ecological significance of nitrogen cycling by tubificid communities in shallow eutrophic lakes of...
TRANSCRIPT

Ecological significance of nitrogen cycling by tubificid communities in shallow
eutrophic lakes of the Danube Delta
Rıs�noveanu Geta*, Carmen Postolache & Anghelut��a V�adineanuDepartment of Systems Ecology, University of Bucharest, Spl. Independentei 91-95, 76201 Bucharest, Romania
(*Author for correspondence: Fax: +40-21-4112310, E-mail: [email protected])
Received 16 April 2003; in revised form 14 January 2004; accepted 16 January 2004
Keywords: tubificids, nitrogen cycle, ecological significane, Danube Delta
Abstract
Importance of tubificid populations on nitrogen cycle in two categories of shallow eutrophic lakes in theDanube Delta was quantitatively assessed for the 1992–1993 period. The structure of the primary producersin the studied lakes was used to discriminate between the two categories: (i) lakes dominated by macro-phytes (A1) and (ii) lakes dominated by phytoplankton (A2). In both categories tubificid worms representedimportant fraction of the entire benthic community (35 and 32%, respectively, as number of individuals).They influence the sediment–water exchange of nutrients. The main processes involved are excretion ofnutrients and their continuous release from sediments by molecular diffusion or through channels createdby bioturbation. Inorganic nitrogen released from bottom sediments may regulate nitrogen load in thewater body and thus, phytoplankton production. In 1992–1993, nitrogen stocks in tubificid biomass ac-counted for 5.3% in A1 lakes and 15.6% in A2 lakes of the amount stocked in phytoplankton, and only for1.2 and 2.9% respectively, of the nitrogen load in water body. Nitrogen excretion rates ranged between60.52 and 153.74 mg N m)2 year)1, and release rates from sediments between 378.26 and960.87 mg N m)2 year)1, the lowest values being recorded for A2 category. Differences are related totubificid biomass, structure and abundance of primary producers and to nutrient load in different eco-systems. Ratios between release rate of inorganic nitrogen by tubificid worms and sedimentation rate oforganic nitrogen in the two categories of lakes were 8.3 and 6.4% respectively. Contribution of nitrogenreleased daily from sediments to the dissolved inorganic nitrogen load in the water column was less than0.5%. However, in A1 and A2 lakes, the released nitrogen had a potential to sustain 24.74 and 8.01%,respectively, of the annual phytoplankton production. These values suggest the significance of tubificids inkeeping the eutrophication process at a high level, especially during the periods when nitrogen is the mainlimiting factor for phytoplankton production.
Introduction
Wetlands of deltas have been shown to trap, store,transform and release nutrients, thus drasticallymodifying the chemistry of the water flowingthrough them and impacting water quality down-stream (Whigham et al., 1988; Cristofor et al.,1993). Nitrogen and phosphorus are the mostcrucial nutrients in this respect because they can
have strongly negative impacts on water qualityand may limit the growth of primary producers(V�adineanu & Cristofor, 1987). Although phos-phorus is most responsible for regulating phyto-plankton production in freshwater lakes, nitrogenmay also be in short supply in highly eutrophiclakes (Claesson & Ryding, 1977). Thus, the degree
Hydrobiologia 524: 193–202, 2004.� 2004 Kluwer Academic Publishers. Printed in the Netherlands. 193

of inorganic nitrogen shortage in lake watermay be important in regulating phytoplank-ton production (V�adineanu et al., 1989). The re-lease of inorganic nitrogen from bottomsediments may regulate nitrogen load in lakes(Tatrai, 1987; Fukuhara & Yasuda, 1989). Thisprocess has a special significance in shallow lakeswhere thermal stratification is easily destroyed bythe wind action. Nitrogen can be introduced di-rectly into epilimnetic water and is thus readilyavailable for uptake by phytoplankton. Thus, inshallow-lake ecosystems the bottom sediments areessential to sustain active biological productionthat is well coupled with nitrogen and phosphoruscycling.
The benthos (comprised mostly by tubificidsand chironomids) may influence nutrient dynam-ics of such lakes: (i) by nutrient sequestration inbiomass and transfer of nutrients through foodwebs to other trophic levels, (ii) through excretionof nutrient compounds and also (iii) throughcontinuous release of nutrients from sedimentsthrough channels created by bioturbation activity(Davis et al., 1975; Fukuhara & Sakamoto, 1987;Sakamoto et al., 1989). Although there have beensome extensive experimental studies on nitrogenexcretion and release rates by different benthicspecies (Gardner et al., 1983; Tatrai, l987;Fukuhara & Yasuda, 1989; Rıs�noveanu et al.,2001), little effort has been done to assessthe zoobenthos community contribution to thenutrient cycle in natural lake ecosystems and thenutrient budget at the ecosystem level. Neverthe-less, an integrated assessment of the communities’role at the ecosystem level is of primary impor-tance for scientific guidance of managerial deci-sions.
In this study we report the results of an eco-system approach undertaken to quantitatively as-sess the role of tubificids in the nitrogen budget ofeutrophic shallow lakes of the Danube Delta andthe ecological significance of this process. To an-swer such a question, rates of nutrient excretionand release rates by macrozoobenthos are relatedto the biomass of macroinvertebrates in sedimentsand compared to: (i) sedimentation rates of or-ganic nitrogen; (ii) nitrogen mineralization rates byother biotic components; (iii) nitrogen productionat the phytoplankton level; (iv) nutrient loads inthe water column.
Description of the site studied
The Danube Delta is one of the most importantwetland areas in Europe because of its ecologicalvalue, surface area and geographical position at theterminal zone of the second largest river in Europe.It functions as an interface between the DanubeRiver System and the Northwestern Black Sea. TheDanube Delta receives the drainage from805 300 km2 and discharges annually about 76% ofthe total freshwater input to the sea (Gas�tescu,1993). The area of flooded wetlands of the DanubeDelta has decreased by 25% since 1965. Conse-quently, the filtering potential of the Danube wet-lands has decreased as well. Several authors(V�adineanu & Cristofor, 1987; Cristofor et al.,1993) reported increasing nitrogen and phosphorusloads in the Danube River during the last decades.Increased eutrophication induced remarkablechanges of both water quality and structure of theprimary producers and, subsequently, along thefood webs (V�adineanu et al., 2002).
During the reference period (1980–1982) in al-most all lakes the primary producers were repre-sented by both the phytoplankton and aquaticvegetation. Lentic ecosystems of the Danube Deltahave followed two different pathways since then.The first one evolved towards a structure thatpreserved a balanced energy flow through phyto-plankton and aquatic vegetation only in the firststages (early 1980s). During the late 1980s thestructure of the primary producers shifted towardsa very significant increase of macrophytes–epi-phyte complex abundance and therefore of theenergy flow at this level (over 75% of the totalsolar energy flow through the primary producers).The second pathway leads since early 1980s to-wards a structural organization dominated by thephytoplankton (10 500 kcal m)2 year)1) (V�adi-neanu et al., 2002). During this period the benthiccommunities have simplified, with only chirono-mid and tubificid species increasing in abundance(Botnariuc et al., 1987; R�s�noveanu et al., 1998). Adecline of benthic species richness was also re-corded (R�s�noveanu et al., 1997).
We assessed the role of tubificids in the nitro-gen cycling for the two categories of ecosystems:A1 – those in which the primary producers aredominated by macrophytes (Lakes Isacova,B�acl�anes�ti and Bogdaproste), and A2 – those in
194

which the primary producers are represented byphytoplankton (Lakes Puiu, Ros�u, Babina, Ma-tit�a and Merhei) (Fig. 1). All the eight lakesstudied are representative of the Danube Deltalentic ecosystems. These lakes are included in thelong-term ecological research network and de-tailed characterization of their morphological,physical and chemical variables were provided byseveral authors (Cristofor, 1987; V�adineanu &Cristofor, 1987; Cristofor et al., 1993). This studyis part of an integrated research programmeimplemented in these ecosystems.
Material and methods
Zoobenthos standing crop
Three benthic sample units were taken from eachof five sites randomly dispersed across a lake, and
this series was repeated 11 times from March 1992to October 1993. In 1992 a set of six samples wastaken at intervals of latest 2 months, except forJanuary when an ice layer covered the water.During 1993 the samples were taken monthly be-tween May and October. All the sampling ofzoobenthos, phytoplankton and macrophytes wereconducted at the same times in the eight lakes ofthe Danube Delta. Concurrent surveys of envi-ronmental conditions in the lake water were con-ducted. The samples were collected with a 50 cm2
corer, washed through a sieve (mesh size 250 lm),and the retained sediments and fauna were pre-served in 4% formaldehyde. All of the faunalspecimens were hand sorted from sediments undera Zeiss stereomicroscope and transferred to sepa-rate containers filled with 70� alcohol. The faunawere subsequently identified to species level in thelaboratory. Identification of the taxa and
Figure 1. Geographical position of the Danube Delta and location of studied lakes.
195

measurements were made using a phase contrastmicroscope.
Wet weight of each worm was measured usingan OHAUS electromicrobalance accurate to1 · 10)7 g. Dry weights were obtained after dryinganimals for 24 h at 60 �C. Secondary productionwas calculated according to the method describedby Hamilton (1969) (Rıs�noveanu & V�adineanu,2000). Production values were considered to esti-mate the nitrogen transferred towards other tro-phic levels.
Nutrient stocks in biomass
Nitrogen stocks in the biomass of tubificids weremeasured on qualitative samples taken with amovable dredge. Samples were washed through a230 · 230 lm mesh. Small amounts of sedimentswere added to water in a large, white tray. Livingindividuals were gently collected with tweezers andtransferred for 24 h in vials containing lake waterfor emptying their guts. Over 80% of individualswere identified as Potamothrix hammoniensis, theothers belonging to Limnodrilus spp. Dry weightwas obtained after drying animals for 24 h at60 �C. Between 51.4 and 259 mg dried biomasses(from each lake) were mineralized and nitrogencontent per biomass unit was determined. Nitro-gen stocks were assessed by multiplication of thesevalues with the total biomass of the dominanttubificid populations.
Stocks in phytoplankton and macrophytes
Sedimentation rate
Chlorophyll a concentration was determined byspectrophotometry on acetone extracts of phyto-plankton, quantitatively concentrated in field on0.8 lm membrane filters. The acetone extractionlasted 24 h in darkness, at 7 �C. For conversion ofchlorophyll a content to biomass and carboncontent, we referred to conversion factors ob-tained by V�adineanu et al. (1985) for the sameecosystems. A C:N ratio of 5 (the lowest value inthe literature) was assumed for nitrogen stockassessment in phytoplankton (Vollenweider, 1969).
The macrophytes biomass during the studyperiod and the nitrogen stocks per biomass unit(Sarbu, 1997) were used for the assessment of
macrophytes production and stocks in terms ofnitrogen.
The annual sedimentation rate of organicnitrogen through the water column was consideredto be 70% of the phytoplankton production(V�adineanu et al., 1985) and the entire macro-phytes production.
Excretion and release rates
Since metabolic rate is related to individual size,the excretion rates used for assessing the metabolicmineralization of nitrogen by tubificid worms weredifferentiated as follows: (i) for the individualshaving dry weights less than 0.25 mg/individualwe used the excretion rates reported by Rıs�nov-eanu & V�adineanu (2000) and Rıs�noveanu et al.(2001) for the tubificid populations from DanubeRiver (0.0045 lg P-PO�
4 mg)1 h)1, 0.138 lgN-NHþ
4 mg)1 h)1, 0.065 lg N-NO�3 mg)1 h)1 (ii)
for the individuals of more than 0.25 mg dryweight we used the rates reported for Limnodrilusspp. by Fukuhara & Sakamoto (1987) (0.003 lgP-PO�
4 mg)1 h)1, 0.07 lg N-NHþ4 mg)1 h)1). As
Devine & Vanni (2002) showed the nutrientflux through benthic invertebrate is affectedmore by total biomass and temperature than byspecies composition, we assumed that therewould not be significant differences between theexcretion rates obtained for Limnodrilus spp.and those for P. hammoniensis. For the relation-ship of excretion to temperature we referred tothe Q10 coefficients from Fukuhara and Yasuda(1989).
Time required for an individual animal to ex-crete an amount of nitrogen equal to its bodynitrogen content (body equivalent excretion time,BEET) was estimated from data of body nitrogencontent and average daily excretion rates at thetemperature range in the studied lakes (providedby the monitoring programme). Mean monthlytemperatures at the sediment–water interface wereused.
The release rates were assessed indirectly, basedon the ratio of excretion to release rates in litera-ture. The value of 0.16 for this ratio was used dueto the fact that it was obtained by Fukuhara &Sakamoto (1987) in experiments with tubificidworms at densities of less than 4500 ind m)2,
196
densities that are similar to those recorded in theDanube Delta lakes.
Chemical analyses
Tubificid dried biomasses were mineralized withsulphuric acid and hydrogen peroxide. Solutionsobtained were neutralized and analysed by thespecific method for ammonium (Kempers, 1974).Water analyses were performed on filtered sam-ples, using colorimetric methods: ammonium bythe indophenol-blue method, and nitrate by phe-nol–disulphonic acid method (Bremner, 1965).
Results
Lake conditions during the study period
The main indicators of the trophic state of the twotrophic categories of ecosystems are presented inTable 1. Throughout the survey period themonthly average of total reactive phosphorus(TRP) load varied between 13.3 and 95.3 lg P l)1
in the A1 ecosystems and between 9.2 and
39.1 lg P l)1 in the A2 ecosystems. The monthlyaverage of dissolved inorganic nitrogen (DIN)load varied in similar ranges in the two categoriesof ecosystems (between 151.4 and 650.4 lg N l)1).Low values of N:P ratios were observed, especiallyduring the summer. The lowest value (2.8) wasrecorded in A1 ecosystems in August. The con-centrations of chlorophyll a ranged between 6.1and 91.8 lg l)1 and the relative transparency(Secchi transparency to depth: T/D) had low val-ues (frequently under 40%). The highest values ofthe sedimented organic matter (SOM) were re-corded in May–June and September.
Biomass of zoobenthos
In the ecosystems where macrophytes dominatedamong the primary producers (A1) the oligochaeteassemblages represented an average of 35% asnumber and 25.6% as biomass of the benthicfauna. In the ecosystems where phytoplanktondominated the primary producers (A2), the oligo-chaete assemblages represented an average of 32%as number and 15.2% as biomass of the benthicfauna. Figure 2 illustrates the abundance and
Table 1. The monthly average values of the main state indicators: total reactive phosphorous (TRP), dissolved inorganic nitrogen
(DIN), ratio of nitrogen to phosphorous (N/P), chlorophyll a (Chl a), ratio of Secchi transparency to depth (T/D) and sedimented
organic matter (SOM) in two categories of lakes (A1 and A2) of the Danube Delta, during 1992–1993
A1 TRP (lg l)1) DIN (lg l)1) N/P Chl a (lg l)1) T/D SOM (%)
March 39.0 421.7 10.8 28.7 0.3 3.3
April 24.2 192.7 15.6 15.7 0.5 3.4
May 29.0 151.4 6.8 6.1 0.5 10.3
June 39.7 629.1 19.0 6.1 0.8 35.3
July 48.3 321.4 12.8 47.2 0.3 3.1
August 95.3 264.0 2.8 46.8 0.2 4.2
September 37.7 650.4 29.5 59.7 0.3 12.8
October 13.3 501.3 40.8 28.7 0.2 8.1
A2
March 9.2 620.1 67.6 36.2 0.4 2.3
April 11.2 324.8 88.2 23.2 0.3 3.2
May 26.9 392.7 14.9 7.8 0.4 10.1
June 39.1 328.2 12.5 14.9 0.2 23.5
July 18.2 201.9 11.9 51.9 0.1 6.0
August 25.1 591.4 25.6 31.3 0.2 3.6
September 9.2 171.0 49.3 91.8 0.1 18.6
October 13.7 433.7 32.3 27.2 0.1 11.1
197
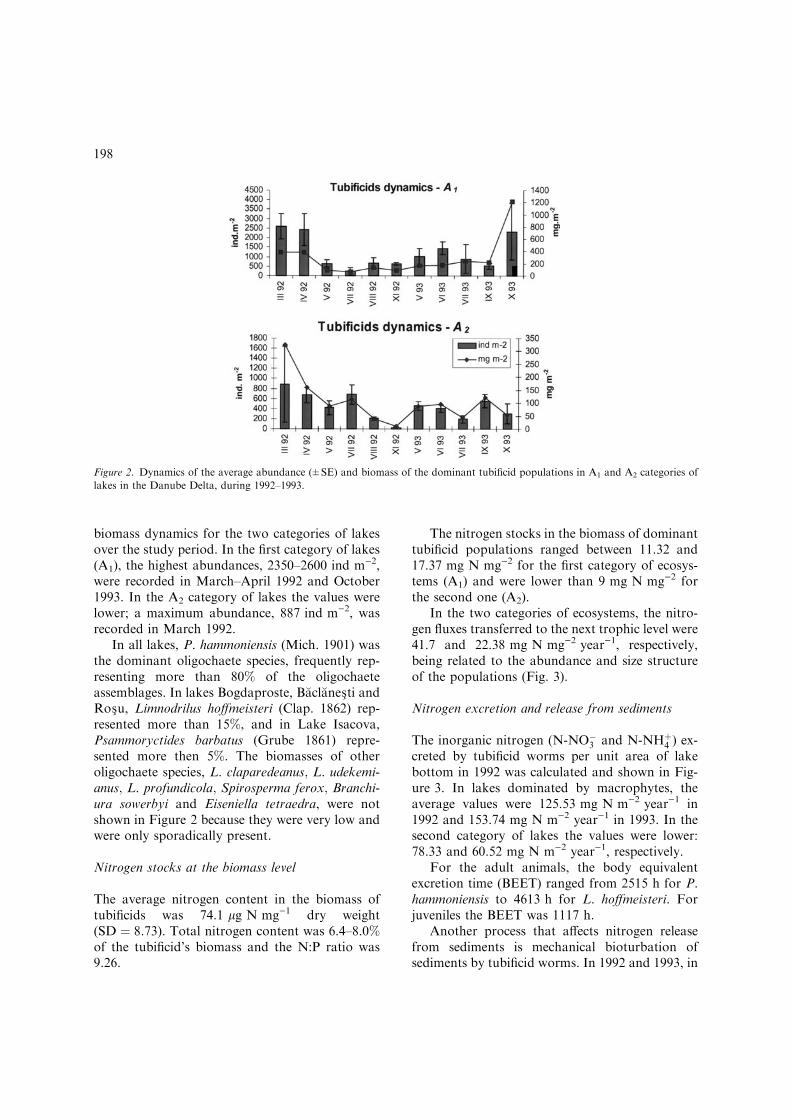
biomass dynamics for the two categories of lakesover the study period. In the first category of lakes(A1), the highest abundances, 2350–2600 ind m)2,were recorded in March–April 1992 and October1993. In the A2 category of lakes the values werelower; a maximum abundance, 887 ind m)2, wasrecorded in March 1992.
In all lakes, P. hammoniensis (Mich. 1901) wasthe dominant oligochaete species, frequently rep-resenting more than 80% of the oligochaeteassemblages. In lakes Bogdaproste, B�acl�anes�ti andRos�u, Limnodrilus hoffmeisteri (Clap. 1862) rep-resented more than 15%, and in Lake Isacova,Psammoryctides barbatus (Grube 1861) repre-sented more then 5%. The biomasses of otheroligochaete species, L. claparedeanus; L. udekemi-anus; L. profundicola; Spirosperma ferox; Branchi-ura sowerbyi and Eiseniella tetraedra, were notshown in Figure 2 because they were very low andwere only sporadically present.
Nitrogen stocks at the biomass level
The average nitrogen content in the biomass oftubificids was 74.1 lg N mg)1 dry weight(SD ¼ 8.73). Total nitrogen content was 6.4–8.0%of the tubificid’s biomass and the N:P ratio was9.26.
The nitrogen stocks in the biomass of dominanttubificid populations ranged between 11.32 and17.37 mg N mg)2 for the first category of ecosys-tems (A1) and were lower than 9 mg N mg)2 forthe second one (A2).
In the two categories of ecosystems, the nitro-gen fluxes transferred to the next trophic level were41.7 and 22.38 mg N mg)2 year)1, respectively,being related to the abundance and size structureof the populations (Fig. 3).
Nitrogen excretion and release from sediments
The inorganic nitrogen (N-NO�3 and N-NHþ
4 ) ex-creted by tubificid worms per unit area of lakebottom in 1992 was calculated and shown in Fig-ure 3. In lakes dominated by macrophytes, theaverage values were 125.53 mg N m)2 year)1 in1992 and 153.74 mg N m)2 year)1 in 1993. In thesecond category of lakes the values were lower:78.33 and 60.52 mg N m)2 year)1, respectively.
For the adult animals, the body equivalentexcretion time (BEET) ranged from 2515 h for P.hammoniensis to 4613 h for L. hoffmeisteri. Forjuveniles the BEET was 1117 h.
Another process that affects nitrogen releasefrom sediments is mechanical bioturbation ofsediments by tubificid worms. In 1992 and 1993, in
Figure 2. Dynamics of the average abundance (±SE) and biomass of the dominant tubificid populations in A1 and A2 categories of
lakes in the Danube Delta, during 1992–1993.
198
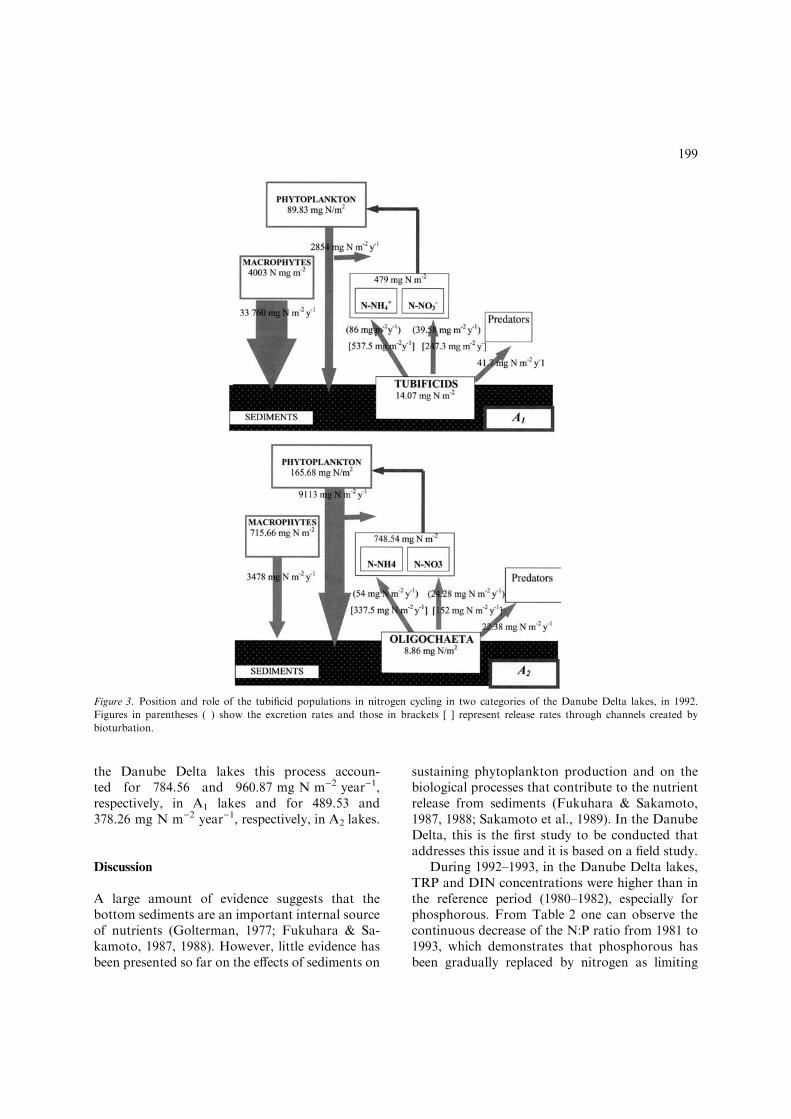
the Danube Delta lakes this process accoun-ted for 784.56 and 960.87 mg N m)2 year)1,respectively, in A1 lakes and for 489.53 and378.26 mg N m)2 year)1, respectively, in A2 lakes.
Discussion
A large amount of evidence suggests that thebottom sediments are an important internal sourceof nutrients (Golterman, 1977; Fukuhara & Sa-kamoto, 1987, 1988). However, little evidence hasbeen presented so far on the effects of sediments on
sustaining phytoplankton production and on thebiological processes that contribute to the nutrientrelease from sediments (Fukuhara & Sakamoto,1987, 1988; Sakamoto et al., 1989). In the DanubeDelta, this is the first study to be conducted thataddresses this issue and it is based on a field study.
During 1992–1993, in the Danube Delta lakes,TRP and DIN concentrations were higher than inthe reference period (1980–1982), especially forphosphorous. From Table 2 one can observe thecontinuous decrease of the N:P ratio from 1981 to1993, which demonstrates that phosphorous hasbeen gradually replaced by nitrogen as limiting
Figure 3. Position and role of the tubificid populations in nitrogen cycling in two categories of the Danube Delta lakes, in 1992.
Figures in parentheses ( ) show the excretion rates and those in brackets [ ] represent release rates through channels created by
bioturbation.
199

factor for phytoplankton development. Thisstatement is also supported by the changes ob-served in the phytoplankton composition, fromthe dominance of green algae to blue-green algae(V�adineanu et al., 1989). The chlorophyll a contentwas also maintained at high levels, which inducedlow values (below 40%) for relative transparency.In these particular conditions tubificids accountedfor 25.6% from the biomass of the benthic com-munities in A1 type of lakes and only for 15.2% inA2 type, representing the second largest groupwithin the communities.
The nitrogen content in tubificid biomass,ranged between 6.4 and 8%. Fukuhara & Yasuda(1989) reported an average content of nitrogen of8.55% of the biomass of Limnodrilus spp. and anitrogen to phosphorus ratio of 8.9 which are onlyslightly different of those obtained for the tubific-ids in the Danube Delta. In the A1 and A2 cate-gories of lakes nitrogen stocks in biomassaccounted for 5.3 and 15.6% respectively, from theamount stocked in phytoplankton and only for 1.2and 2.9% respectively, from the nitrogen load inwater body. The differences are related to tubificidbiomass, structure and abundance of primaryproducers and to nutrient load in different lakes(Fig. 3). Tubificids excrete nutrients through ne-phridia located in each segment of their body.Therefore, nutrients are released into both thewater column and sediments. The present resultsshow that the excreted rates of inorganic nitrogenwere higher in the ecosystems dominated by mac-rophytes (up to 153.74 mg N m)2 year)1 in 1993).However, due to the fact that not all the excretaare directly released into lake water phase but alsowithin the sediments, the values presented in thispaper correspond to the potential values.
Nitrogen mineralized in sediments is easily re-leased into a water column through channels cre-ated from zoobenthos bioturbation. Theexcretion-to-release ratio for ammonium aroseinterest in considering the release process of thischemical species (Fukuhara and Sakamoto, 1987,1988). The values of excretion-to-release ratioobtained in experimental conditions suggest thatthe excretion process plays a minor role inammonium release from sediments. That means,other processes may be responsible for greater re-lease of ammonium from sediments. As Fukuhara& Sakamoto (1987, 1988) and Sakamoto et al.(1989) reported, the ammonium release from sed-iments by bacterial activities might not be themajor process affecting nitrogen release, as it ismost active in a few millimetres thin layer at thesediment surface. Based on experimental evidence,they suggest that the bioturbation activities ofmacrozoobenthic animals significantly increase thediffusion of ammonium from sediments, and thatthe release rates increase with the increase of ani-mal density. However, the values reported in lit-erature for the inorganic nitrogen release rates bythe entire benthic communities through bioturba-tion vary in a wide range, from 252 mg to873 · 102 mg N m)2year)1 (recalculated fromFukuhara & Sakamoto, 1988). Most likely, thedifferences are related to those in the biomassdensities of zoobenthos communities. Accordingwith Fukuhara & Sakamoto (1988) the watertemperature has less influence on nitrogen releasefrom sediment, and the biomass of the zoobenthoscommunities is of importance to regulate theinorganic nitrogen release from sediments. Underexperimental conditions, with different densities ofLimnodrilus spp., Fukuhara & Sakamoto (1987)obtained excretion-to-release ratios less than 1.For densities of 20 ind/pot (equivalent to anabundance less than 4500 ind m)2) they reported aratio of 0.16 ± 0.03. Given the abundance re-corded for the tubificid populations in the DanubeDelta lakes, we considered this value to beappropriate for assessing the nitrogen release ratesfrom sediments. Calculated values in the lakesstudied ranged between 378.26 and960.87 mg N m)2 year)1. We were not able tocompare our data with the reported values becauseof differences in employed methods, temperatureand season. Besides, they refer to the entire benthic
Table 2. The range values of the main trophic parameters in
three different periods in the Danube Delta lakes
Parameter 1981–1982 1986–1987 1991–1993
TRP (l gl)1) 3–34 10–208 3–116
DIN (l gl)1) 90–800 250–800 90–1000
N/P 14–116 <7–59 <3–45
CHLa (l gl)1) 2–65 13–190 2–120
SOM (%) 12 15–25 2–45
(TPR – total reactive phosphorous, DIN – dissolved inorganic
nitrogen, N/P – nitrogen to phosphorous ratio, CHLa –
chlorophyll a, SOM – sedimented organic matter).
200
communities and not only to oligochaete com-munities.
The mineralization of nitrogen in sediments isconsidered to be of similar importance for phyto-plankton production as mineralized nitrogen inwater column (Fukuhara & Sakamoto, 1988). Inthe Danube Delta lakes, ratios of average excretedinorganic nitrogen by tubificids to sedimented or-ganic nitrogen were 1.32% for the first category oflakes, and 1.02% for the second category of lakes.Ratios of release rate of inorganic nitrogen bytubificids to sedimentation rate of organic nitrogenwere 8.26 and 6.39%, respectively (Table 3). Con-sidering the tubificid’s fraction of the entire ben-thic fauna and the low values obtained for thesedimentation to release ratio of nitrogen it seemsthat in the Danube Delta lakes nitrogen sedimen-tation was somehow unbalanced with the inor-ganic nitrogen release from sediments. In a certainmeasure, this fact could be induced by the 100% ofmacrophyte biomass we have considered to reachsediments. Due to this fact these values have to beconsidered as minimum potential values.
The nitrogen excreted and released from sedi-ments per day can contribute with less than 0.07%and respectively 0.5% of the load of DIN in thewhole water column due to the occurrence of inor-ganic nitrogen in high concentration in lake water.
The average amount excreted daily by tubific-ids accounted for less than 0.5% of the nitrogen
stocks in the phytoplankton standing crop.Even the average amounts released daily bythe tubificids in the A1 and A2 category oflakes accounted for 2.61 and 0.85% respectively,of the nitrogen standing stocks at the phyto-plankton level, the released nitrogen from sedi-ments is of significant importance for thephytoplankton production. The released nitrogenhad a potential to sustain 24.74 and 8.01%respectively, of the annual phytoplankton pro-duction.
The value of the released nitrogen to sustainphytoplankton production suggests the impor-tance of tubificids in keeping the eutrophicationprocess at a high level, especially during the peri-ods when nitrogen is the main limiting factor forphytoplankton production.
References
Botnariuc, N., G. Ignat, I. Diaconu & A. V�adineanu, 1987. L’evolution de l’etat trophique des ecosystemes aquatiques
caracteristiques au Delta du Danube: 5. La structure et la
dynamique de la faune benthonique. Revue Roumaine de
Biologie -Biologie Animale 32: 121–127.
Bremner, JM., 1965. Inorganic forms of nitrogen. In Black,
C. A. et al. (eds), Methods of Soil Analysis. American
Society of Argon, Inc., Madison, WI: 1179–1237.
Claesson, A. & S. O. Ryding, 1977. Nitrogen – a growth lim-
iting nutrient in eutrophic lakes. Progresses in Water Tech-
nology 8: 291—199.
Table 3. Ratios of inorganic nitrogen excretion and release rates to organic nitrogen sedimentation rate, standing stocks of dissolved
inorganic nitrogen in the water column (DIN), phytoplankton standing stocks and production rates in two categories of lakes (A1 and
A2) in the Danube Delta, in 1992
Category of lakes Daily excreted · 100 (%) Excreted 1992 · 100 (%)
DIN Phytoplankton
standing stock
Phytoplankton
production
Sedimented
(phytoplankton)
Sedimented
(phytoplankton
& macrophytes)
A1 0.07 0.42 2.77 3.96 1.32
A2 0.03 0.14 0.9 1.28 1.02
Daily released · 100 (%) Released 1992 · 100 (%)
DIN Phytoplankton
standing stock
Phytoplankton
production
Sedimented
(phytoplankton)
Sedimented
(phytoplankton
& macrophytes)
A1 0.41 2.61 17.32 24.74 8.26
A2 0.17 0.85 5.61 8.01 6.39
201

Cristofor, S., 1987. L’ evolution de l’etat trophique des
ecosystemes aquatiques caracteristiques au Delta du Dan-
ube: 6. Responses de la vegetation submerse en fonction de
la reserve de nutrients et du regime hidrologique. Revue
Roumaine de Biologie – Biologie Animale 32: 129–138.
Cristofor, S., A. Vadineanu & G. Ignat, 1993. Importance of
flood zones for nitrogen and phosphorus dynamic in the
Danube Delta. Hydrobiologia 251: 143–148.
Davis R. B., D. L. Thurlow & F. E. Brewster, 1975. Effects of
burrowing tubificid worms on the exchange of phosphorus
between lake sediment and overlying water. Verhandlungen.
Proceedings of International Association of Theoretical and
Applied Limnology 19: 382–394.
Devine J. A. & M. J. Vanni, 2002. Spatial and seasonal varia-
tion in nutrient excretion by benthic invertebrates in a
eutrophic reservoir. Freshwater Biology 47: 1107–1121.
Fukuhara, H. & M. Sakamoto, 1987. Enhancement of inor-
ganic nitrogen and phosphate release from lake sediment
by tubificid worms and chironomid larvae. Oikos 48: 312–
320.
Fukuhara, H. & M. Sakamoto, 1988. Ecological significance of
bioturbation of zoobenthos community in nitrogen release
from bottom sediments in a shallow eutrophic lake. Archiv
fur Hydrobiolgie 113: 425–445.
Fukuhara, H. & K. Yasuda, 1989. Ammonium excretion by
some freshwater zoobenthos from an eutrophic lake. Hyd-
robiologia 173: 1–8.
Gardner, W. S., T. F. Nalepa, D.S. Slavenus & G. A. Laird,
1983. Patterns and rates of nitrogen release by benthic Chi-
ronomidae and Oligochaeta. Canadian Journal of Fisheries
and Aquatic Sciences 40: 259–266.
Gates�cu, P., 1993. The Danube Delta: geographical character-
istics and ecological recovery. Geo Journal 29: 57–67.
Golterman, H. L. (ed.), 1977. Interactions between Sediments
and Freshwater. Dr W. Junk Publishers, The Hague.
Hamilton, L. A., 1969. On estimating annual production.
Reprinted from Limnology and Oceanography 14: 771–782.
Kempers A. J., 1974. Determination of sub-microquantities of
ammonium and nitrates in soil with phenol, sodium nitro-
prusside and hipochlorite. Geoderma 12: 201–206.
Rıs�noveanu, G. & A. V�adineanu, 2000. Evaluarea rolului
asociatiilor si populatiilor in functionarea sistemelor eco-
logice. Studiu de caz: oligochetele bentonice din Dunarea
inferioara si Delta Dunarii. Ars-Docendi, Bucharest.
Rıs�noveanu, G., G. Ignat, A. V�adineanu, S. Cristofor, C. Ci-
ubuc, G. Nafornita & M. Popescu, 1997. The state of the
benthic community of the Danube Delta lakes – a conse-
quence of eutrophication. Revue Roumaine de Biologie –
Biologie Animale 42: 227–235.
Rıs�noveanu, G., C. Postolache & A. V�adineanu, 2001. Nitro-
gen and phosphorous excretion rates by Tubificids from the
Danube River. In Weber, J. et al. (eds), Biogeochemical
Processes and Cycling of Elements in the Environment.
Wroclaw, Poland: 205–206.
Rıs�noveanu, G., A. V�adineanu & G. Ignat, 1998. The structure
of the benthic oligochaeta communities in the Danube Delta
lakes. Revue Roumaine de Biologie – Biologie Animale 44:
1–12.
Sakamoto, M., H. Hayashi, A. Otsiki, K. Aoyama, Y. Wa-
tanabe, T. Hanazato, T. Iwakuma & M. Yasuno, 1989. Role
of bottom sediments in sustaining plankton production in a
lake ecosystem. Experimental demonstration using enclosed
water bodies in a shallow eutrophic lake. Ecological Re-
search 4: 1–16.
Sarbu, A., 1997. Rolul macrofitelor submerse in circuitul unor
elemente minerale in ecosistemele acvatice din Delta Dun�a-
rii. Ph.D. Thesis, University of Bucharest.
Tatrai, I., 1987. The role of fish and benthos in the nitrogen
budget of Lake Balaton, Hungary. Archiv fur Hydrobiolgie
110: 291–302.
V�adineanu, A. & S. Cristofor, 1987. L’evolution de l’etat
trophique des ecosystems aquatiques caracteristiques du
Delta du Danube. 1. Le regime hidrologique, transparence
Secchi et la reserve de phosphore et d’azote. Revue Rou-
maine de Biologie – Biologie Animale 32: 83–91.
V�adineanu, A., Hiel’ekueta, S. Cristofor & I. Rugea, 1985.
Consideratii asupra dinamicii carbonului organic particulat
in unele ecosisteme acvatice ale Deltei Dunarii. Delta Du-
narii - Studii si comunicari de ecologie, Tulcea: 155–167.
V�adineanu, A., N. Botnariuc, S. Cristofor, G. Ignat & C.
Dorobantu, 1989. Tranzitii ale starii trofice a ecosistemelor
acvatice din Delta Dun�arii ın perioada 1981–1987. Ocrotirea
Naturii si Mediului Inconjurator 33: 27–34.
V�adineanu, A., S. Cristofor, G. Ignat, C. Ciubuc, G. Rıs�nov-eanu, F. Bodescu & N. Botnariuc, 2002. Structural and
functional changes within the benthic communities from
Danube Delta Lakes, Verhandlungen. Proceedings of
International Association of Theoretical and Applied Lim-
nology 27: 2571–2576.
Vollenweider, R. A., 1969. Sampling techniques and methods
for estimating quantity and quality of phytoplankton. In
Vollenweider, R. A., J. F. Talling & D. F. Westlake (eds), A
Manual on Methods for Measuring Primary Production in
Aquatic Environments Including a Chapter on Bacteria.
Blackwell Scientific Publishers, Oxford: 4–40.
Whigham, D. F., C. Chitterling & B. Palmer, 1988. Impacts of
freshwater wetlands on water quality: a landscape perspec-
tive. Environmental Management 12: 663–671.
202