–369.Paradis, E., Claude, J. & Strimmer, K. 2004. APE: analyses of
phylogenetics and evolution in R language. Bioinformatics 20:
289–290.Poulin, R. 2005. Relative infection levels and taxonomic dis-
tances among the host species used by a parasite: insights
into parasite specialization. Parasitology 130: 109–115.
Poulin, R. & Mouillot, D. 2004. The relationship between spe-
cialization and local abundance: the case of helminth para-
sites of birds. Oecologia 140: 372–378.Poulin, P., Brodeur, J. & Moore, J. 1994. Parasite manipulation
of host behaviour: should hosts always loose? Oikos 70: 479–484.
R Development Core Team. 2011. R: A Language and Environ-
ment for Statistical Computing. R Foundation for Statistical
Computing, Vienna. ISBN 3-900051-07-0, URL http://www.
R-project.org/.
Rosenzweig, M.L. 1981. A theory of habitat selection. Ecology
62: 327–335.Rosenzweig, M.L. 1987. Habitat selection as a source of biolog-
ical diversity. Evol. Ecol. 1: 315–330.Rosenzweig, M.L. 1991. Habitat selection and population inter-
actions: the search of mechanism. Am. Nat. 137: 5–28.Sarfati, M., Krasnov, B.R., Ghazaryan, L., Khokhlova, I.S.,
Fielden, L.J. & Degen, A.A. 2005. Energy costs of blood
digestion in a host-specific haematophagous parasite. J. Exp.
Biol. 208: 2489–2496.Shargal, E., Kronfeld-Schor, N. & Dayan, T. 2000. Population
biology and spatial relationships of coexisting spiny mice (Ac-
omys) in Israel. J. Mammal. 81: 1046–1052.Shudo, E. & Iwasa, Y. 2001. Inducible defense against patho-
gens and parasites: optimal choice among multiple options.
J. Theor. Biol. 209: 233–247.Tabor, S.P., Williams, D.F., Germano, D.J. & Thomas, R.E. 1993.
Fleas (Siphonaptera) infesting giant kangaroo rats (Dipodomys
ingens) on the Elkhorn and Carrizo plains, San Luis Obispo
County, California. J. Med. Entomol. 30: 291–294.Wenzel, R.L. & Tipton, V.J. 1966. Some relationships between
mammal hosts and their ectoparasites. In: Ectoparasites of
Panama (R.L. Wenzel & V.J. Tipton, eds.), pp. 677–723. FieldMus. Nat. Hist., Chicago.
Data deposited at Dryad: doi:10.5061/dryad.8p6f2
Received 29 May 2012; accepted 25 June 2012
ª 2 01 2 THE AUTHORS . J . E VOL . B I OL . do i : 1 0 . 1 11 1 / j . 1 4 2 0 - 9 1 0 1 . 2 0 1 2 . 0 2 5 77 . x
JOURNAL OF EVOLUT IONARY B IO LOGY ª 20 1 2 EUROPEAN SOC I E TY FOR EVOLUT IONARY B IO LOGY
Ectoparasite fitness in auxiliary hosts 9