early seedling demography in balsam fir seedling banks
TRANSCRIPT

Early seedling demography in balsam firseedling banks
Robin Duchesneau and Hubert Morin
Abstract: Following the 1995 mast year, we conducted detailed seedling demography surveys to characterize someecological factors affecting germination and early establishment in understory balsam fir (Abies balsamea(L.) Mill.)seedling banks and verify if recruitment from a cohort can potentially renew the balsam fir seedling bank. Twenty-fiveseed traps paired with 32 demography subplots were located in each of four permanent plots and regularly monitoredover a 2-year period. Spatial patterns of newly emerged seedlings generally corresponded to those found for seeddispersal. However, at the end of the monitoring period, the spatial pattern of surviving seedlings was independent ofthe seed dispersal pattern. Seedbed types and understory light environments were the main factors explaining seedlingmortality. Sphagnum mosses and decaying wood were the most favorable seedbeds for early establishment. The 0- to25-cm seedling populations grew in size because of the addition of new recruits. If, in the years to come, mortalityrates of the 1995 cohorts were similar to those of already established 0- to 25-cm seedlings, mast years of seedproduction, at least during endemic periods, would be crucial for recruitment and renewal of balsam fir seedling banks.
Résumé: Suite à la forte année semencière de 1995, le suivi démographique d’une cohorte de semis de sapin baumier(Abies balsamea(L.) Mill.) a été effectué afin de caractériser quelques facteurs écologiques qui affectent lagermination et l’établissement initial dans les banques de semis de sapin baumier sous couvert et de vérifier si lerecrutement à partir d’une cohorte peut permettre le renouvellement des banques de semis. Vingt-cinq trappes à grainespairées à 32 quadrats démographiques ont été localisées dans chacune de quatre places échantillons permanentes, et unsuivi a été assuré régulièrement pendant 2 ans. De façon générale, la distribution des graines correspondait aux patronsd’abondance spatiaux des nouveaux émergents. Toutefois, à la fin de la période d’étude, les patrons d’abondancespatiaux des semis ayant survécu ne correspondaient plus à ceux de la distribution des graines. La nature des lits degermination et l’environnement lumineux se sont avérés les critères principaux selon lesquels on pouvait expliquer lamortalité des semis. La sphaigne et le bois en décomposition constituaient les lits de germination les plus favorables àl’établissement initial. Les populations de semis de 0 à 25 cm ontaugmenté suite à l’addition de nouvelles recrues. Si,dans les années à venir, le taux de mortalité descohortes de 1995 se comportait de façon similaire à celui des semis de0 à 25 cm déjà installés, les fortes années semencières, du moins pendant les périodes endémiques, seraient crucialespour assurer un recrutement et un renouvellement des banques de semis du sapin baumier.Duchesneau and Morin
1509
Introduction
Research carried out in balsam fir forests (Abies balsama(L.) Mill.) in New Brunswick (Morris 1963; MacLean 1984;Baskerville 1975), Ontario (Blais 1954; Ghent et al. 1957),and Quebec’s northern boreal forests (Morin 1990, 1994)have corroborated Baskerville’s (1975) hypothesis that sprucebudworm (Choristoneura fumiferana(Clem.)) outbreaks andbalsam fir forests often form a relatively interdependentautoregulating system. In this type of long-term cyclical pat-tern, spruce budworm outbreaks are natural disturbances thatfavor the growth of pre-established natural regeneration fol-lowing the defoliation of mature firs (MacLean 1988). Theregenerative success of balsam fir following this type of re-curring disturbance depends primarily on its seedling bank.As stated, fir seedlings are capable of avoiding damagecaused during outbreaks (Osawa 1994) and surviving under
the canopy for long periods without losing their ability forrapid growth following gap openings (Ruel and Huot 1993)and light availability (Parent and Messier 1995).
A recent study on the seedling bank dynamics of balsamfir demonstrated a great similarity of the seedling age struc-tures throughout 14 stands of various ages (Morin andLaprise 1997). Two hypotheses were proposed to explain theseedling age structure: (i) turnover where the seedling bankis always being renewed, i.e., young seedlings readily re-place older dying ones, and (ii ) the bank is established andgrows to a certain point then stagnates. It is also possiblethat a balsam fir seedling bank builds up rapidly during a20- to 30-year period and that the availability of seedling re-generation niches quickly becomes limiting (Ghent 1958).Osawa (1994) has shown that a lack of seedlings in youngerage-classes could be related to the impact of the last sprucebudworm outbreak on cone and consequently seed produc-tion. It is also likely that the phenomenon of growth stagna-tion plays a more important role during outbreak periodswhile the seedling bank relies on renewal in endemic peri-ods (Morin and Laprise 1997).
To understand the seedling bank dynamics of balsam fir,factors that determine recruitment must be identified. Harper(1977) and Grubb (1977), among others, have suggested that
Can. J. For. Res.29: 1502–1509 (1999) © 1999 NRC Canada
1502
Received August 20, 1998. Accepted March 28, 1999.
R. Duchesneau and H. Morin.1 Département des sciencesfondamentales, Université du Québec à Chicoutimi,Chicoutimi, QC G7H 2B1, Canada.
1Corresponding author. e-mail: [email protected]
I:\cjfr\cjfr29\cjfr-10\X99-090.vpThursday, October 28, 1999 4:05:11 PM
Color profile: DisabledComposite Default screen

the events that control recruitment usually occur during theperiod that encompasses seed dispersal, germination, andseedling establishment. Other authors attempting to synthe-size the functional aspects of recruitment in different plantpopulations have concluded that dispersal and recruitmentare complex processes (Schupp and Fuentes 1995; Houle1995). Year-to-year variation in temperature, rainfall, andseed production; instability in microsite suitability; and in-terdependency of factors limiting recruitment are some ofthe difficulties that may be encountered while examining re-cruitment in seedling populations (De Steven 1991; Erikssonand Ehrlen 1992). Part of the problem is the difficulty indetermining proper spatiotemporal scales of observation(Schupp and Fuentes 1995). On the other hand, Houle(1995) has made it clear that, to understand how patterns ofrecruitment develop through time, it is essential to considerthe demographic consequences of seed dispersal on allstages from seed to new adult.
Since Harper et al. (1961) developed the concept of “safesites,” many authors have tried to identify the main ecologi-cal factors that influence germination and early establish-ment. Of the factors examined, the density of neighbors(Fowler 1988; Taylor and Aarssen 1989), the availability ofseedbeds (Groot 1988; Oswald and Neuenschwander 1993;Fleming and Mossa 1994; Szewczyk and Szwargrzyk 1996),abiotic conditions (Houle 1992), predation (Schupp 1988;Whelan et al. 1990), and light environment (Li and Burton1994) are amongst the most important.
In the present study, the spatial and temporal dynamics ofgermination and early establishment are examined for balsamfir seedling banks in a boreal fir forest found in the northernzone of Quebec. More specifically: (i) we examine whetherthere is a correspondence between spatial patterns of seedabundance and spatial distribution of emergents and seedlingshaving survived two seasons post-emergence, and (ii ) weidentify safe sites for germination and early establishment withrespect to seedbeds and the understory light environment.
In spring, the forest floor in our study sites is generallyvery moist. Furthermore, our sites have a high density ofseed trees. For these reasons, we hypothesized that the spa-tial pattern of newly emerged seedlings was directly relatedto the spatial distribution of seedfall. We also hypothesizedthat seedlings emerging on different seedbeds and, in differ-ent understory light conditions, had different survival proba-bilities for early establishment.
Study area
This study was conducted in four permanent sample plots lo-cated near the 50th parallel (72°35′; 49°48′), about 100 km north of
Lac Saint-Jean, Quebec. The study area is in Thibault’s (1987)black spruce (Picea mariana(Mill.) BSP) – moss ecological region(No. 12b) of the boreal zone. Some balsam fir stands are found inmountainous regions and were greatly damaged by the last sprucebudworm outbreaks. Balsam fir forests of the region are composedof a mosaic of patches of different ages dating back to one of thelast three outbreaks that occurred during the 19th and 20th centu-ries (Morin 1994). Good seed crops occur at intervals of 2–4 years,with very low or no seed production during intervening years (Mor-ris 1951; Ghent 1958). But during any one year a great number ofseeds may be empty because of inadequate pollination (Morris 1951).
The chronology of spruce budworm outbreaks established bydendrochronological techniques indicates that the last defoliationstarted around 1974 and that growth reduction lasted until the late1980s (Morin and Laprise 1990). This result is in good agreementwith detailed aerial defoliation surveys, which indicate that the de-foliation started in 1974 in all the study areas and that it lasted 2–6years. Each stand had been greatly damaged by the last sprucebudworm outbreak. The vegetation association is similar to the bal-sam fir – white birch (Betula payriferaMarsh.) type of the moresouthern balsam fir – white birch zone (Grandtner 1966). Themean annual temperature is –0.7°C ranging from –19°C in Januaryto 15.8°C in July, and the mean annual precipitation is421.7 cmwith 356.6 cm falling as snow (Environment Canada 1992).
The balsam fir stands in this study have never been logged orprotected from spruce budworm defoliation by spraying treat-ments. Sample plots were established in stands with different agestructures. These age structures were previously determined ac-cording to age distribution, growth curves, and the ecological rela-tionship between those two parameters and the spruce budwormoutbreaks (Morin 1994). One 20 × 20 m quadrat was located in aneven-aged unimodal stand that experienced an outbreak from 1944to 1953 (site 1B), and two others were located in stands that wereattacked during the 1909–1923 outbreak (sites 2E and 4B). Thefourth quadrat was located in a stand with a bimodal age structureassociated with both the 1909–1923 and 1944–1953 outbreaks (site4A) (Table 1).
Methods
Caveats and termsIn this study, we use the term “seed dispersal” in a broad sense
to denote both a process (movement of seeds) and an outcome(pattern of seed deposition). The term “germination” is commonlymeasured as the number of seedlings emerging from the soil sur-face. The term “emergents” refers to the newly emerged seedlings,and “early established” is used for those having survived to the endof the second growing season. We use the terms “best seedbed”and “worst seedbed” to describe a seedbed in which the probabili-ties of germination and of seedling survival were found to be high-est and lowest, respectively. Since this study was part of a long-term project, the choice of labels for stand identification (sites 4A,2E, 4B, and 1B) was not arbitrary. They represent the same standsas those previously studied by Morin and Laprise (1997).
© 1999 NRC Canada
Duchesneau and Morin 1503
Site 4A Site 2E Site 4B Site 1B
Establishment year 1922 1841 1883 1934Spruce budworm outbreaks at the origin of
the populations1909–19231944–1953
1909–1923 1909–1923 1944–1953
Age distribution Bimodal Unimodal Unimodal UnimodalTotal density (stems/ha) 7600 1675 2475 17 150Balsam fir density (stems/ha) 7300 1475 2325 16 400Balsam fir basal area (% living + dead) 80 86 76 91
Table 1. Ecological characteristics of the four study sites (modified from Morin and Laprise 1997).
I:\cjfr\cjfr29\cjfr-10\X99-090.vpThursday, October 28, 1999 4:05:12 PM
Color profile: DisabledComposite Default screen
Seed rainTwenty-five seed traps were systematically established at 5-m
intervals in each 20 × 20 m quadrat. The traps were constructed bycovering galvanized steel pails (23.5 cm in diameter) with 2-cmwire mesh for protection against seed predation. To cover the au-tumn 1994 to spring 1995 seed dissemination period, the trapswere emptied and seeds counted four times (September 22, Octo-ber 6 and 20, 1994, and June 5, 1995). Since it is generally ac-knowledged that balsam fir does not maintain a persistent soil seedbank (Thomas and Wein 1985; Houle and Payette 1991), seedfallin this period represents the sum of propagules potentially capableof producing the emergents of the seedling cohort of 1995.
Seedling emergence and survivalThirty-two 60 × 60 cm demography subplots were systemati-
cally established in staggered rows in each 400-m2 quadrat. De-mography subplots were placed within 1 m of the closest seed trap.Demography subplots were surveyed every 2 weeks from earlyJune to late November in both 1995 and 1996. Percent seedlingsurvival for the 1995 seedling cohort was calculated as the ratiobetween seedling number and the number still alive at the end ofthe sampling. Total number of 0- to 25-cm balsam fir seedlingswas also counted.
All seedlings were mapped and the associated seedbed was re-corded (Table 2). The total surface coverage of different seedbedswas also determined for each subplot. The seedbeds found in ourstudy sites were litter, decaying and nondecaying wood, feathermosses on litter, feather mosses on decaying and on nondecayingwood, and also sphagnum moss on litter and on decaying andnondecaying wood. Decaying wood was defined as wood thatcould be easily penetrated with a finger to a depth of 3–5 cm.
Light environmentThe light environment was assessed by the overcast sky method
described in Messier and Puttonen (1995) and Parent and Messier(1996). Instantaneous measurements (Qo) were taken at the centerof each demography subplot at a height of 15 cm aboveground.These measurements were conducted with a LI-COR LI-190SBquantum sensor under completely overcast sky conditions. A sec-ond sensor taking continuous measurements with a data logger wasplaced in the center of an adjacent clearcut to record full light con-ditions (Qi). Percent photosynthetic photon flux density (%PPFD =(Qo/Qi) × 100) represents mean daily light (in the wave band of
400–700 nm) penetration under the forest canopy. Measurementswere made in mid-August.
Statistical analysisThe spatial distribution of seedfall was determined using
Moran’s I coefficient (Legendre and Fortin 1989). Moran’sI is ameasure of spatial autocorrelation and generally behaves like aPearson’s correlation coefficient; its value ranges from –1 to 1,where the expected value in the absence of significant spatialautocorrelation is around zero (Oden and Sokal 1986). The dis-tance classes were 5 m.
Relationships between the spatial patterns of seed abundanceand the spatial distribution of emergents and of 2-year-old seed-lings, at the demography subplot scale, were tested using Pearson’scorrelation coefficients. Seed abundance in subplots was deter-mined from seed counts of the closest seed trap.
To assess the relative quality of seedbeds for germination,Gtests of goodness of fit for single-classification frequency distribu-tions followed by simultaneous test procedures (STP) were per-formed (Sokal and Rohlf 1981). The expected frequencies, basedon a hypothesis extrinsic to the data, were determined by evaluat-ing the number of seeds estimated to have fallen on the total sur-face of each seedbed. This was done by using the number of seedsfound in the closest trap to a demography subplot and by estimat-ing how many seeds had fallen per seedbed surface in that subplot.Then, we summed these seed estimates for the total surface of eachseedbed found in all demography subplots. The sensitivity and pre-cision of this approach depends on how well seed abundance fromtraps related to the estimates of seed abundance for the differentseedbed surfaces.
G tests were also performed to assess the relative importance ofseedbeds in the establishment of seedlings. Distributions used inthe analysis represented the number of seedlings having emergedand survived on each seedbed. When significant heterogeneityamong replicates (seedbeds) was found, the STP procedure wasused to group seedbeds.
All data analyses were performed with SYSTAT version 5.2.1software (SYSTAT, Inc. 1992) except for spatial autocorrelation,which was done withLe progiciel R(Legendre and Vaudor 1991).
Results
Seedling banks had three different understory light envi-ronments: one relatively closed canopy site (1B, 2.83%
© 1999 NRC Canada
1504 Can. J. For. Res. Vol. 29, 1999
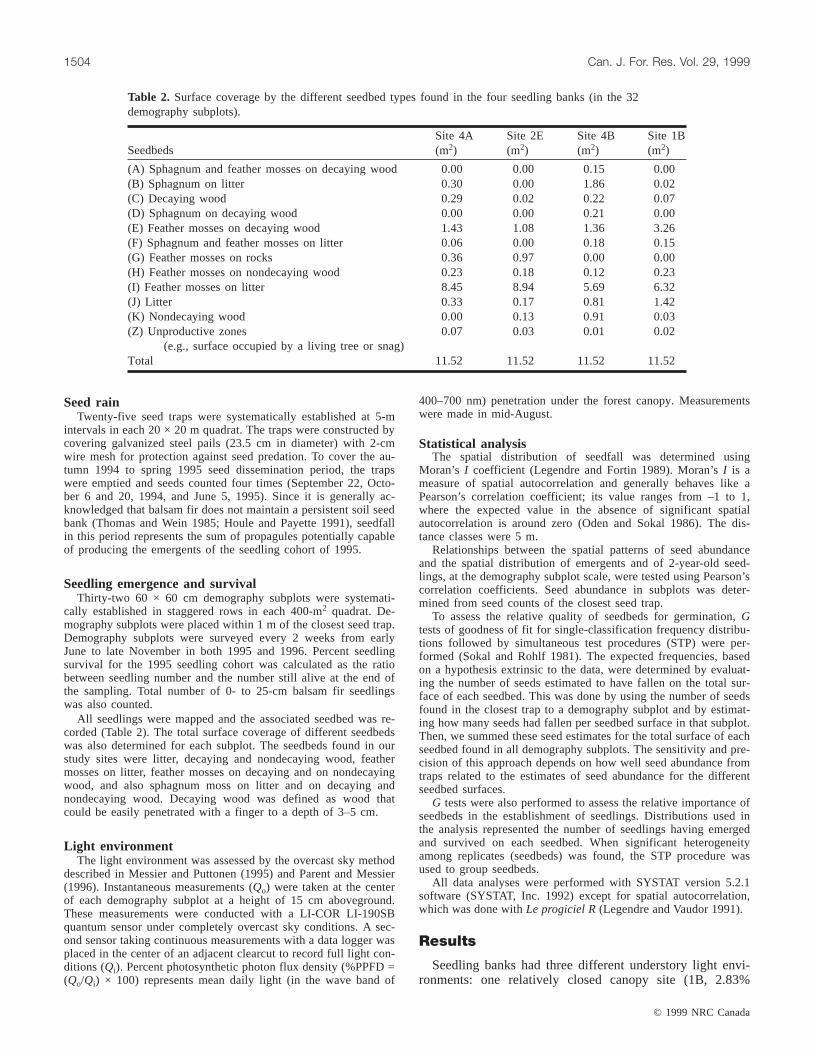
SeedbedsSite 4A(m2)
Site 2E(m2)
Site 4B(m2)
Site 1B(m2)
(A) Sphagnum and feather mosses on decaying wood 0.00 0.00 0.15 0.00(B) Sphagnum on litter 0.30 0.00 1.86 0.02(C) Decaying wood 0.29 0.02 0.22 0.07(D) Sphagnum on decaying wood 0.00 0.00 0.21 0.00(E) Feather mosses on decaying wood 1.43 1.08 1.36 3.26(F) Sphagnum and feather mosses on litter 0.06 0.00 0.18 0.15(G) Feather mosses on rocks 0.36 0.97 0.00 0.00(H) Feather mosses on nondecaying wood 0.23 0.18 0.12 0.23(I) Feather mosses on litter 8.45 8.94 5.69 6.32(J) Litter 0.33 0.17 0.81 1.42(K) Nondecaying wood 0.00 0.13 0.91 0.03(Z) Unproductive zones
(e.g., surface occupied by a living tree or snag)0.07 0.03 0.01 0.02
Total 11.52 11.52 11.52 11.52
Table 2. Surface coverage by the different seedbed types found in the four seedling banks (in the 32demography subplots).
I:\cjfr\cjfr29\cjfr-10\X99-090.vpThursday, October 28, 1999 4:05:13 PM
Color profile: DisabledComposite Default screen

PPFD), two intermediate (4A, 6.04% PPFD; 2E, 11.17%PPFD), and one relatively open (4B, 37.00% PPFD) (Fig. 1).There were also notable differences in the number of seeds,emergents, and early established seedlings between the foursites (Table 3).
Seed rainSeedfall varied between 985.18 and 5471.87 seeds/m2 for
the 1995 seed rain. Spatial autocorrelation analysis for allsites revealed no significant distance classes (p < 0.05). Thecorrelograms usually associated with spatial analysis are notshown since none of the Moran’sI coefficients were signifi-cant at the Bonferroni-corrected level. Since no spatial struc-ture could be detected for the 1995 seed rain, seeddistribution on the forest floor was considered to be random.
Seedling demographyIn all study sites, germination started around June 19 and
ended on August 28, maximum seedling germination oc-curred on July 3, and highest mortality was observed on Au-gust 14, 1995. Overwinter mortality was cumulative and wasrecorded in the early spring of 1996. Survival curves of thefour seedling cohorts show a similar tendency: high mortal-ity in the first 10–16 weeks postgermination followed bystabilization of mortality in the remaining period (Fig. 2).Seedling survival percentages at the last census date were17.73, 12.96, 11.08, and 4.57% for sites 4B, 4A, 2E, and 1B,respectively.
Correspondence between patterns of seed dispersal andrecruitment
The relationship between spatial patterns of seed andseedling abundance was determined at the demography sub-plot level. The notion of “correspondence” refers to a signif-icant positive correlation between seed abundance in a subplotand emergents and (or) early established seedlings found inthe same subplot. Pearson’s correlation coefficients areshown in Table 4 and represent the intensity of the relation-ship between the variables. At the germination level, a sig-nificant relationship of medium intensity was found in threeof the four study sites: 4E, 2E, and 1B. This relationship dis-sipated with time, since only one of the sites (2E) showed acorrespondence between spatial patterns of seed arrival andseedlings having survived after two growing seasons.
Seedbeds and understory lightBoth G tests of goodness of fit for germination and early
establishment levels refuted the hypothesis that the observedfrequency distribution conformed to that of the theoreticaldistribution (Fig. 3). Sphagnum moss provided the bestseedbed for germination. Eight percent of the seeds that fellon this type of seedbed successfully emerged. The worstseedbeds for germination were feather mosses on rocks,feather mosses on nondecayed wood, and nondecayed wood.
© 1999 NRC Canada
Duchesneau and Morin 1505
Fig. 1. Percent photosynthetic photon flux density (%PPFD) inthe understory of the four study sites. Means and standard errorsare calculated from 32 instantaneous measurements taken underovercast sky conditions.
Site 4A Site 2E Site 4B Site 1B
Seeds/m2 (Autumn 1994 – Spring 1995) 4194.97±53.01 4058.88±33.07 985.18±9.91 5471.87±52.06Emergents/m2 (1995) 170.83±48.53 47.05±14.82 33.33±7.68 324.65±69.24Established seedlings/m2 (1996) 22.14±17.60 5.21±3.73 5.91±2.11 14.84±6.34%PPFD 6.04±2.99 11.17±0.89 37.00±12.66 2.83±1.58
Table 3. Abundance of seedfall, emergents, early established seedlings, and percent photosyntetic flux density (%PPFD)for the study sites (means are calculated from 25 seed traps and 32 demography subplots per site).
Fig. 2. Survival curves of the 1995 balsam fir seedling cohort.Relative percentage of seedling survival is defined as (number ofseedlings found at the time (weeks postgermination)/total numberof seedlings) × 100.
I:\cjfr\cjfr29\cjfr-10\X99-090.vpThursday, October 28, 1999 4:05:15 PM
Color profile: DisabledComposite Default screen
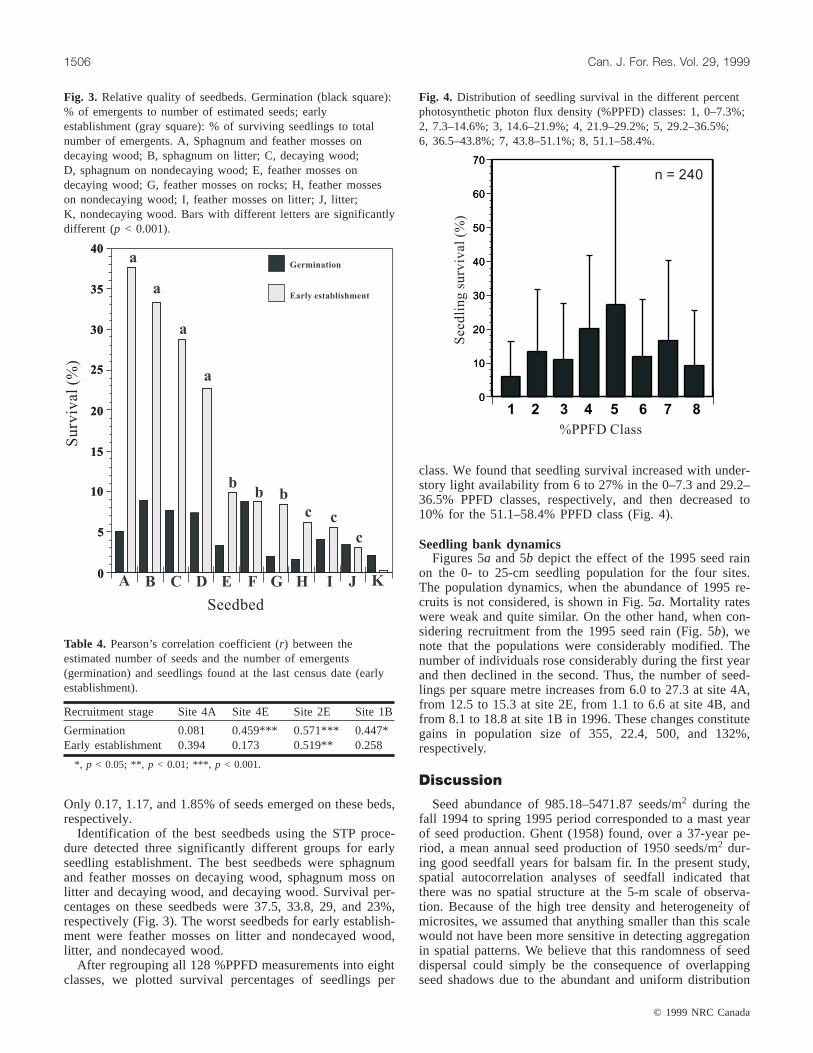
Only 0.17, 1.17, and 1.85% of seeds emerged on these beds,respectively.
Identification of the best seedbeds using the STP proce-dure detected three significantly different groups for earlyseedling establishment. The best seedbeds were sphagnumand feather mosses on decaying wood, sphagnum moss onlitter and decaying wood, and decaying wood. Survival per-centages on these seedbeds were 37.5, 33.8, 29, and 23%,respectively (Fig. 3). The worst seedbeds for early establish-ment were feather mosses on litter and nondecayed wood,litter, and nondecayed wood.
After regrouping all 128 %PPFD measurements into eightclasses, we plotted survival percentages of seedlings per
class. We found that seedling survival increased with under-story light availability from 6 to 27% in the 0–7.3 and 29.2–36.5% PPFD classes, respectively, and then decreased to10% for the 51.1–58.4% PPFD class (Fig. 4).
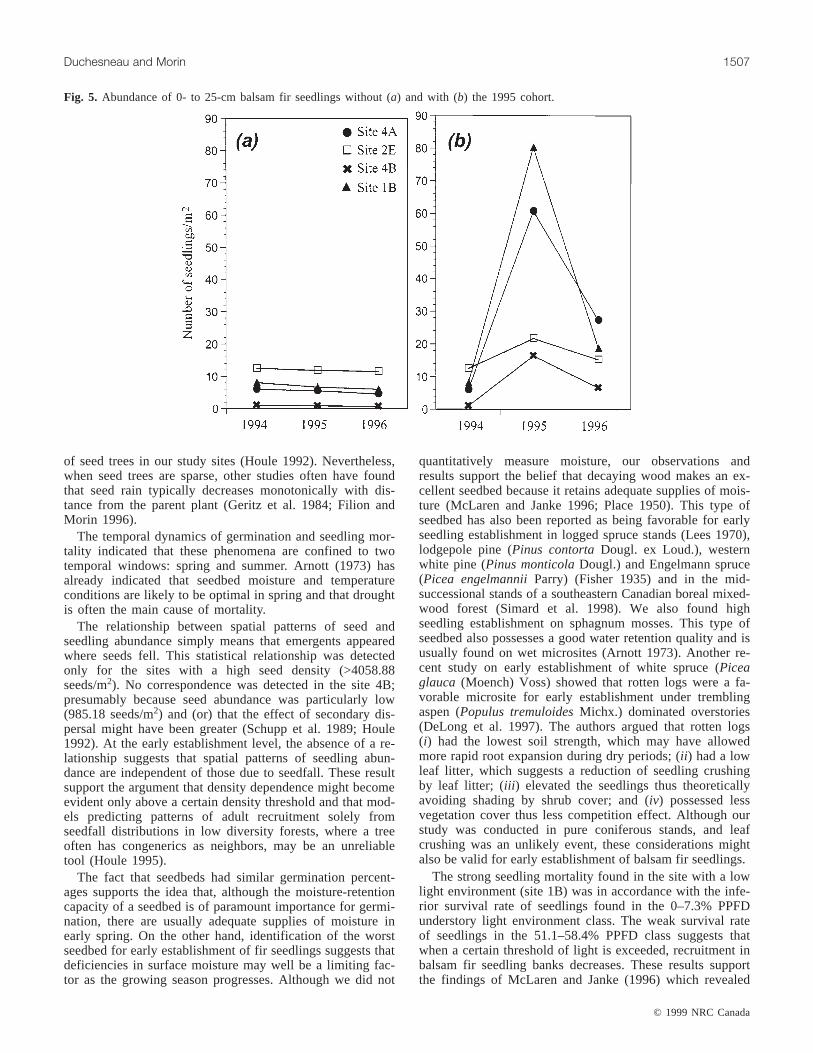
Seedling bank dynamicsFigures 5a and 5b depict the effect of the 1995 seed rain
on the 0- to 25-cm seedling population for the four sites.The population dynamics, when the abundance of 1995 re-cruits is not considered, is shown in Fig. 5a. Mortality rateswere weak and quite similar. On the other hand, when con-sidering recruitment from the 1995 seed rain (Fig. 5b), wenote that the populations were considerably modified. Thenumber of individuals rose considerably during the first yearand then declined in the second. Thus, the number of seed-lings per square metre increases from 6.0 to 27.3 at site 4A,from 12.5 to 15.3 at site 2E, from 1.1 to 6.6 at site 4B, andfrom 8.1 to 18.8 at site 1B in 1996. These changes constitutegains in population size of 355, 22.4, 500, and 132%,respectively.
Discussion
Seed abundance of 985.18–5471.87 seeds/m2 during thefall 1994 to spring 1995 period corresponded to a mast yearof seed production. Ghent (1958) found, over a 37-year pe-riod, a mean annual seed production of 1950 seeds/m2 dur-ing good seedfall years for balsam fir. In the present study,spatial autocorrelation analyses of seedfall indicated thatthere was no spatial structure at the 5-m scale of observa-tion. Because of the high tree density and heterogeneity ofmicrosites, we assumed that anything smaller than this scalewould not have been more sensitive in detecting aggregationin spatial patterns. We believe that this randomness of seeddispersal could simply be the consequence of overlappingseed shadows due to the abundant and uniform distribution
© 1999 NRC Canada
1506 Can. J. For. Res. Vol. 29, 1999
Recruitment stage Site 4A Site 4E Site 2E Site 1B
Germination 0.081 0.459*** 0.571*** 0.447*Early establishment 0.394 0.173 0.519** 0.258
*, p < 0.05; **, p < 0.01; ***, p < 0.001.
Table 4. Pearson’s correlation coefficient (r) between theestimated number of seeds and the number of emergents(germination) and seedlings found at the last census date (earlyestablishment).
Fig. 3. Relative quality of seedbeds. Germination (black square):% of emergents to number of estimated seeds; earlyestablishment (gray square): % of surviving seedlings to totalnumber of emergents. A, Sphagnum and feather mosses ondecaying wood; B, sphagnum on litter; C, decaying wood;D, sphagnum on nondecaying wood; E, feather mosses ondecaying wood; G, feather mosses on rocks; H, feather mosseson nondecaying wood; I, feather mosses on litter; J, litter;K, nondecaying wood. Bars with different letters are significantlydifferent (p < 0.001).
Fig. 4. Distribution of seedling survival in the different percentphotosynthetic photon flux density (%PPFD) classes: 1, 0–7.3%;2, 7.3–14.6%; 3, 14.6–21.9%; 4, 21.9–29.2%; 5, 29.2–36.5%;6, 36.5–43.8%; 7, 43.8–51.1%; 8, 51.1–58.4%.
I:\cjfr\cjfr29\cjfr-10\X99-090.vpThursday, October 28, 1999 4:05:17 PM
Color profile: DisabledComposite Default screen
of seed trees in our study sites (Houle 1992). Nevertheless,when seed trees are sparse, other studies often have foundthat seed rain typically decreases monotonically with dis-tance from the parent plant (Geritz et al. 1984; Filion andMorin 1996).
The temporal dynamics of germination and seedling mor-tality indicated that these phenomena are confined to twotemporal windows: spring and summer. Arnott (1973) hasalready indicated that seedbed moisture and temperatureconditions are likely to be optimal in spring and that droughtis often the main cause of mortality.
The relationship between spatial patterns of seed andseedling abundance simply means that emergents appearedwhere seeds fell. This statistical relationship was detectedonly for the sites with a high seed density (>4058.88seeds/m2). No correspondence was detected in the site 4B;presumably because seed abundance was particularly low(985.18 seeds/m2) and (or) that the effect of secondary dis-persal might have been greater (Schupp et al. 1989; Houle1992). At the early establishment level, the absence of a re-lationship suggests that spatial patterns of seedling abun-dance are independent of those due to seedfall. These resultsupport the argument that density dependence might becomeevident only above a certain density threshold and that mod-els predicting patterns of adult recruitment solely fromseedfall distributions in low diversity forests, where a treeoften has congenerics as neighbors, may be an unreliabletool (Houle 1995).
The fact that seedbeds had similar germination percent-ages supports the idea that, although the moisture-retentioncapacity of a seedbed is of paramount importance for germi-nation, there are usually adequate supplies of moisture inearly spring. On the other hand, identification of the worstseedbed for early establishment of fir seedlings suggests thatdeficiencies in surface moisture may well be a limiting fac-tor as the growing season progresses. Although we did not
quantitatively measure moisture, our observations andresults support the belief that decaying wood makes an ex-cellent seedbed because it retains adequate supplies of mois-ture (McLaren and Janke 1996; Place 1950). This type ofseedbed has also been reported as being favorable for earlyseedling establishment in logged spruce stands (Lees 1970),lodgepole pine (Pinus contortaDougl. ex Loud.), westernwhite pine (Pinus monticolaDougl.) and Engelmann spruce(Picea engelmanniiParry) (Fisher 1935) and in the mid-successional stands of a southeastern Canadian boreal mixed-wood forest (Simard et al. 1998). We also found highseedling establishment on sphagnum mosses. This type ofseedbed also possesses a good water retention quality and isusually found on wet microsites (Arnott 1973). Another re-cent study on early establishment of white spruce (Piceaglauca (Moench) Voss) showed that rotten logs were a fa-vorable microsite for early establishment under tremblingaspen (Populus tremuloidesMichx.) dominated overstories(DeLong et al. 1997). The authors argued that rotten logs(i) had the lowest soil strength, which may have allowedmore rapid root expansion during dry periods; (ii ) had a lowleaf litter, which suggests a reduction of seedling crushingby leaf litter; (iii ) elevated the seedlings thus theoreticallyavoiding shading by shrub cover; and (iv) possessed lessvegetation cover thus less competition effect. Although ourstudy was conducted in pure coniferous stands, and leafcrushing was an unlikely event, these considerations mightalso be valid for early establishment of balsam fir seedlings.
The strong seedling mortality found in the site with a lowlight environment (site 1B) was in accordance with the infe-rior survival rate of seedlings found in the 0–7.3% PPFDunderstory light environment class. The weak survival rateof seedlings in the 51.1–58.4% PPFD class suggests thatwhen a certain threshold of light is exceeded, recruitment inbalsam fir seedling banks decreases. These results supportthe findings of McLaren and Janke (1996) which revealed
© 1999 NRC Canada
Duchesneau and Morin 1507
Fig. 5. Abundance of 0- to 25-cm balsam fir seedlings without (a) and with (b) the 1995 cohort.
I:\cjfr\cjfr29\cjfr-10\X99-090.vpThursday, October 28, 1999 4:05:20 PM
Color profile: DisabledComposite Default screen

© 1999 NRC Canada
1508 Can. J. For. Res. Vol. 29, 1999
that a 40–80% closure of the canopy was beneficial for ger-mination and early establishment of balsam fir.
The sampling method used in this study did not permit usto statistically verify if the seedbeds were spatially distrib-uted in accordance to light availability. Furthermore, be-cause sites were not replicated, we could not use strongstatistical inference to show that seedling survival was siterelated. Nevertheless, we found that the site with the highestlight environment had a greater total surface of the bestseedbeds (Table 2) and a relatively better seedling survivalthan that of the site with the lowest light environment (Fig. 2).Therefore, we believe that recruitment in our seedling banksdepends not only on the abundance of seedfall, but on sitesuitability or availability of best seedbeds, which in turnmight somehow be related to understory light conditions.
The detailed demographic survey of the seedling cohortsindicates that recruitment, even following a mast year ofproduction, remains limited. Yet, addition of new recruitsmodified the structure of the banks to the extent that the to-tal 0- to 25-cm population grew in size instead of decreas-ing. Factors such as seedbed type, understory light, andquantity of seedfall production would then be of paramountimportance when considering seedling bank dynamics in bo-real balsam fir forests. For example, it took approximately167 seeds/m2 to establish one seedling in the site having amean understory light environment of 37% PPFD and 340seeds/m2 for the site with an understory of 2.83% PPFD.
If in the years to come, mortality rates of the 1995 cohortsare similar to those of already established 0- to 25-cm seed-lings, we believe that mast years of seed production, at leastduring endemic periods or in very low understory light envi-ronments, may be crucial for recruitment and renewal of bal-sam fir seedling banks. This does not exclude the possibilitythat two phenomena, renewal and lack of growth, mightoccur simultaneously. This would lead to the balsam firs’ re-generative success following recurring spruce budworm out-breaks (Morin and Laprise 1997). In this case, throughoutthe epidemic periods there would be no cone or seed produc-tion and hence no recruitment, making stagnation of theseedling bank an important factor in assuring the renewal ofbalsam fir forests.
Finally, if we are to fully understand the seedling bankdynamics of boreal fir forests, further long-term studies needto address the relationship between seed rain, site suitability,understory light availability, patterns of seed dispersal andtheir demographic consequences at all sequential stagesfrom seed to new adult.
References
Arnott, J.T. 1973. Germination and seedling establishment.In Pro-ceedings of Direct Seeding Symposium, Sept. 1973, Timmins,Ont. Edited byJ.H. Cayford. Can. For. Serv. Publ. No. 1339.pp. 55–66.
Baskerville, G. 1975. Spruce budworm—super sylviculturist. For.Chron.51: 4–6.
Blais, J.R. 1954. The recurrence of spruce budworm infestation inthe past century in the lac Seul area of northwestern Ontario.Ecology,35: 62–71.
DeLong, H.B., Lieffers, V.J., and Blenis, P.V. 1997. Microsite ef-fects on first-year establishment and overwinter survival of
white spruce in aspen-dominated boreal mixedwoods. Can. J.For. Res.27: 1452–1457.
De Steven, D. 1991. Experiments on mechanisms of tree establish-ment in old-field succession: seedling survival and growth.Ecology,72: 1076–1088.
Environnement Canada. 1992. Sommaire métérologique mensuel,Chibougamau–Chapais, janvier–décembre 1992. ServiceEnvironnement atmosphérique, Environnement Canada, Ottawa,Ont.
Eriksson, O., and Ehrlén, J. 1992. Seed and microsite limitation ofrecruitment in plant populations. Oecologia,91: 360–364.
Fisher, G.M. 1935. Comparative germination of tree species onvarious kinds of surface soil material in the western white pinetype. Ecology,16: 605–611.
Filion, J., and Morin, H. 1996. Distribution spatiale de larégénération de l’épinette noire 8 ans après un feu en forêtboréale (Québec). Can. J. For. Res.26: 601–610.
Fleming, R.L., and Mossa, D.S. 1994. Direct seeding of blackspruce in northwestern Ontario: seedbed relationships. For.Chron.70: 151–158.
Fowler, N.L. 1988. What is a safe site? Neighbor, litter, germina-tion date, and patch effects. Ecology,64: 947–961.
Geritz, S.A.H., de Jong, T.J., and Klinkhamer, P.G.L. 1984. The ef-ficacy of dispersal in relation to safe site area and seed produc-tion. Oecologia,62: 219–221.
Ghent, A.W. 1958. Studies of regeneration in forest strands devas-tated by the spruce budworm II. Age height growth, and relatedstudies of balsam fir seedlings. For. Sci.4: 135–146.
Ghent, A.W., Fraser, D.A., and Thomas, J.B. 1957. Studies of re-generation in forest strands devastated by the spruce budworm I.Evidence of trends in forest succession during the first decadefollowing budworm devastation. For. Sci.3: 184–208.
Grandtner, M.M. 1966. La végétation forestière du Québecméridional. Presses de l’Université Laval, Québec, Que.pp. 219–220.
Groot, A. 1988. Methods for estimating seedbed receptivity and forpredicting seedling stocking and density in broadcast seeding.Can. J. For. Res.18: 1541–1559.
Grubb, P.J. 1977. The maintenance of species richness in plantcommunities: the importance of the regeneration niche. Biol.Rev. Camb. Philos. Soc.52: 107–145.
Harper, J.L. 1977. Population biology of plants. Academic Press,London.
Harper, J.L., Clatworthy, J.N., McNaughton, I.H., and Sagar, G.R.1961. The evolution of closely related species living in the samearea. Evolution,15: 209–227.
Houle, G. 1992. The reproductive ecology ofAbies balsamea, Acersaccharumand Betula alleghaniensisin the Tantaré EcologicalReserve, Quebec. J. Ecol.80: 611–623.
Houle, G. 1995. Seed dispersal and seedling recruitment: the miss-ing link(s). Écoscience,2: 238–244.
Houle, G., and Payette, S. 1991. Seed dynamics ofAbies balsameaandAcer saccharumin a deciduous forest of northeastern NorthAmerica. Am. J. Bot.78: 895–905.
Lees, J.C. 1970. Natural regeneration of white spruce under spruce–aspen shelterwood, B-18a forest section, Alberta. Can. For.Serv. Publ. No. 1274. pp. 16–32.
Legendre, P., and Fortin, M.J. 1989. Spatial pattern and ecologicalanalysis. Vegetatio,80: 107–138.
Legendre, P., and Vaudor, A. 1991. Le progiciel R. Analysemultidimensionnelle, analyse spatiale. Département des sciencesbiologiques, Université de Montréal, Montréal, Que.
Li, X.L., and Burton, P.J. 1994. Interactive effects of light and
I:\cjfr\cjfr29\cjfr-10\X99-090.vpThursday, October 28, 1999 4:05:21 PM
Color profile: DisabledComposite Default screen

© 1999 NRC Canada
Duchesneau and Morin 1509
stratification on the germination of some British Colombia coni-fers. Can. J. Bot.72: 1635–1646.
MacLean, D.A. 1984. Effects of spruce budworm outbreaks on theproductivity and stability of balsam fir forests. For. Chron.60:273–279.
MacLean, D.A. 1988. Effects of spruce budworm outbreaks onvegetation, structure, and succession of balsam fir forests ofCape Breton Island, Canada.In Plant form and vegetation struc-ture. Edited byM.J.A. Werger, P.J.M. Van Der Aart, H.J. Dur-ing, and J.T.A. Verhoeven. SPB Academic Publishing, TheHague, the Netherlands. pp. 253–261.
McLaren, B.E. and Janke, R.A. 1996. Seedbed and canopy covereffects on balsam fir seedling establishment in Isle Royale Na-tional Park. Can. J. For. Res.26: 782–793.
Messier, C., and Puttonen, P. 1995. Spatial and temporal variationin the light environment of developing Scots pine stands: the ba-sis for a quick and efficient method of characterizing light. Can.J. For. Res.25: 343–354.
Morin, H. 1990. Analyse dendroécologique d’une sapinière issued’un chablis dans la zone boréale, Québec. Can. J. For. Res.20:1753–1758.
Morin, H. 1994. Dynamics of balsam fir forests in relation tospruce budworm outbreaks in the Boreal Zone of Quebec. Can.J. For. Res.24: 730–741.
Morin, H., and Laprise, D. 1990. Histoire récente des épidémies dela tordeuse des bourgeons de l’épinette au nord du lac Saint-Jean (Québec) : une analyse dendrochronologique. Can. J. For.Res.20: 1–8.
Morin, H., and Laprise, D. 1997. Seedling bank dynamics in borealbalsam fir forests. Can. J. For. Res.27: 1442–1451.
Morris, R.F. 1951. The effects of flowering on the foliage produc-tion and growth of balsam fir. For. Chron.27: 40–57.
Morris, R.F. 1963. The dynamics of epidemic spruce budwormpopulations. Mem. Entomol. Soc. Can.31: 430–460.
Oden, N.L., and Sokal, R.R. 1986. Directional autocorrelation: anextension of spatial correlograms to two dimensions. Syst. Zool.35: 608–617.
Osawa, A. 1994. Seedling responses to forest canopy disturbancefollowing a spruce budworm outbreak in Maine. Can. J. For.Res.24: 850–859.
Oswald, B.P., and Neuenschwander, L.F. 1993. Microsite variabil-
ity and safe site description for western larch germination andestablishment. Bull. Torrey Bot. Club,120: 148–156.
Parent, S., and Messier, C. 1995. Effets d’un gradient de lumièresur la croissance en hauteur et la morphologie de la cime dusapin baumier régénéré naturellement. Can. J. For. Res.25:878–885.
Parent, S., and Messier, C. 1996. A simple and efficient method toestimate microsite light availability under a forest canopy. Can.J. For. Res.26: 151–154.
Place, I.C.M. 1950. Comparative moisture regimes of humus androtten wood. Silv. Leafl. For. Branch Can. No. 64. pp. 4–18.
Ruel, J.C., and Huot, M. 1993. Impact de la tordeuse desbourgeons de l’épinette sur la régénération des sapinières aprèscoupe à blanc. For. Chron.69: 163–172.
Schupp, E.W. 1988. Seed and early seedling predation in theunderstory and in treefall gaps. Oikos,51: 71–78.
Schupp, E.W., and Fuentes, M. 1995. Spatial patterns of seed dis-persal and the unification of plant population ecology.Écoscience,2: 267–275.
Schupp, E.W., Howe, H.F., Augspurger, C.K., and Levey, D.J.1989. Arrival and survival in tropical treefall gaps. Ecology,70:562–564.
Simard, M.J., Bergerons, Y., and Sirois, L. 1998. Conifer seedlingrecruitment in a southeastern Canadian boreal forest: the impor-tance of substrate. J. Veg. Sci.9: 575–582.
Sokal, R.R., and Rohlf, F.J. 1981. Biometry. W.H. Freeman & Co.,New York.
SYSTAT, Inc. 1992. Statistics. 5th ed. SYSTAT, Inc., Evanston, Ill.Szewczyk, J., and Szwagrzyk, J. 1996. Tree regeneration on rotten
wood and on soil in an old-growth stand. Vegetatio,122: 37–46.Taylor, K.M., and Aarssen, L.W. 1989. Neighbor effects in mast
year seedlings ofAcer saccharum. Am. J. Bot.76: 546–554.Thibault, M. 1987. Les régions écologiques du Québec méridional.
Deuxième approximation. Carte. Service de la recherche,ministère de l’Énergie et des Ressources du Québec, Que.
Thomas, P.A., and Wein, R.W. 1985. Delayed emergence of fourconifer species on postfire seedbeds in eastern Canada. Can. J.For. Res.15: 727–729.
Whelan, C.J., Willson, M.F., Tuma, C.A., and Souza-Pinto, I.1990. Spatial and temporal patterns of postdispersal seed preda-tion. Can. J. Bot.69: 428–436.
I:\cjfr\cjfr29\cjfr-10\X99-090.vpThursday, October 28, 1999 4:05:22 PM
Color profile: DisabledComposite Default screen