dynamics of senescence-related qtls in potato
TRANSCRIPT

Dynamics of senescence-related QTLs in potato
Paula Ximena Hurtado • Sabine K. Schnabel • Alon Zaban •
Merja Vetelainen • Elina Virtanen • Paul H. C. Eilers •
Fred A. van Eeuwijk • Richard G. F. Visser • Chris Maliepaard
Received: 27 February 2010 / Accepted: 17 May 2011 / Published online: 12 June 2011
� The Author(s) 2011. This article is published with open access at Springerlink.com
Abstract The study of quantitative trait’s expression
over time helps to understand developmental processes
which occur in the course of the growing season.
Temperature and other environmental factors play an
important role. The dynamics of haulm senescence was
observed in a diploid potato mapping population in two
consecutive years (2004 and 2005) under field condi-
tions in Finland. The available time series data were
used in a smoothed generalized linear model to
characterize curves describing the senescence devel-
opment in terms of its onset, mean and maximum
progression rate and inflection point. These character-
istics together with the individual time points were
used in a Quantitative trait loci (QTL) analysis.
Although QTLs occurring early in the senescence
process coincided with QTLs for onset of senescence,
the analysis of the time points made it difficult to study
senescence as a continuous trait. Characteristics esti-
mated from the senescence curve allowed us to study it
as a developmental process and provide a meaningful
biological interpretation to the results. Stable QTLs in
the two experimental years were identified for pro-
gression rate and year-specific QTLs were detected for
onset of senescence and inflection point. Specific
interactions between loci controlling senescence
development were also found. Epistatic interaction
between QTLs on chromosomes 4, 5 and 7 were
detected in 2004 and pleiotopic effects of QTLs on
chromosomes 3 and 4 were observed in 2005.
Keywords Beta-thermal time � Epistasis �Functional QTL mapping � Smoothing �Time series � GLM
Introduction
The study of developmental processes in plants
requires the evaluation of traits over time taking into
P. X. Hurtado (&) � R. G. F. Visser � C. Maliepaard
Wageningen UR Plant Breeding, Wageningen University,
P.O. Box 386, 6700 AJ Wageningen, The Netherlands
e-mail: [email protected]
P. X. Hurtado � S. K. Schnabel � P. H. C. Eilers �F. A. van Eeuwijk
Biometris—Applied Statistics, Wageningen University,
P.O. Box 100, 6700 AC Wageningen, The Netherlands
A. Zaban � M. Vetelainen � E. Virtanen
AgriFood Research Finland (MTT), North Ostrobothnia
Research Station, Ruukki, Finland
P. H. C. Eilers
Department of Biostatistics, Erasmus Medical Center,
P.O. Box 2040, 3000 CA Rotterdam, The Netherlands
P. X. Hurtado
C.T. de Wit Graduate School for Production Ecology
and Resource Conservation (PE&RC), Wageningen
University, P.O. Box 386, 6708 PB Wageningen,
The Netherlands
S. K. Schnabel � F. A. van Eeuwijk � R. G. F. Visser
Centre for BioSystems Genomics, P.O. Box 98,
6700 AB Wageningen, The Netherlands
123
Euphytica (2012) 183:289–302
DOI 10.1007/s10681-011-0464-4

account the continuous nature of development.
Quantitative traits normally involved in these pro-
cesses require assessments at several time points
during the life cycle. However, conventional exper-
iments include evaluations at a single fixed time point
during the growing season. Quantitative trait loci
(QTL) analysis using the data collected from those
experiments give just an impression of the loci
affecting the trait at a particular developmental stage.
The understanding of the genetic basis controlling
quantitative traits improves when the evolution of the
trait during the life cycle and the developmental
pattern are considered. In addition, environmental
factors affecting crop development, such as temper-
ature and photoperiod, must be also taken into
account. Therefore, growth models flexible enough
to be adapted to the usually non-linear trait responses
over time have to be implemented. Examples of
commonly used models in biology are the exponen-
tial growth models and the family of s-shaped curves
(Schnute 1981). These models have the advantage of
describing the development of the trait in terms of
curve parameters with a biological interpretation. The
differences in growth trajectories between genotypes
are reflected by genotype-specific curve parameters.
Growth models and QTL analysis can be combined
by modeling growth curve parameters in terms of
QTLs in one-step (Ma et al. 2002; Malosetti et al.
2006) or two-step model approaches (Reymond et al.
2003; Yin et al. 1995). In the former, logistic growth
curves and QTL mapping are combined, modeling
growth parameters as a function of the QTL geno-
types described by molecular marker scores. In the
latter, first genotype-specific parameters are esti-
mated from the growth curves, thereafter they are
used as phenotypic traits in a conventional QTL
analysis. In both approaches, the accurate estimation
of the curve parameters plays an important role to
find meaningful results.
In our study, haulm senescence was assessed at
several time points during the growing season on a
diploid potato mapping population evaluated for two
consecutive years in Finland (2004 and 2005). We are
presenting here an alternative two-step approach:
modeling first in a flexible way the curve trajectories
with a smoothed generalized linear model (GLM)
procedure. We used the collected time series data to
model the development of senescence in terms of time
and temperature. To allow a better comparability of the
trait across the years, we converted the calendar days
after planting into beta-thermal days after planting
(TAP) using a non-linear temperature effect function
described by Khan et al. (2011). Once the senescence
trajectory was defined, the characteristics of the curve-
such as onset, maximum and average progression rate
and inflection point- were estimated from the first and
second derivative of the curve. These characteristics
were used as phenotypic traits describing senescence
development in the QTL analysis. In this way, the
characterization of the smoothed GLM curves allowed
us to map the genetic basis of the senescence process
giving a meaningful biological interpretation to the
results.
Materials and methods
Description of the C9E potato population
The evaluation of haulm senescence was done on a
diploid backcross potato population (C9E) composed
of 250 genotypes. The C9E population is the result
of a cross between two diploid potato clones. The
female parent, the C clone US-W5337.3 (Hanneman
and Peloquin 1967) is a hybrid between Solanum
phureja (PI225696) and S. tuberosum dihaploid
US-W42. The male parent, the E clone (Jacobsen
1980), is a hybrid between VH34211 (a S. vernei–S.
tuberosum backcross) and the C clone itself.
S. phureja is characterized by a lack of dormancy,
early maturity and short day tuberization induction.
In contrast, S. tuberosum varieties have long dor-
mancy, variable maturity and long day tuberization
induction (Hawkes 1990; Ewing and Struik 1992).
The different day length requirements for tuberiza-
tion of S. phurja and S. tuberosum has provided a
source of genetic variation in the C9E population
making it suitable for the study of developmental
processes under different photoperiods.
Experimental design and description of the haulm
senescence evaluation
The very long day conditions of northern Finland
were used to conduct an experiment in two consec-
utive years (2004 and 2005) in the experimental field
of AgriFood Research Finland (MTT) at the North
Ostrobothnia Research Station in Ruukki (64�420N,
290 Euphytica (2012) 183:289–302
123

25�000E). The experiment performed in 2004 was
previously described by Zaban et al. (2006). Sets of
197 and 222 genotypes of the C9E population were
planted in plots of three plants per genotype, where
the genotypes were randomized, on June 1st 2004 and
May 16th 2005, respectively. We acknowledge that in
these trials there was no real replication of genotypes,
but only pseudo-replications. Each plant was evalu-
ated at several time points spaced in intervals of
3–7 days. In 2004 the observation period was 52 days
with evaluations at 77, 81, 86, 91, 95, 100, 105, 109,
116, 123, 129 days after planting (DAP). In the
second year the observation period was shorter
(31 days) and haulm senescence was evaluated at
84, 88, 93, 98, 101, 106, 109, 112 and 115 DAP. The
process of senescence was defined as the period
between the last observation at which the plant was
entirely green and the first date at which the plant was
dead (Celis-Gamboa 2002). The progress of the trait
was measured using the scale described by Celis-
Gamboa et al. (2003) in which 1 = green plant;
2 = upper leaves with the first signs of yellowing
(light green); 3 = yellow leaves; 4 = 25% of haulm
tissue brown; 5 = 50% of haulm tissue brown;
6 = more than 75% of haulm tissue brown; 7 = dead
plant.
Description of Beta-thermal time estimation
Crop development is mainly affected by temperature
and can be modified by other factors such as
photoperiod (Hodges 1991). Previous studies have
shown that potato growth is influenced by tempera-
ture, where warmer temperatures favor vegetative
growth accelerating reproductive and vegetative
development (Haun 1975; Benoit et al. 1986; Struik
and Ewing 1995). Whereas, lower temperatures
facilitate tuber growth (Marinus and Boadlaender
1975). The effect of temperature on crop develop-
ment rate is often described by using a thermal time
concept. Various non-linear models have been devel-
oped to describe temperature response of develop-
mental processes in plants (Johnson and Thornley
1985; Gao et al. 1992; Yin et al. 1995). In our study,
the daily contribution of temperature to plant devel-
opment in 2004 was different from 2005 due to the
fluctuations in daily air temperature. A summary of
the meteorological conditions reported by the station
in Ruukki (Finland) during the growing season in the
two experimental years is presented in Table 1. The
growing season defined as the period between the last
killing frost of spring and the first killing frost of
autumn (Allaby 1998) was defined in 2004 from
April 16 to November 10 and in 2005 from May 3 to
November 14. To allow for a better comparability of
senescence across the years, we converted the
calendar DAP into beta-thermal days after planting
(TAP) using the non-linear temperature effect func-
tion g(T) described by Khan et al. (2011):
gðTÞ ¼ Tc � T
Tc � To
� �T � Tb
To � Tb
� �To�TbTc�To
" #ct
ð1Þ
where the three main temperatures, base, optimum
and ceiling temperature for phenological develop-
ment of potato, were defined as Tb = 5.5�C,
To = 23.4�C and Tc = 34.6�C, respectively. The
temperature response curvature coefficient was esti-
mated as ct = 1.7 according to Khan et al. (2011).
Because the function g(T) is non-linear and temper-
ature fluctuates daily, g(T) was estimated using the
average daily air temperature to obtain the daily
value. The cumulative TAP, combining temperature
and time (beta-thermal time, BTT), was the scale of
the x-axis used to compare senescence development
in the two experimental years.
Table 1 Summary of meteorological conditions reported in Ruukki, Finland during the growing season in 2004 and 2005
Year 2004 2005
Average daily air temperature (�C) 12.7 14.5
Temperature range (�C) 2.6–20.5 4.8–21.9
Amount of rainfall (mm) 514 359
Average daily rainfall (mm) 2.1 1.5
Average daily radiation (kJ/cm2) 142.9 155.05
Radiation range (kJ/cm2) 19.20–297.20 35.04–280.72
Euphytica (2012) 183:289–302 291
123

Description of the smoothed generalized linear
model procedure
To estimate the senescence curve non-parametrically,
a variant of penalized B-splines, P-splines (Eilers and
Marx 1996), was used. Then we interpreted a senes-
cence value y as having observed y ‘‘successes’’ in n
binomial trials and we estimated a smooth curve for the
probability p of a ‘‘success’’. To guarantee that
0 \ p \ 1, we worked with the linear predictor. In
this case it is the logit, g = log(p/(1-p)), which has no
restrictions on its range. This is standard practice in
logistic regression and generalized linear models
(Nelder and Wedderburn 1972). Smooth logistic
regression with P-splines was explained by Eilers
and Marx (1996). They also showed that automatic
interpolation was obtained. We exploited this prop-
erty by working with zero degree B-splines including a
knot on each beta-thermal day (TAP units) of the
domain we study. The penalized log-likelihood in this
situation is
l� ¼X
i
wi½yi logpiþðni� yiÞ logð1� piÞþ logni
yi
� ��
� k2
Xi
ðD2giÞ2�j2
Xi
g2i ð2Þ
with pi ¼ 1= 1þ e�gið Þ:The weight is determined by
wi ¼ npi 1� pið Þ following GLM methodology for
our situation. The second term in the equation above
is the penalty on the linear predictor g. The parameter
k tunes its weight: the larger k, the smoother the
result. We used the operator notation to indicate
repeated differences: D2gi ¼ gi� 2gi�1 þ gi�2 ¼gi� gi�1ð Þ� gi�1� gi�2ð Þ:
One can show that for very large k the estimated
curve for p approaches the logistic curve. Further-
more we used a ridge penalty on g tuned by the
parameter j to avoid numerical instabilities, when gbecomes too large. To estimate g, the iterative
weighted linear regression algorithm (Nelder and
Wedderburn 1972) was modified, to account for the
penalties.
The fitting procedure resulted in a smooth curve
for the development of haulm senescence over time
for each genotype. This flexible functional form using
splines allowed to model different shapes of these
curves (e.g., for early and late genotypes) within the
same framework.
Characteristics estimated from the senescence
curves
Once the senescence curves were fitted, some charac-
teristics of the curves describing the ageing process
were estimated to allow the study of senescence as a
continuous trait changing in time. The characteristics
estimated from the curve have a meaningful biological
interpretation for the senescence development and
facilitate the understanding of the QTL mapping
results. We calculated the first differences on the curve
values (first derivative) as a proxy of the slope of the
curve. The mean slope or mean progression rate
(mprate) is reflecting the average rate of change of the
senescence curve during the whole observation period
giving an idea of how fast a genotype is experiencing
the senescence process. The maximum slope (prate)
explains the maximum rate of change of the senescence
curve when the plant has completed half of the
senescence process (between 4 and 5 in the senescence
scale). The higher the value, the faster full senescence
is reached during the observation period. The inflection
point (ipoint) reflects the point in time when half of the
senescence process has been reached and the trajectory
curve change from convex to concave shape indicating
the beginning of the final stage. Additional character-
istics were deduced from the second differences of the
curve values (second derivative), including the max-
imum and minimum change of the slope, which are
interpreted as the onset and the end of the senescence
process. The onset is indicating the beginning of the
senescence process in terms of time or more accurately
in our case in terms of BTT. The lower the value, the
earlier the senescence process starts.
Repeatability estimation
Data from the three plants within a plot for individual
genotypes were used for a repeatability estimation,
calculated for each year. The repeatability was
defined by the ratio of genetic (genotypic) to
phenotypic variance. Phenotypic variance was equal
to the sum of the genetic variance and a third times
the between-plants-within-plot variance.
G9E interactions
The estimation of genotype by year interactions
included 186 genotypes evaluated in the two
292 Euphytica (2012) 183:289–302
123

consecutive years using a two-way analysis of
variance with genotype and environment fixed in
the model. The curve characteristics (mprate, prate,
ipoint and onset) were used as response variables, y,
and the genotype (G), year (E) and Genotype 9 Year
(G 9 E) interaction as the explanatory variables in
the model, which also includes an error term (based
on the variation between the three plants per
genotype):
y ¼ Gþ E þ G� E þ error: ð3Þ
Description of the genetic map and molecular data
The C9E population was genotyped using amplified
fragment length polymorphism, AFLP (Celis-Gam-
boa 2002), simple sequence repeat, SSR, and cleav-
age amplified polymorphism, CAPS (Werij et al.
2007). Our study included a subset of dominant and
co-dominant markers with the expected segregation
ratios 1:1 and 1:1:1:1, respectively. JoinMap 4 (Van
Ooijen 2006) was used to construct the C and E
parental maps using 164 and 198 markers, respec-
tively, each with 12 linkage groups (LG) previously
reported by Celis-Gamboa (2002). The C map
consisted of 135 markers spanning 917.4 cM. Four
of the 12 LG were split in two sub-groups and one
was split in 3 sub-groups due to the large distances
between adjacent markers (more than 30 cM). The E
map consisted of 132 markers spanning 629.8 cM
and 2 of the 12 LG were split in 2 sub-groups. Since
the maternal (C) and paternal (E) maps were not
integrated due to the expected differences in the
recombination frequencies between the two parents
(with genetic background from two different Solanum
species), the separated parental maps were used to
perform the QTL analysis. Co-dominant markers
present in both parental maps (C and E) were used to
identify the same LG in the two maps. The assign-
ment of linkage groups was done using as a reference
the maps of Celis-Gamboa (2002), each LG is
preceded by the letter C or E according to the
parental map, followed by the LG number.
QTL analysis using curve characteristics
and single time points
The detection of QTLs, estimation of QTL main
effects and their interactions was done in two steps.
In the first step, the detection of QTLs was performed
separately for each type of phenotypic data available.
In the analysis per time point the senescence scores,
from 1 to 7, were used in the QTL mapping. In the
analysis of the characteristics estimated from the
curves (mprate, prate, ipoint and onset), each char-
acteristic was used as a quantitative trait. The use of
model parameters in QTL mapping has been intro-
duced in functional mapping for the analysis of time-
series traits. It incorporates biological principles
(defined with mathematical functions) into the frame-
work for QTL mapping (Wu et al. 2003; Wu and Lin
2006).
A nonparametric QTL analysis was performed for
the time point data using the rank sum test of
Kruskal–Wallis (KW). The curve characteristics were
analyzed using both KW and interval mapping (IM).
Both procedures are available in MapQTL 6 (Van
Ooijen 2009). For the curve characteristics, QTLs
detected by KW and IM were the same. For
transparent comparison of QTL analysis on time
points and curve characteristics, in this paper we
present only results from KW.
QTL mapping was performed separately on the
parental maps and the criterion for detecting QTLs
was set at a significance level of P B 0.005 (Van
Ooijen 2006). To identify the detected QTLs, the
origin of the map (C and E parent) and the linkage
group are indicated in the QTL name using italic
letters. The QTLs were named according to the
parental map and the linkage group in which they
were identified. As an example, the most significant
QTL on linkage group 4 in the C parent was called
C4 and in the case of common markers detected in
both parents, like E5 and C5, the QTL was called Ch5
(corresponding to a QTL on chromosome 5).
QTL 9 QTL interactions
IM was used for detecting main effect QTLs for the
curve characteristics mprate, prate, ipoint and onset.
The presence of epistatic interactions was investigated
per year by fitting linear models (Genstat release 13.2,
Payne et al. 2010) containing pairwise interactions
between those QTLs for whom earlier main effects
were detected for any of the four curve characteristics.
A full model was defined with each curve character-
istic as response variable, y, and the main effect QTLs
Euphytica (2012) 183:289–302 293
123

together with all possible QTL 9 QTL interactions as
fixed explanatory variables.
y ¼ QTLsþ QTL� QTL interactionsþ error ð4ÞFrom the full model (4), for each curve character-
istic a subset of model terms was created. This list
contained the significant main effects QTLs, all
significant epistatic interactions, plus non-significant
main effect QTLs whenever these QTLs were
involved in significant epistatic interactions. A final
model was fitted for each curve characteristic
including only the earlier selected terms.
Results
Curve fitting
The transformation of normal calendar days after
planting into TAP and their cumulative values gener-
ated a new scale (BTT) to measure the senescence
development in terms of time and accumulated tem-
perature. BTT went from 33.3 to 41.85 and from 37.1 to
50.8 during the observation period in 2004 (77–129
DAP) and 2005 (84–115 DAP), respectively. This
scale was used as the x-axis to fit the curves and
visualize the progression of haulm senescence scored
from 1 to 7. The use of the smoothed GLM procedure in
our study allowed flexible modeling of different curve
shapes. Different types of curves were observed in the
C9E population according to the maturity type of each
genotype (Fig. 1a). The description of the five catego-
ries (very early, early, intermediate, late and very late)
according to the duration of the plant cycle was
reported by Celis-Gamboa (2002). In early genotypes
we observed a logistic s-shape in the curve trajectory
while in the late genotypes we had only the exponential
part of the curves often corresponding to completion of
only about half of the senescence process.
After fitting the curves for each genotype, several
characteristics describing the ageing process were
estimated to be used in the QTL analysis. The first
differences on the curves allowed the estimation of
mprate, prate and ipoint (Fig. 1b) and the second
derivative was used to determine the onset of the
senescence process (Fig. 1c). The minimum second
derivative was calculated for the early and interme-
diate genotypes but it was not accurately estimated
for the late genotypes, therefore it was not considered
in this study. However, it can be interpreted as the
end of the senescence process and experiments with a
longer observation period could make use of this
characteristic as well to describe the final stage of
senescence.
Repeatability estimation
Once the curve characteristics were calculated, the
repeatability of each one was estimated for each year.
Fig. 1 Characterization of senescence development in the
C9E population represented by three genotypes with different
maturity type: CE78 (early: red-top line), CE47 (intermediate:
blue-middle line) and CE697 (late: yellow-lowest line). a Fitted
curves showing the senescence development. b First derivative
of the curves in which the maximum slope represents the
progression rate and the dashed lines represent the time when
half of the senescence process has been completed (inflection
point) in each genotype. c Second derivative of the curves in
which the maximum second difference is interpreted as the
onset of senescence and it is indicated with a dashed line in
each genotype. BTT beta-thermal time representing cumulative
TAP beta-thermal days after planting
294 Euphytica (2012) 183:289–302
123

High values were observed for all of them in the
2 years with values between 0.89 and 0.98. Repeat-
ability was also estimated in each time point and
values higher than 0.92 were obtained during the
observation period in each year. The high repeatabil-
ity values observed in the curve characteristics and in
the individual time points are due to the experimental
design.
G9E interactions
A set of 186 genotypes evaluated in 2004 and 2005
was used to estimate the year effect, the genotype
main effect and G9E interaction for prate, mprate,
ipoint and onset according to model (3). There was a
clear year effect for all four traits (P \ 0.001). We
observed in 2005 a senescence period lasting longer
than in 2004 with a slow progression rate. In 2004,
the average mprate and prate were higher (0.36 and
0.86) than in 2005 (0.24 and 0.55), whereas the
average onset and ipoint were higher in 2005 (45.97
and 49.90) than in 2004 (37.28 and 39.65). The
observation period in 2005 was not long enough for
late genotypes to complete the senescence process.
Figure 2 shows the senescence fitted curves of three
genotypes with different maturity type (CE78: early,
CE47: intermediate and CE697: late) in 2004 and
2005 to exemplify the effect of genotype and year in
the C9E population. Significant interactions between
genotypes and year (P-value \ 0.001) were also
observed for each curve characteristic. Larger differ-
ences in mprate, ipoint and onset were observed
between early and late genotypes in 2005, whereas
the differences between early and late genotypes
were larger for prate in 2004. Early genotypes had a
fast senescence development and late genotypes
became very late in 2005.
QTL analysis: characteristics of the curves
and single time points
QTLs were detected in each parental map using the
individual time points and the characteristics esti-
mated from the curves in each year, considering as a
significance threshold a P-value from the Kruskal–
Wallis test lower than 0.005. Only the LGs in which
QTLs were identified are shown in Fig. 3 and two
levels of significance are highlighted (0.001 \ P B
0.005, P B 0.001). In the analysis per time point, 10
and 9 time points were used in the QTL analysis in
2004 and 2005, respectively. In the first year, the first
time point was excluded from the analysis because all
the genotypes were still green. In the analysis using
the curve characteristics, although four of them were
used for the QTL mapping using KW and IM, results
are presented only for onset, prate and ipoint with
Kruskal–Wallis. It is due to the fact that high
correlations were observed between mprate and prate
in the two consecutive years (0.85 and 0.80, respec-
tively) and the same QTLs were detected for both
curve characteristics using KW and IM.
Comparing the results using time points and curve
characteristics, we observed that QTLs detected in
early time points, coincided with QTLs on LG C4,
C5A and E3 associated with onset of senescence.
QTLs occurring in more advanced stages of the
senescence development coincided with QTLs for
prate and ipoint as shown in C5 and E5 (they
correspond to the same LG in both parents and hence
are termed Ch5).
Environment (year)-specific QTLs were found on
C1 and C5A associated with prate in 2005 and onset
in 2004, respectively. QTLs associated with ipoint
were also related with onset in 2005 as it is shown in
E7 and Ch5. The pleiotropic QTLs detected for ipoint
and onset on both LG explained the high correlation
(0.8) observed between the two traits in that year. On
the other hand, QTLs consistently found in C4 in the
two experimental years were associated with onset
and the more significant P-value was observed in
2005 (P \ 0.0001).
Fig. 2 Comparison of senescence development in Finland
during 2004 (solid line) and 2005 (dashed line) represented by
three genotypes with different maturity type: CE78 (early: red-
top line), CE47 (intermediate: blue-middle line) and CE697
(late: yellow-lowest line). The x-axis corresponds to BTT units
adjusted for comparison of the 2 years
Euphytica (2012) 183:289–302 295
123

a
296 Euphytica (2012) 183:289–302
123

QTL9QTL interactions
The most significant QTLs for prate, ipoint and onset
in each linkage group and all the pairwise interactions
between them were used in a linear model to investi-
gate epistatic interactions, within a year. The full
model was based on the joined set of main effect QTLs
as detected for any of the three curve characteristics.
Fig. 3 Graphical representation of the QTLs detected in the C
(a) and E (b) parental maps. Results from the QTL analysis using
data from the field experiments in 2004 and 2005 are presented in
the left and right panel respectively. Only the linkage groups in
which significant QTLs were detected using individual time
points (in 2004: 81,86,91,95,100,105,109,116,123 and 129DAP;
in 2005:84,88,93,98,101,106,109,112 and 115 DAP) and curve
characteristics (onset, ipoint, prate) are presented. The colors in
the linkage groups indicate the significance levels (white:
P [ 0.005, yellow: 0.001 \ P B 0.005, brown: P B 0.001)
Fig. 3 continued
b
Euphytica (2012) 183:289–302 297
123

Therefore, the full model was the same for prate,
ipoint and onset and it was set according to model (4)
including only two-way interactions betweenQTLs:
y ¼E3þ Ch5þ E7 þ C1þ C4þ C5A
þ E3� Ch5þ E3� E7 þ Ch5� E7
þ C1� C4þ C1� C5Aþ C4� C5A
þ E3� C1þ E3� C4þ E3� C5A
þ Ch5� C1þ Ch5� C4þ Ch5� C5A
þ E7 � C1þ E7 � C4þ E7 � C5Aþ error
QTL main effects, QTL9QTL interactions and
their corresponding genotypic classes for prate,
ipoint and onset are presented in Table 2. To go
from the full models to the final models, only
significant terms in the full models were retained
(P \ 0.05). The P-values for the retained terms and
adjusted R2 in the final models for each curve
characteristic are presented in Table 3. For instance,
after fitting the full model for prate in 2004, 6
terms were retained in the final model (Table 2):
two epistatic interactions (E3 9 C5a and Ch5 9
C4), one main effect QTL (Ch5) and three non-
significant main effect QTLs involved in the
epistatic interactions (E3, C4 and C5a). In the final
model, only the interaction Ch5 9 C4 showed to be
significant and the main effect of Ch5 and C5a
(Table 3).
In 2004 and 2005, a main effect of Ch5 on prate,
ipoint and onset was observed. It is related to a QTL
in the same region previously reported as associated
with plant maturity type (Celis-Gamboa 2002), which
is a trait reflecting the general development of the
plant.
In 2005 no epistatic interactions were found
between the QTLs but pleiotropic effects of E3 on
ipoint and onset, and C4 on prate and onset (Table 3).
In 2004, C4 associated with onset of senescence
interacted epistatically with Ch5 associated with
prate and ipoint (Table 3). In the interaction
C4 9 Ch5, eight genotypic classes are present (‘aa
ee’, ‘aa ef’, ‘aa eg’, ‘aa fg’, ‘ab ee’, ‘ab ef’, ‘ab eg’,
‘ab fg’) as shown in Table 2 and the average
senescence curves per class are presented in Fig. 4.
The genotypic classes including ‘ab’ (fitted curves
with dashed line) had a faster onset and senescence
development than the classes in which ‘aa’ (fitted
curves with solid line) was present. The curve ‘ab ee’
showed a higher prate than ‘aa ee’ and the genotypes
in this class had in general a faster onset of
senescence and died earlier. The classes ‘aa fg’ and
‘ab fg’ were associated with late senescence devel-
opment and the prate and ipoint of these two classes
were the lowest. Genotypes in these two classes were
at almost half of the maximum senescence when the
observation period ended. Genotypes in the class ‘aa
fg’ showed the latest senescence development among
all the genotypic classes. Interestingly the class ‘ab
fg’ showed the earliest onset but this was followed by
a delayed senescence and the genotypes in this group
had the latest senescence.
Another epistatic interaction was observed in 2004
between C4 associated with onset and E7 related with
ipoint (Table 3). Figure 5 shows the average senes-
cence curves observed for the interaction C4 9 E7
with eight genotypic classes (‘aa ii’, ‘aa ij’, ‘aa ik’,
‘aa jk’, ‘ab ii’, ‘ab ij’, ‘ab ik’, ‘ab jk’) as shown in
Table 2. The classes including ‘ab’ (fitted curves with
dotted line) had in general, a faster senescence
development than classes in which ‘aa’ (solid line)
was present except for the class ‘aa ik’. In this class
earlier onset of the senescence process was observed
with a faster prate and ipoint while in the group ‘ab
ik’ a delayed onset with a slower prate is present.
Interestingly, big differences were also observed
between the classes ‘aa jk’ and ‘ab jk’. The class
‘aa jk’ showed the same trajectory than the class ‘aa
ii’ and both of them have late onset and ipoint making
the senescence process slower than in the other
classes. On the other hand, the class ‘ab jk’ showed
the earliest onset with a delayed senescence that
ended up simultaneously with the classes exhibiting
the slowest prate (‘aa ii’ and ‘aa jk’).
Discussion
The smoothed GLM procedure in our study allows
flexible modeling of different senescence curves in
the mapping population. Genotypes with early devel-
opment showed an s-shape senescence curve,
whereas genotypes with slow development exhibited
an exponential curve during the observation period.
In both cases the curve trajectories would eventually
reach the maximum of 7, which is the highest
senescence score. In the case of late genotypes, the
observation period was not long enough to complete
298 Euphytica (2012) 183:289–302
123

the senescence process. Four characteristics esti-
mated from the curves (onset, prate, mprate and
ipoint) allowed us to explain senescence in terms of
development. Whereas individual time points only
gave us an impression of the trait at selected different
moments during the growing season. In the first
case sensible biological interpretations could be
made and the results were compared with previous
studies describing the senescence process in the same
population evaluated in the Netherlands (Celis-
Gamboa 2002).
The weather and the general conditions of the
experiment had a clear effect on the performance of
the population in the 2 years of field experimentation
in Finland. Clearly the senescence period lasted
longer in 2005 and showed a slower progression
rate. This was probably due to the higher daily
air temperature observed that year. Marinus and
Table 2 Main effect QTLs and QTL 9 QTL interactions included in the full models for prate, ipoint and onset of senescence in
2004 and 2005
Variable Segregation typea Possible genotypic classesb 2004 2005
prate ipoint onset prate ipoint onset
E3 ‘‘nn 9 np’’ nn,np 0.86 0.032 0.003 0.856 0.006 0.011
Ch5 ‘‘ef 9 eg’’ ee,ef,e.g,fg \0.001 \0.001 0.007 \0.001 \0.001 \0.001
E7 ‘‘ij 9 ik’’ ii,ij,ik,jk 0.886 0.007 0.021 0.223 0.056 0.004
C1 ‘‘ab 9 aa’’ aa,ab 0.841 0.614 0.719 0.03 0.034 0.969
C4 ‘‘ab 9 aa’’ aa,ab 0.689 0.159 0.007 0.01 0.475 0.006
C5a ‘‘ab 9 aa’’ aa,ab 0.081 0.428 \0.001 0.763 0.076 0.154
E3 9 Ch5 ‘‘nn 9 np’’ 9 ‘‘ef 9 eg’’ nn ee, nn ef, nn e.g, nn fg, np ee,np ef, np e.g, np fg
0.838 0.117 0.293 0.829 0.486 0.563
E3 9 E7 ‘‘nn 9 np’’ 9 ‘‘ij 9 ik’’ nn ii, nn ij, nn ik, nn jk, np ii, npij, np ik, np jk
0.44 0.943 0.427 0.264 0.61 0.471
Ch5 9 E7 ‘‘ef 9 eg’’ 9 ‘‘ij 9 ik’’ ee ii, ee ij, ee ik, ee jk, ef ii, ef ij,ef ik, ef jk, e.g ii, e.g ij, e.g ik,e.g jk fg ii, fg ij, fg ik, fg jk
0.658 0.121 0.161 0.399 0.561 0.515
C1 9 C4 ‘‘ab 9 aa’’ 9 ‘‘ab 9 aa’’ aa aa, aa ab, ab aa, ab ab 0.93 0.178 0.139 0.566 0.398 0.925
C1 9 C5a ‘‘ab 9 aa’’ 9 ‘‘ab 9 aa’’ aa aa, aa ab, ab aa, ab ab 0.267 0.789 0.269 0.115 0.891 0.744
C4 9 C5a ‘‘ab 9 aa’’ 9 ‘‘ab 9 aa’’ aa aa, aa ab, ab aa, ab ab 0.302 0.701 0.264 0.426 0.075 0.325
E3 9 C1 ‘‘nn 9 np’’ 9 ‘‘ab 9 aa’’ nn aa, nn ab, np aa, np ab 0.392 0.33 0.62 0.396 0.75 0.906
E3 9 C4 ‘‘nn 9 np’’ 9 ‘‘ab 9 aa’’ nn aa, nn ab, np aa, np ab 0.373 0.701 0.895 0.909 0.784 0.574
E3 9 C5a ‘‘nn 9 np’’ 9 ‘‘ab 9 aa’’ nn aa, nn ab, np aa, np ab 0.019 0.657 0.037 0.5 0.366 0.922
Ch5 9 C1 ‘‘ef 9 eg’’ 9 ‘‘ab 9 aa’’ aa ee, aa ef, aa e.g, aa fg, ab ee,ab ef, ab e.g, ab fg
0.989 0.805 0.793 0.704 0.397 0.385
Ch5 9 C4 ‘‘ef 9 eg’’ 9 ‘‘ab 9 aa’’ aa ee, aa ef, aa e.g, aa fg, ab ee,ab ef, ab e.g, ab fg
0.043 0.039 0.47 0.022 0.736 0.977
Ch5 9 C5a ‘‘ef 9 eg’’ 9 ‘‘ab 9 aa’’ aa ee, aa ef, aa e.g, aa fg, ab ee,ab ef, ab e.g, ab fg
0.247 0.479 0.553 0.962 0.203 0.669
E7 9 C1 ‘‘ij 9 ik’’ 9 ‘‘ab 9 aa’’ aa ii, aa ij, aa ik, aa jk, ab ii, abij, ab ik, ab jk
0.414 0.566 0.959 0.904 0.57 0.948
E7 9 C4 ‘‘ij 9 ik’’ 9 ‘‘ab 9 aa’’ aa ii, aa ij, aa ik, aa jk, ab ii, abij, ab ik, ab jk
0.601 0.105 0.02 0.636 0.08 0.204
E7 9 C5a ‘‘ij 9 ik’’ 9 ‘‘ab 9 aa’’ aa ii, aa ij, aa ik, aa jk, ab ii, abij, ab ik, ab jk
0.294 0.475 0.88 0.691 0.265 0.708
Adjusted
R231.5 47.7 29.4 13.9 49.5 30.2
a Segregation type codes for a CP population according to MapQTL 6 (Van Ooijen 2009)b Genotype codes for a CP population, depending on the locus segregation type (Van Ooijen 2009) and the QTL 9 QTL interaction
Euphytica (2012) 183:289–302 299
123
Boadlaender (1975) reported that high temperatures
accelerate the reproductive and vegetative develop-
ment of potato plants and delay the senescence
process. Our results are therefore in line with their
findings.
Comparing the QTL analysis using the individual
time points and the curve characteristics, QTLs
detected at early time points were associated with
onset of senescence. QTLs occurring in intermediate
to late time points coincided with QTLs associated
with prate and ipoint. Although the results are
comparable with both phenotypic data, the analysis
using the characteristics of fitted curves offers a
biological interpretation of the results in terms of
senescence development. In our study, four curve
characteristics described the whole process and
allowed us to study the progression of the trait.
When the C 9 E population was evaluated in the
Netherlands, QTLs on chromosome 5 were identified
for different traits related with senescence and life
cycle. Malosetti et al. (2006) reported two QTLs on
chromosome 5 associated with mid-senescence and
rate of senescence. Celis-Gamboa (2002) identified
three QTLs on the same chromosome associated with
the duration of the plant cycle and one of them was
found associated to plant maturity type explaining
40% of the phenotypic variance. In this study, we
detected QTLs associated with senescence develop-
ment on chromosome 5. These are in the same region
where the latter QTL was reported by Celis-Gamboa
(2002), suggesting a pleiotropic effect of this region
for prate, ipoint, onset and plant maturity.
An interesting QTL associated with onset was
found on C4, whereas no QTLs have been previously
reported for developmental traits on this chromo-
some. A wide range of functional genes involved in
resistance against various pathogens have been
reported on chromosome 4, among others a major
late blight resistance gene cluster (Celebi-Toprak
et al. 2002; Li et al. 1998; Park et al. 2005; Zimnoch-
Guzowska et al. 2000).
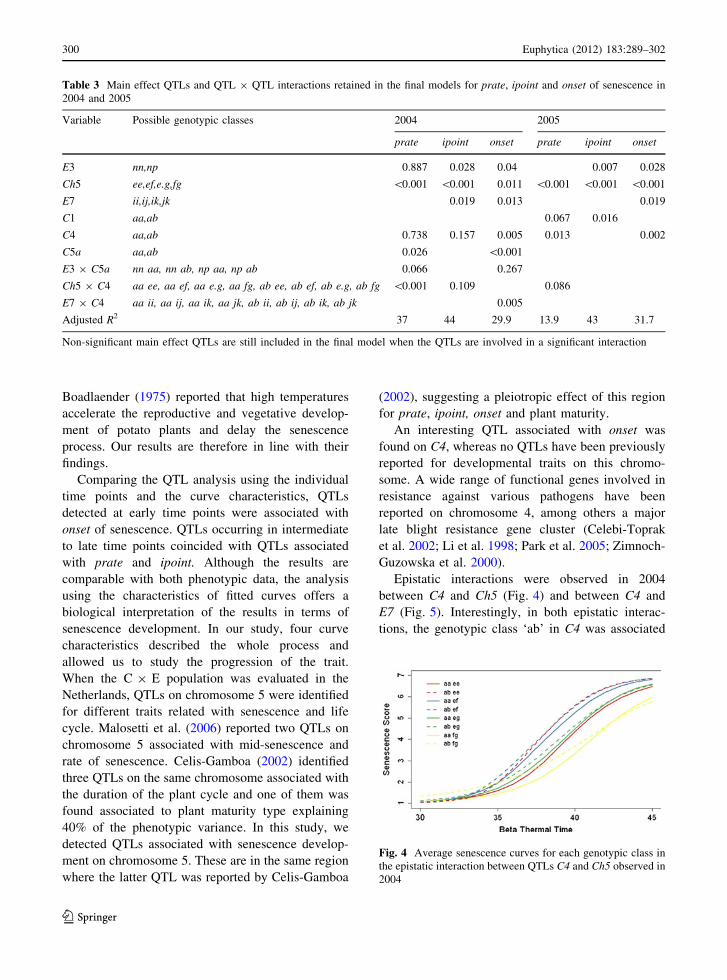
Epistatic interactions were observed in 2004
between C4 and Ch5 (Fig. 4) and between C4 and
E7 (Fig. 5). Interestingly, in both epistatic interac-
tions, the genotypic class ‘ab’ in C4 was associated
Table 3 Main effect QTLs and QTL 9 QTL interactions retained in the final models for prate, ipoint and onset of senescence in
2004 and 2005
Variable Possible genotypic classes 2004 2005
prate ipoint onset prate ipoint onset
E3 nn,np 0.887 0.028 0.04 0.007 0.028
Ch5 ee,ef,e.g,fg \0.001 \0.001 0.011 \0.001 \0.001 \0.001
E7 ii,ij,ik,jk 0.019 0.013 0.019
C1 aa,ab 0.067 0.016
C4 aa,ab 0.738 0.157 0.005 0.013 0.002
C5a aa,ab 0.026 \0.001
E3 9 C5a nn aa, nn ab, np aa, np ab 0.066 0.267
Ch5 9 C4 aa ee, aa ef, aa e.g, aa fg, ab ee, ab ef, ab e.g, ab fg \0.001 0.109 0.086
E7 9 C4 aa ii, aa ij, aa ik, aa jk, ab ii, ab ij, ab ik, ab jk 0.005
Adjusted R2 37 44 29.9 13.9 43 31.7
Non-significant main effect QTLs are still included in the final model when the QTLs are involved in a significant interaction
Fig. 4 Average senescence curves for each genotypic class in
the epistatic interaction between QTLs C4 and Ch5 observed in
2004
300 Euphytica (2012) 183:289–302
123

with early development of the senescence process. In
the interaction C4 9 Ch5, the genotypic classes ‘ab
ee’ and ‘ab ef’ associated with early senescence
showed delayed onset but fast prate and ipoint
whereas the class ‘ab fg’’ showed early onset but a
senescence period lasting longer. In 2005 no epistatic
interactions were observed but main effects of C1 and
pleiotropic effects of E3 and C4 on ipoint/onset and
prate/onset, respectively.
In summary, by modeling haulm senescence in the
C 9 E potato population using a smoothed GLM
procedure, we were able to characterize the curve in
terms of developmental traits and to identify QTLs
associated with the senescence process. Pleiotropic
effects and epistatic interactions between QTLs were
detected when two-way interactions were studied.
Delayed senescence was associated with particular
genotypic classes (C4 9 Ch5: ‘ab fg’ and C4 9 E7:
‘aa ii’ ‘aa jk’) and some classes showing early onset
turned to have delayed senescence development
(C4 9 Ch5: ‘ab fg’ and C4 9 E7: ‘ab jk’).
Different regulatory genes have been reported for
onset and progression rate of senescence in Arabid-
opsis thaliana (Gepstein et al. 2003) and it will be
interesting to compare the genes involved in the same
process in different crops. The complexity of
QTL 9 E and QTL 9 QTL 9 E as well as the
performance of genotypes under different environ-
ments will be considered in a further study providing
insights for the better understanding of adaptation and
developmental processes in potato.
Acknowledgments We thank Prof. Paul Struik (Centre for
Crop Systems Analysis, Wageningen University) for the
discussion on the use of beta-thermal time in this study. Part
of this project was (co)financed by the Centre for BioSystems
Genomics (CBSG) which is part of the Netherlands Genomics
Initiative/Netherlands Organisation for Scientific Research.
Open Access This article is distributed under the terms of the
Creative Commons Attribution Noncommercial License which
permits any noncommercial use, distribution, and reproduction
in any medium, provided the original author(s) and source are
credited.
References
Allaby M (1998) A dictionary of plant sciences. Encyclope-
dia.com. http://www.encyclopedia.com. Accessed 30
November 2009
Benoit RG, Grant WJ, Devine OJ (1986) Potato top growth as
influenced by day–night temperature differences. Agron J
78:264–269
Celebi-Toprak F, Slack SA, Jahn MM (2002) A new gene,
Nytbr, for hypersensitivity to Potato virus Y from Solanumtuberosum maps to chromosome IV. Theor Appl Genet
104:669–674
Celis-Gamboa BC (2002) The life cycle of the potato (Solanumtuberosum L.): from crop physiology to genetics. PhD
Thesis. Wageningen University, Wageningen
Celis-Gamboa C, Struik PC, Jacobsen E, Visser RGF (2003)
Temporal dynamics of tuber formation and related
processes in a crossing population of potato (Solanumtuberosum). Ann Appl Biol 143:175–186
Eilers PHC, Marx BD (1996) Flexible smoothing with
B-splines and penalties. Stat Sci 11(2):89–121
Ewing EE, Struik PC (1992) Tuber formation in potato:
induction, initiation and growth. Hortic Rev 14:89–198
Gao LZ, Jin ZQ, Huang Y, Zhang LZ (1992) Rice clock
model—a computer simulation model of rice develop-
ment. Agric For Meteorol 60:1–16
Gepstein S, Sabehi G, Carp M-J, Hajouj T, Nesher MFO, Yariv
I, Dor C, Bassani M (2003) Large-scale identification of
leaf senescence-associated genes. Plant J 36:629–642
Hanneman RE, Peloquin SJ (1967) Crossability of 24 chro-
mosome potato hybrids with 48 chromosome cultivars.
Eur Potato J 10:62–73
Haun JR (1975) Potato growth-environment relationship. Agric
Meteorol 15:325–332
Hawkes JG (1990) The potato: evolution, biodiversity and
genetic resources. Belhaven press, London, p 259
Hodges T (1991) Temperature and water stress effects on
phenology. In: Hodges T (ed) Predicting crop phenology.
CRC Press, Boca Raton, pp 7–13
Jacobsen E (1980) Increase of diplandroid formation and seed
set in 4x 9 2x crosses in potatoes by genetical manipu-
lation of dihaploids and some theoretical consequences.
Z Pflanzenzuecht 85:110–121
Johnson IR, Thornley JHM (1985) Temperature dependence of
plant and crop processes. Ann Bot 55:l–24
Khan MS, Struik PC, van der Putten PEL, van Eck HJ, van
Eeuwijk FA, Yin X (2011) Analyzing genetic variation in
Fig. 5 Average senescence curves for each genotypic class in
the epistatic interaction between QTLs C4 and E7 observed in
2004
Euphytica (2012) 183:289–302 301
123

potato (Solanum tuberosum) canopy dynamics using
standard cultivars and a segregating population. (Under
review)
Li X, van Eck HJ, Rouppe van der Voort JNAM, Huigen DJ,
Stam P, Jacobsen E (1998) Autotetraploids and genetic
mapping using common AFLP markers: the R2 allele
conferring resistance to Phytophthora infestans mapped
on potato chromosome 4. Theor Appl Genet 96:
1121–1128
Ma C-X, Casella G, Wu R (2002) Functional mapping of
quantitative trait loci underlying the character process: a
theoretical framework. Genetics 161:1751–1762
Malosetti M, Visser RGF, Celis-Gamboa C, van Eeuwijk FA
(2006) QTL methodology for response curves on the basis
of non-linear mixed models, with an illustration to
senescence in potato. Theor Appl Genet 113:288–300
Marinus J, Boadlaender KBA (1975) Response of some potato
varieties to temperature. Potato Res 18:189–204
Nelder JA, Wedderburn RWM (1972) Generalized linear
models. J Royal Stat Soc, Series A 135:370–384
Park TH, Gros A, Sikkema A, Vleeshouwers VGAA, Muskens
M, Allefs S, Jacobsen E, Visser RGF, van der Vossen
EAG (2005) The late blight resistance locus Rpi-blb3from Solanum bulbocastanum belongs to a major late
blight R gene cluster on chromosome 4 of potato. Mol
Plant Microbe Interact 18:722–729
Payne RW, Murray DA, Harding SA, Baird DB, Soutar DM
(2010) An introduction to GenStat for Windows, 13th edn.
VSN International, Hemel Hempstead, UK
Reymond M, Muller B, Leonardi A, Charcosset A, Tardieu F
(2003) Combining quantitative trait loci analysis and an
ecophysiological model to analyze the genetic variability
of the responses of maize leaf growth to temperature and
water deficit. Plant Physiol 131:664–675
Schnute J (1981) A versatile growth model with statistically
stable parameters. Can J Fish Aquat Sci 38:1128–1140
Struik PC, Ewing EE (1995) Crop physiology of potato:
responses to photoperiod and temperature relevant to crop
modeling. In: Haverkort AJ, MacKerron DKL (eds)
Potato ecology and modeling of crops under conditions
limiting growth. Kluwer Acad Publ, Boston, pp 19–40
van Ooijen JW (2006) JoinMap 4, Software for the calculation
of genetic linkage maps in experimental populations.
Kyazma B.V, Wageningen, the Netherlands
van Ooijen JW (2009) MapQTL 6, Software for the mapping of
quantitative trait loci in experimental populations of dip-
loid species. Kyazma B.V, Wageningen, the Netherlands
Werij JS, Kloosterman B, Celis-Gamboa C, de Vos CHR,
America T, Visser RGF, Bachem C (2007) Unravelling
enzymatic discoloration in potato through a combined
approach of candidate genes, QTL, and expression anal-
ysis. Theor Appl Genet 115:245–252
Wu RL, Lin M (2006) Functional mapping—how to map and
study the genetic architecture of dynamic complex traits.
Nat Rev Gene 7:229–237
Wu RL, Ma CX, Zhao W, Casella G (2003) Functional map-
ping of quantitative trait loci underlying growth rates: a
parametric model. Physiol Genomics 14:241–249
Yin X, Kropff MJ, McLaren G, Visperas RM (1995) A non-
linear model for crop development as a function of tem-
perature. Agric For Meteorol 77:1–16
Zaban A, Vetelainen M, Celis-Gamboa C, van Berloo R,
Haggman H, Visser RGF (2006) Physiological and
genetic aspects of a diploid potato population in the
Netherlands and Northern Finland. Maataloustieteen Pai-
vat 2006:1–7
Zimnoch-Guzowska E, Marczewski E, Lebecka R, Flis B,
Schafer-Pregl R, Salamini F, Gebhardt C (2000) QTL
analysis of new sources of resistance to Erwinia caroto-vora ssp. atroseptica in potato done by AFLP, RFLP, and
resistance-gene-like markers. Crop Sci 40:1156–1167
302 Euphytica (2012) 183:289–302
123