
APPLIED MICROBIOLOGY, Aug. 1971, p. 171-176Copyright © 1971 American Society for Microbiology
Vol. 22, No. 2Printed in U.S.A.
Production of Double Zones of Hemolysis by CertainStrains of Hemolytic Streptococci of Groups A, B, C,
and G on Heart Infusion AgarROBERT C. NOBLE AND KENNETH L. VOSTI
Division of Infectious Diseases, Department of Medicine, Stanford Medical Center, Stanford, California 94305
Received for publication 16 February 1971
This report describes the appearance of a double zone of hemolysis around surfacecolonies of certain strains of streptococci of groups A, B, C, and G incubatedaerobically on sheep blood-Heart Infusion Agar. The occurrence of the alteredhemolytic pattern was related to peroxide production by the organism. Anaerobicconditions and the incorporation of catalase into the agar abolished the double-zone pattern and caused reversion of the organisms to a beta-hemolytic pattern. Thedouble-zone pattern can be confused with alpha hemolysis on surface growth.
Patterns of hemolysis as described by Brown(2) have, in general, been adopted as an initialstep in screening for Lancefield group A strep-tococci. Often the observation of these hemolyticpatterns is made on streak blood-agar plates,whereas Brown's description of the patterns wasintended to describe subsurface growth in pourplates. Beta-hemolytic streptococcal variants withabsent or incomplete hemolytic patterns have beendescribed previously around surface colonies onsheep blood-agar plates (2, 5-7, 9, 11, 16, 17, 19).The present report describes an additional varia-tion in the pattern of hemolysis around surfacecolonies of Lancefield group A, B, C, and G onheart infusion-blood-agar plates and suggests areason for its appearance.
MATERIALS AND METHODSOrganism. Several strains of streptococci were ex-
amined for their hemolytic patterns. Many of theserological group A and all of the serological groupsB, C, and G were fresh isolates from the StanfordMedical Center bacteriology laboratory. A number ofgroup A, M type 12 strains were donated by RichardFacklam of the Center for Disease Control, Atlanta,Ga. W. R. Maxted of the Central Public HealthLaboratory, Colindale, England, donated anothergroup of type 12 streptococci including strain no.R53/1077. K. L. Vosti of Stanford donated the strainB12/0, a type 12, and VT3, a type 3, group A strep-tococcus. Rebecca Lancefield of the RockefellerUniversity donated two group A, M type 3 strains,D5BX/11/3, 9,/20/62, and F208 (16 October 1963).
Media. The following commercial solid media(Difco) were used in preparing blood-agar plates:Heart Infusion agar, Heart Infusion agar with 2%
neopeptone, tryptic soy agar, nutrient agar, proteoseno. 3 agar, and Tryptose blood agar base. The agarplates were supplemented with either sheep, horse,rabbit, or human sterile defibrinated blood. The con-centration of defibrinated blood in the plates was 5%except in a few instances when 10% was used. BrainHeart Infusion broth (Difco) was used for growth ingrouping and typing the streptococci.
Hemolysis. The criteria of Brown were followed inobserving subsurface hemolysis (2).
Differing growth conditions. Streak or pour plates ofthe test cultures were normally carried out underaerobic conditions at 37 C for at least 48 hr. Theinocula were normally made from a 5% sheep blood-Heart Infusion agar (SBHIA) plate. The anaerobicgrowth was investigated in some cultures by incuba-tion in 10% COz-90% N2 gas exchange jars and the"Gaspak" system (BBL; reference 1).The effect of refrigeration on some strains was
tested by incubation of the cultures at 37 C for 18hr followed by refrigeration for 24 hr and then byincubation for another 18 hr. In some instances, thiscycle was repeated.
Catalase experiments. Crude sterile solution of beefliver catalase (Sigma) was incorporated into 5%SBHIA plates (1,660 units/ml) just before the additionof the sheep cells. Similar plates were prepared withheat-inactivated catalase. The plates were streakedwith the test organism, incubated aerobically for 48hr at 37 C, and examined for their hemolytic patterns.
Peroxide experiments. Hydrogen peroxide (3%)was diluted 1:10 in normal saline, and 3.0 ml wasoverlaid on SBHIA plates for 5 min. A plate overlaidwith normal saline served as a control. The plateswere drained, dried in an incubator for 30 min, andthen streaked with a known beta-hemolytic group Astreptococcus. The plates were examined at 24 and 48hr after incubation at 37 C.
171
on January 26, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from
NOBLE AND VOSTI
A
*........
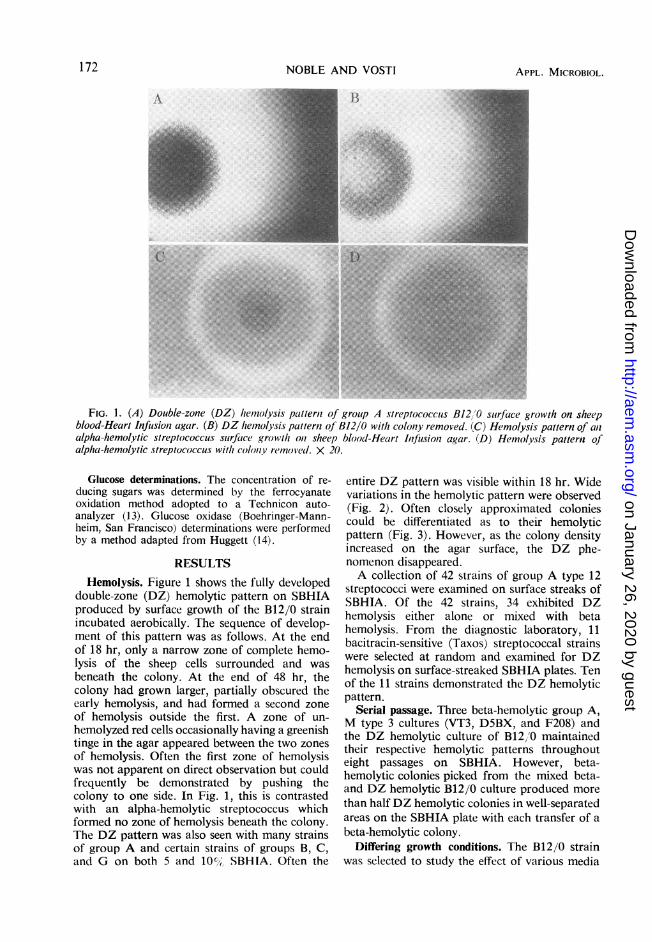
FIG. 1. (A) Double-zone (DZ) heinolysis patterli of group A streptococcus B12,'0 suirface growvth on sheepblood-Heart Infusionz agar. (B) DZ hemnolysis pattern oj B12/0 with colony removed. (C) Hemolysis pattern of anlalpha-hemolytic streptococcus suirfaice growth oni sheep blood-Heart Infusion agar. (D) Hemolysis patteri: ofalpha-hemolytic streptococcus with colonY removed. X 20.
Glucose determinations. The concentration of re-ducing sugars was determined by the ferrocyanateoxidation method adopted to a Technicon auto-analyzer (13). Glucose oxidase (Boehringer-Mann-heim, San Francisco) determinations were performedby a method adapted from Huggett (14).
RESULTSHemolysis. Figure 1 shows the fully developed
double-zone (DZ) hemolytic pattern on SBHIAproduced by surface growth of the B12/0 strainincubated aerobically. The sequence of develop-ment of this pattern was as follows. At the endof 18 hr, only a narrow zone of complete hemo-lysis of the sheep cells surrounded and wasbeneath the colony. At the end of 48 hr, thecolony had grown larger, partially obscured theearly hemolysis, and had formed a second zoneof hemolysis outside the first. A zone of un-hemolyzed red cells occasionally having a greenishtinge in the agar appeared between the two zonesof hemolysis. Often the first zone of hemolysiswas not apparent on direct observation but couldfrequently be demonstrated by pushing thecolony to one side. In Fig. 1, this is contrastedwith an alpha-hemolytic streptococcus whichformed no zone of hemolysis beneath the colony.The DZ pattern was also seen with many strainsof group A and certain strains of groups B, C,and G on both 5 and 10%i, SBHIA. Often the
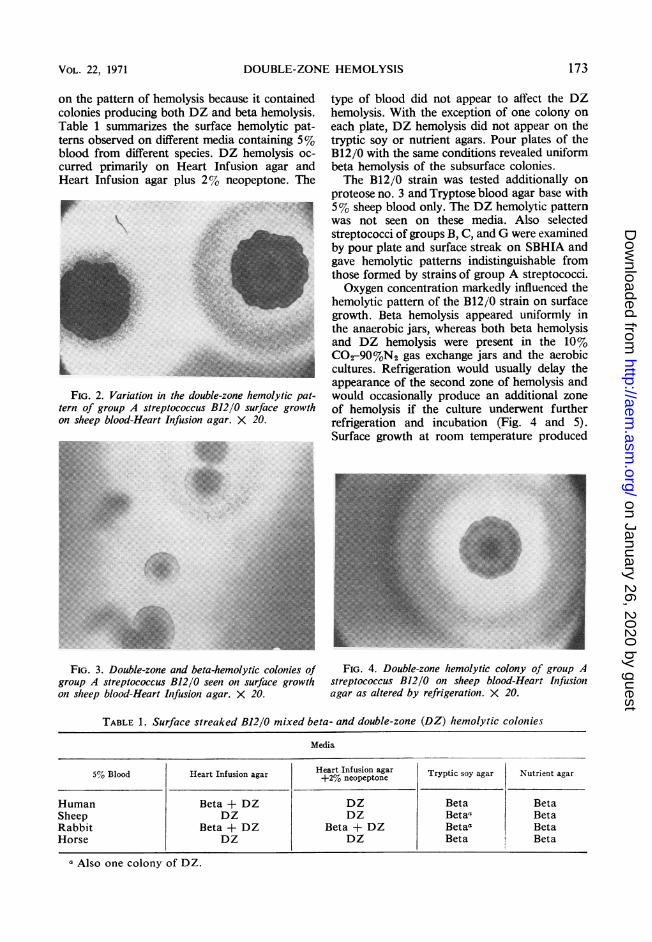
entire DZ pattern was visible within 18 hr. Widevariations in the hemolytic pattern were observed(Fig. 2). Often closely approximated coloniescould be differentiated as to their hemolyticpattern (Fig. 3). However, as the colony densityincreased on the agar surface, the DZ phe-nomenon disappeared.A collection of 42 strains of group A type 12
streptococci were examined on surface streaks ofSBHIA. Of the 42 strains, 34 exhibited DZhemolysis either alone or mixed with betahemolysis. From the diagnostic laboratory, 11bacitracin-sensitive (Taxos) streptococcal strainswere selected at random and examined for DZhemolysis on surface-streaked SBHIA plates. Tenof the 11 strains demonstrated the DZ hemolyticpattern.
Serial passage. Three beta-hemolytic group A,M type 3 cultures (VT3, D5BX, and F208) andthe DZ hemolytic culture of B12/'0 maintainedtheir respective hemolytic patterns throughouteight passages on SBHIA. However, beta-hemolytic colonies picked from the mixed beta-and DZ hemolytic B12/0 culture produced morethan halfDZ hemolytic colonies in well-separatedareas on the SBHIA plate with each transfer of abeta-hemolytic colony.
Differing growth conditions. The B12/0 strainwas selected to study the effect of various media
172 APPL. MICROBIOL.
on January 26, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from
DOUBLE-ZON
on the pattern of hemolysis because it containedcolonies producing both DZ and beta hemolysis.Table 1 summarizes the surface hemolytic pat-terns observed on different media containing 5%blood from different species. DZ hemolysis oc-cuffed primarily on Heart Infusion agar andHeart Infusion agar plus 2% neopeptone. The
s t:... ::::.|..
.|. l.:: -
*::i: t;e w[: ... w: :.: ::-: x: :.
F R S .: :., ,:;
... :::.,^>. k O S.
.S.:'..°'':itL.
FIG. 2. Variation in the double-zone hemolytic pat-tern of group A streptococcus B12/0 surface growthon sheep blood-Heart Infusion agar. X 20.~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~._E......._~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~.|no........
FIG 3. Double-zone and beta-hemolytic colonies of
group A streptococcus B12/0 seen on surface growth
on sheep blood-Heart Iifusion agar. X 20
HE HEMOLYSIS 173
type of blood did not appear to affect the DZhemolysis. With the exception of one colony oneach plate, DZ hemolysis did not appear on thetryptic soy or nutrient agars. Pour plates of theB12/0 with the same conditions revealed uniformbeta hemolysis of the subsurface colonies.The B12/0 strain was tested additionally on
proteose no. 3 and Tryptose blood agar base with5% sheep blood only. The DZ hemolytic patternwas not seen on these media. Also selectedstreptococci of groups B, C, and G were examinedby pour plate and surface streak on SBHIA andgave hemolytic patterns indistinguishable fromthose formed by strains of group A streptococci.Oxygen concentration markedly influenced the
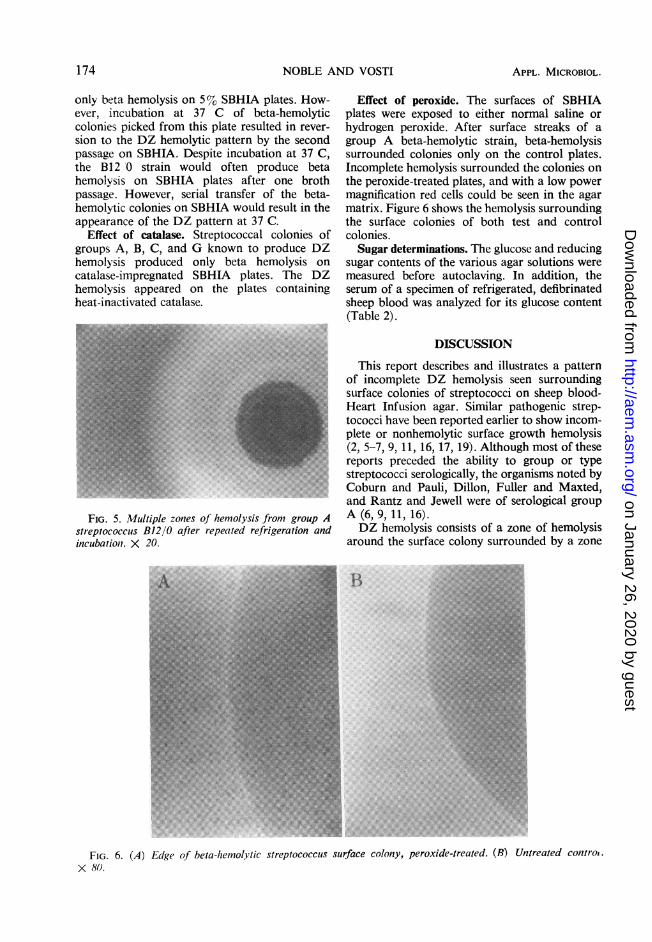
hemolytic pattern of the B12/0 strain on surfacegrowth. Beta hemolysis appeared uniformly inthe anaerobic jars, whereas both beta hemolysisand DZ hemolysis were present in the 10%C02-90%N2 gas exchange jars and the aerobiccultures. Refrigeration would usually delay theappearance of the second zone of hemolysis andwould occasionally produce an additional zoneof hemolysis if the culture underwent furtherrefrigeration and incubation (Fig. 4 and 5).Surface growth at room temperature produced
FiG. 4. Double-zone hemolytic colony of group Astreptococcus B12/0 on sheep blood-Hear.t Infu.sionagar as altered by refrigeration. X 20.
TABLE 1. Surface streaked B12/0 mixed beta- and double-zone (DZ) hemolytic colonies
Media
5% Blood Heart Infusion agar Heart Infusion agar Tryptic soy agar Nutrient agarBlood Heart nfusion agar+2% neopeptone
Human Beta + DZ DZ Beta BetaSheep DZ DZ Betaa BetaRabbit Beta + DZ Beta + DZ Betaa BetaHorse DZ DZ Beta Beta
a Also one colony of DZ.
VOL. 22, 1971
on January 26, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from
NOBLE AND VOSTI
only beta hemolysis on 5% SBHIA plates. How-ever incubation at 37 C of beta-hemolyticcolonies picked from this plate resulted in rever-sion to the DZ hemolytic pattern by the secondpassage on SBHIA. Despite incubation at 37 C,the B12 0 strain would often produce betahemolysis on SBHIA plates after one brothpassage. However, serial transfer of the beta-hemolytic colonies on SBHIA would result in theappearance of the DZ pattern at 37 C.
Effect of catalase. Streptococcal colonies ofgroups A, B, C, and G known to produce DZhemolysis produced only beta hemolysis oncatalase-impregnated SBHIA plates. The DZhemolysis appeared on the plates containingheat-inactivated catalase.
X:s~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~......FIG. 5. Multiple zones of hemolysis from group A
streptococcus B12/0 after repeated refrigeration andincubation. X 20.
Effect of peroxide. The surfaces of SBHIAplates were exposed to either normal saline orhydrogen peroxide. After surface streaks of agroup A beta-hemolytic strain, beta-hemolysissurrounded colonies only on the control plates.Incomplete hemolysis surrounded the colonies onthe peroxide-treated plates, and with a low powermagnification red cells could be seen in the agarmatrix. Figure 6 shows the hemolysis surroundingthe surface colonies of both test and controlcolonies.Sugar determinations. The glucose and reducing
sugar contents of the various agar solutions weremeasured before autoclaving. In addition, theserum of a specimen of refrigerated, defibrinatedsheep blood was analyzed for its glucose content(Table 2).
DISCUSSION
This report describes and illustrates a patternof incomplete DZ hemolysis seen surroundingsurface colonies of streptococci on sheep blood-Heart Infusion agar. Similar pathogenic strep-tococci have been reported earlier to show incom-plete or nonhemolytic surface growth hemolysis(2, 5-7, 9, 11, 16, 17, 19). Although most of thesereports preceded the ability to group or typestreptococci serologically, the organisms noted byCoburn and Pauli, Dillon, Fuller and Maxted,and Rantz and Jewell were of serological groupA (6, 9,11, 16).DZ hemolysis consists of a zone of hemolysis
around the surface colony surrounded by a zone
B 0 t-t 5tE 1g§
FIG. 6. (A) Edge of beta-hemolytic streptococcus surface colony, peroxide-treated. (B) Untreated control.X 80.
174 APPL. MICROBIOL.
rR im;4.-
on January 26, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

DOUBLE ZONE HEMOLYSIS
TABLE 2. Glucose and reducing sugar contents of various agar sohitionzs before autoclaving
Type 12,Reducing~ ~~ rou A
Agar solution Reducing Glucose sheep Other components (8)sugar ~~~~~surface
hemolysis
Tryptic soy.............. 64a Oa B Tryptose, soytoneNeopeptone............. 70 3.7 DZ Beef Heart Infusion-neopeptoneNutrient................. 31 0 B Peptone, beef extractHeart Infusion........... 64 4.8 DZ Beef Heart Infusion, TryptoseProteose no. 3........... 82 55 B Peptone, dextroseTryptose blood.......... 57 20 B Tryptose, beef extractSheep blood............. 35
a Expressed as milligrams per 100 ml.
of intact red cells which is enclosed by an outerzone of hemolysis. This should not be confusedwith either the alpha prime hemolysis which is asubsurface phenomenon with "fixed corpusclesnext to the colony" (2) or the subsurface DZhemolysis described by Brown for group Borganisms (3, 4). This subsurface pattern usuallyrequires refrigeration for its development. The"reverse alpha zone" pattern of hemolysis alsodescribed by Brown for Strep. Brig. III aa(mucosus) mimics the DZ pattern (2). However,the organism is in other respects alpha-hemolytic,and the pattern is seen only with anaerobicincubation and refrigeration of subsurfacecolonies. One instance of another pattern similarto the DZ pattern was described by Brown withstreptococcus B-7a. This pattern also occurredwith aerobic surface incubation; however, in pourplates, it was clear that the organism was alpha-hemolytic in character. The subsurface hemolysisseparates this pattern from the DZ hemolyticpatterns described in the present report, sincestrains with the latter pattern reverted to betahemolysis on subsurface growth in pour plates.The ability of sugar to suppress hemolysis on
blood plates has been known for a long time.Ruediger in 1906 noted that the addition ofglucose at 1,000 mg/100 ml to agar previouslyfree from glucose would abolish beta hemolysisand resulted in a green coloration surroundingthe streptococcal colony (17). Fuller and Maxtedfound that their green variant streptococci becamebeta-hemolytic if reducing sugar was exhaustedfrom their growth medium. With the addition ofglucose at 100 mg/100 ml of medium, the green-ing around the variants would reappear (11). Inthe present study, both DZ and beta hemolysistook place at agar glucose levels below 55 mg/100ml and reducing sugar levels below 82 mg/100mg. Since the DZ hemolysis took place at glucoselevels below those at which beta hemolysis oc-curred, it seems unlikely that the sugar content
of these agars could account for the productionof the DZ hemolysis.
Several lines of evidence suggest that the DZhemolysis is related to peroxide production in thepresence of oxygen by the surface colonies ofstreptococci. The DZ hemolytic pattern revertsto a beta-hemolytic pattern with the exclusion ofoxygen or the incorporation of catalase into themedium. In addition, Fuller and Maxted haveshown that group A, type 3 streptococci do notproduce peroxide (12), and these organisms alsodo not exhibit DZ hemolysis on Heart Infusionagar. In another study, the same authors ex-amined nonhemolytic variants of group A strep-tococci (11). The beta-hemolytic pattern of theirorganisms was also restored by catalase incorpora-tion and oxygen exclusion. In addition in theirnonhemolytic greening variants, the peroxideproduction of the streptococci preceded thehemolysis production by the organisms. Otherstudies in broth of hemolysin and peroxide pro-duction also indicate that the production of thesematerials does not appear to be simultaneous(19).Although DZ hemolysis may be caused by non-
syncronous production of the hemolysin andperoxide, the inner ring of unhemolyzed cor-puscles requires further explanation. It is possiblethat peroxide affects the red cell in such a way asto prevent the action of the hemolysin. Brownhas noted that corpuscles which surround alpha-hemolytic streptococci and which have acquireda greenish or brownish discoloration are no longersubject to hemolysis by beta-hemolytic strep-tococci (2). Before the full development of theDZ hemolytic pattern, the inner hemolytic zoneis often surrounded by an outer zone of greenishdiscolored corpuscles. This discolored zone be-comes the inner ring of unhemolyzed corpusclesand its final width often decides whether itsgreenish discoloration remains. The capacity ofa beta-hemolytic, group A streptococcus to pro-
VOL. 22, 1971 175
on January 26, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

176 NOBLE AND VOSTI
duce a greening reaction has been demonstratedby Stinebring and Morton (18). With a cello-phane sack technique, they demonstrated identicalrates of conversion of oxyhemoglobin to methe-moglobin by a group A and a Viridans strep-tococcus (18). In addition in the present report,dilute solutions of peroxide overlaid on SBHIAplates prevented complete hemolysis of the cor-puscles surrounding the beta-hemolytic colonies.Thus, it is probable that the DZ hemolysis patternstarts with early hemolysis production whichcauses the clear zone surrounding the colony.Subsequent peroxide formation by the colonyprobably fixes the red cells at the periphery of thehemolyzed area. This in turn prevents lysis of thecorpuscles when the hemolysin again diffuses out-ward to form the second zone of hemolysis.The expression of the DZ pattern seems to be
limited to the Heart Infusion and neopeptoneagars of those agars tested. Beef heart infusion isa common ingredient of both these agars. In someof the reports previously cited, the blood-agar basewas not specified, but, in many, meat infusionwas a common ingredient (2, 5-7, 16). However,the medium used by Fuller contained trypticdigest of ox hearts and that used by Dillon con-tained a papain digest of ox liver (9, 11, 15).
Because of the wide variation in the DZhemolytic pattern and the common occurrence ofgreening in the unhemolyzed zone, confusionbetween alpha and DZ hemolysis on surfacestreaks is likely to occur. Fortunately in heavygrowths of DZ hemolytic organisms in groups A,B, C, and G, the densely concentrated coloniesappeared beta-hemolytic and only the relativelywell-separated colonies demonstrated the DZpattern. Also alpha-hemolytic surface coloniesdo not typically have an inner zone of hemolysiswith lysin underlying the colony. However, acharacteristic shared by the surface DZ coloniesand the alpha-hemolytic colonies is the formationof additional zones of hemolysis with alternateincubation and refrigeration as demonstrated byBrown (2). This DZ phenomenon has not pre-viously been demonstrated with group A orga-nisms.The phenomenon of the DZ hemolytic pattern
should be considered in the selection of a blood-agar base used in screening for beta hemolysis.If either Heart Infusion or neopeptone agars areto be used, reduced oxygen conditions must beprovided by anaerobic jars, pour plates, or sub-surface cuts into the agar as suggested by Fry(10). It should be noted that the gas exchangemethod did not prevent the DZ hemolysis. Theuse of such media without providing the cor-responding reduced oxygen environment may re-
APPL. MICROBIOL.
sult in confusion of the relatively nonpathogenicalpha-hemolytic streptococci with potentiallypathogenic DZ hemolytic streptococci of groupsA, B, C, and G.
ACKNOWLEDGMENTS
We thank Paul L. Wolf and George W. H. Bailey for theirassistance in performing the sugar determinations.
This investigation was supported by a National Institutes ofHealth Special Fellowship award no. 5 F03 A144651-02 and byPublic Health Service grants AI-06964 and TI-AI-185 fromn theNational Institute of Allergy and Infectious Diseases.
LITERATURE CITED
1. Brewer, J. H., and D. C. AlIglier. 1966. Safe self-contaiinedcarbon dioxide-hydrogen anaerobic system. Appl. Micro-biol. 14:985-988.
2. Brown, J. H. 1919. The use of blood agatr for the study ofstreptococci. Rockefeller Institute Medical Research Mono-graph No. 9.
3. Brown, J. H. 1937. Appearance of double-zone beta hemolyticstreptococci in blood agar. J. Bacteriol. 34:35-48.
4. Brown, J. H. 1939. Double-zonie beta-hemolytic streptococci:their cultural characteristics, serological grouping, occur-rence and pathogenic significance. J. Bacteriol. 37:133-144.
5. Clawson, B. J. 1920. Further observations on varieties ofstreptococci with reference to hemolysis. J. Infec. Dis. 27:368-377.
6. Coburn, A., and R. Pauli. 1941. The interaction of host andbacterium in the developmiient of communicability ofStreptococcuis haennolyticus. J. Exp. Med. 73:551-570.
7. Colebrook, L., S. D. Elliott, W. R. Maxted, C. W. Morley,and M. Mortell. 1942. Infection by non-haemolytic groupA streptococci. Lancet 2:30-31.
8. Difco Laboratories. 1959. Difco Mauiual, 9th ed. Difco Labo-ratories, Detroit.
9. Dillon, H. C. 1968. Impetigo conitagiosa: suppurative and nonl-suppurative complicationis. Am-ier. J. Dis. Child. 115:530-541.
10. Fry, R. M. 1933. Anaerobic mnethods for the identification ofhaemolytic streptococci. J. Pathol. BRcteriol. 37:337-340.
11. Fuller, A. T., and W. R. Maxted. 1939. The production ofhemolysin and peroxide by haemnolytic streptococci in rela-tion to the non-haemolytic variants of group A. J. Pathol.Bacteriol. 49:83-94.
12. Fuller, A. T., and W. R. Maxted. 1939. A type of group Ahaemolytic streptococcus which fails to form peroxide.Brit. J. Exp. Pathol. 20:177-181.
13. Hoffman, W. S. 1937. A rapid photoelectric method for thedetermination of glucose in blood and urine. J. Biol. Chemn.120:51-55.
14. Huggett, A. St. G., and D. A. Nixon. 1957. Use of glucoseoxidase, peroxidase and O-dianiisidine in determination ofblood and urinary glucose. Lancet 2:368-369.
15. Oxoid Limited. 1969. Oxoid manual, 3rd ed. Oxoid Limllited,London.
16. Rantz, L. A., and M. L. Jewell. 1940. The relationship ofserological groups A, B and C of Lancefield to the type ofhemolysis produced by streptococci in poured blood agarplates. J. Bacteriol. 40:1-8.
17. Ruediger, G. V. 1906. The cause of green coloration of bac-terial colonies in blood-agar plates. J. Infec. Dis. 3:663-665.
18. Stinebring, W. R., and H. E. Morton. 1951. Studies of theaction of streptococci on blood with their application to
metabolism and variation of streptococci. J. Bacteriol. 62:395-403.
19. Todd, E. W. 1928. The conversion of hemolytic streptococcito non-hemolytic form. J. Exp. Med. 48:493-511.
on January 26, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from



![Streptococcus pseudoporcinus subacute mitral valve ......Streptococcus pseudoporcinus is a β-hemolytic Streptococcus with a wide-zone hemolysis first identified in 2006 [1]. Prior](https://cdn.vdocuments.site/doc/165x107/5f419238528e526df81403df/streptococcus-pseudoporcinus-subacute-mitral-valve-streptococcus-pseudoporcinus.jpg)



