ecological risk assessment (era) and hormesis
TRANSCRIPT

The Science of the Total Environment 288(2002) 131–140
0048-9697/02/$ - see front matter� 2002 Elsevier Science B.V. All rights reserved.PII: S0048-9697Ž01.01120-2
Ecological risk assessment(ERA) and hormesis
Peter M. Chapman*
EVS Environment Consultants, 195 Pemberton Avenue, North Vancouver, BC, Canada V7P 2R4
Received 24 May 2001; accepted 31 October 2001
Abstract
Based on our current state of knowledge, the significance and importance of hormesis is likely to be greater forecotoxicology, a component of ecological risk assessment(ERA), than for the overall process of ERA. Appropriatelydetermining the role of hormesis in ERA will require extension of hormesis beyond chemical stressors to abiotic(e.g.habitat) and biotic stressors(e.g. species introductions, organism interactions). It will also require determining for allstressors whether at both individual and higher levels of organization, hormesis has positive, neutral or adverseeffects. This determination must be made for model organisms, populations and communities. Adverse effects are theleast likely, however, neutral effects cannot be ruled out. Presently, consideration of hormetic effects in ERA is mostappropriate in a detailed level ecological risk assessment(DLERA), the most complex form of ERA. It is notappropriate in either problem formulation or a screening level ERA(SLERA). Further, for hormetic effects to berecognized and accepted fully into ERA may require a paradigm shift. Three on-going paradigm shifts to whichhormesis could be linked are: recognition of the low utility of no-observed effects concentrations(NOECs);recognition of the need for special treatment of essential element doseyconcentration-responses, which are similar tohormetic responses; and, the replacement of environmental toxicology with ecological toxicology(ecotoxicology). �2002 Elsevier Science B.V. All rights reserved.
Keywords: Hormesis; Subsidy–stress; Ecological risk assessment; Ecotoxicology; Exposure–response relationships
1. Introduction
Hormetic responses have been reported in hun-dreds of studies for a broad range of species(protozoa, bacteria, fungi, plants, invertebrates,and vertebrates including humans), biological end-points (e.g. survival, growth, reproduction), andboth inorganic and organic chemicals(Calabreseand Baldwin, 1997). Despite this, it has had a
*Corresponding author. Tel.:q1-604-986-4331; fax:q1-604-662-8548.
E-mail address: [email protected](P.M. Chapman).
long, but not always distinguished history(Cala-brese and Baldwin, 2000). It has too often beenassociated with homeopathy, marginalized, andyorignored. However, perhaps because of its notalways distinguished history, its potential roles inecological risk assessment(ERA) remain to bedetermined. The purpose of this paper is to buildon the findings of Chapman(1998, 1999, 2001a)and explore potential future roles that hormesiscould legitimately and usefully play in ERA andtwo of its key components, ecotoxicology andecology.

132 P.M. Chapman / The Science of the Total Environment 288 (2002) 131–140
2. Terms: hormesis, ecotoxicology, ecology andera
2.1. Hormesis
Hormesis can be generally defined as a stimu-latory effect(e.g. an increase in growth or fecund-ity) that typically extends 30–60% above controllevels, and over a 10-fold range below the no-observed-effects-concentration(NOEC). Hormesishas been hypothesized to be overcompensation toan alteration in homeostasis(Stebbing, 1998,2000). In other words, under low stress, organismsnot only repair any damage, but also overcompen-sate and reduce background damage more effec-tively. A formal definition has been proposed byCalabrese and Baldwin(2002): ‘an adaptiveresponse characterized by biphasic dose responsesof generally similar quantitative features withrespect to amplitude and range of the stimulatoryresponse that are either directly induced or theresult of compensatory biological processes fol-lowing an initial disruption of homeostasis’.Whether hormesis results in beneficial or harmfuleffects is, appropriately, not part of this definition.
2.2. Toxicology
Toxicology involves exposing an organism orbiological system to a stressor and determiningeither a response(e.g. toxicity) andyor in the caseof a chemical, uptake of that chemical into biolog-ical tissues(bioaccumulation). Ecotoxicology isbest defined in functional terms; it is distinguishedfrom environmental toxicology by Chapman(2001b). Ecotoxicology involves laboratory orfield testing that focuses on organisms importantin the food chainycommunity structure and func-tion. It is not restricted to individual species tests,and is not solely concerned with chemical effects,but may also include consideration of potentiallymore important stressors such as habitat loss andspecies introductions. Testing is conducted by tox-icologists but with appropriate input from ecolo-gists and, in some cases, other specializeddisciplines.
2.3. Ecology
Ecology is a scientific discipline, focusing on:interactions between organisms; distributions andabundances of organisms; the functioning of bio-logical populations and communities; and the proc-esses that affect all of these parameters(Andrewartha and Birch, 1954). Initially, ecologi-cal studies were concerned with elucidating naturalrelationships; however, as anthropogenic influenc-es on natural communities have increased, so haveecological studies expanded to include both naturaland anthropogenic changes to populations andcommunities. Typically, ecology has been primar-ily concerned with low dose effects, while toxi-cology has been primarily concerned with highdose effects.
2.4. Ecological risk assessment (ERA)
Ecological risk assessment is an iterative proc-ess, a framework providing a basis for eventualrisk management. It typically involves three tiers:problem formulation or hazard assessment(initialplanning and information gathering); effects andexposure assessment(data gathering and analysis);and risk characterization(assimilation and integra-tion). ERA differs from human health risk assess-ment (HHRA) based on the fact that it mustconsider a very large number of species ratherthan a single species(human beings). Further,whereas a HHRA aims to protect individuals, anERA is not concerned with individuals with thepossible exception of individuals of endangeredspecies, but rather with populations and with eco-system process and function. An ERA can beperformed to two levels of detail: a screening levelERA (SLERA); or a detailed level ERA(DLERA)(Hill et al., 2000 — Table 1). Both(eco)toxicology and ecology are key componentsof ERA.
3. Hormesis in (eco)toxicology
Hormesis is still not fully understood by far toomany (eco)toxicologists. Most ignore it. Somewho do not, nevertheless misunderstand it andascribe anomalous concentration–response curves

133P.M. Chapman / The Science of the Total Environment 288 (2002) 131–140
Table 1Some generic differences: Screening Level Ecological RiskAssessment(SLERA) and Detailed Level Ecological RiskAssessment(DLERA)a
Parameter SLERA DLERA
Relative level of effort Low HighLevel of conservation High Decreased
(over- (reasonablyprotective) protective)
Level of uncertainty High DecreasedHazard quotients(HQs) Generic Site-specificExtrapolations Broad Limited
Adapted from Hill et al.(2000).a
to hormesis when this is not the case. For example,it is well-known that increasing water hardness(mgyl calcium carbonate) ameliorates the bioa-vailability and toxicity of most metals(CCME,1999). Toxicity testing involving elevated metalsconcentrations in hard water can result in expo-sure–response curves that appear to be hormetic.There may be no toxicity at the highest testexposures in very hard water. However, toxicitycan appear at intermediate exposures due to dilu-tion effects. Dilution of the highest test exposureswill decrease not only metals concentrations, butalso water hardness. If water hardness is decreasedsuch that it no longer ameliorates metals bioavail-ability, toxicity may occur. At lower exposures,metals may be diluted sufficiently that despiteeven lower water hardness, no toxicity will result.Hence, the appearance of a hormetic exposure–response curve over a segment of the exposurerange can be provided by extrinsic factors(e.g.water hardness modifying toxicity), rather than byintrinsic factors (i.e. attempts to maintainhomeostasis).
But there are also too many instances of actualhormesis that are not recorded because tests arenot conducted with a sufficiently low range ofexposures. However, convincing researchers andtoxicity testing laboratories to extend their testingrange downward is difficult for two reasons. First,there is a long tradition in toxicology of focusingonly on higher exposures. Second, and much moreimportant, adding additional test exposures haseconomic(cost) consequences, which are currentlydifficult to justify. Only if hormesis is shown to
have real significance that needs to be taken intoaccount in risk and other assessments as mandatedby regulatory agencies, will low exposure testingbecome a standard feature of ecotoxicology andthus of ERA.
To some extent, there is a circular logic involvedhere. For hormesis to become accepted, it needsto be demonstrated in tests by individual research-ers and laboratories, and not merely reported inliterature summaries. However, for such demon-strations to become common, publication of hor-metic curves in ecotoxicology papers must becomemore common. Yet hormetic curves are typicallynot published, and papers debating the importanceand relevance of hormesis are not common in theecological toxicity literature. One way to breakthis circle of inaction might be to develop a well-advertised web site where hormetic data can beentered and evaluated, and examples of differentexposure–response curves and advice on theirinterpretation, particularly in a regulatory context,can be obtained.
Biostatistical tools for dealing with hormesisalso need to be developed, and existing tools needto be more widely used. It is not widely appreci-ated that Van Ewijk and Hoekstra(1993) havepublished a regression-based method for determin-ing EC values at low exposures that incorporates50
hormesis. Hormesis has been shown to occur andhas been evaluated by some authors using thisequation(e.g. Muyssen and Janssen, 2001). Otherregression-based methodology has been proposedby Bailer and Oris(1997, 2000) and Bailer et al.(2000a,b). Unfortunately, most researchers andtoxicity testing laboratories do not use such regres-sion-based methodology, and too many are proba-bly not aware it exists. The tendency is to use ‘offthe shelf’ methodology, approved by the regulatoryagencies that mandate the vast majority of toxicitytest data. Again, until this changes hormesis willnot be recognized in most toxicity test results,even if low enough exposures are tested. Table 2outlines the differences required to move frompresent ecotoxicology testing which does notexplicitly recognize hormesis to future testingwhich does. Additional detail is provided by Bailer(2001).

134 P.M. Chapman / The Science of the Total Environment 288 (2002) 131–140
Table 2Implications of hormesis for ecotoxicology
No allowance for hormesis Allowance made for hormesis(present testing) (future testing?)
Exposure-response relationships assumed, Exposure–response relationships investigated,not investigated not assumed
High toxicity test exposures Both high and low toxicity test exposures
Few exposure concentrations, More exposure concentrations,regimented spacing appropriate statistical power and spacing
Pooling of controls and No pooling of controls orlower test exposures possible lower test exposures
Range finders to pick test exposure ranges Use of environmental fate modelsto pick test exposures
Assumption of monotonic No preconceived assumptionsor linear data pattern; or confining statistics, e.g. application ofstatistics used to force this pattern quadratic model to generalized linear model
(Kerr and Meador, 1996);use of regression-type models allowing forhormesis(Van Ewijk and Hoekstra, 1993;Bailer and Oris, 1997, 2000;Bailer et al., 2000a,b)
4. Ecology
Hormesis is basically simply a response tostress. The form of stress may make no differenceto the hormetic response, at least at the individuallevel. For example, hormetic responses by humansare well documented(Yerkes and Dodson, 1908;Revelle and Loftus, 1990) in response to lowlevels of non-chemical stress(e.g. deadlines, pres-entations, etc.).
However, although chemical hormesis has beendemonstrated, there is less evidence that physicalandyor biological stressors can also cause horme-sis. Such evidence is important as, arguably, theseecological(rather than toxicological) stressors aremore important overall to the health and persist-ence of populations and communities than arechemical stressors(with the obvious exception ofchemical ‘hot-spots’) (Chapman, 1995).
There is some evidence that hormesis may notbe just a generalized toxicological phenomenon,but also a generalized ecological phenomenon, forinstance the intermediate disturbance hypothesis inecology (Chapman, 1998, 1999) and other ana-logues (Gentile, 2000). The intermediate distur-
bance hypothesis proposed historically by Tansleyand Adamson(1925) and more recently by Paineand Vadas(1969), describes both natural biotic(e.g. predation, competition) and physical stressors(e.g. fires, extreme weather events) as necessaryto maintain ecosystems at optimum levels overlarge spatial and temporal scales. Specifically,species richness is predicted to be greatest follow-ing moderate levels of disturbance(O’Neill et al.,1986; Collins et al., 1995). However, the inter-mediate disturbance hypothesis does not appear toapply to all situations, possibly due to differentiallife-history features and responses to disturbance(Underwood, 2000).
Another hypothesis linking stimulatory effectswith low levels of stress, chemical or non-chemi-cal, is the subsidy–stress gradient(Odum, 1971;Knight, 1981, 1983). Subsidy–stress effects havebeen shown in relation to biotic interactions andhabitat changes(Hargrave, 1970; Cooper, 1973;Flint and Goldman, 1975).
Anthropogenic stress may also induce hormesisin natural communities. A good example is offeredby soil microbial communities exposed to heavymetal stress. Traditionally, it was assumed that a

135P.M. Chapman / The Science of the Total Environment 288 (2002) 131–140
linear relationship existed; diversity decreased asstress increased. Giller et al.(1998) insteadhypothesize from the available literature, that lowlevels of stress result in the predominance of afew competitive species with consequent lack ofdiversity. As stress increases, these species losetheir competitive advantage and diversity increas-es. At higher stress, progressive species extinctionsoccur, and diversity decreases.
Bartell (2000) addressed the question: ‘Areecosystems hormetic?’ He noted that ecosystem-level hormesis could involve key ecological proc-esses such as nutrient cycling, usage of energyinputs, and stability. He argued that hormesis maywell be occurring at the ecosystem level andoffered examples including the following: a prob-ability of increased phytoplankton production as afunction of copper exposure for coastal marineecosystems; increased species diversity resultingfrom habitat changes; and increased fish speciesrelated to eutrophication. He believes there isenough evidence to more fully explore the possi-bility of ecosystem-level hormesis and its possibleimplications.
However, Gentile and van der Schalie(2000)point out the difficulties inherent in evaluatingoutcomes from hormesis as beneficial or harmfulfor ecological systems. Specifically, they note thatan increase in nutrients in a nutrient-poor lake mayalter species composition(a negative for speciesthat are replaced) and also increase populations ofsport fish (a positive for those species and forhumans fishing them). Similarly, crayfish survivebetter at sub-lethal concentrations of zinc whenthis metal kills the fungus on their gills(Giesy etal., 1980). Thus hormetic responses may comprisedirect or indirect(secondary) effects resulting froma direct toxic effect.
Further, stimulation of a single life history trait,such as growth or fecundity, can at times beexplained by a trade-off between traits(Stearns,1989, 1992). A positive effect on one measuredlife history trait may in fact be at the expense ofanother life history trait, resulting in no net gainfor the organism. It is not uncommon with daphn-ids or other animals for exposure to a toxicantconcentration to increase clutch size(hormesis),but reproduction may be delayed or egg size
reduced(Stearns, 1992; Ebert, 1993, 1994). Asnoted by Giesy(2001), alterations in the timingor number of eggs, even if apparently positive(e.g. more eggs), can have negative repercussions(e.g. loss of phase with males, more but smallerand less ‘fit’ eggs).
To incorporate hormesis into ecology and fullyunderstand its significancewi.e. is it a significantpositive attribute, neutral(irrelevantytrivial), or asignificant negative attributex requires knowledgeof:
● Individual-level consequences of suborganismhormesis — including any trade-offs betweenlife history traits
● Population-level consequences of individualhormesis — including any trade-offs betweenorganisms
● Processes(abiotic, biotic) regulating populationsize and health — which may be affected(positively, negatively or not at all) by hormesis
● Minimum viable population size(and genericconstraints) — which must be maintained andwhich can be affected(again, positively, nega-tively or not at all) by hormesis.
Clearly, it is not possible to fully understand allof the above, in all cases, for all organisms. Thusactual data must be supplemented by modeling(e.g. Caswell, 2000; Wang and Smith, 2000).
5. Ecological risk assessment (ERA)
Risk assessments are conducted in a tieredmanner, with initial assessments(problem formu-lation, SLERA) tending to be most conservativeand thus, to have the highest level of uncertainty.Uncertainty is reduced in successive tiers. Gentileand van der Schalie(2000) note that ‘it is unlikelythat hormesis is a critical factor in most ecologicalrisk assessments, given the magnitude of otheruncertainties inherent in the process’(similar com-ments are provided by Giesy(2001)). They furtherargue that default adjustments for hormetic effectson chemical exposure are unlikely to be practical.
If Gentile and van der Schalie(2000) and Giesy(2001) are correct, hormesis is best incorporatedinto an ERA where realism will be highest, andwhere conservatism and uncertainty will be lowest.
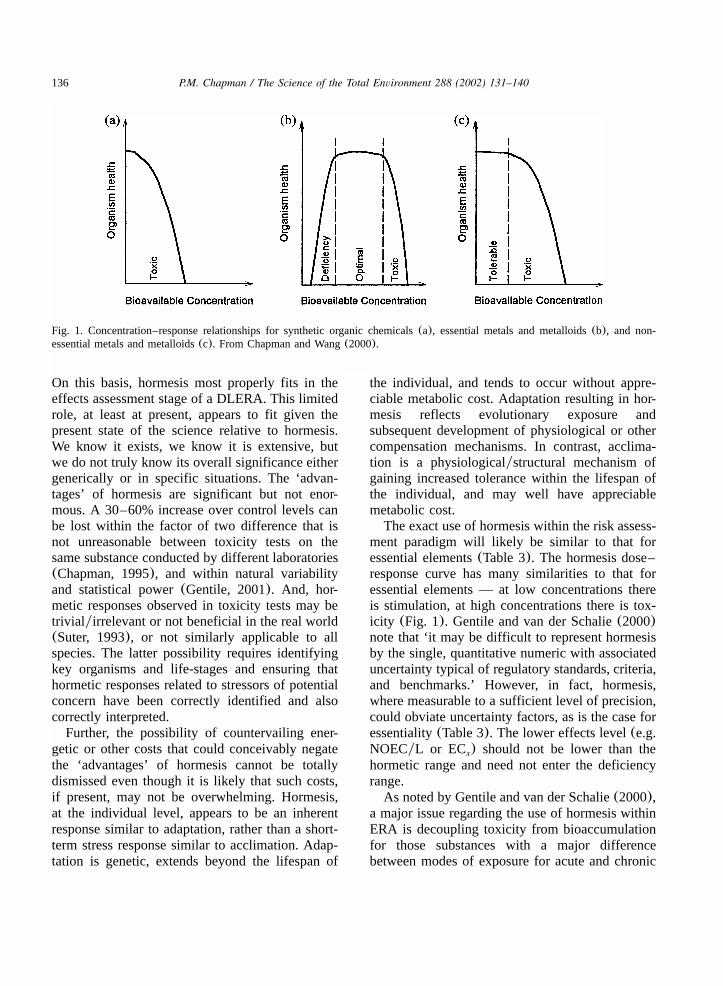
136 P.M. Chapman / The Science of the Total Environment 288 (2002) 131–140
Fig. 1. Concentration–response relationships for synthetic organic chemicals(a), essential metals and metalloids(b), and non-essential metals and metalloids(c). From Chapman and Wang(2000).
On this basis, hormesis most properly fits in theeffects assessment stage of a DLERA. This limitedrole, at least at present, appears to fit given thepresent state of the science relative to hormesis.We know it exists, we know it is extensive, butwe do not truly know its overall significance eithergenerically or in specific situations. The ‘advan-tages’ of hormesis are significant but not enor-mous. A 30–60% increase over control levels canbe lost within the factor of two difference that isnot unreasonable between toxicity tests on thesame substance conducted by different laboratories(Chapman, 1995), and within natural variabilityand statistical power(Gentile, 2001). And, hor-metic responses observed in toxicity tests may betrivialyirrelevant or not beneficial in the real world(Suter, 1993), or not similarly applicable to allspecies. The latter possibility requires identifyingkey organisms and life-stages and ensuring thathormetic responses related to stressors of potentialconcern have been correctly identified and alsocorrectly interpreted.
Further, the possibility of countervailing ener-getic or other costs that could conceivably negatethe ‘advantages’ of hormesis cannot be totallydismissed even though it is likely that such costs,if present, may not be overwhelming. Hormesis,at the individual level, appears to be an inherentresponse similar to adaptation, rather than a short-term stress response similar to acclimation. Adap-tation is genetic, extends beyond the lifespan of
the individual, and tends to occur without appre-ciable metabolic cost. Adaptation resulting in hor-mesis reflects evolutionary exposure andsubsequent development of physiological or othercompensation mechanisms. In contrast, acclima-tion is a physiologicalystructural mechanism ofgaining increased tolerance within the lifespan ofthe individual, and may well have appreciablemetabolic cost.
The exact use of hormesis within the risk assess-ment paradigm will likely be similar to that foressential elements(Table 3). The hormesis dose–response curve has many similarities to that foressential elements — at low concentrations thereis stimulation, at high concentrations there is tox-icity (Fig. 1). Gentile and van der Schalie(2000)note that ‘it may be difficult to represent hormesisby the single, quantitative numeric with associateduncertainty typical of regulatory standards, criteria,and benchmarks.’ However, in fact, hormesis,where measurable to a sufficient level of precision,could obviate uncertainty factors, as is the case foressentiality(Table 3). The lower effects level(e.g.NOECyL or EC ) should not be lower than thex
hormetic range and need not enter the deficiencyrange.
As noted by Gentile and van der Schalie(2000),a major issue regarding the use of hormesis withinERA is decoupling toxicity from bioaccumulationfor those substances with a major differencebetween modes of exposure for acute and chronic

137P.M. Chapman / The Science of the Total Environment 288 (2002) 131–140
Table 3Implications of hormesis for ERA; similar implications apply to the incorporation of essentiality(e.g. essential elements) into ERA
Step Present focus Future focus
Problem formulationy ●Few and generally high ●More exposures; both high andhazard assessment toxicity test exposures low exposure concentrations
Exposure ●Upper exposure bounds ●Distribution of actual dosesyassessment concentrations from exposure
(both upper andlower bounds possible)
Effects assessment ●Few and generally high ●More exposures; both high andtoxicity test exposures low exposure concentrations●Assumption of monotonicy ●No preconceived assumptionslinear data pattern (or confining statistics)●Estimate risk of exceeding ●Effects estimated bya threshold value exposure–response modeling
Risk characterization ●Application of ●No safety factors(if hormeticsafety factors thresholds can be determined)●Effect of a stressor on a ●Net effect of a stressor ondefined endpoint organisms and communities●Upper uncertainty bounds ●Upper and lower
uncertainty bounds●Likelihood of adverse ●Likelihood of adverse andeffects beneficial effects●Linear, no threshold ●No linear, no thresholddose–response dose–responsedefault assumptions default assumptions●Point estimates of ●Risk always characterizedrisk possible as a range
Risk management ●Chemical concentrations ●Chemical concentrations at orat or below background below hormetic level
toxicity. For most substances and many organisms,chronic and acute toxicity both result primarilyfrom water column exposures. However, for somesubstances such as the organic forms of mercuryand selenium, PCBs, DDT, chronic exposureresults primarily if not entirely from dietary uptake.And for some organisms(e.g. those burrowinginto contaminated sediments), dietary uptake canbe an extremely important uptake route not onlyfor these, but also for other substances. This issuemust be addressed in ERA, specifically by identi-fying stressor–response pathways that are mostlikely to be significantly affected by hormesis andthose (e.g. dietary uptake) for which hormeticresponses are inappropriate. As Gentile and vander Schalie(2000) point out, it would be inappro-priate to apply a toxicologically derived lower
effects level that allowed bioaccumulation to levelsof concern.
Because of the issues discussed above, Gentileand van der Schalie(2000) conclude that ‘hor-mesis will likely not be a very important factor inmost ecological risk assessments.’ As is the casewith most generalizations, this will not always betrue, and there will be instances where hormeticeffects result in a shift in ‘bright lines’ that moreappropriately reflect the balance between over- andunder-protection. The true need is to focus ERAefforts on those instances where this is truly thecase and not waste time or resources on trivialcases, which will solely denigrate hormesis as ageneral and acceptable scientific principle that isuseful for ERA.

138 P.M. Chapman / The Science of the Total Environment 288 (2002) 131–140
In fact, as noted by Menzie(2001), some ERAsdo incorporate hormetic phenomena directly foressential nutrients such as zinc, biodegradation ofpetroleum hydrocarbons, and natural phenomenasuch as periodic flooding and fire. Although themechanisms governing hormesis are different,ERAs and associated policies(i.e. risk manage-ment) recognizing these phenomena acknowledgethat (Menzie, 2001) ‘exposure and response arenot always directly related«less is not alwaysbetter and may in some cases be worse.’ For otherstressors generally considered harmful(e.g. cad-mium, mercury), environmental policies considerassimilative capacity or the ability of organisms towithstand a certain level of stress. In this casehormesis is considered implicitly rather thanexplicitly (Menzie, 2001).
An alternative, general DLERA scheme forhormetic substances could involve:
● Estimating the exposure range of selectedorganisms to stressors of potential concern(SOPCs) in appropriate habitatsycommunities.
● Establishing hormetic doseyconcentration–response curves.
● Determining the extent and range of hormesisto the selected organisms.
● Determining whether a given, low doseresponse has a positive, neutral or negativeeffect on the overall health of both individualorganisms and populationsycommunities.
● Factoring this information into the risk charac-terization (e.g. if effect is positive, baselineshould be maximal response, and if not, baselinewould revert to default: responses in unexposedorganisms or populationsycommunities).
6. Possible complementary paradigm shifts
Incorporation of hormesis into ERA may requirea paradigm shift. Given that the ‘advantages’ ofhormesis are not large, a paradigm shift based onhormesis alone appears unlikely. However, linkinghormesis to other phenomena in a ‘combinedparadigm shift’ offers good possibilities for mov-ing forward. There are at least three different suchphenomena to which hormesis could be linked.
First, a philosophical change in toxicology end-points is underway. The utility of no-observed-effect-concentrationsylevels (NOECyLs) is inquestion in scientific and regulatory arenas, forreasons explained by Chapman et al.(1996) andSuter (1996). There is strong pressure to movephilosophically from the idea of no effect, to theidea of a limited level of effectwe.g. a set effectsconcentration or EC , or a no-effects-concentrationx
(NEC) that does not need to equal zero — Koo-ijmann, 1996x. Thus, the International StandardsOrganization Working Group ISO TC147ySC5 —Water Quality, Biological Methods has agreed thatthe NOEC will be phased out of standards(Sep-tember 18–24, 2000 meeting, Antalya, Turkey).This philosophical shift mirrors that engenderedby hormesis in that total elimination of chemicalsis not sought, but rather the focus is on theirdecrease to levels where they are not an issue.
Second, increasing recognition that essential ele-ments cannot be treated similarly to non-essentialelements is also resulting in a philosophicalchange. It is increasingly being recognized that forsome chemicals, adverse effects occur not onlywith excess but also with deficiency. The regula-tory implication is that lower effects bounds, bethey NOEC or EC values, cannot be set belowx
deficiency levels. Similarly, the lower bounds forhormetic substances need not be set below hor-metic levels. This lower bounding of effects valueshas an important consequence; if the lower boundsare known, there is no need for uncertainty(i.e.safety) factors, instead the boundaries can be setmuch more precisely and without concern for over-or under-conservatism(Chapman et al., 1998).
Third, there is a continuing shift in the field oftoxicology from reductionist to more holisticapproaches. As noted by Chapman(2001b), envi-ronmental toxicology is giving way to ecologicaltoxicology. Rather than toxicologists being primar-ily concerned with getting toxicity tests doneaccording to preset formulae, the emphasis isshifting to ensuring that the tests are as ecologi-cally relevant as possible. Hormesis is a demon-strable, inherent phenomenon common to a verywide variety of organisms and endpoints. Whereasit can be ignored by environmental toxicology, it

139P.M. Chapman / The Science of the Total Environment 288 (2002) 131–140
cannot be ignored by ecotoxicology, which is akey component of ERA.
However, in the context of these potential par-adigm shifts, hormesis may not be the most appro-priate umbrella term for unusual exposure–response relationships at low doses(Menzie, 2001;Suter, 2001; Giesy, 2001). A change in terminol-ogy may be necessary both because hormesis isgenerally associated with single species adaptiveresponses, and because we cannot assume higher-level hormetic effects are mediated by the samemechanisms as lower-level effects.
7. Conclusions
The implications of hormesis for environmentalrisk assessment may be potentially profound: pres-ent techniques used to estimate the no-effect-concentration(NEC) may not be beneficial tobiota or the ecosystem. Effectively, some level ofstress may be desirable and the current emphasison zero discharge of any chemicals may well beless than beneficial to at least some if not manyorganisms. However, the profundity of these impli-cations requires non-trivial changes in both ourphilosophical and methodological approaches toecological risk assessment. This will not be aneasy process. It will require addressing two keyissues: first, defining exposure–response relation-ships within the range of real world exposures;and second, understanding the mechanistic basisybases for hormesis(Suter, 2001; Bailer, 2001;Giesy, 2001; Gentile, 2001) to allow predictionsof hormetic responses. It may also require a changein terminology, specifically development of a suit-able umbrella term for low dose exposure-responses that include single-species hormeticresponses.
Acknowledgments
I thank Ed Calabrese for encouraging me towrite an earlier version of this paper(Chapman,2001a) and Bill van der Schalie for encouragingme to expand it. I also thank the seven individualswho commented on Chapman(2001a) in theBELLE Newsletter. This expansion of my originalpaper benefited greatly from their insightful com-
ments. Further improvements were provided by ananonymous referee. Final word processing wasdone by Jackie Gelling (EVS EnvironmentConsultants).
References
Andrewartha HG, Birch LC. The distribution and abundanceof animals. Chicago, IL, USA: University of Chicago Press,1954.
Bailer AJ. Experiments, analyses and decisions: hormesis inecotoxicology. BELLE Newslett 2001;10(1):9–11.
Bailer AJ, Oris JT. Estimating inhibition concentrations fordifferent response scales using generalized linear models.Environ Toxicol Chem 1997;16:1554–1559.
Bailer AJ, Oris JT. Defining the baseline for inhibition con-centration calculations for hormetic hazards. J Appl Toxicol2000;20:121–125.
Bailer AJ, Hughes MR, Denton D, Oris JT. An empiricalcomparison of effective concentration estimators for evalu-ating aquatic toxicity test responses. Environ Toxicol Chem2000a;19:141–150.
Bailer AJ, Elmore RT, Shumate BJ, Oris JT. A simulationstudy of characteristics of statistical estimators of inhibitionconcentrations. Environ Toxicol Chem 2000b;19:3068–3073.
Bartell SM. Are ecosystems hormetic? Hum Ecol Risk Assess2000;6:237–243.
Calabrese EJ, Baldwin LA. The dose determines the stimula-tion (and poison): development of a chemical hormesisdatabase. Int J Toxicol 1997;16:545–559.
Calabrese EJ, Baldwin LA. History of chemical hormesis.Hum Exp Toxicol 2000;19:2–31.
Calabrese EJ, Baldwin LA. Defining hormesis. BELLE News-lett 2002;11(in press).
Caswell H. Matrix population models, 2nd ed. Sunderland,MA, USA: Sinauer Associates, 2000.
CCME. Canadian environmental quality guidelines. Winnipeg,MN: Canadian Council of Ministers of the Environment,1999.
Chapman PM. Ecotoxicology and pollution — key issues. MarPollut Bull 1995;31:167–177.
Chapman PM. New and emerging issues in ecotoxicology —the shape of testing to come? Australas J Ecotoxicol1998;4:1–7.
Chapman PM. Whole effluent toxicity testing — usefulness,level of protection, and risk assessment. Environ ToxicolChem 1999;19:3–13.
Chapman PM. The implications of hormesis to ecotoxicologyand ecological risk assessment(ERA). BELLE Newslett2001a;10(1):2–9.
Chapman PM. Integrating toxicology and benthic ecology:Putting the eco back into ecotoxicology. Mar Pollut Bull2001b;(in press).

140 P.M. Chapman / The Science of the Total Environment 288 (2002) 131–140
Chapman PM, Wang F. Issues in ecological risk assessment ofmetals and metalloids. Hum Ecol Risk Assess 2000;6:965–988.
Chapman PM, Cardwell RS, Chapman PF. A warning: NOECsare inappropriate for regulatory use. Environ Toxicol Chem1996;15:77–79.
Chapman PM, Fairbrother A, Brown D. A critical evaluationof safety(uncertainty) factors for ecological risk assessment.Environ Toxicol Chem 1998;17:99–108.
Collins SL, Glenn SM, Gibson DJ. Experimental analysis ofintermediate disturbance and initial floristic composition:decoupling cause and effect. Ecology 1995;76:486–492.
Cooper DC. Enhancement of net primary productivity byherbivore grazing in aquatic laboratory microcosms. LimnolOceanogr 1973;18:31–37.
Ebert D. The trade-off between offspring size and number inDaphnia magna — The influence of genetic, environmentaland maternal effects. Arch Fur Hydrobiol(Suppl.)1993;90:453–473.
Ebert D. A maturation size threshold and phenotypic plasticityof age and size at maturity inDaphnia magna. Oikos1994;69:309–317.
Flint RW, Goldman CR. The effects of a benthic grazer onthe primary productivity of the littoral zone of Lake Tahoe.Limnol Oceanogr 1975;20:935–944.
Gentile JH. Hormesis in ecology and ecological assessments.Hum Ecol Risk Assess 2000;6:223–226.
Gentile JH. The implications of hormesis to ecotoxicology andecological risk assessment(ERA). BELLE Newslett2001;10(1):12–13.
Gentile JH, van der Schalie WH. Hormesis and ecological riskassessment: fact or fantasy? Hum Ecol Risk Assess2000;6:227–236.
Giesy JP. Hormesis — does it have relevance at the population-, community- or ecosystem-levels of organization? BELLENewslett 2001;10(1):14–17.
Giesy JP, Bowling JW, Kania HJ. Cadmium and zinc accu-mulation and elimination by freshwater crayfish. Arch Envi-ron Contam Toxicol 1980;9:683–697.
Giller KE, Witter E, McGrath SP. Toxicity of heavy metals tomicroorganisms and microbial processes in agricultural soils:a review. Soil Biol Biochem 1998;30:1389–1414.
Hargrave BT. The effect of a deposit-feeding amphipod on themetabolism of benthic microflora. Limnol Oceanogr1970;15:21–30.
Hill RA, Chapman PM, Mann GS, Lawrence GS. Level ofdetail in ecological risk assessment. Mar Pollut Bull2000;40:471–477.
Kerr DR, Meador JP. Modelling dose response using general-ized linear models. Environ Toxicol Chem 1996;15:395–401.
Knight RL. A control hypothesis for ecosystems — energeticsand quantification with the toxic metal cadmium. In: MitschWJ, Bosserman RW, Klopatek JM, editors. Energy andecological modeling. Developments in environmental mod-
elling. Amsterdam: Elsevier Scientific Publishing Co,1981:601–615.
Knight RL. Energy basis of ecosystem control at SilverSprings, Florida. In: Fontaine TD, Bartell SM, editors.Dynamics of lotic ecosystems. Ann Arbor, MI, USA: AnnArbor Science, 1983:161–179.
Kooijmann SALM. An alternative for NOEC exists, but thestandard model has to be abandoned first. Oikos1996;75:310–316.
Menzie CA. Hormesis in ecological risk assessment: a usefulconcept, a confusing term, andyor a distraction? BELLENewslett 2001;10(1):17–19.
Muyssen BTA, Janssen CR. Zinc acclimation and its effect onthe zinc tolerance ofRaphidocelis subcapitata andChlorellavulgaris in laboratory experiments. Chemosphere2001;45:507–514.
Odum EP. Fundamentals of ecology. Philadelphia, PA, USA:W.B. Saunders Co, 1971.
O’Neill RV, DeAngelis DL, Waide JB, Allen TFH. A hierar-chical concept of ecosystems. Princeton, NJ, USA: PrincetonUniversity Press, 1986.
Paine RT, Vadas RL. The effects of grazing by sea urchins,Strongylocentrotus spp. on benthic algal populations. LimnolOceanogr 1969;14:710–719.
Revelle W, Loftus DA. Individual differences and arousal:implications for the study of mood and memory. CognEmotion 1990;4:209–237.
Stearns SC. Trade-offs in life history evolution. Funct Ecol1989;3:259–268.
Stearns SC. The evolution of life histories. New York: OxfordUniversity Press, 1992.
Stebbing ARD. A theory for growth hormesis. Mutat Res1998;403:249–258.
Stebbing ARD. Maia hypothesis — growth control and toxi-cology. Hum Ecol Risk Assess 2000;6:301–311.
Suter GW. Ecological risk assessment. Chelsea, MI, USA:Lewis Publishers, 1993.
Suter GW. Abuse of hypothesis testing in ecological riskassessment. Hum Ecol Risk Assess 1996;2:331–349.
Suter GW. Focus on exposure–response relationships, andcomplex forms will come naturally. BELLE Newslett2001;10(1):22.
Tansley AG, Adamson RS. Studies on the vegetation of theEnglish chalk. III. The chalk grasslands of the Hampshire–Sussex border. J Ecol 1925;13:177–223.
Underwood AJ. Experimental ecology of rocky intertidalhabitats: what are we learning? J Exp Mar Biol Ecol2000;250:51–76.
Van Ewijk PH, Hoekstra JA. Calculation of the EC and its50
confidence interval when subtoxic stimulus is present. Eco-toxicol Environ Saf 1993;25:25–32.
Wang SCD, Smith EP. Adjusting for mortality effects inchronic toxicity: mixture model approach. Environ ToxicolChem 2000;19:204–209.
Yerkes RM, Dodson JD. The relationship of strength ofstimulus to rapidity of habit formation. J Comp NeurolPsychol 1908;18:459–482.