Download - Effects of presurgical weight reduction on the development of hypothalamic hyperphagia in rats

BEHAVIORAL BIOLOGY 21,412--417 (1977)
BRIEF REPORT
Effects of Presurgical Weight Reduction on the Develop- ment of Hypothalamic Hyperphagia in Rats 1
A N T H O N Y S C L A F A N I
Department of Psychology, Brooklyn College of the City University of New York, Brooklyn, New York 11210
Ventromedial hypothalamic (VMH) knife cuts or sham cuts were made in female rats maintained either at ad libitum body weights or at 80% o fa d libitum weights. Following surgery, when food was freely available to all subjects, the VMH-cut rats overate relative to the controls, irrespective of whether their body weights were at normal or below-normal levels. In contrast to the hyperphagic effect of the VMH cuts, presurgical weight reduction did not result in elevated food intake, although it did increase postsurgical weight gain. These findings are discussed in light of recent motivational studies of the VMH hyperphagia syndrome.
It is well known that lesions or knife cuts in the ventromedial hypo- thalamus (VMH) produce overeating and obesity in animals fed a palat- able diet (Brobeck et al., 1943; Sclafani and Grossman, 1969). Recent studies have now demonstrated that VMH-damaged animals also display an enhanced motivation to eat palatable foods when tested under nonde- prived conditions (Beatty, 1973; Kent and Peters, 1973; Sclafani and Kluge, 1974; Sclafani, 1972; unpublished observations). However, when food deprived and maintained at 80 to 90% of preoperative body weights, VMH-damaged rats do not appear more motivated than do intact animals to obtain food (Kent and Peters, 1973; Porter and Allen, 1972; Sclafani, 1971; Sclafani and Kluge, 1974). These findings suggest that hypothalamic hyperphagia may be displayed only in rats at or above-normal ad libitum weight levels. Deprivation to below-normal body weights, by increasing the hunger drive of both intact and VMH-damaged rats, may temporarily prevent the VMH animals from overeating relative to controls. Some support for this hypothesis is provided by findings obtained with VMH
1 This research was supported by National Institute of Mental Health Grant MH 21563 and by the Research Foundation of the City University of New York Grant 10103.
412
Copyright @ 1977 by Academic Press, lnc All rights of reproduction Jn any form reserved ISSN 0091-6773

WEIGHT LOSS AND HYPOTHALAMIC HYPERPHAGIA 413
knife-cut and control rats switched from an unpalatable quinine diet to a palatable high-fat diet (Sclafani et al., 1976). The quinine-adulterated food suppressed the intake and body weight of both groups to below-normal levels, and, when offered the high-fat diet, the VMH and control rats initially overate to the same degree. However, as they gained weight, the control rats reduced their intake of the high-fat diet, while the VMH animals continued to overeat and became hyperphagic relative to the controls.
The purpose of the present experiment was to test more directly the hypothesis that body weight reduction to below-normal levels temporarily suppresses the appearance of hypothalamic hyperphagia. This was ac- complished by comparing the effects of VMH knife cuts and sham surgery on the food intake of rats maintained at either ad libitum or 80% of ad libitum body weights.
Method. Subjects. Thirty-three adult female rats of the CFN strain (Charles River, Mass.) were used as subjects. The rats were individually housed in wire-mesh cages in a colony room under a 12:12 light-dark cycle. Data from six rats are not included since they died shortly after surgery.
Procedure. All rats were initially given ad libitum access to Pu- rina Lab chow and tap water. The subjects were then divided into two groups equated for body weight, and 14 rats were given restricted amounts of food designed to maintain their body weights at approximately 80% ofad libitum levels, while the remaining 13 rats continued to receive food ad libitum. After 15 days on this schedule VMH knife cuts were made in eight of the deprived (VMH-DEP group) and seven of the non- deprived (VMH-AD LIB group) rats, while the remaining subjects were given sham knife cuts (CON-DEP, n = 6; CON-AD LIB, n = 6). Surgery was performed using the encephalotomy technique described by Sclafani (1971) and Equi-Thesin anesthesia (0.25 ml/0.1 kg body wt). The coordi- nates for the bilateral parasagittal knife cuts were AP: 8.0 to 5.0; H: +0.5 to -2.5; L = 0.8 (de Groot, 1967).
After surgery all rats were given 20_ + 1 g of food, which was equivalent to the ad libitum intake of the nondeprived subjects prior to surgery. The reason for the limited amount of food was to prevent rats operated on early in the day from eating more than rats operated on later in the day. During the next 13 days all rats were given ad libitum access to Purina chow. Food intake and body weight were recorded to the nearest gram throughout the experiment.
Histological verification of the knife cuts is not available since the subjects are currently being used in another experiment. This type of knife cut has been extensively performed in my laboratory, however, and histological description of such cuts can be found in previous reports (Sclafani and Grossman, 1969; Sclafani, 1971; Sclafani et al., 1976).
414 ANTHONY SCLAFANI
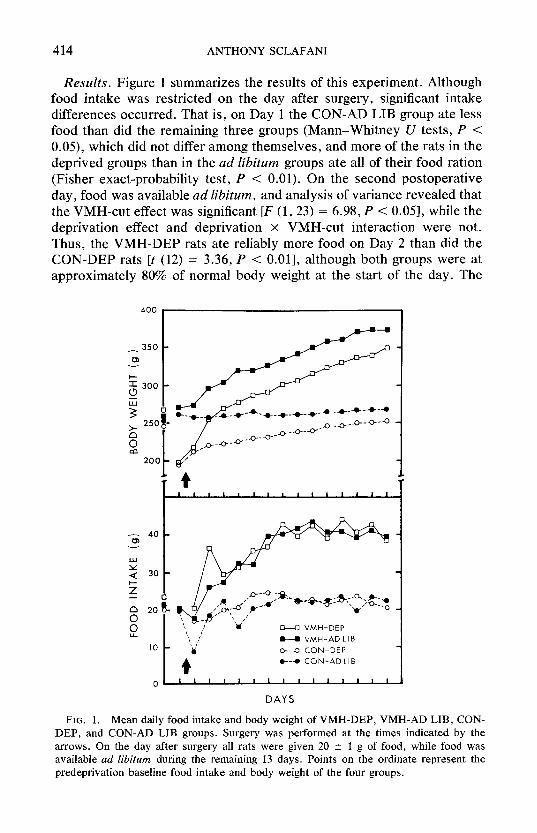
Results. Figure 1 summarizes the results of this experiment. Although food intake was restricted on the day after surgery, significant intake differences occurred. That is, on Day 1 the CON-AD LIB group ate less food than did the remaining three groups (Mann-Whi tney U tests, P < 0.05), which did not differ among themselves, and more of the rats in the deprived groups than in the ad libitum groups ate all of their food ration (Fisher exact-probabili ty test, P < 0.01). On the second postoperat ive day, food was available ad libitum, and analysis of variance revealed that the VMH-cut effect was significant IF (1, 23) = 6.98, P < 0.05], while the deprivation effect and deprivation x VMH-cut interaction were not. Thus, the VMH-DEP rats ate reliably more food on Day 2 than did the CON-DEP rats It (12) = 3.36, P < 0.01], although both groups were at approximately 80% of normal body weight at the start of the day. The
4 0 0
350
I,-- _~300
2OO
i _O. 41- -O- "O" "O" -O
f
v < 3o i-.-
Z ~ . ~._-o-2~.~ " ~.. ,~.~ .o..~. °
8 20 i ;~: /" O ", Z " "w' ~VMH-DEP " ~ V M H - A D L I B
10 ' " O----O C O N - D E P ~j~ 11--41 C O N - A D LIB
0 J I I I I I 1 I I I I I I I I
DAYS
FIG. 1. Mean daily food intake and body weight of VMH-DEP, VMH-AD LIB, CON- DEP, and CON-AD LIB groups. Surgery was performed at the times indicated by the arrows. On the day after surgery all rats were given 20 +- 1 g of food, while food was available ad libitum during the remaining 13 days. Points on the ordinate represent the predeprivation baseline food intake and body weight of the four groups.

WEIGHT LOSS AND HYPOTHALAMIC HYPERPHAGIA 415
VMH-DEP rats also consumed more food than did the VMH-AD LIB subjects, but this difference was not reliable.
During the remaining 12 days of the experiment the two VMH groups continued to overeat relative to the controls, and, within each surgical condition, there was no difference between the intakes of the deprived and ad libitum groups. Total food consumption over the 2-week post- operative period for the VMH-AD LIB, VMH-DEP, CON-AD LIB, and CON-DEP groups was 503, 519,290, and 302 g, respectively, and analysis of these data indicated that only the VMH-cut effect was significant IF (1, 23) = 65.28, P < 0.001].
As illustrated in Fig. 1, over the first 2 postoperative days, the VMH rats gained more weight than did the control rats [F (1, 23) = 35.0, P < 0.001], and within each surgical group the deprived rats gained more than did the ad lib rats [F (1, 23) = 34.7, P < 0.001]. Following this initial period, the VMH-cut rats continued to gain more weight than did the controls, with the rate of gain of the VMH-DEP group being slightly but not significantly greater than that of the VMH-AD LIB group (7.8 vs 6.5 g/day). The CON-DEP rats also gained at a faster rate than did the CON-AD LIB rats during Days 2-14, and in this case the difference was statistically significant [2.5 vs 0.9 g/day; t(10) = 4.89, P < 0.01]. The total weight gain during the 2 postoperative weeks for the VMH-AD LIB, VMH-DEP, CON-AD LIB, and CON-DEP groups was 103,155, 5, and 56 g, respectively, and analysis of variance indicated that both the VMH-cut effect IF (1, 23) = 131.9,P < 0.001] and the deprivation effect [F (1, 23) = 34.1, P < 0.001] were highly significant. By the end of the experiment the deprived rats still weighed slightly less than did the ad lib rats, but analysis of the Day 14 body weights revealed that only the VMH-cut effect was reliable [F (1, 23) = 55.5, P < 0.001].
Discussion. The results of this experiment fail to confirm the hypothesis that weight reduction to below-normal levels inhibits or delays the appearance of hypothalamic hyperphagia. The VMH-DEP group was hyperphagic relative to the CON-DEP group beginning the very first day that food was available ad libitum (Day 2), even though both groups were at 80% of normal body weight. In fact, rather than interfering with the development of hypothalamic hyperphagia, the effects of the presurgical weight loss on postoperative food intake and body weight gain were fully additive to the effects of the VMH-knife cuts (see below).
The results further revealed that food deprivation and weight loss, contrary to the effects of VMH cuts, do not result in significantly elevated food intakes, although they reliably increase body weight gain during refeeding. That is, over the 2-week postoperative period, the CON-DEP groups ate only 12 g more food, but gained 51 g more weight than did the CON-AD LIB group, and the VMH-DEP group ate only 16 g more food, but gained 48 g more weight than did the VMH-AD LIB group. These results are compatible with the recent report of Levitsky et al. (1976) that

416 ANTHONY SCLAFANI
rats deprived from 24 to 96 hr recover only about 1 day's intake of food when allowed to refeed, although their body weights return to normal ad libitum levels. Therefore, in both intact as well as VMH-damaged rats, increased food intake is only partially responsible for the recovery of weight following a fast. Apparently of greater importance for weight recovery are changes in food utilization and/or energy expenditure during refeeding (see Levitsky et al., 1976).
The finding that VMH-damaged rats deprived to a percentage of their preoperative weights overeat relative to similarily deprived controls when given food ad libitum, but do not bar press more for food than do controls during short-term tests (Kent and Peters, 1973; Porter and Allen, 1972; Sclafani, 1971: Sclafani and Kluge, 1974), indicates that short-term oper- ant tests of food motivation do not predict 24-hr food intake. This conclu- sion is further supported by the findings obtained with intact rats: Increas- ing the level of food deprivation in normal animals increases their operant responding for food (see Bolles, 1967), but does not appreciably increase their daily food intake during refeeding (Levitsky et al., 1976). Thus, it appears that short-term operant performance for food and 24-hr ad libitum intake are under the control of different neural mechanisms or processes (see Colier et al., 1977). Furthermore, damage to the ventromedial hypo- thalamus appears to disinhibit only the mechanism controlling ad libitum food intake, and this effect is independent of and additive to the effects produced by food deprivation and weight loss.
In contrast to the failure of presurgical weight reduction to prevent temporarily VMH-cut rats from overeating relative to controls, Sclafani et al. (1976) observed that VMH-cut and control rats are initially equally hyperphagic when switched from a bitter quinine diet to a palatable high-fat diet. Sclafani et aI. (1976) interpreted this effect as a result of the reduced body weights maintained by the rats on the quinine diet. In light of the present findings, however, it appears that the hyperphagia initially displayed by both the control and VMH-cut rats was more a response to the improved palatability of the diet than to their reduced body weight. In fact, it is known that intact rats will overeat and gain excessive weight when fed very palatable diets (Mallet, 1964: Sclafani and Springer, 1976: Sclafani and Gorman, 1977), and the dependence of hypothalamic hyper- phagia on diet palatability is well documented (Gale and Sclafani, 1977; Ferguson and Keesey, 1975; Sclafani et al., 1976). Therefore, it seems that the appetite aroused by good tasting foods may be a more potent and durable stimulus for overeating in intact and VMH-damaged rats than is the hunger produced by food deprivation and weight loss.
REFERENCES Beatty, W. W. (1973). Influence of type of reinforcement on operant responding by rats with
ventromedial lesions. Physiol. Behav. 10, 841-846. Bolles, R. C. (1967). "Theory of Motivation." New York: Harper & Row.

WEIGHT LOSS AND HYPOTHALAMIC HYPERPHAGIA 417
Brobeck, J. R., Tepperman, J., and Long, C. N. H. (1943). Experimental hypothalamic hyperphagia in the albino rat. Yale J. Biol. Med. 15, 831-853.
Collier, G., Hirsch, E., and Kanarek, R. (1977). The operant revisited. In W. K. Honig and J. E. R. Staddon (Eds.), "Handbook of Operant Behavior," pp. 28-52. Englewood Cliffs. N. J.: Prentice-Hall
De Groot, J. (1967). "The Rat Forebrain in Stereotaxic Coordinates." Amsterdam: N. V. Noord-Hollansche Uitgevers.
Ferguson, N. B. L., and Keesey, R. E. (1975). Effect of a quinine-adulterated diet upon body weight maintenance in male rats with ventromedial hypothalamic lesions. J. Comp. Physiol. Psychol. 95, 478-488.
Gale, S. K., and Sclafani, A. (1977). Comparison of ovarian and hypothalamic obesity syndromes in the female rat: Effects of diet palatability on food intake and body weight. J. Comp. Physiol. Psychol. 91, 381-392.
Kent, M. A., and Peters, R. H. (1973). Effects of ventromedial hypothalamic lesions on hunger-motivated behavior in rats. J. Comp. Physiol. Psychol. 83, 92-97.
Levitsky, D. A., Faust, I., and Glassman, M. (1976). The ingestion of food and the recovery of body weight following fasting in the naive rat. Physiol. Behav. 17, 575-580.
Mailer, O. (1964). The effect of hypothalamic and dietary obesity on taste preferences in rats. Life Sci. 3, 1281-1291.
Porter, J. H., and Allen, J. D. (1972). Food-motivated performance as a function of weight loss in hypothalamic hyperphagic rats. Psychon. Sci. 28, 285-288.
Sclafani, A. (1971). Neural pathways involved in the ventromedial hypothalamlc lesion syndrome in the rat. J. Comp. Physiol. Psychol. 77, 70-96.
Sclafani, A. (1972). The effects of food deprivation and palatability on the latency to eat in normal and hyperphagic rats. Physiol. Behav. 8, 977-979.
Sclafani, A. (1976). Appetite and hunger in experimental obesity syndromes. In D. Novin, W. Wyrwicka, and G. A. Bray (Eds.), "Hunger: Basic Mechanisms and Clinical Implications," pp. 281-295. New York: Raven Press.
Sclafani, A., and Gorman, A. N. (1977). Effects of age, sex and prior body weight on the development of dietary obesity in adult rats. Physiol. Behav., in press.
Sclafani, A., and Grossman, S. P. (1969). Hyperphagia produced by knife cuts between the medial and lateral hypothalamus in the rat. Physiol. Behav. 4, 533-537.
Sclafani, A., and Kluge, L. (1974). Food motivation and body weight levels in hypothalamic hyperphagic rats: A dual lipostat model of hunger and appetite. J. Comp. Physiol. Psychol. 86, 28-46.
Sclafani, A., and Springer, D. (1976). Dietary obesity in adult rats: Similarities to hypothalamic and human obesity syndromes. Physiol. Behav. 17, 461-471.
Sclafani, A., Springer, D., and Kluge, L. (1976). Effects of quinine adulteration on the food intake and body weight of obese and non-obese hypothalamic hyperphagic rats. Physiol. Behav. 16, 631-640.







