dynamic spoiiie assembly mediates septal membrane...
TRANSCRIPT

10.1101/gad.1925210Access the most recent version at doi: 2010 24: 1160-1172Genes Dev.
Tinya C. Fleming, Jae Yen Shin, Sang-Hyuk Lee, et al.
sporulationBacillus subtilisduring Dynamic SpoIIIE assembly mediates septal membrane fission
MaterialSupplemental http://genesdev.cshlp.org/content/suppl/2010/05/26/24.11.1160.DC1.html
References http://genesdev.cshlp.org/content/24/11/1160.full.html#ref-list-1
This article cites 33 articles, 13 of which can be accessed free at:
serviceEmail alerting
click heretop right corner of the article orReceive free email alerts when new articles cite this article - sign up in the box at the
http://genesdev.cshlp.org/subscriptions go to: Genes & DevelopmentTo subscribe to
Copyright © 2010 by Cold Spring Harbor Laboratory Press
Cold Spring Harbor Laboratory Press on June 13, 2010 - Published by genesdev.cshlp.orgDownloaded from

Dynamic SpoIIIE assembly mediatesseptal membrane fission duringBacillus subtilis sporulation
Tinya C. Fleming,1 Jae Yen Shin,2,3 Sang-Hyuk Lee,2,4 Eric Becker,1 Kerwyn Casey Huang,5
Carlos Bustamante,2,3,4,6,7 and Kit Pogliano1,8
1Division of Biological Sciences, University of California at San Diego, La Jolla, California 92093, USA; 2Department of Physics,University of California at Berkeley, Berkeley, California 94720, USA; 3Howard Hughes Medical Institute, University ofCalifornia at Berkeley, Berkeley, California 94720, USA; 4Physical Bioscience Division, Lawrence Berkeley National Laboratory,Berkeley, California 94720, USA; 5Department of Bioengineering, Stanford University, Stanford, California 94305, USA;6Department of Molecular and Cell Biology, University of California at Berkeley, Berkeley, California 94720, USA;7Department of Chemistry, University of California at Berkeley, Berkeley, California 94720, USA
SpoIIIE is an FtsK-related protein that transports the forespore chromosome across the Bacillus subtilissporulation septum. We use membrane photobleaching and protoplast assays to demonstrate that SpoIIIE isrequired for septal membrane fission in the presence of trapped DNA, and that DNA is transported across separatedaughter cell membranes, suggesting that SpoIIIE forms a channel that partitions the daughter cell membranes.Our results reveal a close correlation between septal membrane fission and the assembly of a stable SpoIIIEtranslocation complex at the septal midpoint. Time-lapse epifluorescence, total internal reflection fluorescence(TIRF) microscopy, and live-cell photoactivation localization microscopy (PALM) demonstrate that the SpoIIIEtransmembrane domain mediates dynamic localization to active division sites, whereas the assembly of a stablefocus also requires the cytoplasmic domain. The transmembrane domain fails to completely separate themembrane, and it assembles unstable foci. TIRF microscopy and biophysical modeling of fluorescence recoveryafter photobleaching (FRAP) data suggest that this unstable protein transitions between disassembled andassembled oligomeric states. We propose a new model for the role of SpoIIIE assembly in septal membrane fissionthat has strong implications for how the chromosome terminus crosses the septum.
[Keywords: SpoIIIE; DNA translocase; membrane fission; FRAP; PALM]
Supplemental material is available at http://www.genesdev.org.
Received March 12, 2010; revised version accepted April 15, 2010.
The final step of cell division is the separation of daughtercell membranes (septal membrane fission). Althoughseptal membrane fission is universally conserved in allcellular organisms, it remains poorly understood, partic-ularly in prokaryotes. Septal membrane fission in plantand animal cells requires vesicular transport, and it islikely that fusion of vesicles that span the intracellularbridge completes cell division (Finger and White 2002;Jurgens 2005). Bacterial division commences with mem-brane constriction, as in animal cells, but there is noevidence for vesicular transport in bacteria, suggestingthat they mediate septal membrane fission by a funda-mentally different mechanism than eukaryotes.Bacillus subtilis sporulation provides a unique oppor-
tunity to study two distinct membrane fission events in
a bacterium. First, at the onset of sporulation, the celldivides near the cell pole and traps one chromosome inthe septum (Fig. 1A). Septal membrane fission is thereforecomplicated by this trapped chromosome, which is trans-located into the forespore by the SpoIIIE DNA translocase(Wu and Errington 1994; Errington 2001). Next, the mem-brane of the larger mother cell migrates around thesmaller forespore in a phagocytosis-like process calledengulfment (for review, see Errington 2003; Hilbert andPiggot 2004). Ultimately the leading edges of the mothercell membrane converge on the distal side of the fore-spore, and a second membrane fission event pinches off adetached, internalized forespore (Sharp and Pogliano1999).Prior data suggested that bothmembrane fission events
of sporulation involve the SpoIIIE DNA translocase.SpoIIIE is required for cytoplasmic compartmentalizationof the two daughter cells (Wu and Errington 1994; Liuet al. 2006) and for engulfment membrane fission (Sharp
8Corresponding author.E-MAIL [email protected]; FAX (858) 822-5740.Article is online at http://www.genesdev.org/cgi/doi/10.1101/gad.1925210.
1160 GENES & DEVELOPMENT 24:1160–1172 ! 2010 by Cold Spring Harbor Laboratory Press ISSN 0890-9369/10; www.genesdev.org
Cold Spring Harbor Laboratory Press on June 13, 2010 - Published by genesdev.cshlp.orgDownloaded from
and Pogliano 1999, 2003; Liu et al. 2006), suggesting thatSpoIIIE might play a general role in membrane fissionduring sporulation. The localization of SpoIIIE to both theseptal midpoint (the site of DNA translocation) and to thepole of the engulfed forespore before engulfment mem-brane fission indicates that the protein is in the rightplace at the right time to directly participate in bothevents (Sharp and Pogliano 1999;Wu and Errington 1997).
In addition, genetic studies have demonstrated that DNAtranslocation depends primarily on the cytoplasmicmotor domain, while compartmentalization and engulf-ment membrane fission depend primarily on theN-terminal transmembrane domain (Sharp and Pogliano1999, 2003; Liu et al. 2006), indicating that these twofunctions are genetically separable. The close correlationbetween defects in cytoplasmic compartmentalization
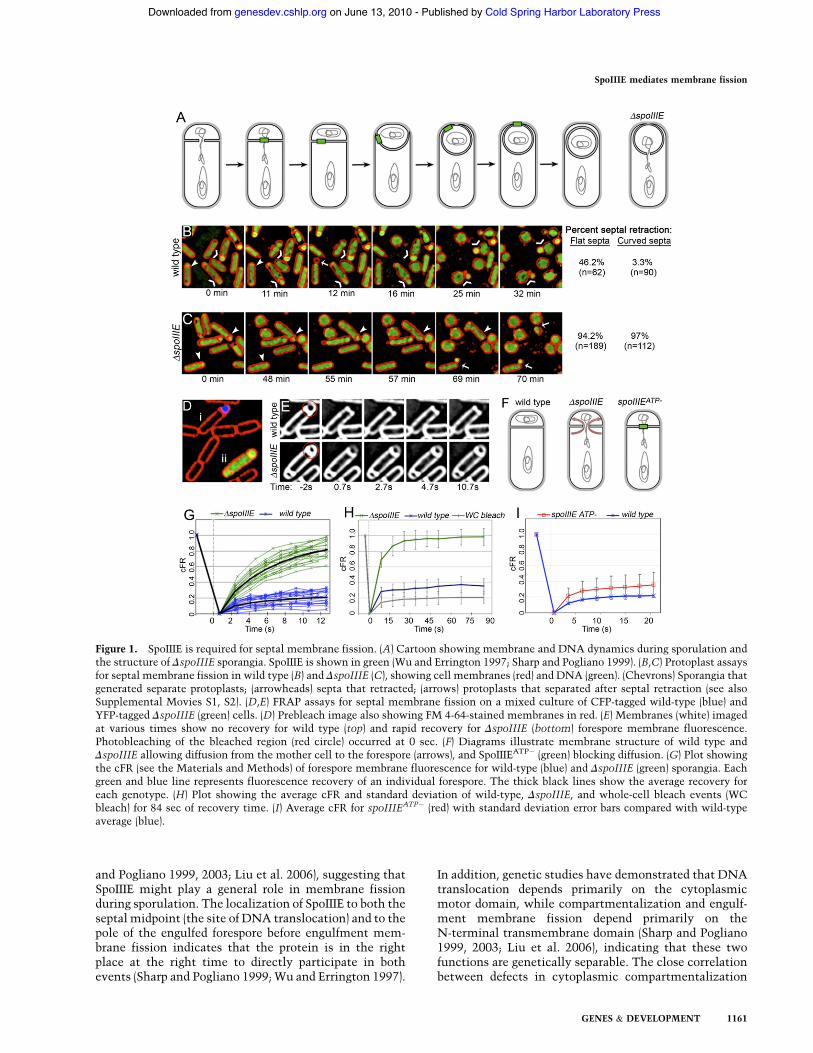
Figure 1. SpoIIIE is required for septal membrane fission. (A) Cartoon showing membrane and DNA dynamics during sporulation andthe structure of DspoIIIE sporangia. SpoIIIE is shown in green (Wu and Errington 1997; Sharp and Pogliano 1999). (B,C) Protoplast assaysfor septal membrane fission in wild type (B) and DspoIIIE (C), showing cell membranes (red) and DNA (green). (Chevrons) Sporangia thatgenerated separate protoplasts; (arrowheads) septa that retracted; (arrows) protoplasts that separated after septal retraction (see alsoSupplemental Movies S1, S2). (D,E) FRAP assays for septal membrane fission on a mixed culture of CFP-tagged wild-type (blue) andYFP-tagged DspoIIIE (green) cells. (D) Prebleach image also showing FM 4-64-stained membranes in red. (E) Membranes (white) imagedat various times show no recovery for wild type (top) and rapid recovery for DspoIIIE (bottom) forespore membrane fluorescence.Photobleaching of the bleached region (red circle) occurred at 0 sec. (F) Diagrams illustrate membrane structure of wild type andDspoIIIE allowing diffusion from the mother cell to the forespore (arrows), and SpoIIIEATP! (green) blocking diffusion. (G) Plot showingthe cFR (see the Materials and Methods) of forespore membrane fluorescence for wild-type (blue) and DspoIIIE (green) sporangia. Eachgreen and blue line represents fluorescence recovery of an individual forespore. The thick black lines show the average recovery foreach genotype. (H) Plot showing the average cFR and standard deviation of wild-type, DspoIIIE, and whole-cell bleach events (WCbleach) for 84 sec of recovery time. (I) Average cFR for spoIIIEATP! (red) with standard deviation error bars compared with wild-typeaverage (blue).
SpoIIIE mediates membrane fission
GENES & DEVELOPMENT 1161
Cold Spring Harbor Laboratory Press on June 13, 2010 - Published by genesdev.cshlp.orgDownloaded from

and in engulfment membrane fission suggested that thecytoplasmic compartmentalization defect of spoIIIE mu-tants might be due to incomplete septal membranefission (Liu et al. 2006).Based on these results, we proposed that, when the
sporulation septum completes constriction, SpoIIIE as-sembles a transmembrane channel that insulates thenegatively charged DNA from the hydrophobic lipidbilayer and its hydrophilic head groups. One model forthis structure is a paired transmembrane channel withsubunits in both daughter cell membranes encircling thetrapped DNA strands (Liu et al. 2006), which could alsomediate the temporary partitioning of the forespore andmother cell membrane during DNA translocation. In sup-port of this model, a recent study found that wild-typesporangia displayed compartmentalized forespore mem-branes during DNA translocation, and that a translocase-defective, localization-proficient spoIIIE mutant main-tained partitioned membranes when DNA traversed theseptum (Burton et al. 2007). However, this study alsoconcluded that SpoIIIE was not required for septal mem-brane fission, based on observations suggesting a DspoIIIEmutant showed normal septal membrane fission (Burtonet al. 2007).Here we confirm the observation that SpoIIIE maintains
membrane compartmentalization during DNA transloca-tion. However, protoplast and fluorescence recovery afterphotobleaching (FRAP) assays demonstrate that, in theabsence of SpoIIIE, the forespore and mother cell mem-branes remain contiguous, indicating defects in septalmembrane fission. Biophysical modeling of lipid diffusionindicates that FRAP can be used as a quantitative assay formembrane geometry and compartmentalization. In addi-tion, we compare FRAP results with assessments of SpoIIIEassembly by several microscopy techniques and finda correlation between the ability of SpoIIIE to assembleand its ability to partition daughter cell membranes. Ourdata indicate that SpoIIIE assembly is a multistep process,with initial dynamic localization to sites of active celldivision mediated by the transmembrane domain, fol-lowed by the assembly of a stable translocation complex,which requires both the transmembrane and the cyto-plasmic motor domain. These results resolve conflictingobservations regarding the role of the transmembraneand motor domains in SpoIIIE assembly, and providemechanistic insight into septal membrane fission duringsporulation.
Results
SpoIIIE is required for septal membrane fissionduring sporulation
We used two methods to determine if SpoIIIE is requiredfor septal membrane fission. First, we treated cells withlysozyme to digest peptidoglycan, which causes rod-shaped Bacillus cells to become spherical protoplasts. Ifseptal membrane fission is complete, the forespore willform a separate protoplast from the mother cell. If septalmembrane fission is incomplete, the septum will retract
as peptidoglycan is digested. This protoplast assayshowed that 46% of flat septa in wild-type cells retract(Fig. 1B, arrowhead), suggesting that many had incom-plete septa with septal openings smaller than the resolu-tion limit of epifluorescence microscopy. Engulfmentstarts after septation, and is readily visualized by thecurving of the septum around the forespore. We foundthat just 3% of curved wild-type septa retracted, confirm-ing that septation was incomplete in very few cells. Incontrast, 97% of curved septa in DspoIIIE cells retracted(Fig. 1C, arrowheads). Thus, even after the onset ofengulfment, most DspoIIIE sporangia fail to completeseptal membrane fission.Protoplast assays also appeared to subject cells to phys-
ical stresses that can pinch off several separated pro-toplasts from a single cell. Specifically, some sporangiafirst show septal retraction and later pinch off a forespore-sized protoplast at the forespore pole (Fig. 1B,C, arrows).We therefore used a second, more direct method: stainingcells with the membrane dye FM 4-64, and then perform-ing a FRAP analysis by photobleaching only the foresporemembrane (Fig. 1D,E). If septal membrane fission is com-plete, then the forespore membrane fluorescence will failto recover because the dye will be unable to diffuse acrossthe septum. However, if septal membrane fission isincomplete, forespore and mother cell membranes re-main connected, allowing FM 4-64 to diffuse from theunbleached mother cell into the bleached forespore (Fig.1F). In this case, forespore fluorescence should fully re-cover due to equilibration of membrane dye between theforespore and mother cell.We performed simultaneous FRAP of forespore mem-
branes of wild-type sporangia expressing a CFP reporterand DspoIIIE sporangia expressing a YFP reporter toobserve fluorescence recovery side by side (Fig. 1D). Webleached forespores with slightly curved septa to selectsporangia that had completed cell division. Rapid re-covery of forespore fluorescence was observed in onlythe DspoIIIE strain, suggesting that the membranes ofthis strain are contiguous, whereas wild-type membranesare separated (Fig. 1E). Identical results were obtainedwith two published spoIIIE deletions (Wu and Errington1994; Pogliano et al. 1997; Sharp and Pogliano 1999) andwith the complete deletion of spoIIIE constructed for thisstudy (Supplemental Figure S1).We next quantified the results in a manner that allows
the results of many individual cells to be overlayed andthe average recovery and standard deviations to becalculated (Fig. 1G–I). FRAP analysis of wild type showedlimited recovery, with an average recovery of 20% mem-brane fluorescence (corrected fraction recovery [cFR] =0.2) after 12 sec (Fig. 1G) and 35% after 84 sec (Fig. 1H).The whole-cell bleach control also showed a slight re-covery (cFR = 0.2 at 84 sec), perhaps due to exchange offluorescent FM 4-64 from the medium to the cells ordue to reversible photobleaching of some FM 4-64 mol-ecules. The slightly greater fluorescence recovery ofbleached wild-type forespores than bleached whole cellslikely results from incomplete septal bleaching, since theforespore membrane cannot be distinguished from the
Fleming et al.
1162 GENES & DEVELOPMENT
Cold Spring Harbor Laboratory Press on June 13, 2010 - Published by genesdev.cshlp.orgDownloaded from
mother cell membrane at the septum. The failure of wild-type forespores to fully recover indicates that the fore-spore and mother cell membranes are separated (Fig. 1F).In contrast, analysis of DspoIIIE sporangia showed com-plete recovery to 100% of possible fluorescence (cFR = 1)with an average time to half recovery (t1/2) of ;5 sec (Fig.1G–H). This indicates that the forespore and mother cellmembranes remain contiguous in the absence of SpoIIIE(Fig. 1F). Thus, two separate assays demonstrate thatSpoIIIE is required for septal membrane fission duringB. subtilis sporulation.
SpoIIIE is required for septal membrane fission onlyin the presence of trapped DNA
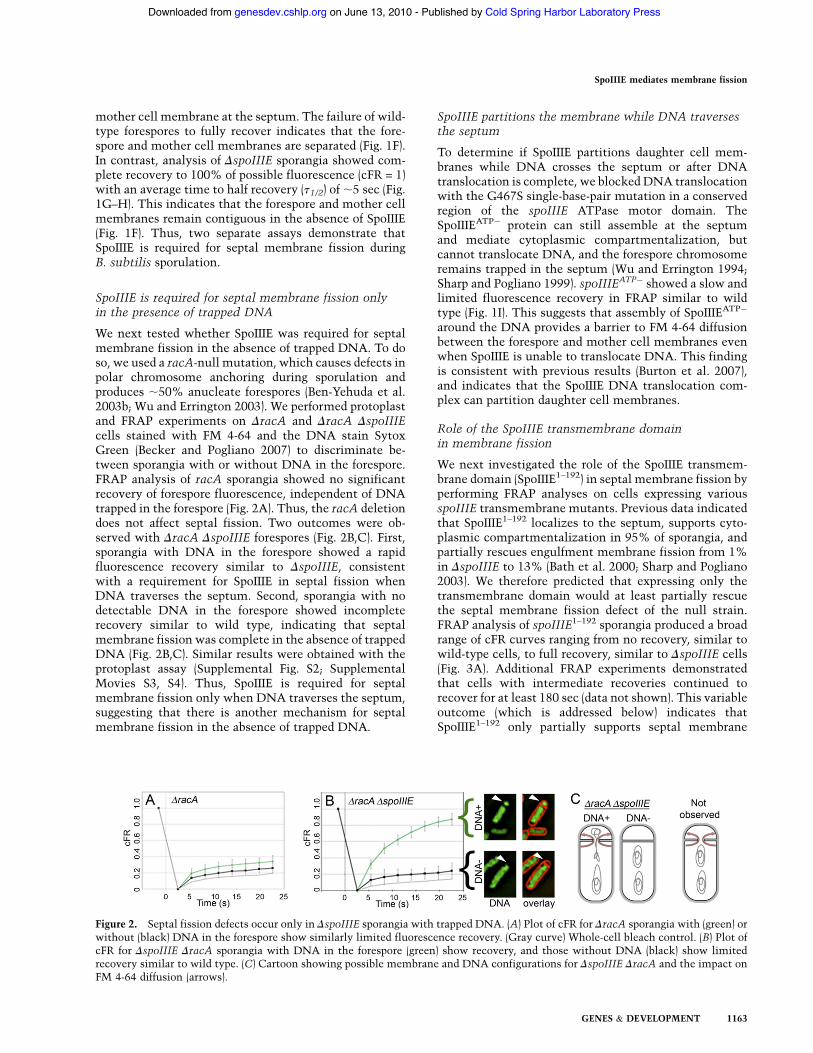
We next tested whether SpoIIIE was required for septalmembrane fission in the absence of trapped DNA. To doso, we used a racA-null mutation, which causes defects inpolar chromosome anchoring during sporulation andproduces ;50% anucleate forespores (Ben-Yehuda et al.2003b; Wu and Errington 2003). We performed protoplastand FRAP experiments on DracA and DracA DspoIIIEcells stained with FM 4-64 and the DNA stain SytoxGreen (Becker and Pogliano 2007) to discriminate be-tween sporangia with or without DNA in the forespore.FRAP analysis of racA sporangia showed no significantrecovery of forespore fluorescence, independent of DNAtrapped in the forespore (Fig. 2A). Thus, the racA deletiondoes not affect septal fission. Two outcomes were ob-served with DracA DspoIIIE forespores (Fig. 2B,C). First,sporangia with DNA in the forespore showed a rapidfluorescence recovery similar to DspoIIIE, consistentwith a requirement for SpoIIIE in septal fission whenDNA traverses the septum. Second, sporangia with nodetectable DNA in the forespore showed incompleterecovery similar to wild type, indicating that septalmembrane fission was complete in the absence of trappedDNA (Fig. 2B,C). Similar results were obtained with theprotoplast assay (Supplemental Fig. S2; SupplementalMovies S3, S4). Thus, SpoIIIE is required for septalmembrane fission only when DNA traverses the septum,suggesting that there is another mechanism for septalmembrane fission in the absence of trapped DNA.
SpoIIIE partitions the membrane while DNA traversesthe septum
To determine if SpoIIIE partitions daughter cell mem-branes while DNA crosses the septum or after DNAtranslocation is complete, we blocked DNA translocationwith the G467S single-base-pair mutation in a conservedregion of the spoIIIE ATPase motor domain. TheSpoIIIEATP! protein can still assemble at the septumand mediate cytoplasmic compartmentalization, butcannot translocate DNA, and the forespore chromosomeremains trapped in the septum (Wu and Errington 1994;Sharp and Pogliano 1999). spoIIIEATP! showed a slow andlimited fluorescence recovery in FRAP similar to wildtype (Fig. 1I). This suggests that assembly of SpoIIIEATP!
around the DNA provides a barrier to FM 4-64 diffusionbetween the forespore and mother cell membranes evenwhen SpoIIIE is unable to translocate DNA. This findingis consistent with previous results (Burton et al. 2007),and indicates that the SpoIIIE DNA translocation com-plex can partition daughter cell membranes.
Role of the SpoIIIE transmembrane domainin membrane fission
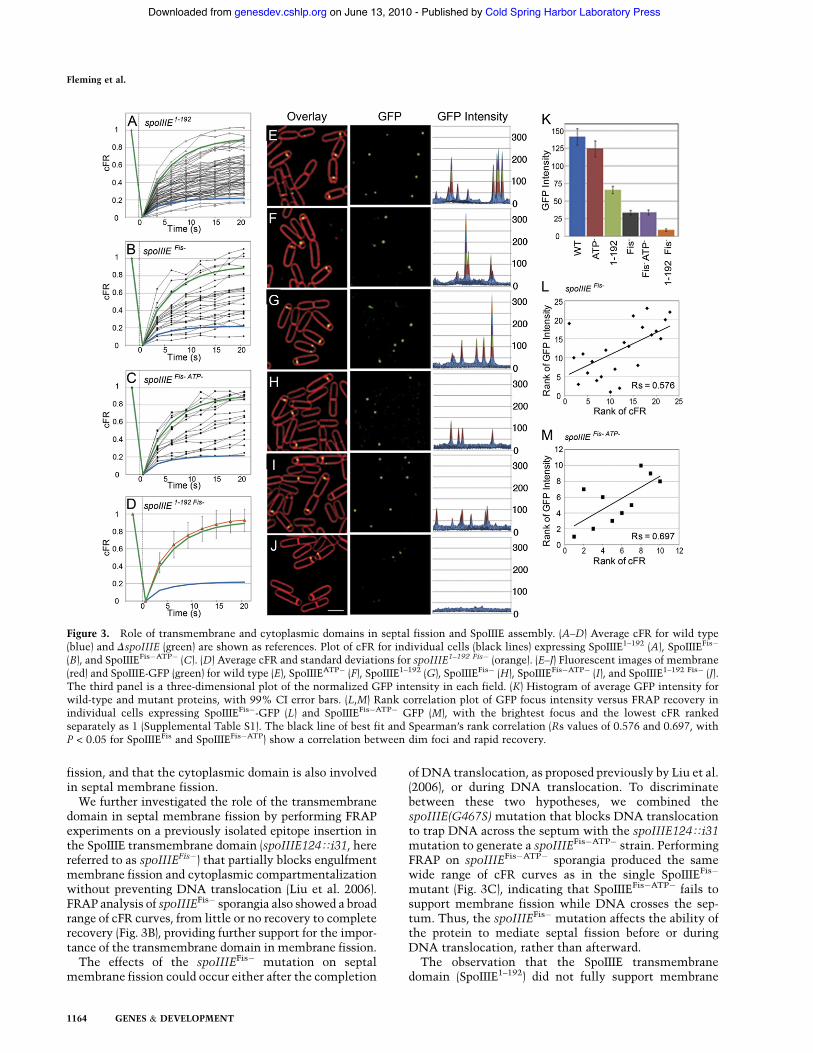
We next investigated the role of the SpoIIIE transmem-brane domain (SpoIIIE1–192) in septal membrane fission byperforming FRAP analyses on cells expressing variousspoIIIE transmembrane mutants. Previous data indicatedthat SpoIIIE1–192 localizes to the septum, supports cyto-plasmic compartmentalization in 95% of sporangia, andpartially rescues engulfment membrane fission from 1%in DspoIIIE to 13% (Bath et al. 2000; Sharp and Pogliano2003). We therefore predicted that expressing only thetransmembrane domain would at least partially rescuethe septal membrane fission defect of the null strain.FRAP analysis of spoIIIE1–192 sporangia produced a broadrange of cFR curves ranging from no recovery, similar towild-type cells, to full recovery, similar to DspoIIIE cells(Fig. 3A). Additional FRAP experiments demonstratedthat cells with intermediate recoveries continued torecover for at least 180 sec (data not shown). This variableoutcome (which is addressed below) indicates thatSpoIIIE1–192 only partially supports septal membrane
Figure 2. Septal fission defects occur only in DspoIIIE sporangia with trapped DNA. (A) Plot of cFR for DracA sporangia with (green) orwithout (black) DNA in the forespore show similarly limited fluorescence recovery. (Gray curve) Whole-cell bleach control. (B) Plot ofcFR for DspoIIIE DracA sporangia with DNA in the forespore (green) show recovery, and those without DNA (black) show limitedrecovery similar to wild type. (C) Cartoon showing possible membrane and DNA configurations for DspoIIIE DracA and the impact onFM 4-64 diffusion (arrows).
SpoIIIE mediates membrane fission
GENES & DEVELOPMENT 1163
Cold Spring Harbor Laboratory Press on June 13, 2010 - Published by genesdev.cshlp.orgDownloaded from
fission, and that the cytoplasmic domain is also involvedin septal membrane fission.We further investigated the role of the transmembrane
domain in septal membrane fission by performing FRAPexperiments on a previously isolated epitope insertion inthe SpoIIIE transmembrane domain (spoIIIE124Ti31, herereferred to as spoIIIEFis!) that partially blocks engulfmentmembrane fission and cytoplasmic compartmentalizationwithout preventing DNA translocation (Liu et al. 2006).FRAP analysis of spoIIIEFis! sporangia also showed a broadrange of cFR curves, from little or no recovery to completerecovery (Fig. 3B), providing further support for the impor-tance of the transmembrane domain in membrane fission.The effects of the spoIIIEFis! mutation on septal
membrane fission could occur either after the completion
of DNA translocation, as proposed previously by Liu et al.(2006), or during DNA translocation. To discriminatebetween these two hypotheses, we combined thespoIIIE(G467S) mutation that blocks DNA translocationto trap DNA across the septum with the spoIIIE124Ti31mutation to generate a spoIIIEFis!ATP! strain. PerformingFRAP on spoIIIEFis!ATP! sporangia produced the samewide range of cFR curves as in the single SpoIIIEFis!
mutant (Fig. 3C), indicating that SpoIIIEFis!ATP! fails tosupport membrane fission while DNA crosses the sep-tum. Thus, the spoIIIEFis! mutation affects the ability ofthe protein to mediate septal fission before or duringDNA translocation, rather than afterward.The observation that the SpoIIIE transmembrane
domain (SpoIIIE1–192) did not fully support membrane
Figure 3. Role of transmembrane and cytoplasmic domains in septal fission and SpoIIIE assembly. (A–D) Average cFR for wild type(blue) and DspoIIIE (green) are shown as references. Plot of cFR for individual cells (black lines) expressing SpoIIIE1–192 (A), SpoIIIEFis!
(B), and SpoIIIEFis!ATP! (C). (D) Average cFR and standard deviations for spoIIIE1–192 Fis! (orange). (E–J) Fluorescent images of membrane(red) and SpoIIIE-GFP (green) for wild type (E), SpoIIIEATP! (F), SpoIIIE1–192 (G), SpoIIIEFis! (H), SpoIIIEFis!ATP! (I), and SpoIIIE1–192 Fis! (J).The third panel is a three-dimensional plot of the normalized GFP intensity in each field. (K) Histogram of average GFP intensity forwild-type and mutant proteins, with 99% CI error bars. (L,M) Rank correlation plot of GFP focus intensity versus FRAP recovery inindividual cells expressing SpoIIIEFis!-GFP (L) and SpoIIIEFis!ATP! GFP (M), with the brightest focus and the lowest cFR rankedseparately as 1 (Supplemental Table S1). The black line of best fit and Spearman’s rank correlation (Rs values of 0.576 and 0.697, withP < 0.05 for SpoIIIEFis and SpoIIIEFis!ATP) show a correlation between dim foci and rapid recovery.
Fleming et al.
1164 GENES & DEVELOPMENT
Cold Spring Harbor Laboratory Press on June 13, 2010 - Published by genesdev.cshlp.orgDownloaded from

fission suggests that the cytoplasmic domain contributesto septal membrane fission. To further investigate therespective contributions of the cytoplasmic and trans-membrane domains to SpoIIIE assembly, we deletedthe cytoplasmic domain from SpoIIIEFis! to produceSpoIIIE1–192 Fis!. FRAP analysis of this strain demon-strated rapid recovery similar to the spoIIIE-null strain(Fig. 3D), with no sporangia showing an intermediaterecovery as in the full-length SpoIIIEFis! protein orSpoIIIE1–192. Thus, deletion of the cytoplasmic domainfrom a partially fission-defective mutant enhances themembrane fission defect. These results suggest that thetransmembrane domain and the cytoplasmic domaincooperate to mediate assembly of a SpoIIIE complex thatcan partition the forespore and mother cell membranes.
Correlation of mutant SpoIIIE protein assemblyand FRAP recovery
The wide range of FRAP recovery curves suggested thatindividual cells might vary in their ability to assemblea SpoIIIE DNA translocation complex that mediates septalmembrane fission. To test this hypothesis, we performedtwo experiments to determine if there was a correlationbetween the ability of various SpoIIIE mutant proteinsto assemble at the septum and their ability to completeseptal membrane fission. First, we inferred the state ofSpoIIIE assembly at the septal midpoint by quantifying thefluorescence intensity of wild-type and mutant SpoIIIE-GFP foci. The two membrane fission-proficient proteins,wild-type SpoIIIE-GFP and SpoIIIEATP!-GFP, showedequivalently high GFP focus intensities (Fig. 3E,F,K). Twomutations that cause intermediate septal fission defects,spoIIIE1–192 and spoIIIEFis!, caused a decrease in averagefluorescence intensities to 53% and 24% of wild-typelevels, respectively (Fig. 3G–I,K). The mutant completelydefective for membrane fission, SpoIIIE1–192 Fis-GFP, as-sembled few detectable foci, which were very dim (Fig.3J,K). These results suggest a correlation between decreas-ing GFP focus intensity and a decreased ability to mediatemembrane fission, which we verified does not result fromchanges in proteins levels (Supplemental Fig. S3). Second,we assessed the correlation between GFP focus intensityand FRAP outcomes in individual cells of the spoIIIEFis!
and spoIIIEFis!ATP! strains, which show a broad range ofFRAP recovery curves and focus intensities. A Spearman’srank correlation test indicated a significant correlationbetween the assembly of a bright SpoIIIE focus at theseptum and membrane compartmentalization (Fig. 3L,M)in which cells with dimmer GFP foci tended to have fasterFRAP recovery rates (Supplemental Table S1). The corre-lation between GFP focus intensity and FRAP recoverysuggests a direct involvement of SpoIIIE in septal mem-brane fission during sporulation.
Multistep pathway for SpoIIIE assembly
To further explore the connection between SpoIIIE as-sembly and membrane fission, we used fluorescence mi-croscopy to follow SpoIIIE assembly. We first performed
time-lapse epifluorescence microscopy over a compara-tively long time scale of at least 30 min. This revealeda dynamic population of SpoIIIE molecules at the divisionsite both before and during constriction (Fig. 4A; Supple-mental Fig. S4). The SpoIIIE focus was first observed ;3min after the apparent completion of septation (Fig. 4A),although this estimate is imprecise due to the limitedspatial resolution of epifluorescence microscopy, whichlimits the ability to visualize small openings in theseptum. Nevertheless, this suggests that the SpoIIIE focusrapidly assembles from a dynamic population of mole-cules at the septum.We next used total internal reflection fluorescence
(TIRF) microscopy to explore SpoIIIE dynamics withincreased temporal resolution. The increased sensitivityof TIRF allows visualization of fewer molecules, anda shorter exposure time allows visualization of rapidlymoving molecules. To compensate for the shallow illu-mination plane, we used partial TIRF in which the laserwas aligned so that stable SpoIIIE foci at the septalmidpoint (;400 nm into the cell) were clearly imaged.TIRF imaging demonstrated that SpoIIIE initially local-izes to incomplete septa as dynamic foci (Fig. 4B,D).Septal kymographs, in which slices across the septum ateach time point are cropped and stacked, more clearlydemonstrated movement of SpoIIIE, and suggested thatfoci either assembled and disassembled on opposite sidesof the septum, or moved from one side to the otherwithin seconds (Fig. 4B,D). Thus, the first step of SpoIIIEassembly is dynamic localization to active division sites,which is challenging to visualize with epifluorescencemicroscopy but is clearly imaged by TIRF. Cells with flatsepta showed two distinct patterns. Most (85%) showeda stable focus that moved little throughout the experi-ment (Fig. 4C,E). However, ;15% showed SpoIIIE-GFPthat moved within the septum (Fig. 4E), indicating thateither septation is not fully complete in these cells, orthe SpoIIIE focus has not yet fully assembled, as con-firmed by live-cell photoactivation localization micros-copy (PALM) (Fig. 5).
The transmembrane domain assemblesan unstable focus
We next examined the dynamics of the SpoIIIE transmem-brane domain by TIRF microscopy. SpoIIIE1–192-GFP mol-ecules showed dynamic localization to incomplete septathat was indistinguishable from the wild type (Fig. 4F),indicating that the transmembrane domain is sufficient fordynamic septal localization. Cells in which septation ap-peared to be complete showed dynamic assemblies and fociin frequencies similar to wild type. However, septal kymo-graphs revealed that SpoIIIE1–192-GFP foci were less stablethan the wild-type foci. Specifically, SpoIIIE1–192-GFP focioften showed frequent flares emanating out from thecentral focus (Fig. 4G). Indeed, even the most stable trans-membrane domain focus observed (Fig. 4G, cell 6) showedan occasional diffuse focus not seen in wild type. Thissuggests that the SpoIIIE transmembrane domain assem-bles unstable foci.
SpoIIIE mediates membrane fission
GENES & DEVELOPMENT 1165
Cold Spring Harbor Laboratory Press on June 13, 2010 - Published by genesdev.cshlp.orgDownloaded from

SpoIIIE localizes to the leading edgeof constricting septa
To improve the spatial resolution of SpoIIIE imagingexperiments, we next performed PALM on cells express-ing SpoIIIE, SpoIIIEATP!, and SpoIIIE1–192 fused to tandemdimer EOS (tdEOS) (Wiedenmann et al. 2004). PALMallows localization of individual SpoIIIE-tdEOS moleculeswith 20-nm precision and the visualization of SpoIIIElocalization at higher, non-diffraction-limited spatial res-olution (Betzig et al. 2006; Shroff et al. 2008; McKinneyet al. 2009). We performed PALM on live cells to avoidmembrane perturbations and other artifacts caused bychemical fixation (Supplemental Fig. S5). Live-cell PALMdemonstrated that SpoIIIE is clearly localized to constrict-ing and flat septa compared with the rest of themembrane(Fig. 5A–C,L). This high-resolution imaging clearly shows
SpoIIIE molecules to be confined near the leading edge ofconstricting septa, forming clusters on one or both sides ofincomplete septa (Fig. 5, class I), a phenomenon that ismuch less apparent in diffraction-limited epifluorescenceimages (Fig. 5, inset). Flat septa displayed two classes oflocalization. In 52% of flat septa, two distinct or oneelongated cluster of SpoIIIE-tdEOS was distributed alongthe septal membrane (Fig. 5B, class II). These are likelyincomplete septa with an opening smaller than the reso-lution of the conventional light microscope, since theyoccur at a frequency similar to septal retraction in wild-type cells in the protoplast assay (Fig. 1B). Another 40%show a single cluster of SpoIIIE molecules at the septalmidpoint, while 8% display too few molecules to beclassified (Fig. 5C, class III). These septa have likelycompleted septation and assembled a SpoIIIE DNA trans-location complex. Thus, live-cell PALM demonstrates
Figure 4. SpoIIIE dynamics. (A) Time-lapse epifluorescence microscopy of SpoIIIE-GFP. Images of FM 4-64 staining (top) and GFPfluorescence (bottom) were collected every 90 sec with time (minutes) shown below the images. The asterick denotes the apparentcompletion of septation, based on membrane images (see also Supplemental Movie S5). (B,C) Diagrams show construction of septalkymographs in cells with unstable (B) or stable (C) foci. Slices the width of septum were taken for each time point and then stacked,with the first time point at the top and the last at the bottom. (D–G) TIRF microscopy of SpoIIIE-GFP (D,E) and SpoIIIE1–192-GFP (F,G)sporangia with clearly incomplete septa (D,F) or flat septa (E,G), based on FM 4-64 staining. Images were collected approximately every0.5 sec for >37 sec. Septal kymographs for nine individual cells, including three incomplete and six flat septa, were generated bystacking 1 3 18 pixel slices of the septum for each time. Arrowheads indicate the times on the kymographs that correspond to the fullcell images shown to the left of the kymographs. The first image shows the membrane, and the three following images show TIRF GFPfluorescence at the indicated times (see also Supplemental Movies S6, S7).
Fleming et al.
1166 GENES & DEVELOPMENT
Cold Spring Harbor Laboratory Press on June 13, 2010 - Published by genesdev.cshlp.orgDownloaded from
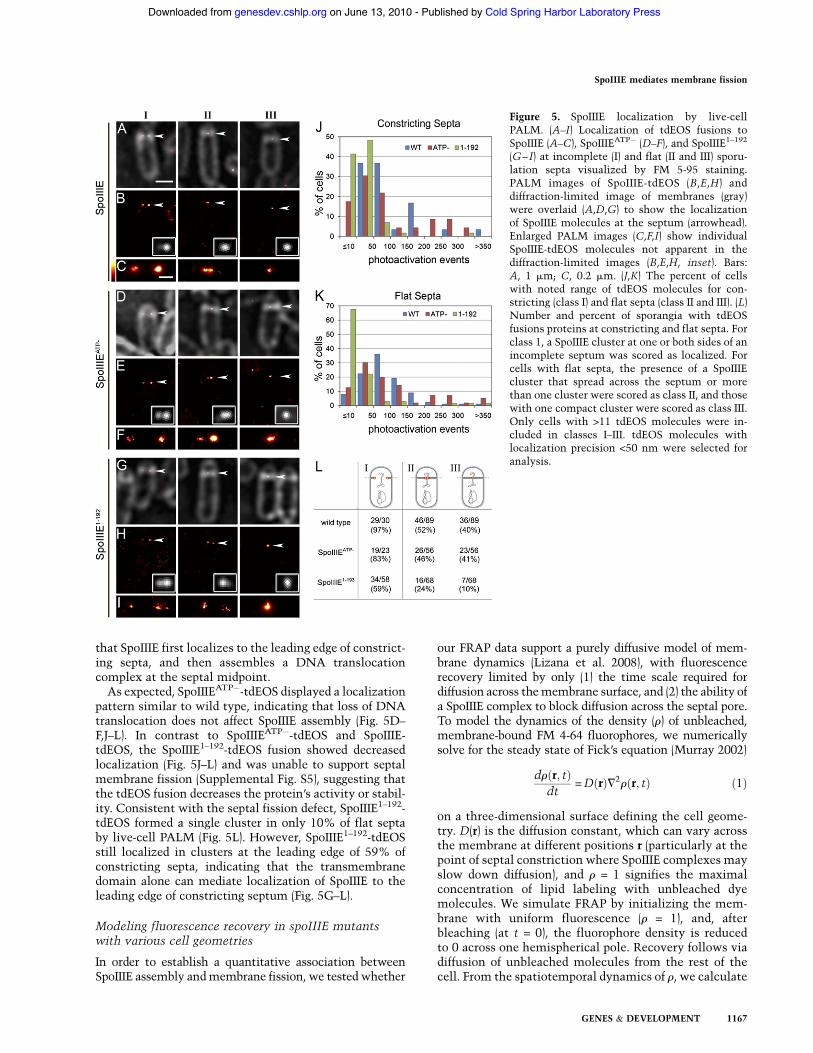
that SpoIIIE first localizes to the leading edge of constrict-ing septa, and then assembles a DNA translocationcomplex at the septal midpoint.As expected, SpoIIIEATP!-tdEOS displayed a localization
pattern similar to wild type, indicating that loss of DNAtranslocation does not affect SpoIIIE assembly (Fig. 5D–F,J–L). In contrast to SpoIIIEATP!-tdEOS and SpoIIIE-tdEOS, the SpoIIIE1–192-tdEOS fusion showed decreasedlocalization (Fig. 5J–L) and was unable to support septalmembrane fission (Supplemental Fig. S5), suggesting thatthe tdEOS fusion decreases the protein’s activity or stabil-ity. Consistent with the septal fission defect, SpoIIIE1–192-tdEOS formed a single cluster in only 10% of flat septaby live-cell PALM (Fig. 5L). However, SpoIIIE1–192-tdEOSstill localized in clusters at the leading edge of 59% ofconstricting septa, indicating that the transmembranedomain alone can mediate localization of SpoIIIE to theleading edge of constricting septum (Fig. 5G–L).
Modeling fluorescence recovery in spoIIIE mutantswith various cell geometries
In order to establish a quantitative association betweenSpoIIIE assembly andmembrane fission, we tested whether
our FRAP data support a purely diffusive model of mem-brane dynamics (Lizana et al. 2008), with fluorescencerecovery limited by only (1) the time scale required fordiffusion across themembrane surface, and (2) the ability ofa SpoIIIE complex to block diffusion across the septal pore.To model the dynamics of the density (r) of unbleached,membrane-bound FM 4-64 fluorophores, we numericallysolve for the steady state of Fick’s equation (Murray 2002)
dr"r; t#dt
=D"r#=2r"r; t# "1#
on a three-dimensional surface defining the cell geome-try. D(r) is the diffusion constant, which can vary acrossthe membrane at different positions r (particularly at thepoint of septal constriction where SpoIIIE complexes mayslow down diffusion), and r = 1 signifies the maximalconcentration of lipid labeling with unbleached dyemolecules. We simulate FRAP by initializing the mem-brane with uniform fluorescence (r = 1), and, afterbleaching (at t = 0), the fluorophore density is reducedto 0 across one hemispherical pole. Recovery follows viadiffusion of unbleached molecules from the rest of thecell. From the spatiotemporal dynamics of r, we calculate
Figure 5. SpoIIIE localization by live-cellPALM. (A–I) Localization of tdEOS fusions toSpoIIIE (A–C), SpoIIIEATP! (D–F), and SpoIIIE1–192
(G–I) at incomplete (I) and flat (II and III) sporu-lation septa visualized by FM 5-95 staining.PALM images of SpoIIIE-tdEOS (B,E,H) anddiffraction-limited image of membranes (gray)were overlaid (A,D,G) to show the localizationof SpoIIIE molecules at the septum (arrowhead).Enlarged PALM images (C,F,I) show individualSpoIIIE-tdEOS molecules not apparent in thediffraction-limited images (B,E,H, inset). Bars:A, 1 mm; C, 0.2 mm. (J,K) The percent of cellswith noted range of tdEOS molecules for con-stricting (class I) and flat septa (class II and III). (L)Number and percent of sporangia with tdEOSfusions proteins at constricting and flat septa. Forclass 1, a SpoIIIE cluster at one or both sides of anincomplete septum was scored as localized. Forcells with flat septa, the presence of a SpoIIIEcluster that spread across the septum or morethan one cluster were scored as class II, and thosewith one compact cluster were scored as class III.Only cells with >11 tdEOS molecules were in-cluded in classes I–III. tdEOS molecules withlocalization precision <50 nm were selected foranalysis.
SpoIIIE mediates membrane fission
GENES & DEVELOPMENT 1167
Cold Spring Harbor Laboratory Press on June 13, 2010 - Published by genesdev.cshlp.orgDownloaded from
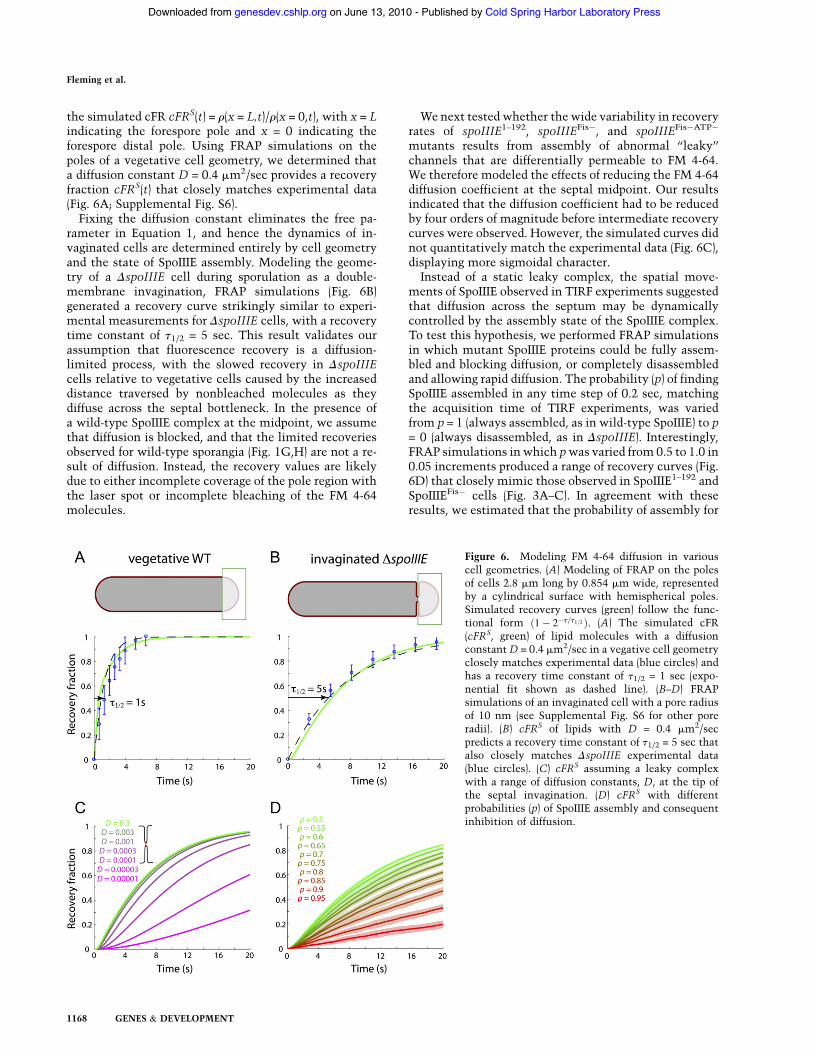
the simulated cFR cFRS(t) = r(x = L,t)/r(x = 0,t), with x = Lindicating the forespore pole and x = 0 indicating theforespore distal pole. Using FRAP simulations on thepoles of a vegetative cell geometry, we determined thata diffusion constant D = 0.4 mm2/sec provides a recoveryfraction cFRS(t) that closely matches experimental data(Fig. 6A; Supplemental Fig. S6).Fixing the diffusion constant eliminates the free pa-
rameter in Equation 1, and hence the dynamics of in-vaginated cells are determined entirely by cell geometryand the state of SpoIIIE assembly. Modeling the geome-try of a DspoIIIE cell during sporulation as a double-membrane invagination, FRAP simulations (Fig. 6B)generated a recovery curve strikingly similar to experi-mental measurements for DspoIIIE cells, with a recoverytime constant of t1/2 = 5 sec. This result validates ourassumption that fluorescence recovery is a diffusion-limited process, with the slowed recovery in DspoIIIEcells relative to vegetative cells caused by the increaseddistance traversed by nonbleached molecules as theydiffuse across the septal bottleneck. In the presence ofa wild-type SpoIIIE complex at the midpoint, we assumethat diffusion is blocked, and that the limited recoveriesobserved for wild-type sporangia (Fig. 1G,H) are not a re-sult of diffusion. Instead, the recovery values are likelydue to either incomplete coverage of the pole region withthe laser spot or incomplete bleaching of the FM 4-64molecules.
We next tested whether the wide variability in recoveryrates of spoIIIE1–192, spoIIIEFis!, and spoIIIEFis!ATP!
mutants results from assembly of abnormal ‘‘leaky’’channels that are differentially permeable to FM 4-64.We therefore modeled the effects of reducing the FM 4-64diffusion coefficient at the septal midpoint. Our resultsindicated that the diffusion coefficient had to be reducedby four orders of magnitude before intermediate recoverycurves were observed. However, the simulated curves didnot quantitatively match the experimental data (Fig. 6C),displaying more sigmoidal character.Instead of a static leaky complex, the spatial move-
ments of SpoIIIE observed in TIRF experiments suggestedthat diffusion across the septum may be dynamicallycontrolled by the assembly state of the SpoIIIE complex.To test this hypothesis, we performed FRAP simulationsin which mutant SpoIIIE proteins could be fully assem-bled and blocking diffusion, or completely disassembledand allowing rapid diffusion. The probability (p) of findingSpoIIIE assembled in any time step of 0.2 sec, matchingthe acquisition time of TIRF experiments, was variedfrom p = 1 (always assembled, as in wild-type SpoIIIE) to p= 0 (always disassembled, as in DspoIIIE). Interestingly,FRAP simulations in which pwas varied from 0.5 to 1.0 in0.05 increments produced a range of recovery curves (Fig.6D) that closely mimic those observed in SpoIIIE1–192 andSpoIIIEFis! cells (Fig. 3A–C). In agreement with theseresults, we estimated that the probability of assembly for
Figure 6. Modeling FM 4-64 diffusion in variouscell geometries. (A) Modeling of FRAP on the polesof cells 2.8 mm long by 0.854 mm wide, representedby a cylindrical surface with hemispherical poles.Simulated recovery curves (green) follow the func-tional form "1! 2!t=t1=2 #. (A) The simulated cFR(cFRS, green) of lipid molecules with a diffusionconstantD = 0.4 mm2/sec in a vegative cell geometryclosely matches experimental data (blue circles) andhas a recovery time constant of t1/2 = 1 sec (expo-nential fit shown as dashed line). (B–D) FRAPsimulations of an invaginated cell with a pore radiusof 10 nm (see Supplemental Fig. S6 for other poreradii). (B) cFRS of lipids with D = 0.4 mm2/secpredicts a recovery time constant of t1/2 = 5 sec thatalso closely matches DspoIIIE experimental data(blue circles). (C) cFRS assuming a leaky complexwith a range of diffusion constants, D, at the tip ofthe septal invagination. (D) cFRS with differentprobabilities (p) of SpoIIIE assembly and consequentinhibition of diffusion.
Fleming et al.
1168 GENES & DEVELOPMENT
Cold Spring Harbor Laboratory Press on June 13, 2010 - Published by genesdev.cshlp.orgDownloaded from

the cells represented by the kymographs in Figure 4G is0.22, 0.55, 0.78, 0.88, 0.96, and 0.98 (for cells 1–6, re-spectively). Taken together, these simulations supporta model in which the intermediate FRAP curves observedin spoIIIE1–192, spoIIIEFis!, and spoIIIEFis!ATP! strains re-flect repeated disassembly and assembly of a SpoIIIEchannel at the septal midpoint, rather than the staticassembly of a channel that is only partially permeable tolipid molecules.
Discussion
Our data indicate that SpoIIIE assembles a DNA trans-location complex via a multistep pathway that requiresboth the transmembrane domain, which mediates dy-namic localization to division sites and the assembly ofunstable foci, and the cytoplasmic domain, which stabi-lizes the DNA translocation complex. We further dem-onstrated that the absence of SpoIIIE causes a severedefect in septal membrane fission when DNA traversesthe septum, that daughter cell membranes are separatedduringDNA translocation, and that there is a quantitativeassociation between septal membrane fission and theassembly of a stable DNA translocation complex. Thesefindings provide the first direct experimental evidencethat SpoIIIE assembly mediates septal membrane fissionduring sporulation (Liu et al. 2006; Burton et al. 2007).The first step in the SpoIIIE assembly pathway is
dynamic localization to active division sites, a process me-diated by the transmembrane domain (Fig. 7A–C, step 1).Epifluorescencemicroscopy initially suggested that SpoIIIElocalizes as a ring at constricting septa (Sharp and Pogliano2002). However, the improved temporal resolution of TIRFmicroscopy reveals that these ‘‘rings’’ represent the blurredmotion of rapidly movingmolecules imaged with exposuretimes that are long relative to the rate of movement. Theimproved spatial resolution of live-cell PALM indicatesthat SpoIIIE localizes specifically to the leading edge ofconstricting septa. It is tempting to speculate that thislocalization might be mediated by the ability of the trans-membrane domain to sense regions of increasedmembranecurvature, as demonstrated for several membrane proteinsand wedge-shaped lipids (Huang et al. 2006; Ramamurthiet al. 2009).The second step in SpoIIIE assembly is the formation of
a focus at the septal midpoint, a process mediated by thetransmembrane domain. Live-cell PALM demonstratesthat there are similar numbers of SpoIIIE moleculesmoving at constricting septa as in the focus, suggestingthat most SpoIIIE molecules at the nascent septumultimately assemble into the septal focus. In keeping withprevious studies (Bath et al. 2000; Sharp and Pogliano2003; Liu et al. 2006), we find that the transmembranedomain is capable of assembling foci at the septal mid-point, although TIRFmicroscopy demonstrates that thesefoci are less stable than those assembled by the full-lengthprotein. Our data therefore suggest a third step in SpoIIIEassembly: stabilization of the transmembrane assemblyby the cytoplasmic domain. We suggest that this stepinvolves the engagement of the DNA motor domain with
DNA that traverses the septum, which stabilizes theSpoIIIE complex for the duration of DNA translocation(Fig. 7A–C, steps 3, 4). This stabilization might alsorequire the linker domain, which appears to stabilize themotor domain of Escherichia coli FtsK (Dubarry and Barre2010). This DNA-dependent stabilization of the SpoIIIEfocus likely explains previous observations that SpoIIIEassembles only in the presence of trapped DNA (Ben-Yehuda et al. 2003a), since transient foci and dynamicSpoIIIE molecules at nascent septa are difficult to image.
Figure 7. Model for SpoIIIE function in septal membranefission. Diagrams of septation show the whole cell (A), withthe region of interest indicated by red circles, and the septum asa side view (B) and as a top view (C), showing the lipid bilayer(green circles), DNA (gold), and SpoIIIE (blue). SpoIIIE trans-membrane domain is shown as a wedge in the lipid bilayer, withthe linker as a black line and the motor domain as a sphere.(Step 1) Initially, SpoIIIE dynamically localizes to the leadingedge of constricting septa, but does not block lipid diffusion. Asthe membrane contacts DNA, the SpoIIIE transmembranedomain assembles around the trapped DNA (step 2) formingan unstable channel that can separate membranes (step 3). (Step4) The cytoplasmic motor domain then assembles on the DNA(likely only in the mother cell) (Becker and Pogliano 2007),stabilizing the channel and forming a channel that blocksdiffusion between the daughter cell membranes during DNAtranslocation. We show this structure to be a paired channel,although other arrangements are possible. (Step 5) After DNAtranslocation, the motor domain releases DNA, and the com-plex disassociates to allow the terminus across the septum.(Step 6) Finally, irreversible membrane fission occurs.
SpoIIIE mediates membrane fission
GENES & DEVELOPMENT 1169
Cold Spring Harbor Laboratory Press on June 13, 2010 - Published by genesdev.cshlp.orgDownloaded from

SpoIIIE therefore has at least two domains that co-operate to mediate the assembly of the DNA transloca-tion complex: the transmembrane domain that targetsthe protein to the septum (and likely assembles a channelat the septal midpoint), and the DNA motor domain thatcan hexamerize in the presence of DNA even in theabsence of the transmembrane domain (Ptacin et al.2008). A critical question for future studies is how theassembly and disassembly of these domains are coordi-nated and coupled to the initiation and termination ofDNA translocation.Our membrane FRAP experiments and biophysical
modeling results indicate that FM 4-64 diffuses throughthe cytoplasmic membrane with a diffusion coefficient of0.4 mm2/sec, a rate that suggests that the dye moleculesdo not significantly impair free diffusion within thecytoplasmicmembrane due to interactions with localizedproteins or lipids. They also demonstrate that, in theabsence of SpoIIIE, septal membrane fission does notoccur, allowing FM 4-64 to diffuse into the bleachedforespore membrane at rates consistent with the exis-tence of a 10-nm opening in the septum. These resultscontradict a previous study in which fewer cells werebleached and fewer spoIIIE mutants were examined (seeSupplemental Fig. S1 for discussion; Burton et al. 2007).However, consistent with this prior study, we find thattranslocation-defective and assembly-proficient SpoIIIEproteins are capable of blocking lipid diffusion throughthe septum. This suggests that the DNA translocationcomplex moves the DNA across separated daughter cellmembranes. This finding is consistent with the pairedchannel model for SpoIIIE topology (Liu et al. 2006;Burton et al. 2007), which postulates that SpoIIIE formsa channel that spans two lipid bilayers (Fig. 7). Recentexperiments indicate that the transmembrane domain ofthe SpoIIIE homolog FtsK can be replaced with othertransmembrane domains (Dubarry and Barre 2010). Thissuggests that DNA translocation does not depend on theassembly of a channel, which is consistent with ourobservations that the membrane fission-defective SpoIIIEmutant is DNA translocation-proficient (Liu et al. 2006).Our localization studies, FRAP analysis, and biophysi-
cal modeling efforts provide quantitative support for themodel that assembly of SpoIIIE into a focus at the septalmidpoint mediates septal membrane fission (Liu et al.2006). Most compelling of these findings is that theSpoIIIE transmembrane domain, which TIRF microscopyshows to assemble unstable foci, produces a wide range ofFRAP recovery curves. Biophysical modeling of FRAPdata indicates that this outcome can be readily recapitu-lated in simulations in which the SpoIIIE transmembranedomain alternates between fully assembled and disas-sembled states that either completely block or allow FM4-64 diffusion across the septum, respectively (Fig. 7A–C,steps 2, 3). This suggests that the variable FRAP outcomesin different cells reflect subtle changes in the probabilityof assembly of the transmembrane domain in each cell,perhaps due to cell–cell variations in the number ofSpoIIIE molecules at the septum observed by PALM (Fig.5). If SpoIIIE does assemble a channel for the DNA, then
the assembly and disassembly we observe would requirethat the hydrophilic amino acids that line the aqueouspore be masked in the disassembled subunits to preventthe energetically unfavorable interaction of these aminoacids with the hydrophobic core of the lipid bilayer. Thislikely requires either the presence of membrane proteinsthat act as chaperones for the unfolded subunits, orrearrangements of the individual transmembrane seg-ments to bury these regions.Several genetic, biophysical, and cell biological ex-
periments indicate that SpoIIIE translocates the twoarms of the circular chromosome simultaneously (Wuand Errington 1998; Becker and Pogliano 2007; Burtonet al. 2007; Ptacin et al. 2008), although structural stud-ies demonstrate that the DNA motor domain can onlyaccommodate a single dsDNA strand (Massey et al. 2006).It is therefore likely that the septum contains at least twoSpoIIIE channels, one for each arm of the circular chro-mosome that traverses the septum. Translocating a circu-lar chromosome through two adjacent channels posesa difficulty when the final loop of DNA reaches theseptum. Our observation that the absence of the DNA-binding domain produces a channel that can assembleand disassemble suggests that, once the terminus reachesthe septum, the DNA motor domain disassociates fromthe DNA and destabilizes the channel, opening a porethat allows the terminus to pass into the forespore (Fig.7A–C, step 5). After DNA is cleared from the septum,SpoIIIE or another protein involved in septal membranefission in the absence of DNA could permanently sepa-rate the membranes (Fig. 7A–C, step 6).Finally, our data emphasize the importance of thought-
fully considering the caveats of various imaging methodsbefore interpreting cell biological data. First, althoughboth epifluorescence microscopy and live-cell PALMappear to show that SpoIIIE localizes as rings at nascentdivision sites, TIRF microscopy shows that these ringsare artifacts caused by blurring of dynamic moleculesduring long exposure times in epifluorescence micros-copy and the compilation of molecules imaged overseveral minutes into a single image in PALM. Second,although the dynamic assemblies at constricting septaappear less intense than foci in both epifluorescence andTIRF microscopy, live-cell PALM demonstrates thatthere are similar numbers of SpoIIIE molecules at bothassembly stages. Live-cell PALM has two attributes thatlikely account for this difference: the ability to detect andenumerate single fluorescent molecules, which elimi-nates the preferential detection of stationary rather thandynamicmolecules, and themore effective elimination ofnoise during imaging processing that is facilitated by theintense burst of photons emitted by tdEOS during theshort (50-msec) exposure time. Thus, PALM providesboth improved spatial resolution and a more quantitativemeasure of SpoIIIE abundance, TIRF probes SpoIIIE dy-namics at higher temporal resolution to visualize rapidsteps in complex assembly, and epifluorescence providesa broad perspective of SpoIIIE dynamics throughout celldivision and engulfment. Each method reveals differ-ent features of the SpoIIIE assembly pathway, which is
Fleming et al.
1170 GENES & DEVELOPMENT
Cold Spring Harbor Laboratory Press on June 13, 2010 - Published by genesdev.cshlp.orgDownloaded from

characterized by a wide range of movement rates, fromthe very rapid movement around nascent septa (withinseconds) to the more leisurely movement of the focus tothe cell pole during engulfment (within 45 min). Integra-tion of these approaches with biophysical modeling pro-vides amechanistic explanation of membrane fission thatbridges the molecular and cellular scales.
Materials and methods
Strains and culture conditions
Strains are derivatives of B. subtilis PY79 (Supplemental TableS2). Mutations were introduced by transformation with plasmidsor genomic DNA (Dubnau and Davidoff-Abelson 1971). See theSupplemental Material for details of plasmid construction, in-cluding oligonucleotide sequences (Supplemental Table S3).Cells were grown in 20%–25% LB, and sporulation was inducedby resuspension (Becker and Pogliano 2007) at 37°C unless notedotherwise.
Microscopy
Except for PALM data, microscopy was performed on an AppliedPrecision optical sectioning microscope (Liu et al. 2006) usingsoftWoRx version 3.3.6 (Applied Precision) and ImageJ foranalysis. Cells were sporulated in the presence of 0.5 mg/mLmembrane dye FM 4-64 and 0.5 mM nucleic acid stain SytoxGreen (Invitrogen), as needed. Samples were immobilized onpoly-L-lysine-treated coverslips, except for TIRF and time-lapsemicroscopy, which were performed on agarose pads (Becker andPogliano 2007). For time lapse, cells were placed on the agarosepad 1.5–2 h after resuspension and were incubated at 30°C duringimaging. Images were collected every 90 sec for >30 min. TIRFmicroscopy used an Olympus 1003 1.65 Apo objective, immer-sion oil n = 1.78 (Cargille Laboratories), and sapphire coverslips(Olympus). TIRF images were collected approximately every 500msec, until the sample photobleached using a 488-nm argonlaser attached to an Applied Precision TIRF module set at 70%power for pulses of 0.2 sec. Kymographs were generated withImageJ, making a montage of 0.06-mm 3 1.14-mm slices throughthe septum at each time.
Protoplast assay
Two hours to 2.5 h after initiation of sporulation, cells wereconcentrated 203 in SMM buffer, placed on a slide, and mixedwith lysozyme (final concentration, 1 mg/mL) (Broder andPogliano 2006). Images were taken every 60 sec until all cellsformed protoplasts, which was variable. Sporangia were scoredas having either complete septal fission (if the septum remainedthroughout protoplast formation to generate two protoplasts) orincomplete septal fission (by septal retraction).
Quantification of FRAP
To maintain sufficient fluorescence to measure recovery afterphotobleaching, we sporulated cells in 2 mg/mL FM 4-64. After2–2.5 h of sporulation at 37°C or 3 h of sporulation at 30°C, cellswere pelleted by microcentrifugation at 7000 rpm for 10 sec,washed twice in sporulation media, resuspended in 1/20th vol offresh sporulation media, and mounted on a poly-Lysine cover-slip. Photobleaching was achieved with one 0.3-sec pulse froma 488-nm argon laser set to 20%–30% power. FRAP quantifica-tion was performed as described by Broder and Pogliano (2006)
with the following modifications. To compare multiple FRAPevents on a single graph, we calculated the cFR by determiningthe relative intensity of the bleached region to the unbleachedregion, and defining the bleach event as cFR = 0 and completerecovery as cFR = 1 for each photobleached sporangia (Supple-mental Material; Wu et al. 2006). Average cFR curves wereobtained by averaging fluorescence recovery values at the sametime points for each strain. The equation cFR(t) = A(1 ! e!t/t1)(y0+ Be!t/t2) was used for curve fitting.
Quantification of GFP fluorescence intensity
To avoid bleed-through of FM 4-64 fluorescence in the FITCchannel, we sporulated cells in 0.25 mg/mL FM4-64. Images ofthe medial focal plane were taken 2 h after resuspension with a3-sec exposure for GFP. GFP intensities were first modified by3 3 3 binning using ImageJ to smooth fluorescence intensitywhen the peak is spread over 2 pixels versus centered over1 pixel. GFP foci were identified using the ImageJ threshold tool.Only foci from sporangia with flat or slightly curved septa werequantified. The maximum fluorescence values for at least 200foci for each strain were corrected for background fluorescence,normalized to the photosensor reading for each image to accountfor any changes in bulb intensity, and then averaged.
PALM acquisition and analysis
The PALM instrument was built on a commercial Olympus IX71fluorescence microscope with two lasers, wavelengths 561 nm(Compass 561, 20 mW, Coherent, Inc.) and 405 nm (Cube, 405nm, 100 mW, Coherent, Inc.), and an Olympus UAPO150xO/TIRFM-SP NA1.45 objective. Fluorescence images were acquiredby a low-noise, light-sensitive EMCCD (DV887ECS-BV, Andor).Cells were sporulated by resuspension at 30°C, and the mem-branes were stained with 0.05 mg/mL FM 5-95 and harvested at3 h after sporulation. After two washes and then concentra-tion, cells mixed with gold nanospheres (790122, Microspheres-Nanospheres) were immobilized on poly-L-lysine-treated cov-erslips. FM 5-95-labeled membranes were imaged with thefluorescence filter set (FF01-560/25-25 and FF01-684/25-25, Sem-rock), and then photobleached by illumination with intense561-nm laser (;4.2 mW) for a few minutes prior to the PALMdata acquisition to eliminate background fluorescence. Afterfocusing to achieve partial TIRF, the 405-nm activation laserwas turned on low (1.2–6.0 mW) and PALM data were acquiredusing an emission filter for tdEOS (FF01-588/21-25, Semrock).About 10,000 frames at 50 msec per frame were acquired andanalyzed with programs written inMATLAB based on previouslydescribed algorithms (Betzig et al. 2006). Themolecular positionsof local peaks (approximately eight times brighter than thebackground noise) were determined by fitting to two-dimensionalGaussian PSF, and in-plane sample drift was corrected by the lo-calization of the bright Au-fiducials at nanometer accuracy (Yildizet al. 2003). See the Supplemental Material for further details.
Acknowledgments
We thank Jan Liphardt, Ann McEvoy, and Derek Greenfield fordiscussion and assistance with the PALM instrumentation, andMichael W. Davidson for the gift of the tdEOS plasmid. We alsothank Dan Broder for help with the protoplast experiments. T.F.and E.B. performed the epifluorescence, FRAP, and TIRF exper-iments in the K.P. laboratory. K.C.H. performed the biophysicalmodeling experiments. J.Y.S and S.H.L. conducted PALM exper-iments in the C.B. laboratory. This research was supportedby National Institutes of Health grants GM057045 (to K.P.),
SpoIIIE mediates membrane fission
GENES & DEVELOPMENT 1171
Cold Spring Harbor Laboratory Press on June 13, 2010 - Published by genesdev.cshlp.orgDownloaded from

GM071552 and GM032543 (to C.B.), and GM075000 (to K.C.H),and the Director’s New Innovator Award DP2OD006466 (toK.C.H). T.C.F. is supported by the NIH post-doctoral fellowship5F32GM081174.
References
Bath J, Wu LJ, Errington J, Wang JC. 2000. Role of Bacillussubtilis SpoIIIE in DNA transport across the mother cell-prespore division septum. Science 290: 995–997.
Becker EC, Pogliano K. 2007. Cell-specific SpoIIIE assembly andDNA translocation polarity are dictated by chromosomeorientation. Mol Microbiol 66: 1066–1079.
Ben-Yehuda S, Rudner DZ, Losick R. 2003a. Assembly of theSpoIIIE DNA translocase depends on chromosome trappingin Bacillus subtilis. Curr Biol 13: 2196–2200.
Ben-Yehuda S, Rudner DZ, Losick R. 2003b. RacA, a bacterialprotein that anchors chromosomes to the cell poles. Science299: 532–536.
Betzig E, Patterson GH, Sougrat R, Lindwasser OW, Olenych S,Bonifacino JS, Davidson MW, Lippincott-Schwartz J, HessHF. 2006. Imaging intracellular fluorescent proteins at nano-meter resolution. Science 313: 1642–1645.
Broder DH, Pogliano K. 2006. Forespore engulfment mediated bya ratchet-like mechanism. Cell 126: 917–928.
Burton BM, Marquis KA, Sullivan NL, Rapoport TA, RudnerDZ. 2007. The ATPase SpoIIIE transports DNA across fusedseptal membranes during sporulation in Bacillus subtilis.Cell 131: 1301–1312.
Dubarry N, Barre FX. 2010. Fully efficient chromosome dimerresolution in Escherichia coli cells lacking the integralmembrane domain of FtsK. EMBO J 29: 597–605.
Dubnau D, Davidoff-Abelson R. 1971. Fate of transformingDNA following uptake by competent Bacillus subtilis.I. Formation and properties of the donor–recipient complex.J Mol Biol 56: 209–221.
Errington J. 2001. Septation and chromosome segregation duringsporulation in Bacillus subtilis. Curr Opin Microbiol 4: 660–666.
Errington J. 2003. Regulation of endospore formation in Bacillussubtilis. Nat Rev Microbiol 1: 117–126.
Finger FP, White JG. 2002. Fusion and fission: Membranetrafficking in animal cytokinesis. Cell 108: 727–730.
Hilbert DW, Piggot PJ. 2004. Compartmentalization of geneexpression during Bacillus subtilis spore formation. Micro-biol Mol Biol Rev 68: 234–262.
Huang KC, Mukhopadhyay R, Wingreen NS. 2006. A curvature-mediated mechanism for localization of lipids to bacterialpoles. PLoS Comput Biol 2: e151. doi: 10.1371/journal.pcbi.0020151.
Jurgens G. 2005. Plant cytokinesis: Fission by fusion. TrendsCell Biol 15: 277–283.
Liu NJ, Dutton RJ, Pogliano K. 2006. Evidence that the SpoIIIEDNA translocase participates in membrane fusion duringcytokinesis and engulfment. Mol Microbiol 59: 1097–1113.
Lizana L, Bauer B, Orwar O. 2008. Controlling the rates ofbiochemical reactions and signaling networks by shape andvolume changes. Proc Natl Acad Sci 105: 4099–4104.
Massey TH, Mercogliano CP, Yates J, Sherratt DJ, Lowe J. 2006.Double-stranded DNA translocation: Structure and mecha-nism of hexameric FtsK. Mol Cell 23: 457–469.
McKinney SA, Murphy CS, Hazelwood KL, Davidson MW,Looger LL. 2009. A bright and photostable photoconvertiblefluorescent protein. Nat Methods 6: 131–133.
Murray JD. 2002. Mathematical biology. Springer, New York.
Pogliano K, Hofmeister AE, Losick R. 1997. Disappearanceof the sE transcription factor from the forespore and theSpoIIE phosphatase from the mother cell contributes toestablishment of cell-specific gene expression during sporu-lation in Bacillus subtilis. J Bacteriol 179: 3331–3341.
Ptacin JL, Nollmann M, Becker EC, Cozzarelli NR, Pogliano K,Bustamante C. 2008. Sequence-directed DNA export guideschromosome translocation during sporulation in Bacillussubtilis. Nat Struct Mol Biol 15: 485–493.
Ramamurthi KS, Lecuyer S, Stone HA, Losick R. 2009. Geo-metric cue for protein localization in a bacterium. Science323: 1354–1357.
Sharp MD, Pogliano K. 1999. An in vivo membrane fusion assayimplicates SpoIIIE in the final stages of engulfment duringBacillus subtilis sporulation. Proc Natl Acad Sci 96: 14553–14558.
Sharp MD, Pogliano K. 2002. Role of cell-specific SpoIIIEassembly in polarity of DNA transfer. Science 295: 137–139.
Sharp MD, Pogliano K. 2003. The membrane domain of SpoIIIEis required for membrane fusion during Bacillus subtilissporulation. J Bacteriol 185: 2005–2008.
Shroff H, White H, Betzig E. 2008. Photoactivated localizationmicroscopy (PALM) of adhesion complexes. Curr Protoc CellBiol 41: 4.21.1–4.21.27. doi: 10.1002/0471143030.cb0421s41.
Wiedenmann J, Ivanchenko S, Oswald F, Schmitt F, Rocker C,Salih A, Spindler KD, Nienhaus GU. 2004. EosFP, a fluores-cent marker protein with UV-inducible green-to-red fluores-cence conversion. Proc Natl Acad Sci 101: 15905–15910.
Wu LJ, Errington J. 1994. Bacillus subtilis SpoIIIE proteinrequired for DNA segregation during asymmetric cell di-vision. Science 264: 572–575.
Wu LJ, Errington J. 1997. Septal localization of the SpoIIIEchromosome partitioning protein in Bacillus subtilis. EMBOJ 16: 2161–2169.
Wu LJ, Errington J. 1998. Use of asymmetric cell division andspoIIIE mutants to probe chromosome orientation and orga-nization in Bacillus subtilis. Mol Microbiol 27: 777–786.
Wu LJ, Errington J. 2003. RacA and the Soj–Spo0J systemcombine to effect polar chromosome segregation in sporu-lating Bacillus subtilis. Mol Microbiol 49: 1463–1475.
Wu YX, Masison DC, Eisenberg E, Greene LE. 2006. Applicationof photobleaching for measuring diffusion of prion proteinsin cytosol of yeast cells. Methods 39: 43–49.
Yildiz A, Forkey JN, McKinney SA, Ha T, Goldman YE, SelvinPR. 2003. Myosin V walks hand-over-hand: Single fluoro-phore imaging with 1.5-nm localization. Science 300: 2061–2065.
Fleming et al.
1172 GENES & DEVELOPMENT
Cold Spring Harbor Laboratory Press on June 13, 2010 - Published by genesdev.cshlp.orgDownloaded from