Download - An endornavirus from a hypovirulent strain of the violet root rot fungus, Helicobasidium mompa

Virus Research 118 (2006) 143–149
An endornavirus from a hypovirulent strain of theviolet root rot fungus, Helicobasidium mompa�
Hideki Osaki a,∗, Hitoshi Nakamura a, Atsuko Sasaki a,Naoyuki Matsumoto b, Koji Yoshida a
a National Institute of Fruit Tree Science, Fujimoto, Tsukuba 305-8605, Japanb National Institute for Agro-Environmental Sciences, Kan-non dai, Tsukuba 305-8604, Japan
Received 25 October 2005; received in revised form 8 December 2005; accepted 8 December 2005Available online 18 January 2006
Abstract
We determined the complete nucleotide (nt) sequence (16,614 nt) of a large double-stranded (ds) RNA (referred to as L1 dsRNA), previouslyidentified as the hypovirulence factor from strain V670 of the violet root rot fungus, Helicobasidium mompa. The positive-strand of L1 dsRNAcontained a long open reading frame (ORF) potentially encoding a protein of 5373 amino acids (molecular mass 603,080 Da) with conservedmTwhapan©
K
1
bbaedssip
n
R
0d
otifs characteristic of RNA-dependent RNA polymerase (RdRp) and helicase. The ORF is the longest so far reported in the fungal kingdom.he putative RdRp and helicase were shown to be related to putative RdRps and helicases of members of the genus Endornavirus. As is the caseith endornaviruses, the coding (sense) strand of L1 dsRNA contained a discontinuity (nick) at nt position 2552. A region between the RdRp andelicase domains of the polyprotein also had an amino acid sequence, resembling UDP glycosyltransferases (UGTs) in Oryza sativa endornavirusnd Phytophthora endornavirus 1. Regions in the L1 dsRNA-encoded protein presumed to contain putative helicase, UGT and RdRp motifs wereresent at comparable positions to those in other endornaviruses. L1 dsRNA of H. mompa strain V670 was assigned to the genus Endornavirus,nd here, we designate it as H. mompa endornavirus 1-670 (HmEV1-670). This represents the first report of a fungal endornavirus whose completeucleotide sequence has been determined.
2005 Elsevier B.V. All rights reserved.
eywords: Helicobasidium mompa; Endornavirus; Helicase; RNA-dependent RNA polymerase; dsRNA; Hypovirulence; UDP glycosyltransferase
. Introduction
The violet root rot fungus, Helicobasidium mompa Tanaka, aasidiomycete, occurs on various plants, including 104 specieselonging to 44 families (Ito, 1949). In spite of its serious dam-ge on agricultural crops, especially on fruit trees, there are fewffective control methods so far developed except biennial soilrench of chemicals. Biological control of violet root rot, usingoil microorganisms, is also inefficient, because the complexoil microflora is considered to prevent the establishment ofntroduced antagonists in the rhizosphere. Matsumoto (1998)roposed the use of double-stranded (ds) RNA or mycoviruses
� The nucleotide sequence data will appear in the DDBJ, EMBL and NCBIucleotide sequence databases under the accession number AB218287.∗ Corresponding author at: National Agricultural Research Center for Westernegion, 1-3-1 Sen-yu, Zentsuji, Kagawa 765-8508, Japan. Fax: +81 877 63 1683.
E-mail address: [email protected] (H. Osaki).
to control root diseases of fruit trees because the viruses existwithin the cytoplasm of the host fungus and is free from micro-bial interactions in soil.
dsRNA mycoviruses or dsRNA elements exist in all majorclasses of fungi (Buck, 1986; Nuss and Koltin, 1990) andare classified into four families Partitiviridae, Chrysoviri-dae, Totiviridae, and Hypoviridae. In addition, a novel genus,Mycoreovirus has recently been added to the family Reoviridaeby the ICTV (Mertens et al., 2005). Although a large number ofthe viruses that infect plant pathogenic fungi have been reportedto be avirulent, it is becoming increasingly clear that pheno-typic consequences of harboring specific mycoviruses or cer-tain dsRNA molecules can range from symptomless to severelydebilitating, or from hypovirulence to hypervirulence (Ghabrial,1994; Nuss and Koltin, 1990).
A large dsRNA (referred to as L1 dsRNA) in H. mompastrain V670 was identified as a hypovirulence factor (Ikeda etal., 2003). We subsequently analyzed L1 dsRNA to determine
168-1702/$ – see front matter © 2005 Elsevier B.V. All rights reserved.oi:10.1016/j.virusres.2005.12.004

144 H. Osaki et al. / Virus Research 118 (2006) 143–149
its complete RNA sequence and elucidated that L1 dsRNAbelonged to the genus Endornavirus which was proposed byGibbs et al. (2000) and newly accepted by the ICTV (Gibbset al., 2005). Fukuhara et al. (2003) also determined thepartial sequence of a dsRNA which was closely related toendornaviruses, from another strain of H. mompa. This is thefirst report to describe the complete nucleotide sequence of anendornavirus from a fungus.
2. Materials and methods
2.1. Extraction of dsRNA
A hypovirulent strain V670 of H. mompa from Corylusheterophylla in Japan (Ikeda et al., 2003) was maintained onpotato dextrose agar. For purification of dsRNAs, cultures weregrown on a cellulose membrane overlaid on potato dextroseagar plates at 25 ◦C. The cellulose membrane with mycelia wasthen stripped from the agar and ground to a fine powder in liq-uid nitrogen. dsRNA was extracted according to the method ofArakawa et al. (2002) and separated by electrophoresis on 1.0%agarose gel in TAE buffer (0.04 M Tris, 0.02 M acetic acid, 1 mMNa2EDTA, pH 7.4). Gels were stained with ethidium bromide.For cloning, L1 dsRNA was purified by excising bands from thegel and eluting dsRNA.
2
MpwFccT3DC
program DNASIS Version. 2.1 (Hitachi Software Engineering,Tokyo). Sequence comparison with those available fromnucleic acid and protein databases was performed using theBLAST program (Altschul et al., 1990). Multiple sequencealignments, construction of phylogenetic tree and bootstrappinganalysis were accomplished with the CLUSTAL W programs(Thompson et al., 1994). Neighbour-joining phylogenetictree was displayed using the program TREEVIEW (Page,1996).
3. Results
3.1. Nucleotide sequence of V670 L1 dsRNA and deducedamino acid sequence
The entire sequence of 16,614 nucleotides of L1 dsRNA wasassembled from overlapping clones (Fig. 1). One long openreading frame (ORF) was found in one strand. It has the poten-tial to encode a protein of 5373 amino acids (molecular mass603,080 Da). The first methionine codon at nt 11–13 was in afavourable context for translation initiation, with a purine (A orG) in position -3 and a G in position +4 (GCA G in cDNA ofL1 dsRNA), according to Kozak’s rules (Kozak, 1986; Lutckeet al., 1987). The 3′-untranslated region (UTR) was 485 bp longand lacked a poly(A) tail. The ORF of L1 dsRNA is apparentlytsdsmbt2a1saiT
F edicteV nes oe 73 aap
.2. Cloning and sequencing of dsRNA
cDNA clones were obtained following the method oforiyama et al. (1995). Briefly, random hexamers were used as
rimers for the synthesis of cDNA. Overlapping cDNA clonesere obtained using RT-PCR with sequence-specific primers.inally, the 5′- and 3′-RACE system for rapid amplification ofDNA ends (Invitrogen, Carlsbad) was used to obtain cDNAlones that corresponded to the terminal regions of the dsRNA.he discontinuity of coding strand was also analyzed by′-RACE. DNA sequence was determined using an automatedNA sequencer (PRISM 310NT, Applied Biosystems, Fosterity). Deduced amino acid sequence was predicted using the
ig. 1. Schematic representation of the strategy used in cDNA cloning and pr670. cDNA and RT-PCR clones are represented by filled boxes, 3′-RACE clo
xtremities by open boxes. The positive-strand features a long ORF encoding 53ositive-strand is interrupted by a discontinuity at nt 2552 (arrow).
he longest so far reported in the fungal kingdom. A BLASTearch using the complete amino acid sequence of the pre-icted L1 dsRNA-encoded protein showed significant sequenceimilarities to the predicted polyprotein sequences encoded byembers of the novel genus Endornavirus, recently accepted
y the ICTV (Gibbs et al., 2005). These included Phytoph-hora endornavirus 1 (PEV1, E value: 3e−77; Hacker et al.,005), Oryza sativa endornavirus (OsEV, 4e−72; Moriyama etl., 1995), and Vicia faba endornavirus (VfEV, 8e−69; Pfeiffer,998). Other short reading frames were also found in bothtrands. BLAST searches revealed that their deduced aminocids had no clear similarity to any proteins. These short read-ng frames are considered unlikely to be translated into proteins.he nucleotide sequence is deposited into the DDBJ, EMBL and
d genome organization of the L1 dsRNA from Helicobasidium mompa strainf the extremities are indicated by hatched shading, and 5′-RACE clones of thelong potential protein with a 10 nt long 5′-UTR and a 485 nt long 3′-UTR. The
H. Osaki et al. / Virus Research 118 (2006) 143–149 145
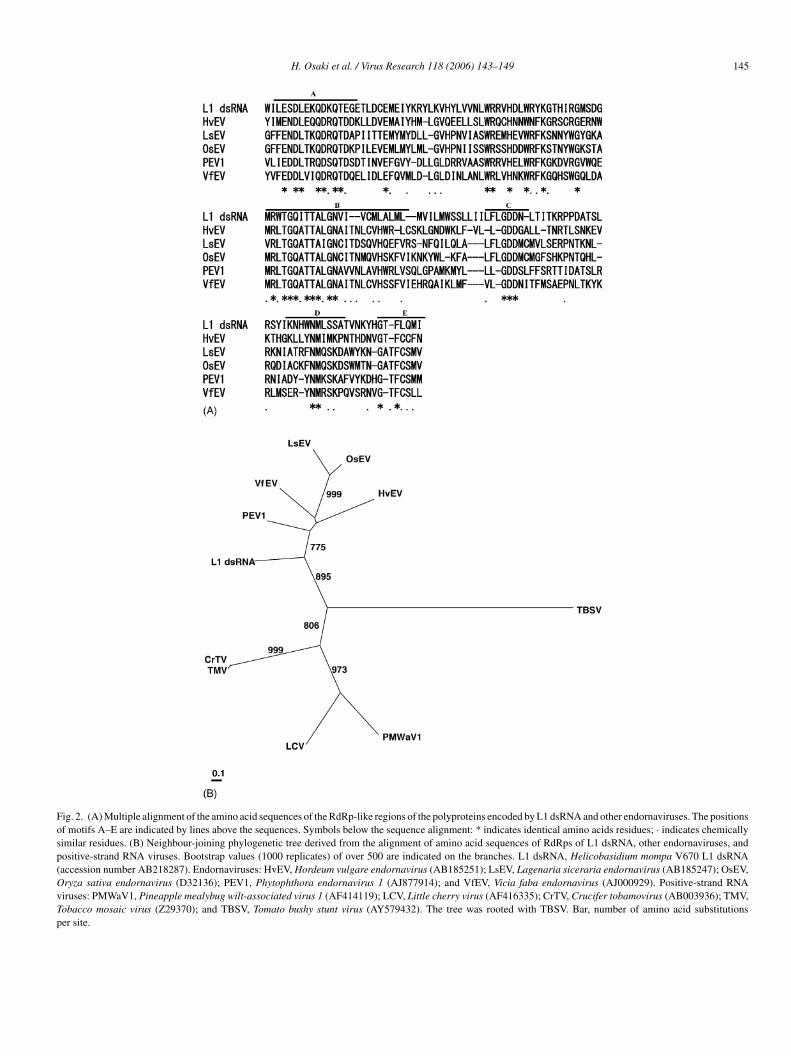
Fig. 2. (A) Multiple alignment of the amino acid sequences of the RdRp-like regions of the polyproteins encoded by L1 dsRNA and other endornaviruses. The positionsof motifs A–E are indicated by lines above the sequences. Symbols below the sequence alignment: * indicates identical amino acids residues; · indicates chemicallysimilar residues. (B) Neighbour-joining phylogenetic tree derived from the alignment of amino acid sequences of RdRps of L1 dsRNA, other endornaviruses, andpositive-strand RNA viruses. Bootstrap values (1000 replicates) of over 500 are indicated on the branches. L1 dsRNA, Helicobasidium mompa V670 L1 dsRNA(accession number AB218287). Endornaviruses: HvEV, Hordeum vulgare endornavirus (AB185251); LsEV, Lagenaria siceraria endornavirus (AB185247); OsEV,Oryza sativa endornavirus (D32136); PEV1, Phytophthora endornavirus 1 (AJ877914); and VfEV, Vicia faba endornavirus (AJ000929). Positive-strand RNAviruses: PMWaV1, Pineapple mealybug wilt-associated virus 1 (AF414119); LCV, Little cherry virus (AF416335); CrTV, Crucifer tobamovirus (AB003936); TMV,Tobacco mosaic virus (Z29370); and TBSV, Tomato bushy stunt virus (AY579432). The tree was rooted with TBSV. Bar, number of amino acid substitutionsper site.

146 H. Osaki et al. / Virus Research 118 (2006) 143–149
Fig. 3. Multiple alignment of amino acid sequences of the helicase-like region of the polyproteins encoded by L1 dsRNA and other endornaviruses. The positions ofmotifs I–VI are indicated by lines above the sequences. Symbols below the sequence alignment: * indicates identical amino acids residues; · indicates chemically similarresidues. L1 dsRNA, Helicobasidium mompa V670 L1 dsRNA (accession number AB218287); OsEV, Oryza sativa endornavirus (D32136); PEV1, Phytophthoraendornavirus 1 (AJ877914); PvEV, Phaseolus vulgaris endornavirus (AB185245); VfEV, Vicia faba endornavirus (AJ000929).
NCBI nucleotide sequence databases under the accession num-ber AB218287.
Endornaviruses are endogenous dsRNA elements withgenomes ranging between 14 and 18 kbp in size and containsingle ORFs spanning most of the length of the RNA (Gibbs etal., 2005). The size of V670 L1 dsRNA, the presence of a sin-gle, long ORF, and sequence similarity indicate that L1 dsRNAbelongs to the genus Endornavirus. Furthermore, a discontinuitypresent in the coding strand of all members of Endornavirus wasalso found to exist in L1 dsRNA at nt 2552, which was similarlypositioned to the discontinuity of VfEV (nt 2735) (Fig. 1).
3.2. Helicase-like, UGT-like, and RdRp-like regions
BLAST searches implied the presence of conserved motifscharacteristic of RNA-dependent RNA polymerase (RdRp) andhelicase in L1 dsRNA. They resembled those of members ofthe genus Endornavirus. Fig. 2A shows an alignment of aminoacid sequences within the conserved motifs A–E (Poch et al.,1989; Bruenn, 1993) of RdRps in six endornaviruses, includ-ing L1 dsRNA. The next most similar alignments were withregions of RdRps of the alpha-like virus superfamily of positive-
strand RNA viruses (Buck, 1996), particularly closterovirusesand tobamoviruses. It supports a suggestion that endornavirusesmight have evolved from an alpha-like virus that has lost itscapsid-coding region (Gibbs et al., 2000). A phylogenetic tree,derived from the alignment of amino acid sequences of theRdRps, is shown in Fig. 2B. L1 dsRNA RdRp obviously clusterswith other endornaviruses in RdRp. Fig. 3 shows an alignment ofamino acid sequences within the conserved motifs I–VI (Kooninand Dolja, 1993) of helicases with known endornaviruses.
The large proteins encoded by the endornaviruses have beenassumed to be polyproteins that are processed by virus-encodedproteinases. No proteinase motifs or proteolytic cleavage siteshave yet been reported for the endornaviruses. However, Hackeret al. (2005) identified cysteine-rich regions, which might playenzymic roles, in the polyproteins of VfEV, OsEV and PEV1.We also found no proteinase motifs, but a cysteine-rich regionthat contained 16 cysteine residues in a sequence of 72 residues(aa 1198–1269) in the L1 dsRNA-encoded large protein.
A BLAST search also revealed that a region between theRdRp and helicase domains of the L1 dsRNA polyprotein hadsignificant amino acid sequence similarity to UDP glycosyl-transferases (UGTs), as was reported in the case of OsEV and

H. Osaki et al. / Virus Research 118 (2006) 143–149 147
Fig. 4. Multiple alignment of amino acid sequences of the UGT motifs A and B of the polyproteins encoded by L1 dsRNA, other endornaviruses, and hypoviruses.Symbols below the sequence alignment: * indicates identical amino acids residues; · indicates chemically similar residues. The numbers of amino acid residuesbetween motifs A and B for each virus are indicated. L1 dsRNA, Helicobasidium mompa V670 L1 dsRNA (accession number AB218287); OsEV, Oryza sativaendornavirus (D32136); PEV1, Phytophthora endornavirus 1 (AJ877914); CHV3, Cryphonectria hypovirus 3 (AF188514) and CHV4, Cryphonectria hypovirus 4(AY307099).
Fig. 5. Comparison of genome organizations of Helicobasidium mompa endornavirus 1-670, Phytophthora endornavirus 1, Oryza sativa endornavirus, and Vicia fabaendornavirus. Long rectangular boxes represent the coding region for the polyproteins. Small boxes indicate the positions of the helicase-like (diagonally hatched),UGT-like (dotted) and RdRp-like (black) regions within the polyproteins. Arrowheads point to the discontinuity in the coding regions. Horizontal lines on either sideof the long rectangular boxes indicate the 5′- and 3′-UTRs.
PEV1. Fig. 4 shows the amino acid sequence alignment of puta-tive UGT motifs A and B (Hacker et al., 2005) encoded by L1dsRNA, other endornaviruses and hypoviruses, Cryphonectriahypoviruses 3 and 4 (CHV3 and 4; Linder-Basso, 2002). Theendornaviruses could not be distinguished from the Cryphonec-tria hypoviruses in UGT motifs except that the motifs were moreclosely positioned in the endornaviruses.
The regions presumed to encode helicase, UGT, and RdRpof the L1 dsRNA polyprotein, as well as the point of discon-tinuity, were all arranged in positions similar to those foundin other endornaviruses (Fig. 5). These observations stronglysupport that L1 dsRNA represents the genome of a member ofEndornavirus.
4. Discussion
Based on sequence analyses, homology searches, phyloge-netic analysis, and genomic organization, we propose that H.mompa V670 L1 dsRNA be assigned to the genus Endor-navirus and designate a new species, H. mompa endornavirus1-670 (HmEV1-670). Endornaviruses exist in plants, i.e., rice(Fukuhara et al., 1993), broad bean (Pfeiffer, 1998), and kidneybean (Wakarchuk and Hamilton, 1990). Recently an endor-navirus, Phytophthora endornavirus 1 (PEV1), has also been
identified in plant-parasitic Phytophthora species (Hacker et al.,2005), belonging to the Chromista kingdom (Cavalier-Smith,1997), which is separately treated from the kingdom of fungi(Hawksworth et al., 1995). Although partial sequence of dsRNAwhich was closely related to endornaviruses from another strainof H. mompa was determined (Fukuhara et al., 2003), this paperis the first report describing complete nucleotide sequence of anendornavirus from fungus. In Endornavirus, HmEV1-670 hasthe second-longest genome following VfEV (17635 bp, Fig. 5).The genome is also longer than those of hypoviruses (9–13 kbp),that is, the longest in the mycoviruses.
The amino acid sequence of the RdRp domain in HmEV1-670 was more closely related to that of the non-plant memberof the genus Endornavirus, PEV1 (43.3% identity), than thoseof endornaviruses from plants (36.2–40.7% identity). However,its helicase domain was more similar to those of endornavirusesfrom plants (25.6–31.1% identity) than that of PEV1 (25.5%identity). These findings suggest the genome heterogeneity ofendornaviruses, also supporting the hypothesis that RNA recom-bination had occurred possibly through horizontal transmissionbetween plants and parasitic organisms.
UGT genes have been known in several families of DNAviruses, and ecdysteroid UGTs are common in baculoviruses(Markine-Goriaynoff et al., 2004). Recently, they were also

148 H. Osaki et al. / Virus Research 118 (2006) 143–149
found in many RNA viruses, e.g. hypoviruses CHV3 and CHV4(Linder-Basso, 2002), and endornaviruses OsEV, and PEV1(Hacker et al., 2005). Viral UGT genes are considered to havebeen acquired from their hosts during the evolutionary processof the virus and the host (Markine-Goriaynoff et al., 2004).Also, plant endornaviruses are transmitted vertically throughseed and pollen, but not horizontally (Fukuhara et al., 1993).On the other hand, HmEV1-670 from a fungus was horizontallytransmitted through cytoplasmic mixing after hyphal anastomo-sis (Ikeda et al., 2003), representing the first evidence that anendornavirus was also horizontally transmitted. Plant-parasiticH. mompa infects a wide range of plants. These observationsled us to assume that fungi might have acquired their ancestralviruses from plants or vice versa, during infection.
The cellular sterol UGT gene is essential for pathogenesis inplant pathogenic fungi; a sterol UGT induced by hard surfacecontact of conidia was identified as a novel fungal virulencefactor in Colletotrichum gloeosporioides (Kim et al., 2002).It is also known that homology-dependent gene silencing ofa pathogenicity gene results in reduced virulence in Phytoph-thora infestans (Latijnhouwers et al., 2004). HmEV1-670 alsoreduces virulence of H. mompa (Ikeda et al., 2003). Gene silenc-ing induced by expression of HmEV1-670 UGT gene mightreduce virulence of H. mompa. However, the genetic backgroundof virulence is totally lacking in this fungus and should be elu-cidated in the future to clarify the mechanism of hypovirulence.
th(sieefe
A
Aa
R
A
A
B
B
B
C
Fukuhara, T., Moriyama, H., Pak, J.Y., Hyakutake, H., Nitta, T., 1993.Enigmatic double-stranded RNA in Japonica rice. Plant Mol. Biol. 21,1121–1130.
Fukuhara, T., Yamamoto, N., Yuki, C., Horiuchi, H., Miyazaki, S., Koga, R.,Moriyama, H., Takeshita, M., Ikeda, K., Arakawa, M., Matsumoto, N.,Arie, T., 2003. Endornavirus, the large dsRNA replicon with plasmid-likeproperties in plants and plant pathogenic fungi. In: Proceedings of theSeventh International Congress of Plant Molecular Biology, Barcelona,Spain, pp. S25–S27 (Abstracts).
Ghabrial, S.A., 1994. New developments in fungal virology. Adv. Virus Res.43, 303–388.
Gibbs, M.J., Koga, R., Moriyama, H., Pfeiffer, P., Fukuhara, T., 2000. Phy-logenetic analysis of some large double-stranded RNA replicons fromplants suggests they evolved from a defective single-stranded RNA virus.J. Gen. Virol. 81, 227–233.
Gibbs, M.J., Pfeiffer, P., Fukuhara, T., 2005. Genus endornavirus. In: Fau-quet, C.M., Mayo, M.A., Maniloff, J., Desselberger, U., Ball, L.A.(Eds.), Virus Taxonomy. Eighth Report of the International Committeefor the Taxonomy of Viruses. Academic Press, San Diego, pp. 603–605.
Hacker, C.V., Brasier, C.M., Buck, K.W., 2005. A double-stranded RNA froma Phytophthora species is related to the plant endornaviruses and containsa putative UDP glycosyltransferase gene. J. Gen. Virol. 86, 1561–1570.
Hawksworth, D.L., Kirk, P.M., Sutton, B.C., Pegler, D.N., 1995. Dictionaryof the Fungi. CAB International, Wallingford, pp. 227–228.
Ikeda, K., Nakamura, H., Matsumoto, N., 2003. Hypovirulent strain of theviolet root rot fungus Helicobasidium mompa. J. Gen. Plant Pathol. 69,385–390.
Ito, K., 1949. Studies on ‘Murasaki-mompa’ disease caused by Helicobasid-ium mompa Tanaka. Bull. Govern. For. Exp. Station 43, 1–126.
Kim, Y.-K., Wang, Y., Liu, Z.M., Kolattukudy, P.E., 2002. Identification of
K
K
L
L
L
M
M
M
M
N
P
P
Most endornaviruses have no obvious effect on the pheno-ype of their hosts. However, HmEV1-670 was found as theypovirulence factor, reducing virulence of the host, H. mompaIkeda et al., 2003). VfEV is associated with cytoplasmic maleterility in V. faba. The sequence information is extremely lim-ted in endornaviruses. Additional reports on the occurrence ofndornaviruses and on their sequence information should revealvolutionary interactions with their hosts. As to endornavirusesrom fungi, when novel viruses are found, they should also bevaluated as biocontrol agents.
cknowledgement
This study was funded by the Research Project for Utilizingdvanced Technologies in the Ministry of Agriculture, Forestry
nd Fisheries.
eferences
ltschul, S.F., Gish, W., Miller, W., Myers, E.W., Lipman, D.J., 1990. Basiclocal alignment search tool. J. Mol. Biol. 215, 403–410.
rakawa, M., Nakamura, H., Uetake, Y., Matsumoto, N., 2002. Presence anddistribution of double-stranded RNA elements in the white root rot fungusRosellinia necatrix. Mycoscience 43, 21–26.
ruenn, J.A., 1993. A closely related group of RNA-dependent RNApolymerases from double-stranded RNA viruses. Nucl. Acids Res. 21,5667–5669.
uck, K.W., 1986. Fungal virology—an overview. In: Buck, K.W. (Ed.),Fungal Virology. CRC Press, Boca Raton, pp. 1–84.
uck, K.W., 1996. Comparison of the replication of positive-stranded RNAviruses of plants and animals. Adv. Virus Res. 47, 159–251.
avalier-Smith, T., 1997. Sagenista and Bigyra, two phyla of heterotrophicheterokont Chromists. Arch Protistenk 148, 253–267.
a hard surface contact-induced gene in Colletotrichum gloeosporioidesconidia as a sterol glycosyltranserase, a novel fungal virulence factor.Plant J. 30, 177–187.
oonin, E.V., Dolja, V.V., 1993. Evolution and taxonomy of positive-strand RNA viruses: implications of comparative analysis of amino acidsequences. Crit. Rev. Biochem. Mol. Biol. 28, 375–430.
ozak, M., 1986. Point mutations define a sequence flanking the AUG ini-tiator codon that modulates translation by eukaryotic ribosomes. Cell 44,283–292.
atijnhouwers, M., Ligterink, W., Vleeshouwers, V.G.A.A., van West, P.,Govers, F., 2004. A G� subunit controls zoospore motility and virulencein the potato late blight pathogen Phytophthora infestans. Mol. Microbiol.51, 925–936.
inder-Basso, D., 2002. Viruses and transposons of the chestnut blight fungus,Cryphonectria parasitica: tools to examine virulence, fungal diversity, andevolution. PhD Dissertation. Rutgers University, New Brunswick, NJ.
utcke, H.A., Chow, K.C., Mickel, F.S., Moss, K.A., Kern, H.F., Scheele,G.A., 1987. Selection of AUG initiation codons differs in plants andanimals. EMBO J. 6, 43–48.
arkine-Goriaynoff, N., Gillet, L., van Etten, J.L., Korres, H., Verma, N.,Vanderplasschen, A., 2004. Glycosyltranserases encoded by viruses. J.Gen. Virol. 85, 2741–2754.
atsumoto, N., 1998. Biological control of root diseases with dsRNA basedon population structure of pathogens. JARQ 32, 31–35.
ertens, P.P.C., Hillman, B.I., Suzuki, N., 2005. Genus Mycoreovirus. In:Fauquet, C.M., Mayo, M.A., Maniloff, J., Desselberger, U., Ball, L.A.(Eds.), Virus Taxonomy. Eighth Report of the International Committeefor the Taxonomy of Viruses. Academic Press, San Diego, pp. 556–560.
oriyama, H., Nitta, T., Fukuhara, T., 1995. Double-stranded RNA in rice:a novel RNA replicon in plants. Mol. Gen. Genet. 248, 364–369.
uss, D.L., Koltin, Y., 1990. Significance of dsRNA genetic elements in plantpathogenic fungi. Annu. Rev. Phytopathol. 28, 37–58.
age, R.D.M., 1996. TreeView: an application to display phylogenetic treeson personal computers. Comput. Appl. Biosci. 12, 357–358.
feiffer, P., 1998. Nucleotide sequence, genetic organization and expressionstrategy of the double-stranded RNA associated with the‘447’ cytoplasmicmale sterility trait in Vicia faba. J. Gen. Virol. 79, 2349–2358.

H. Osaki et al. / Virus Research 118 (2006) 143–149 149
Poch, O., Sauvaget, I., Delarue, M., Tordo, N., 1989. Identification of fourconserved motifs among the RNA-dependent polymerase encoding ele-ments. EMBO J. 8, 3867–3874.
Thompson, J.D., Higgins, D.G., Gibson, T.J., 1994. Clustal W. Improving thesensitivity of progressive multiple sequence alignments through sequence
weighting, position-specific gap penalties and weight matrix choice. Nucl.Acids Res. 22, 4673–4680.
Wakarchuk, D.A., Hamilton, R.I., 1990. Partial nucleotide sequence fromenigmatic dsRNAs in Phaseolus vulgaris. Plant Mol. Biol. 14, 637–639.







