Download - Encyclopedia of Inland Waters || Protists

ProtistsR W Sanders, Temple University, Philadelphia, PA, USA
ã 2009 Elsevier Inc. All rights reserved.
What are Protists?
The protists are an evolutionarily diverse groupthat includes most of what have been historicallycalled protozoa, including ciliates, amoeba, hetero-trophic flagellates, oomycetes, and slime molds plusphytoplankton, periphyton, and algae (red, green,and brown). Protists are primarily unicellular eukary-otes with heterotrophic and/or photosynthetic nutri-tion that are not plants, animals, or fungi. Likeother eukaryotes, protist cells possess at least onemembrane-bound nucleus plus endoplasmic reticu-lum and other organelles that can include mitochon-dria, chloroplasts, and flagella or cilia. Some protistsare colonial or related to multicellular groups, butnone develop tissue. The different ecological rolesfilled by protists within clearly recognized phyla ledto some subgroups being historically described byboth zoological and botanical nomenclature. Forexample, many dinoflagellate genera contain bothphototrophic and heterotrophic species, and in addi-tion to the occurrence of both photosynthetic andheterotrophic euglenids, some of photosyntheticspecies can be induced to lose their chloroplasts andsurvive using dissolved organic matter for energyand carbon.In earlier taxonomic schemes ‘Protista’ was consid-
ered a Kingdom, but advances in systematics haveshown that there are groupings within the protiststhat are not taxa of single origin (monophyletic) orthat do not contain all the descendents of a commonancestor (holophyletic). Hence, protist is not a phy-logentically useful term. Rather, it is a term of conve-nience that describes an assemblage of often distantlyrelated organisms lumped together as a matter ofecological utility. The protists do have lineages thatare properly organized to represent evolutionary rela-tionships. Some clades, which are groups of ancestralspecies and all their known descendents, includeonly protists. Alveolata (ciliates, dinoflagellates,and apicomplexa), Cercozoa (amoeba and amoeba-flagellates with fine pseudopods or filipodia), andEuglenozoa (euglenids, kinetoplastids, and diplone-mids) typify purely protistan clades. Other clades,however, indicate a shared ancestry between someprotists and plants or animals that is closer thantheir evolutionary relationship to other protists.For example, the choanoflagellates group with truefungi, chytrids, and animals as Opisthokonts, and thegreen algae, including common freshwater species
252
like Chlamydomonas and Volvox, group with thePlantae.
As one might expect from such an evolutionarilyand ecologically diverse group, making generaliza-tions about the protists must be a cautionary exercise.Yet, for the practicing field biologist or ecologist,the small size and functional similarities of manyof the protists makes grouping them together a ration-al practice. Biologists often still group the protistsby their primary means of locomotion into flagella-tes, ciliates, and amoeba. These subdivisions areusually easily discernable with light microscopy andare retained here, not because they reflect currentunderstanding of phylogeny, but rather because theyoften reflect niches and form the basis of much ofthe study of community ecology in freshwater protiststo date.
Morphological Groups
Flagellates
Flagellates have one-to-many flagella that function inmotility, attachment to a substrate, and also for feed-ing. They are phylogenetically diverse (Table 1), andare themost numerous of the freshwater protists reach-ing abundances in eutrophic systems that can approach105 cellsml�1 and 106 cells cm�3 in the plankton andsediments, respectively. Purely heterotrophic free-liv-ing flagellates include choanoflagellates and bicosoe-cids. Choanoflagellates, easily recognized by theirdistinctive ‘collar’ of microvilli and a single flagellum,can have a lorica that is sometimes difficult to see withlight microscopy. They are most often attached and/orcolonial in freshwaters. Bicosoecids are heteroconts –possessing two different types of flagella – and areoften attached by one of the flagella to a lorica or tothe substrate. Cercazoans have two flagella, but areamoeboflagellates that use pseudopodia for feeding.Some species traditionally considered as amoeba arenow also classified as Cercazoa (explained later).
Among the several taxa of photosynthetic flagellatesare a number of groups with members that are whollyheterotrophic or that combine photosynthetic andheterotrophic nutrition (mixotrophy). Dinoflagellates,chrysomonads, euglenids, and cryptomonads havenumerous phagotrophic or osmotrophic species thatlack chloroplasts. Phagotrophy of bacteria or otherprotists by chloroplast-containing species has also

Table 1 Some common genera of heterotrophic flagellates
Group Genera Comments
Opisthokonta
Choanomonada Collar flagellates, with a single flagellum surrounded by cytoplasmic ‘fingers’ used
in feeding on bacteria; mostly sessile/attached; Salpingoecawith a cellulose thecaMonosigidae Monosiga
Salpingoecidae SalpingoecaRhizaria
Cercazoa Cercomonas Bacterivorous using pseudopods; amoebo-flagellates
ChromalveolataHaptophyta Chrysochromulina Some haptophytes in marine systems produce ichthyotoxin, although this has not
been demonstrated in fresh waters
Straminopiles
Bicosoecida Bicosoeca Immotile in a lorica, filter feeder on bacteriaChrysophyceae Actinomonas,
Ciliophrys
Pedinellids, single flagellum, usually stalked, filter feeds on bacteria
Paraphysomonas One short and one long flagella; silica spines on cell body used to identify species
requires electron microscopySpumella (Monas) With light microscopy nearly identical to Paraphysomonas.
Ochromonas Typically with chloroplast, which may be reduced in mixotrophic populations using
osmotrophy or phagotrophyCryptophycea Goniomonas,
Chilomonas
Bacterivorous; Chilomonas species were recently reassigned, with freshwater
members assigned as heterotrophic Cryptomonas
Cryptomonas Primarily photosynthetic species, some of these with low rates of phagotrophy
reported (mixotrophy)Alveolata
Dinoflagellata Gymnodinium Two flagella, one horizontal around cell that gives slow spinning motion when
swimming; photosynthetic, predatory and mixotrophic species
ExcavataEuglenozoa
Euglenida Peranema, Entosiphon,
Astasia
Most euglenids have two flagella emerging from an anterior pocket, with one trailing
and usually in contact with the substrate. Colorless species ingest detritus,
bacteria and/or are osmotrophicKinetoplastea Bodo, Rhyncomonas Only two common genera of free-living kinetoplastids; generally glide on substrates
like euglenids and ingest attached bacteria
The higher ranking follows the ‘nameless-rank’ system of Adl et al. (2005), which considers the taxonomic endings to be accidents of history without the
traditional hierarchical meaning of past classification codes. Some rankings are tentative, including the union of Alveolata with Haptophycea,
Stromenopiles and Cryptophysea into Chromalveolata. Most of the higher groups include photosynthetic flagellates, some of which combine
heterotrophic and phototrophic nutrition.
Protists, Bacteria and Fungi: Planktonic and Attached _ Protists 253
been reported for dinoflagellates, chrysomonads, andcryptomonads, and is discussed further in this articleand elsewhere in this volume.Many recent ecological field studies separate fla-
gellates into different groups based primarily on size.This is most common in examinations of the micro-bial food web, where heterotrophic nanoflagellates(HNAN) are enumerated collectively due to theirhigh abundance, importance as bacterivores, and thedifficulty of separating taxa in preserved samples.This size-based assemblage originally describedplankton that was�20 mm in diameter, but flagellates�5mm often dominate this size category. It is difficultto separate individuals this small into different taxa-nomic groups by light microscopy; even divisioninto functional groups of photosynthetic versus het-erotrophic flagellates is problematic without properfixation and slide preparation methods. Conse-quently, care must be taken to determine if reportedresults combined all small protistan cells into
one trophic category or functionally divide groupsinto autotrophic and heterotrophic forms (usuallybased on the presence or absence of chloroplasts).Heterotrophic flagellates larger than 20mm are typi-cally much less abundant than nanoflagellates, andinclude many of the dinoflagellates and euglenids(Figure 1).
Amoeboid Protists
Amoebae are plastic cells that lack a fixed externalmorphology (Figure 1). They move primarily usingpseudopodia with shapes that vary in different taxafrom finely pointed and branching filose extensionsto blunt lobopodia. The pseudopodia are used bothfor movement and for engulfing food particles. Sev-eral amoeboid groups have flagella during temporaryswimming stages for dispersal, and some of these,including the stalked heliozoans, have affiliationwith Cercozoa or Stamenopiles (Table 2).
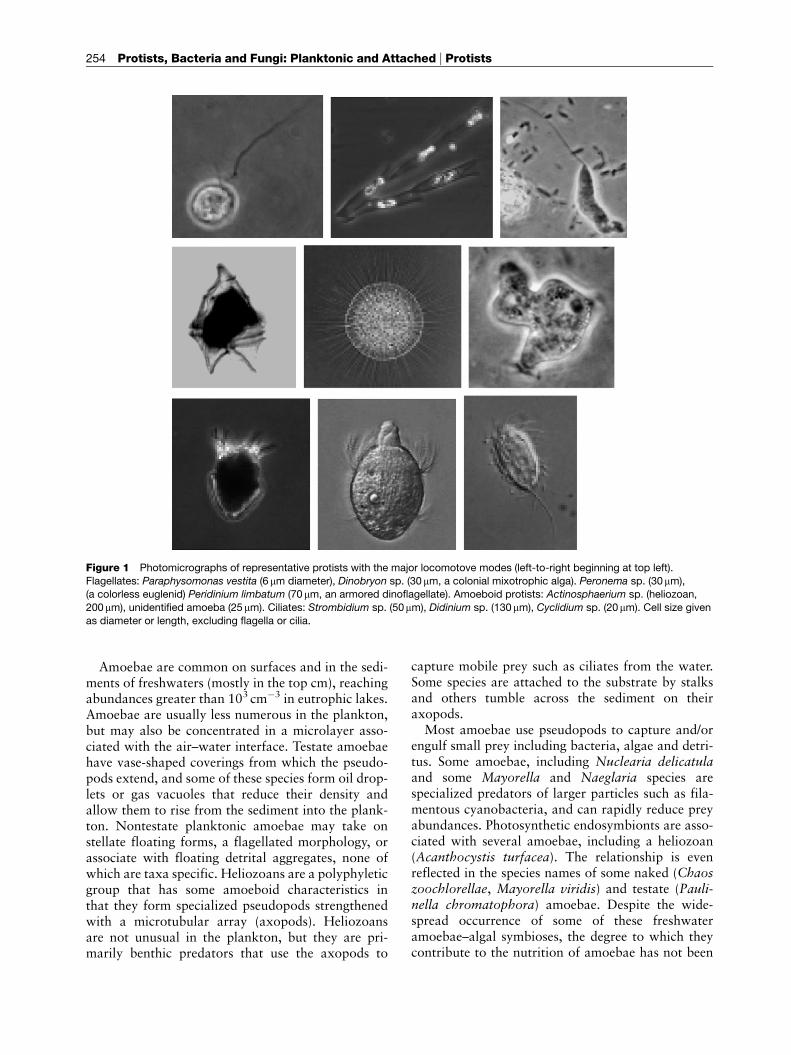
Figure 1 Photomicrographs of representative protists with the major locomotove modes (left-to-right beginning at top left).
Flagellates: Paraphysomonas vestita (6 mm diameter), Dinobryon sp. (30mm, a colonial mixotrophic alga). Peronema sp. (30mm),(a colorless euglenid) Peridinium limbatum (70mm, an armored dinoflagellate). Amoeboid protists: Actinosphaerium sp. (heliozoan,
200mm), unidentified amoeba (25 mm). Ciliates: Strombidium sp. (50mm), Didinium sp. (130mm), Cyclidium sp. (20mm). Cell size given
as diameter or length, excluding flagella or cilia.
254 Protists, Bacteria and Fungi: Planktonic and Attached _ Protists
Amoebae are common on surfaces and in the sedi-ments of freshwaters (mostly in the top cm), reachingabundances greater than 103 cm�3 in eutrophic lakes.Amoebae are usually less numerous in the plankton,but may also be concentrated in a microlayer asso-ciated with the air–water interface. Testate amoebaehave vase-shaped coverings from which the pseudo-pods extend, and some of these species form oil drop-lets or gas vacuoles that reduce their density andallow them to rise from the sediment into the plank-ton. Nontestate planktonic amoebae may take onstellate floating forms, a flagellated morphology, orassociate with floating detrital aggregates, none ofwhich are taxa specific. Heliozoans are a polyphyleticgroup that has some amoeboid characteristics inthat they form specialized pseudopods strengthenedwith a microtubular array (axopods). Heliozoansare not unusual in the plankton, but they are pri-marily benthic predators that use the axopods to
capture mobile prey such as ciliates from the water.Some species are attached to the substrate by stalksand others tumble across the sediment on theiraxopods.
Most amoebae use pseudopods to capture and/orengulf small prey including bacteria, algae and detri-tus. Some amoebae, including Nuclearia delicatulaand some Mayorella and Naeglaria species arespecialized predators of larger particles such as fila-mentous cyanobacteria, and can rapidly reduce preyabundances. Photosynthetic endosymbionts are asso-ciated with several amoebae, including a heliozoan(Acanthocystis turfacea). The relationship is evenreflected in the species names of some naked (Chaoszoochlorellae, Mayorella viridis) and testate (Pauli-nella chromatophora) amoebae. Despite the wide-spread occurrence of some of these freshwateramoebae–algal symbioses, the degree to which theycontribute to the nutrition of amoebae has not been

Table 2 Some common genera of amoeboid protists and their food
Group Genera Comments Food
Amoebozoa
Tubulinea
Tubulinida Amoeba, Chaosa, Hartmannella Bacteria and/or small protists
Testacealobosia Arcella, Difflugiaa Testate/shelled, benthic andplanktonic stages
Omnivores, algae, detritus
Flabellinea
Dactylopodida Mayorellaa Predators of protists, detritusVannellida Vannella, Platyamoeba Benthic and planktonic Primarily bacteria
Acanthamoebidae Acanthamoeba Some species are facultative human
pathogens
Primarily bacteria
ExcavataHeterolobosea
Vahlkampfilidae Naegleria, Tetramitus,
Vahlkampfia
Bacteria, cyanobacteria
OpisthokontaMesomycetozoa
Nucleariida Nuclearia Bacteria, cyanobacteria
Heliozoansb
CercozoaNucleohelea
Clathrulinidae Clathrulina, Hedriocystis Benthic, sessile, often stalked,
sometimes colonial
Fine particulate matter, including
bacteriaStramenopiles
Actinophryidae Actinosphaerium, Actinophrys Planktonic or mobile benthic Other protists and rotifers
Centrohelida
Acanthocystidae Acanthocystis Planktonic Other protists, phytoplanktonHeterophridae Heterophrys, Chlamydaster Planktonic
The higher ranking follows Adl et al. (2005), which considers the taxonomic endings to be accidents of history and without the traditional hierarchical
meaning of past codes of classification. Many amoebae have flagellated stages, and flagellated forms dominate the life stages of some groups (e.g.,
Stramenopiles, Cercazoa).aSome spp. with zoochlorellae or photosynthetic symbionts.bHeliozoans have some general similarities, which include radiating stiffened pseudopodia, but the group is not monophyletic. Some heliozoan species
have flagellated and/or free amoeboid stages and the group falls across taxonomic lines that include flagellates or are of uncertain affinities.
Protists, Bacteria and Fungi: Planktonic and Attached _ Protists 255
well studied. Some amoebae, including Naegleriafowleri and one or more Acanthomoeba are faculta-tive pathogens of humans – the causative agents ofamoeboid meningitis.
Ciliates
Ciliophora is one protistan lineage that was groupedtogether historically and is still recognized as mono-phyletic. Cilia are present during some stage of the lifecycle in all ciliates, and are arranged in lines (kinities).The arrangement of kinities varies from a sparse distri-bution to a dense covering of the whole cell, and is animportant morphological feature for ciliate identifica-tion. Cilia are sometimes arranged compactly to formcompound organelles, such as cirri that are used formovement on surfaces, or as membranelles that directcurrents toward the cytostome (‘mouth’). Similar instructure to eukaryotic flagella, the cilia move wateracross the cell and contribute to both locomotion andfood capture. Another morphologically distinguishingcharacter of ciliates is nuclear dimorphism, in which
each cell has a least one macronucleus and one micro-nucleus. The macronucleus has multiple copies of aciliates genome and controls RNA synthesis for cellu-lar regulation. The diploid micronuclei undergo meio-sis and are exchanged during conjugationwith anothercell when ciliates reproduce sexually.
Most ciliates are motile and have adaptations thatallow them to move into the plankton (Figure 1), butmany species are more commonly associated with thebenthos. There also are species that use stalks, lori-cae, or a gelatinous matrix to attach to substrates.Some attached species are found frequently in thewater column attached to detritus or as epibionts onplanktonic organisms, including fish. Many ciliatesare omnivorous, ingesting detritus, bacteria, andother small protists or even metazoans. Others arespecialized bacterivores, algivores, or carnivores.Bacterivores and algivores typically generate watercurrents with cilia that bring prey toward the cyto-some; others use cilia to sweep attached or settledparticles from detritus or the substrate. Predatoryciliates often use toxicysts or extrusomes, which

256 Protists, Bacteria and Fungi: Planktonic and Attached _ Protists
immobilize active prey. Ciliate species with endosym-biotic algae are fairly common and occasionally mayreach biomass levels equal to that of phytoplankton,thus contributing substantially as primary producers.In addition, there are species that retain functionalchloroplasts from ingested algal prey (kleptoplasty).Although identification at the level of ‘ciliate’ seems
straightforward, ciliature, size, and shape vary consid-erably. One group of sessile and predatory ciliates, thesuctoria, has cilia only during its dispersal stages(Table 3). Additionally, ciliates overlap in size withseveral metazoan groups, such as rotifers, gastrotrichs,and turbellarians, which also are ciliated and can bemistaken for ciliates by beginning microscopists.
Reproduction and Population Growth
Protists reproduce asexually by binary fission, whichcan lead to rapid population growth. For heterotro-phic protists, population growth rates are generally
Table 3 Some common genera of ciliates and their freshwater hab
Group Genera Habitat
Karyorelictea Loxodes Planktonic and ben
Heterotrichea Blepharisma, Stentora Mostly motile, plan
also attached to
SpirotrichiaHypotrichia Aspidisca, Euplotes Mostly motile, ben
Oligotrichia Strombidiuma,
Halteria
Mostly planktonic
Choreotrichea Stobilidium Planktonic
Stichotrichia Stylonychia Mostly planktonic
Armophorea Metopus,
Caenomorpha
Planktonic, anaero
Litostomatea
Haptoria
Didinium, Dileptus,
Lacrymaria
Motile; planktonic
Phyllopharyngea
Suctoria
Tokophyra,
Discophyra
Sessile; cilia only d
dispersal stageNassophorea Nassula,
Pseudomicrothorax
Motile, benthic and
Colpodea Colpoda, Bursaria Motile, benthic or p
Protostomatea Colepsa, Prorodona,Urotricha
Mostly benthic, so
Oligohymenophorea
Hymenostomata Colpidium, Glaucoma,Tetrahymena
Mostly benthic
Peniculia Frontoniaa,
Parameciuma,
Stokesiaa
Benthic and plankt
Scuticociliata Cyclidium,
Pleuronema,
Lembadion
Mostly planktonic
Peritrichia Vorticellaa,Ophridiuma, Epistylis
Sessile, attached bepiplanktonic
The higher ranking follows Adl et al. (2005). Most of the groups include specieaGenera have at least one species with photosynthetic symbionts or sequeste
positively related to food concentration and tempera-ture up to some critical level, and negatively related tospecies cell size. But generation times are highly vari-able and maximum growth rates of some species withrelatively large size can approach those of much smal-ler protists. Populations of many protists can doubleseveral times per day in laboratory situations,although division rates of one to several days areprobably more realistic for natural populations wherefood and predators may limit growth. Protistan pro-ductivity in natural systems, particularly in the ben-thos, is not well documented and is likely higher thanis generally recognized. The high potential growthrates enable heterotrophic protists to persist duringperiods of high predation, and to respond rapidly toincreases in fast-growing prey such as bacteria andphotosynthetic protists.
Sexual reproduction is known for many protistsand it takes varied forms, but the essential featuresare formation of gametic cells or nuclei, fusing of thenuclei and halving of the chromosome number at
itats
Feeding mode/food
thic Omnivores, other protists, bacteria
ktonic, but
substrate
Mostly filter feeders on bacteria, small protists
thic Other protists and bacteria
bic Mostly filter feeders on bacteria, typically
dependent on methanogenic endosymbiontsand benthic Predators on motile protists
uring Predators on motile protists
planktonic Algae, cyanobacteria
lanktonic Mostly filter feeders on bacteria, small protists
me planktonic Omnivores; detritus, protists, decomposinganimals
Mostly filter feeders on bacteria, small protists
onic
y stalk, some
s from several habitats, with the most common habitat listed first.
red chloroplasts.

Protists, Bacteria and Fungi: Planktonic and Attached _ Protists 257
some point in the life cycle. Sex in protists is rarelyassociated with increases in the number of individ-uals; it interrupts asexual reproduction and the rapidpopulation growth associated with binary fission.Sex appears to be essential for preventing senescenceand extinction of clones of several protists, includingParamecium, but is unknown in some others. Autog-amy, in which the meiotic products fuse in the samecell, only stalls senescence in Paramecium, but isthe normal pattern in the heliozoan Actinophrys.When sexual reproduction does occur, it is usuallylinked to environmental change, and in some casesto the formation of cysts.Cyst formation is not always associated with sex in
protists. Some protists undergo binary division incysts, while others do not increase in number duringencystment. Whether or not encystment is associatedwith reproduction, it is frequently a response to envi-ronmental stress. The ciliateColpoda and the amoebaAcanthamoeba, common in temporary bodies ofwater and soil, will encyst due to a variety of signalsthat are linked to drying. Food shortage, increasedsalt concentrations, high temperature, and low oxy-gen all can initiate formation of desiccation-resistantcysts. For species associated with temporary aquatichabitats, this can be an important means of survivalas well as dispersal.
Distribution of Protists
In the Pelagic Zone
Protists are able to position themselves in the watercolumn through several mechanisms, including pro-duction of gas or lipid vacuoles noted previously forthe testate amoebae Difflugia. However, swimmingwith flagella or cilia is the most common way thatmotile algae, ciliates, and heterotrophic flagellatesposition themselves in response to a variety of gradi-ents in a stratified water column. Light requirementsobligate phototrophs to maintain themselves in theeuphotic zone, but many phototrophic and somepigmented heterotrophic organisms, such as ciliatesin the genus Blepharisma, will swim to avoid brightlight. Motile photosynthetic organisms, such as dino-flagellates and ciliates with algal symbionts, areknown to migrate toward the surface in the morningand into deeper, potentially nutrient-richer layerslater in the day. Most heterotrophic protists tend torespond to other environmental parameters such asoxygen or food abundance. Some heterotrophs,including the ciliates Metopus and Caenomorpha(Table 3), are obligate anaerobes and are found only
in anoxic areas of sediments and hypolimnia of eutro-phic waters. The ciliate Loxodes prefers low oxygenenvironments and will migrate out of anoxic sedi-ments and congregate within a specific low oxygenlayer in the overlying water. Loxodes uses geotaxislinked to oxygen concentration to maintain its posi-tion in the oxycline. Algivorous protists will tend toaccumulate in layers where light and nutrient levelsare best for their prey, just as the greatest abundanceof bacterivorous protists, especially heterotrophicflagellates, are associated with areas of higher bacte-rial production.
Aggregates of detritus occur in the water column (aswell as the benthos) and these will often have higherabundances of bacteria. Increases of bacterial preylead to higher abundances of certain heterotrophicprotists on these microhabitats relative to the sur-rounding water – especially in oligotrophic systems.Occurrence of ‘benthic’ protists, such as the flagellateBodo and the ciliate Colpoda, in the water columnmay be due to association with these aggregates.Another distinct habitat, the air–water interface, alsocan be enriched with protists that use the surface as asubstrate. Both naked and testate amoebae, such asArcella, various ciliates and flagellates (e.g., Codono-siga) can hang below or move atop the water’s surfacefeeding on bacteria or phototrophs growing there.
There tend to be strong seasonal changes in theabundance and species composition of aquatic pro-tists. Phototrophic species often decline in winter andincrease again with the longer days and greater lightintensity in the spring. Bacterial activity increases withautotroph activity, since the photosynthetic organismsrelease dissolved organic matter and eventually die,and both of these processes supply substrate for bac-teria. Heterotrophic protists will increase in responseto abundance of algal and bacterial prey, and canequal or exceed zooplankton biomass during periodswhen their prey are abundant. Protistan populationgrowth rate also tends to be positively related to tem-perature so the low photosynthesis and low growthrates of winter lead to minimum abundances, whileprotistan productivity tends to increase in warmerseasons.
In the Benthos
The benthos in both lotic and lentic systems tendsto accumulate organic matter and detritus, whichoffers both a food source and a habitat for benthicprotists. Attached and motile protists can occur inhigh numbers on and in the sediments. Attached het-erotrophic organisms typically create a current to

258 Protists, Bacteria and Fungi: Planktonic and Attached _ Protists
bring suspended food from the water (e.g., peritrichciliates), or allow material to fall or swim intospecialized feeding appendages (suctoria and heliozo-ans). Phototrophic species can form mats or biofilmsthat often have high abundances of cyanobacteriawith or without protistan phototrophs. Motile proto-zoa move on and through these biofilms, and also atthe surface and within the sediment. In sedimentswith relatively high organic matter content, gradientsof oxygen, redox potential, pH, and other chemicalsset up with sediment depth. As noted for the plank-ton, species that prefer oxic, microaerophyllic, oranoxic environments will move to appropriate depthsin the sediment or will even leave the sediment andmove into the water column. The sediments are asource of the anaerobic protozoa that appear in thewater column when anoxic hypolimnia develop, typ-ically during summer, in stratified lakes.
Ecological Roles and the Dynamics ofProtists in Freshwater Systems
Primary Production
Although the influx of terrestrial carbon and its utiliz-ation in aquatic systems suggest that many lentic andlotic systems may tend toward net heterotrophy (i.e.,ecosystem respiration exceeds primary production),photosynthesis still contributes significantly to thefood webs of freshwater systems. Protists, includingperiphyton and phytoplankton, are major contri-butors to total aquatic primary production and toeutrophication. Photosynthetic species dominate theprotistan biomass in the euphotic zones of lakes, butsize, depth and nutrient concentration of a lake deter-mine whether these phytoplankton are the major pri-mary producers. The deeper the lake and the less areain the littoral zone, the more pronounced the phyto-plankton contribution to primary production becausethe bulk of the benthic area is not within the euphoticzone. If dissolved nutrient levels are sufficient, phyto-plankton also can reduce the light reaching sedi-ments, and consequently shade the communities ofperiphytic algae (including photosynthetic protists)and submerged macrophytes. In shallow lakes, eutro-phication may thus lead to a switch from a benthic toa pelagic dominance of primary production. In oligo-trophic systems, benthic protists can contribute moreto whole-lake primary production; light attenuationdoes not so strongly limit photosynthesis in deeperwater and nutrients are available from the sediment.In streams and rivers, water depth and sediment load
similarly affect the relative importance of the periph-yton and phytoplankton, with periphyton contribut-ing relatively more in low- to middle-order streamswhere more light reaches the benthos.
Food Webs and Protists
Advances in the last 30 years that produced dataindicating high abundances and productivity of bac-teria in aquatic systems also triggered hypothesesabout the influence of protists on the populationdynamics of bacteria. The microbial loop concept,as developed for planktonic marine systems, recog-nized that bacterioplankton incorporate dissolvedorganic matter into biomass and that bacteria aresubsequently ingested by heterotrophic protists.Thus, fixed organic carbon that was once consideredunavailable to most pelagic organisms could be recov-ered and funneled into the rest of the pelagic foodweb via a protistan link. Conversely, if there is efficientremineralization by bacteria and protists, little of theorganic matter will immediately transfer to highertrophic levels, though this does not mean that it isnecessarily lost from the system.
Individual feeding rates of protists tend to beinversely correlated with size; nanoflagellates ingest afew to 100 bacteria per flagellate per hour, while someciliates ingest >1000 bacteria per hour. In freshwatersystems, heterotrophic protists tend to be the majorconsumers of bacteria, and heterotrophic flagellatesalone can sometimes remove more than 100% of thedaily planktonic bacterial production. The usual domi-nance of bacterivory by heterotrophic flagellatesreflects their high abundances, but ciliates and mixo-trophic protists also can strongly reduce the abundanceand alter the species composition of bacterioplankton –as can rotifers and cladocerans. These multiple tro-phic pathways moving microbial biomass into thehigher trophic levels suggest that the ‘microbial loop’should be considered a highly dynamic and persis-tent component of aquatic food webs rather than anindependent entity that is distinct from the primaryproduction–herbivore trophic pathway.
The impact of heterotrophic protists feeding onphototrophic protists in the plankton has receivedless attention than the trophic interactions of protistsand bacteria. Many species of ciliates, amoebae, anddinoflagellates ingest phytoplankton, but rotifers andcrustacean zooplankton tend to cause more phyto-plankton grazing mortality. However, because oftheir efficiency in capturing small particles, protistanherbivores are relatively more important if pico-autotrophs dominate primary production. But protists

Protists, Bacteria and Fungi: Planktonic and Attached _ Protists 259
also ingest larger cells, and both amoebae and ciliatescan reduce populations of larger phototrophs. Forexample, the amoeba Nuclearia has been investigatedas a potential biocontrol agent because of its highingestion rates when feeding on filamentous algaeand cyanobacteria. In sediments, protistan herbivoryaccounts for<5%ofperiphytonproduction.Likewise,studies to date suggest that protists ingest only about5% of the daily bacterial production in sedimentsdespite having feeding rates similar to those measuredin the plankton. Under long-term anerobic conditionsbacterivory by benthic protists may be relatively moreimportant as metazoan diversity decreases, but com-parative studies are lacking.Higher (metazoan) trophic levels utilize autotro-
phic and heterotrophic protists in planktonic andbenthic environments. As noted earlier, autotrophicprotists have long been recognized as the base of mostaquatic food webs. Zooplankton and filter-feedingbenthic organisms capture phytoplankton, and brow-sers/scrapers such as snails and various aquatic insectsingest periphyton. Heterotrophic protists, which canbe as abundant as autotrophs and overlap in size, areingested incidentally by these ‘herbivorous’ metazo-ans that feed by either size- or nonselective methods.Some copepods feed selectively on ciliates and it islikely that there are other such discriminating preda-tors. In laboratory experiments, ciliates or HNANalone are usually a sufficient and complete diet formetazoan survival. However, ciliates and HNAN asa sole food source sometimes were insufficient forrobust reproduction suggesting nutritional deficiency.In natural systems, a mixed diet may overcome theselimitations on reproduction observed with food con-sisting of one or a few species.Predation by metazoans and other protists impacts
populations of heterotrophic protists. Ciliates can beimportant predators of HNAN and other ciliates inthe plankton and benthos. Most suctoria and heliozo-ans also feed on other protists. Predator removal(or addition) experiments in lakes demonstrated thatmoderate grazing by crustacean zooplankton canbalance or exceed protistan population growth, andthat selective feeding can alter protistan communitystructure. And though summer ‘clearwater’ phasesobserved in many lakes generally refer to a reductionin phytoplankton, predation by zooplankton alsosubstantially reduces heterotrophic protist popula-tions during these phases as well. In aquatic systems,it is likely that heterotrophic protists contribute to thediet of at least some life stage of most zooplankton,many fish, molluscs, and aquatic insects that also feedon photosynthetic protists.
Nutrient Cycling
The transfer of nutrients through different chemicalstates and ecological compartments (biotic and abi-otic) is tightly linked to microorganisms – especiallyin aquatic systems where protists dominate primaryproduction. Availability of dissolved phosphorus and/or nitrogen limits primary production, and recycling ismost important where allochthonous input of N andP is low. Heterotrophic protists can be efficient nutri-ent remineralizers, depending on the nutrient contentof their prey, and contribute to phytoplankton nutrientdemand in the epilimnion of stratified lakes whenmeasurable dissolved nutrients are low. Recyclingoften leads to high nutrient retention in benthic sys-tems within the photic zone of lakes and streams. Inthe benthos, phototrophic and heterotrophic micro-organisms, both prokaryotic and eukaryotic, tend tolive in close proximity and often within boundarylayers that can lead to localized nutrient limitationand a subsequent importance of recycling.
Summary
Protists represent a ubiquitous, though taxonomicallyill-defined, group of generally microscopic eukaryotesthat include amoeboid, flagellated, and ciliated taxa.They contribute substantially to primary production,food web interactions, and nutrient recycling in aquaticecosystems. All trophic levels, including primary pro-ducers, have protistan members. Heterotrophs feed asbacterivores, herbivores, carnivores, histophages, andparasites, or as omnivores. Several protistan groupshave members that combine photosynthesis andheterotrophy (mixotrophy). Protists are the major pre-dators of bacteria and are prey of benthic and plank-tonic metazoans. Consequently, dissolved organicmatter recovered by bacteria is coupled to metazoansby heterotrophic and mixotrophic protists, while pho-tosynthetic protists (phytoplankton and periphyton)are the base of aquatic metazoan food webs.
Glossary
Kleptoplasty – Retention of functional chloroplasts bya heterotrophic organism after ingestion of an auto-troph. Unlike symbiotic algae, these chloroplasts losefunctionality over time and must be replaced.
See also: Algae; Chrysophytes - Golden Algae; Diatoms;Green Algae; Harmful Algal Blooms; Microbial FoodWebs; Other Phytoflagellates and Groups of LesserImportance; Photosynthetic Periphyton and Surfaces;

260 Protists, Bacteria and Fungi: Planktonic and Attached _ Protists
Phytoplankton Nutrition and Related Mixotrophy;Phytoplankton Population Dynamics: Concepts andPerformance Measurement; Phytoplankton Productivity;Regulators of Biotic Processes in Stream and RiverEcosystems; Trophic Dynamics in Aquatic Ecosystems.
Further Reading
Adl SM, Simpson AGB, Farmer MA, et al. (2005) The new higher
level classification of eukaryotes with emphasis on the taxonomyof protests. Journal of Eukaryotic Microbiology 52: 399–541.
Foissner W, Berger H, and Schaumburg J (1999) Identification andEcology of Limnetic Plankton Ciliates. Munich: Bavarian State
Office for Water Management.Lee JJ, Leedale GF, and Bradbury P (eds.) (2000) Illustrated Guide
to the Protozoa 2nd edn., 2 vols. Lawrence: Society of Proto-
zoologists.Patterson DJ (2000) Free-living Freshwater Protozoa. Boca Raton,
FL: CRC Press.
Taylor WD and Sanders RW (2001) Protozoa. In: Thorp JH and
Covich AP (eds.) Ecology and Classification of North AmericanFreshwater Invertebrates, 2nd edn., pp. 43–96. San Diego:
Academic Press.
Wehr JD and Sheath RG (eds.) (2003) Freshwater Algae of NorthAmerica. San Diego: Academic Press.
Weisse T (2002) The significance of inter- and intraspecific vari-
ation in bacterivorous and herbivorous protists. Antonie vanLeeuwenhoek 81: 327–341.
Relevant Websites
http://megasun.bch.umontreal.ca/protists/protists.html – Protist
Image Data.http://www.microbeworld.org/microbes/protista –MicrobeWorld –
Protista.
http://www.uga.edu/~protozoa – International Society of Protistol-
ogists.







