does soil nutrient availability influence night-time water flux of aspen saplings?
TRANSCRIPT

Do
PHa
b
a
ARRA
KFGNNNS
1
b(Idwupn(te
0h
Environmental and Experimental Botany 82 (2012) 37– 42
Contents lists available at SciVerse ScienceDirect
Environmental and Experimental Botany
journa l h omepa g e: www.elsev ier .com/ locate /envexpbot
oes soil nutrient availability influence night-time water fluxf aspen saplings?
riit Kuppera,∗, Gristin Rohulaa, Liina Saksinga, Arne Sellina, Krista Lõhmusa, Ivika Ostonena,eljä-Sisko Helmisaarib, Anu Sõbera
Institute of Ecology and Earth Sciences, University of Tartu, Lai 40, 51005 Tartu, EstoniaDepartment of Forest Sciences, University of Helsinki, FI-00014 Helsinki, Finland
r t i c l e i n f o
rticle history:eceived 8 June 2011eceived in revised form 22 March 2012ccepted 30 March 2012
eywords:ine-root adaptationrowth chamber experimentight-time water relationsitrogenutrient deficiencyap flux density
a b s t r a c t
Previous investigations have been demonstrated that night-time water flux may increase or remainunchanged at low mineral nutrient availability. At the same time, it is a well-known fact that night-time water flux is relatively high in fast-growing species, which typically grow in fertile soil. To testthe impact of soil nutrient deficiency on night-time water flux and fine-root acclimation in saplings ofhybrid aspen (Populus tremula L. × Populus tremuloides Michx.), a growth chamber experiment was car-ried out. We set up a hypothesis that night-time sap flux density (F) and night-time water use percentagefrom daytime water use (NWU) are more intense in fertile conditions, in order to enhance or sustain thehigh intrinsic growth potential of hybrid aspen. The main limiting element in the low nutrient availabil-ity treatment (low-n) was nitrogen. The nitrogen concentrations of leaves and fine-roots exhibited thestrongest (R2 = 0.95; P < 0.001) positive relationship with NWU and foliar biomass. Both the night-time Fand NWU were several times higher in the case of fertilized soil (high-n treatment) compared to low-ntreatment (P < 0.01). The differences in nocturnal (and in diurnal) F disappeared at the end of the periodof sap flow measurements, when the foliage area of trees was almost full-grown. Endogenous increase
in water flux during predawn hours was observable only in the high-n treatment. Significantly greaterNWU (P < 0.01) and specific fine-root length (P < 0.05), but smaller fine-root biomass (P < 0.05) in saplingsof the high-n treatment potentially allow plants to use mass flow in soil more efficiently for transporta-tion nutrients towards roots and to decrease construction costs for fine-root biomass production. Ourresults suggest that decreased night-time water flux as a result of strong nutrient (especially nitrogen)deficit could be characteristic to fast-growing tree species, which are adapted to grow in fertile soil.. Introduction
In the last decade, the night-time transpiration phenomenon haseen intensively studied as a potential driver of mineral nutrientsespecially nitrogen) from the soil to tree roots and up to the foliage.t has been suggested that increased water flux reduces the nutrientepletion zone of mass flow mobile nutrients like that may other-ise develop in the rhizosphere as a consequence of active nutrientptake. The mass flow of nutrients may be especially important forlants with low root densities, providing a mechanism for accessingutrients in the absence of an extensively proliferated root systemCramer et al., 2008). The reduction of xylem flow is also known
o repress xylem loading of different nitrogen compounds (Delhont al., 1995) and to inhibit the uptake of nitrate and ammonium∗ Corresponding author. Tel.: +372 7 376 219; fax: +372 7 376 222.E-mail address: [email protected] (P. Kupper).
098-8472/$ – see front matter © 2012 Elsevier B.V. All rights reserved.ttp://dx.doi.org/10.1016/j.envexpbot.2012.03.013
© 2012 Elsevier B.V. All rights reserved.
in fine roots of different tree species at night (Gessler et al., 1998;Geßler et al., 2002).
However, the causality between nocturnal transpiration andmineral nutrition has remained rather unclear and has an ambiva-lent nature (Daley and Phillips, 2006; Caird et al., 2007; Dawsonet al., 2007; Howard and Donovan, 2007, 2010; Marks andLechowicz, 2007; Scholz et al., 2007; Christman et al., 2009). Thereis evidence that night-time transpiration increases at low min-eral nutrient (N, P) availability (Scholz et al., 2007) or remainsunchanged (Howard and Donovan, 2007, 2010; Christman et al.,2009). At the same time it is a well-recognized fact that night-timetranspiration is characteristic to shade-intolerant species, whichdo have relatively high overall growth rate and leaf nitrogen con-centration (Daley and Phillips, 2006; Marks and Lechowicz, 2007).Phillips et al. (2010) demonstrated that nocturnal water transport
was greater in Eucalyptus trees with a higher proportion of youngfoliage. This suggests that night-time transpiration could be ben-eficial in periods of high nitrogen demand. Hence, recent findingsseem to support the idea that night-time transpiration should be
3 nd Experimental Botany 82 (2012) 37– 42
hdtctsebeae
nuc((o
2
2
bbps2t2ostww
eameswtaigoAtnwN0g(mC
etwvta
Fig. 1. Average daily course of the stem sap flux density (F) in high nutrient avail-ability treatment (high-n) and low nutrient availability treatment (low-n) and airvapour pressure deficit (VPD) in growth chamber during period of sap flow mea-
8 P. Kupper et al. / Environmental a
igher in trees growing in fertile soils than in the case of nutrienteficiency, because high growth rate and leaf nitrogen concentra-ion will more likely be achieved in fertilized (and well watered)onditions. At the same time, one cannot exclude the possibilityhat trees growing in nutrient deficiency will transpire more inten-ively to acquire additional N and P at night, as suggested by Scholzt al. (2007). Caird et al. (2007) pointed out that the relationshipetween varying night-time leaf conductance responses to nutri-nts and particular life forms or ecological strategies is unknownnd may be related to the underlying causes of nutrient statusffects on day-time leaf conductance.
In the current study, impact of mineral nutrient availability onight-time water flux was analysed on hybrid aspen (Populus trem-la L. × Populus tremuloides Michx.) saplings grown in a climatehamber. We set up a hypothesis that night-time sap flux densityF) and night-time water use percentage from daytime water useNWU) are more intense in fertile conditions, in order to enhancer sustain the high intrinsic growth potential of hybrid aspen.
. Materials and methods
.1. Sample trees and experimental conditions
The experiment was carried out in a double-sided growth cham-er Percival AR-95 HIL (Percival Scientific Inc., Perry, IA) from theeginning of April to the end of July 2009. Eight 3-year-old micro-ropagated hybrid aspen (P. tremula L. × P. tremuloides Michx.)aplings were dug out from Rõka experimental site (Kupper et al.,011), Järvselja Experimental Forest District (58◦24′N, 27◦29′E, alti-ude 40–48 m) in south-eastern Estonia before bud break in April009. The average diameter (at the base of the stem) and heightf the sample trees was 10.2 mm and 1.06 m, respectively. The rootystem of each transplant was washed carefully to remove soil par-icles and the saplings were planted into the 10-l pots. The potsere without drainage holes in order to prevent the leakage ofater and the leaching out of mineral nutrients.
The amount of growth substrate was different for the two nutri-nt availability treatments, being 9 and 7 l for the high nutrientvailability (high-n) and the low nutrient availability (low-n) treat-ents, respectively. The difference in soil volume enabled us to
qualize the plant size/soil volume ratio and also ensured similaroil moisture conditions in both treatments. The pots were filledith a growth substrate of calcimined sphagnum peat with a par-
icle size of up to 20 mm. The organic content of the peat was 98%nd the degree of decomposition was between 2 and 4 accord-ng to the Von Post classification. Each pot contained up to 3%ravel with a particle size of 2–5 mm. The initial concentrationf macro- and micronutrients in the growth substrate (Appendix) was about two times higher in the high-n treatment than in
he low-n treatment, whereas the dominating N source was NO3−
itrogen. In addition, the growth substrate of the high-n treatmentas enriched with a water dissolved fertilizer (containing 6.6%O3
−, 2.4% NH4+, 9% P2O5, 27% K2O, 3% MgO, 0.01% B, 0.01% Cu,
.06% Fe, 0.04% Mn, 0.001% Mo and 0.01% Zn) nine times during arowth period. Each tree was fertilized with a 300 ml of solution0.1%). The average soil water pHH2O measured with a digital pH
eter (Model 60; Lazar Research Laboratories, Inc., Los Angeles,A) was 5.8 and 4.9 for high-n and low-n treatments, respectively.
To water the trees, the pots with saplings were weighed, usuallyvery evening and the amount of daily consumed water was addedo the pots to restore the soil water reserve. The threshold of soil
ater content was set to 60% of the field capacity. The 20% larger soilolume in the pots of the fertilized trees enabled 20% more watero be added to the pots at the same soil water content threshold,nd a presumable decline in soil matric potential (� S) as a result of
surements. Both lines of the F represent the average value of four measured trees.The line of the VPD represents the average value in the growth chamber. The blackstrip expresses night-time.
greater water consumption due to the larger size of fertilized treesto be partially avoided. The average � S did not fall below −25 kPaduring the whole study period. During sap flow measurements thedaily average � S was −11.5 and −12.9 kPa in high-n and low-ntreatments, respectively.
The night/day length in the chamber was 8.5/15.5 h, and the lightwas switched off from 23:00 to 7:30 h. The daily photosyntheticallyactive radiation (PAR) was kept constant at 400–800 �mol m−2 s−1
for the upper foliage of the saplings. Both the relative humidity(RH) and temperature of the air (Ta) varied between the oppositesides of the growth chamber. The average RH and Ta were 71.3%and 20.6 ◦C in left side of the chamber but 65.1% and 22.4 ◦C in theright side of the chamber. However, the trees from the differenttreatments were distributed equally between the two sides of thechamber (four sample trees on both sides: two from high-n and twofrom low-n treatment) and therefore the two nutrient availabil-ity treatments are comparable to each other. The average RH waskept ca 5% lower at night than in the daytime, in order to inducenocturnal transpiration flux in sample trees (Fig. 1). The air tem-perature of the chamber was kept constant during the night andday. Vapour pressure deficit of the atmosphere (VPD) was calcu-lated as the difference between the saturation and ambient vapourpressures using RH and Ta data. RH and Ta were measured with fourHMP45A sensors (Vaisala, Helsinki, Finland) located at two heights(70 and 120 cm) on the two sides of the growth chamber. The read-ings of the HMP45A probes were recorded with a DL2e data logger(Delta-T Devices, Burwell, UK). � S was measured with dielectricwater potential sensors (MPS-1; Decagon Devices, Pullman, WA),one per each pot, located at a depth of 10–15 cm and connected tothe Em50 data loggers (Decagon Devices). The readings of all sen-sors were collected every 1 min and stored as average values every10 min with the data loggers.
2.2. Measurements of sap flow and leaf area
The xylem sap flow of saplings (N = 8) was recorded with asap flow system T4.2 (EMS Brno, Brno, Czech Republic), furnishedwith ‘Baby Kucera’ sap flow sensors for 8–12 and 12–18 mm stem
diameter. Each sample tree was fitted with one sensor installedca 40–50 cm above the ground. The sap flow data was recordedevery 1 min and stored as 10-min averages from 37 to 57 daysafter bud break in May and June. Nevertheless, only the 11 days
nd Exp
(cailwiwummto
moalfo
A
ddlt(
(uctiatfli
(flNt
2
lm
2
uTw(lpI
2
lt
P. Kupper et al. / Environmental a
38–42, 44, 47–48, 54–56 after bud break) without any notablehanges in chamber and soil parameters were included in the datanalysis to estimate stem sap flow. The average values recordedn humid nights (RH > 90%; VPD < 0.3 kPa) were used in the base-ine subtraction procedure. Humid nights were induced during the
hole period of sap flow measurements to exclude the impact ofncreased stem heat storage on the baseline values. In addition,
e weighed the tree containers daily with an electronic scale andsed the weight data to correct the sap flow values over the wholeeasurement period. Hence, a regression between simultaneouslyeasured sap flow and weighed water loss of trees was derived, and
he regression equation was applied to calculate calibrated valuesf the sap flow.
To express the sap flux density per unit projected leaf area (F;mol m−2 s−1), the tree total leaf area was determined on the basis
f intact leaves every 1–3 weeks from April to June. To estimate therea of intact leaves, the length (l) and width (W) of every singleeaf were measured with a ruler; thereafter the leaf area (AL) wasound using an empirical relationship between AL and the productf l and W, derived at the end of the study period in July:
L = 0.539+0.659 × l × W; R2=0.991, P < 0.001, N=50. (1)
To calculate the leaf area values for the sap flow measurementays, a regression analysis was applied on determined the leaf areaataset. Thereafter, regression equations were used to calculate the
eaf area for all sap flow measurement days. The total leaf area ofhe saplings was determined with a LI-3100C optical area meterLI-COR Biosciences, Lincoln, NE) at the end of July.
We used only the averages of daytime (15.5 h) and night-time8.5 h) sap flux density (F) for statistical data analysis. This enableds to reduce the possible impact of stem water capacity on theanopy transpiration estimate. Moreover, we did not detect anyime lag between time series of F and VPD (estimated at 10-minntervals from 10:00 to 13:00 h on 38–40 days after bud break),nd the correlation between F and current VPD was always closerhan that of any shifted time series. Therefore we assume that sapux density is an adequate estimate of the whole-tree transpiration
n the case of our study.Night-time water use percentage from daytime water use
NWU) was calculated as the proportion of mean night-time sapux density compared to mean daytime sap flux density (%). TheWU is a species-specific measure of plant water use, which allows
he estimation of plants’ relative ability to transpire at night.
.3. Biomass analysis
Total tree biomass and biomasses of separate fractions (roots,eaves) were estimated at the end of the study period. The plant
aterial was dried at 70 ◦C for 48 h and weighed.
.4. Fine-root analysis
The root systems of saplings were washed free of soil man-ally with tap water and separated into living and dead roots.he proportion of dead roots was negligible, and hence dead rootsere omitted from further analysis. Four sub-samples of fine roots
d < 1 mm) per tree (N = 8) were taken to estimate specific rootength (SRL; m g−1). For the measurement of SRL and whole-treerojected fine-root area, WinRHIZO Pro 2003b (Regent Instruments
nc., Québec, QC) was used.
.5. Chemical analysis of roots and leaves
In order to evaluate the nutrient status of the trees, roots andeaves were analysed for macro- and micronutrients. The concen-ration of total N in leaves was determined by standard Kjeldahl
erimental Botany 82 (2012) 37– 42 39
procedure using a Kjeltec Auto 1030 Analyser (FOSS Trecator AB,Höganäs, Sweden). The total N concentrations of the fine-root sam-ples were determined with a LECO CHLOW-A1000 analyser (LECOCorporation, St. Joseph, MI). To determine total concentrations ofother elements (K, Ca, Mg, P, S, Fe, Mn, Zn, Cu), fine roots andleaves were extracted by heating (target temperature 175 ◦C) thedried and milled samples in HNO3 (+H2O2) in a microwave oven(Mars 5), based on EPA 3051. After extraction the samples werecooled, filtered and diluted, and the concentrations measured byan inductively coupled plasma optical emission spectrometer ICP-OES (Thermo Scientific, ICAP 6000). Samples for chemical analysiswere taken from dried biomass of fine roots (N = 8) and leaves (N = 8for nitrogen). The concentration of other elements in leaves wasestimated only in 4 saplings.
2.6. Statistical methods
For comparison of the dynamics of sap flux density, night-timewater use percentage, and canopy leaf area within the study period(days: 38–42, 44, 47–48, 54–56 after bud break), a repeated mea-sures analysis of variance (ANOVA) was applied on the datasets.A Student’s t-test for dependent samples was used in the case ofrepeated sap flux density measurements to estimate changes insap flux density at night.
A Student’s t-test for independent samples was applied todifferent measured parameters (Nleaf, Nroot, Proot, Caroot, Mgroot,Sroot, Curoot, Feroot, Znroot, Mnroot, F, SRL, total tree biomass,total foliar biomass, total leaf area, total fine-root area) to esti-mate the differences between high-n and low-n treatments.The assumptions of normality and homogeneity of varianceswas checked using the Kolmogorov–Smirnov D-statistic and theLevene test, respectively. If the conditions of normality andhomogeneity of variables were not met, a nonparametric two-sided Mann–Whitney U-test was used (Kroot, total fine-rootbiomass).
A forward stepwise regression analysis was used to differenti-ate the impact of mineral nutrients (Nleaf, Nroot, Caroot, Proot, Sroot,)and pH on NWU. A linear regression analysis was carried out to esti-mate relationships between dependent (NWU, total foliar biomass)and predictor variables (Nleaf, Nroot, Proot). The normality of theregression residuals was checked using the Kolmogorov–SmirnovD-statistic. All data analyses were carried out using the softwareStatistica, Version 7.0 (StatSoft Inc., Tulsa, OK).
3. Results
Although the daytime stem sap flux density was not statistically(P > 0.05) higher in saplings grown at greater soil nutrient availabil-ity, the night-time F was significantly different (P < 0.01) betweenthe treatments during the observed time period (Fig. 2). Both day-and night-time F were statistically different (P < 0.05) for the dif-ferent treatments at the beginning of the measurement period (onthe 38th day after bud break). In contrast, neither daytime nor noc-turnal sap flux density differed (P > 0.05) between the treatmentsat the end of sap flow measurement period (on the 56th day afterbud break).
The seasonal courses of diurnal and nocturnal F followedchanges in the leaf area of saplings (Fig. 3). As leaf area did not vary(P > 0.05) between the treatments at the beginning of the measure-ment period (from 38 to 42 days after bud break), the difference inF was not directly caused by the difference in leaf area, but rather
by the difference in sap flow per se. The increase in leaf area (about2.5 times) was responsible for the proportional decrease in sap fluxdensity per unit projected leaf area (F) in fertilized trees during sapflow measurements (Fig. 3).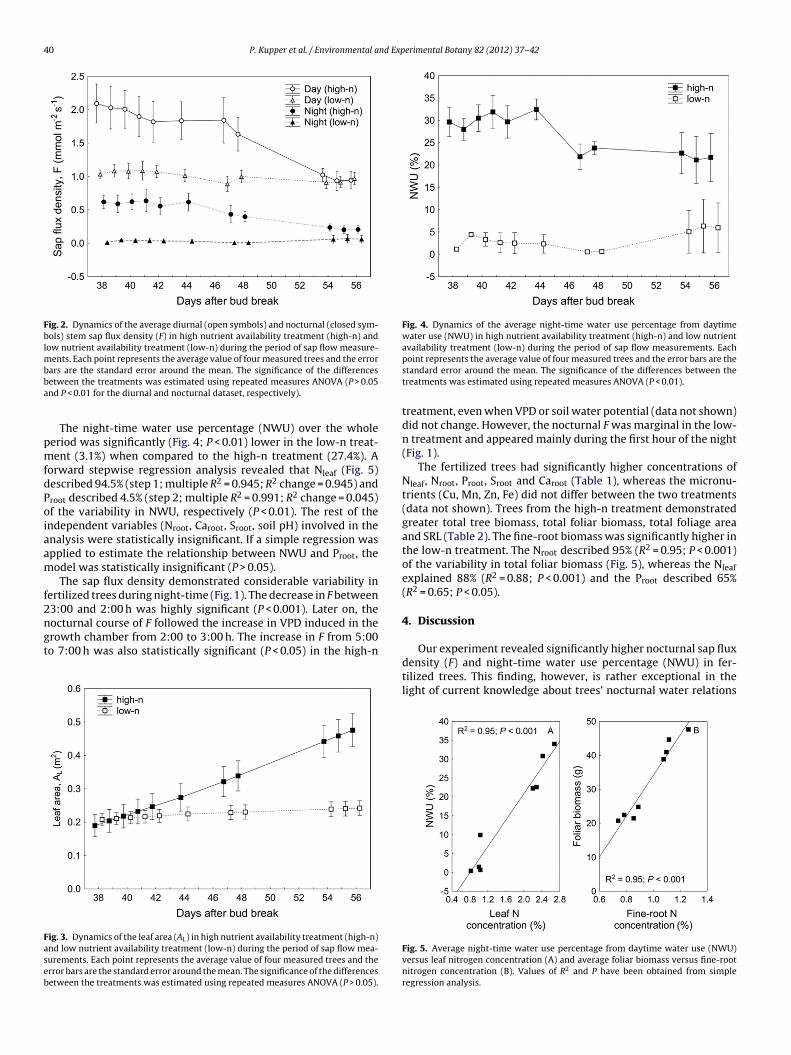
40 P. Kupper et al. / Environmental and Experimental Botany 82 (2012) 37– 42
Fig. 2. Dynamics of the average diurnal (open symbols) and nocturnal (closed sym-bols) stem sap flux density (F) in high nutrient availability treatment (high-n) andlow nutrient availability treatment (low-n) during the period of sap flow measure-ments. Each point represents the average value of four measured trees and the errorbba
pmfdPoiaam
f2ngt
Faseb
Fig. 4. Dynamics of the average night-time water use percentage from daytimewater use (NWU) in high nutrient availability treatment (high-n) and low nutrientavailability treatment (low-n) during the period of sap flow measurements. Each
ars are the standard error around the mean. The significance of the differencesetween the treatments was estimated using repeated measures ANOVA (P > 0.05nd P < 0.01 for the diurnal and nocturnal dataset, respectively).
The night-time water use percentage (NWU) over the wholeeriod was significantly (Fig. 4; P < 0.01) lower in the low-n treat-ent (3.1%) when compared to the high-n treatment (27.4%). A
orward stepwise regression analysis revealed that Nleaf (Fig. 5)escribed 94.5% (step 1; multiple R2 = 0.945; R2 change = 0.945) androot described 4.5% (step 2; multiple R2 = 0.991; R2 change = 0.045)f the variability in NWU, respectively (P < 0.01). The rest of thendependent variables (Nroot, Caroot, Sroot, soil pH) involved in thenalysis were statistically insignificant. If a simple regression waspplied to estimate the relationship between NWU and Proot, theodel was statistically insignificant (P > 0.05).The sap flux density demonstrated considerable variability in
ertilized trees during night-time (Fig. 1). The decrease in F between3:00 and 2:00 h was highly significant (P < 0.001). Later on, the
octurnal course of F followed the increase in VPD induced in therowth chamber from 2:00 to 3:00 h. The increase in F from 5:00o 7:00 h was also statistically significant (P < 0.05) in the high-nig. 3. Dynamics of the leaf area (AL) in high nutrient availability treatment (high-n)nd low nutrient availability treatment (low-n) during the period of sap flow mea-urements. Each point represents the average value of four measured trees and therror bars are the standard error around the mean. The significance of the differencesetween the treatments was estimated using repeated measures ANOVA (P > 0.05).
point represents the average value of four measured trees and the error bars are thestandard error around the mean. The significance of the differences between thetreatments was estimated using repeated measures ANOVA (P < 0.01).
treatment, even when VPD or soil water potential (data not shown)did not change. However, the nocturnal F was marginal in the low-n treatment and appeared mainly during the first hour of the night(Fig. 1).
The fertilized trees had significantly higher concentrations ofNleaf, Nroot, Proot, Sroot and Caroot (Table 1), whereas the micronu-trients (Cu, Mn, Zn, Fe) did not differ between the two treatments(data not shown). Trees from the high-n treatment demonstratedgreater total tree biomass, total foliar biomass, total foliage areaand SRL (Table 2). The fine-root biomass was significantly higher inthe low-n treatment. The Nroot described 95% (R2 = 0.95; P < 0.001)of the variability in total foliar biomass (Fig. 5), whereas the Nleafexplained 88% (R2 = 0.88; P < 0.001) and the Proot described 65%(R2 = 0.65; P < 0.05).
4. Discussion
Our experiment revealed significantly higher nocturnal sap fluxdensity (F) and night-time water use percentage (NWU) in fer-tilized trees. This finding, however, is rather exceptional in thelight of current knowledge about trees’ nocturnal water relations
Fig. 5. Average night-time water use percentage from daytime water use (NWU)versus leaf nitrogen concentration (A) and average foliar biomass versus fine-rootnitrogen concentration (B). Values of R2 and P have been obtained from simpleregression analysis.

P. Kupper et al. / Environmental and Exp
Table 1Concentrations (mg kg−1) of mineral elements in leaves and fine-roots in two nutri-ent availability treatments. The differences of the means were estimated usingStudent’s t-test or Mann–Whitney U-test. na – P-value not applicable because oflimited sample size; ns – statistically not significant at level of 0.05; **P < 0.01;***P < 0.001.
Element High-n Low-n P N
Nleaf 23,900 9700 *** 8Nroot 11,400 8200 *** 8Kleaf 17,955 10,772 na 4Kroot 4902 4252 ns 8Caleaf 14,655 16,895 na 4Caroot 9911 7421 ** 8Mgleaf 1521 3258 na 4Mgroot 2449 2083 ns 8Pleaf 1653 1501 na 4Proot 2271 1538 ** 8
afletf2l2etmibfrtflea2
gttihTtfo(sscD
TAfU*
Sleaf 2215 1799 na 4Sroot 1395 1022 *** 8
s the increased need for nutrients transferred via transpirationux has been observed in nutrient-poor environments (Scholzt al., 2007). Nevertheless, some studies have shown that night-ime stomatal openness and transpiration are characteristic ofast-growing shade-intolerant pioneer species (Daley and Phillips,006; Marks and Lechowicz, 2007) and also for the sun foliage
ocated in the upper canopy position of trees (Kupper and Sellin,009; Sellin and Lubenets, 2010); thus, for trees or canopy lay-rs characterized by high leaf nitrogen concentration. In our study,he highest values of nocturnal F and NWU in the high-n treat-
ent were strongly associated with high foliar N concentration. Its probable that intense day- and night-time transpiration coulde beneficial, and could give an advantage in competition toast-growing species like aspen, which usually occupy nutrient-ich soils. In fertile soils the competition for light is expectedo be high, and in these circumstances nocturnal transpirationalux could have an adaptive nature in natural conditions. How-ver, increased nocturnal transpiration in response to nutrientvailability is probably of species-specific nature (Caird et al.,007).
Increased F in the high-n treatment during intensive foliagerowth period suggests that night-time water flux is greatest whenhe plant need for nitrogen is highest. It has been demonstratedhat the need for nitrogen is related to the nitrate reductase activ-ty, which depends on growth phase of an individual leaf and isighest when the leaf area expands intensively (Marschner, 2002).his could explain why the difference in nocturnal F between thereatments was smallest when the leaf area of the trees was almostull-grown. Increased nocturnal transpiration has been previouslybserved at the beginning of an intensive growth period in PinusGrulke et al., 2004) as well as in Eucalyptus (Phillips et al., 2010)
pecies. Seasonal variability could be one possible explanation whyeveral studies have not detected increased night-time stomatalonductance in diverse species (Caird et al., 2007; Howard andonovan, 2007; Christman et al., 2009), including Populus (Howardable 2verage values of tree variables in two nutrient availability treatments. The dif-
erences of the means were estimated using Student’s t-test or Mann–Whitney-test (N = 8). ns – statistically not significant at level of 0.05; *P < 0.05; **P < 0.01;**P < 0.001.
Variable High-n Low-n P
Total tree biomass, Bt (g) 173 130 *Total foliar biomass, Bf (g) 43.0 22.3 ***Total leaf area, ATL (m2) 0.70 0.29 **Total fine-root biomass, Bfr (g) 6.09 11.7 *Total fine-root area, Afr (cm2) 1236 1650 nsSpecific fine-root length, SRL (m g−1) 96.8 62.8 *
erimental Botany 82 (2012) 37– 42 41
and Donovan, 2010), as a result of higher soil N availability. Anotherexplanation is that the response of water flux to nitrate availabil-ity is biphasic (Wilkinson et al., 2007), and therefore a parabolicresponse of nocturnal transpiration to nitrate availability could beexpected.
There was a significant increase in nocturnal sap flux density inthe high-n treatment in the dark in the early morning, whereas theVPD and soil water potential were constant during the observedtime period. Such an endogenous increase in F was most likelycaused by the gradual opening of the stomata, while it was notobserved in trees grown in nutrient deficiency. Endogenous, grad-ual increases in stomatal opening during predawn hours have beenreported in other species too (Bucci et al., 2004; Dodd et al., 2005;Howard and Donovan, 2007). Such circadian regulation might bean important factor affecting stomatal opening and photosynthe-sis during predawn hours (Caird et al., 2007; Dawson et al., 2007).However, it may also support nitrogen uptake from soil as night-time water flux enables to avoid the accumulation of differentsoluble nitrogen compounds in fine roots where they may inhibitnitrate and ammonium uptake (Gessler et al., 1998; Geßler et al.,2002). Hence, the lack of the endogenous increase in nocturnalwater flux in the low-n treatment may associate with restricteduptake and delivery of nitrogen.
The foliar N concentration was very low in the nutrient defi-ciency treatment according to threshold value given by Van denBurg (1990). However, the average leaf N concentration in fer-tilized trees was sufficient for hybrid aspen growth, being closeto the optimum (Tullus et al., 2010). The average levels of otherfoliar macronutrients (K, Ca, Mg and P) were sufficient for hybridaspen growth (Van den Burg, 1990; Jug et al., 1999) also in theless fertilized (low-n) treatment. The fine-root Ca and S concen-trations in the low-n treatment were higher than the overallaverage concentrations of Ca and S in plant tissues according toMarschner (2002). Although the average level of Pleaf was sufficientfor hybrid aspen growth, we cannot exclude the possibility that sig-nificantly lower fine-root P concentration in the low-n treatmentcould lead to an additional decrease in the uptake and reduc-tion of NO3
− nitrogen in roots, as shown in several crop speciesgrown in phosphorus deficiency (Schjorring, 1986; Rufty et al.,1990).
The two-fold lower fine-root biomass and significantly higherSRL enabled the exploitation of a unit of soil volume at substan-tially smaller cost in the fertilized treatment. Hence, the treestend to invest more resources into development of extensive rootsystem than into morphological adaptations of fine roots in nutri-ent deficiency. However, under high root density conditions theeffect of mass-flow on nitrogen supply is probably reduced (Crameret al., 2008). Thus, we assume that the trees in the high-n treat-ment could benefit relatively more from the mass flow of nitratenitrogen, which solubility and mobility in soil is high (Barber,1995).
In conclusion, the increased night-time sap flux density (F)and night-time water use percentage relative to daytime wateruse (NWU) are characteristic to hybrid aspen in fertile condi-tions, whereas the intensity of water flux depends on the leafarea increment (on seasonal scale) and on endogenous factors(on daily scale). Increased water flux in fertile conditions co-occurs with increased specific fine-root length (SRL), and withdecreased fine-root biomass. These adaptations allow the acqui-sition of nutrients via mass flow more efficiently in fertileconditions, with lower investment in fine-root construction, andthe allocation of more resources into canopy growth. Our find-
ings suggest that decreased night-time water flux as a result ofstrong nutrient (especially nitrogen) deficit could be characteristicto fast-growing tree species, which are adapted to grow in fertilesoil.
4 nd Exp
A
(aEEKtatc
Ams
F
NNPKMBMCMZFS
R
B
B
C
C
C
D
D
2 P. Kupper et al. / Environmental a
cknowledgements
The study was supported by the Estonian Science FoundationGrant Nos. 7736 and 9186), by the Estonian Ministry of Educationnd Research (target financing project SF0180025s12), and by theU through the European Regional Development Fund (Centre ofxcellence in Environmental Adaptation). We are grateful to Dr.rõõt Aasamaa, Ingmar Tulva and Olaf Räim for technical advice on
he measurements. We thank Kätlin Alasepp and Indrek Loolaid forssistance in the lab and Ilmar Part for language revision. We alsohank two anonymous reviewers for critical remarks and helpfulomments on the previous version of the manuscript.
ppendix A. Concentrations (mg kg−1) of fertilizers andineral elements added to sphagnum peat growth
ubstrate in two nutrient availability treatments.
ertilizers and elements High-n Low-n
O3− 606 308
H4+ 432 220
2O5 1189 6162O 2078 1057gO 245 92
8.7 1.3o 41 8.8
u 78 5.3n 46 7.0
n 30 1.8e 128 4.0
23 0
eferences
arber, S.A., 1995. Soil Nutrient Bioavailability: A Mechanistic Approach, 2nd edn.Wiley, New York.
ucci, S.J., Scholz, F.G., Goldstein, G., Meinzer, F.C., Hinojosa, J.A., Hoffman, W.A.,Franco, A.C., 2004. Processes preventing nocturnal equilibration between leafand soil water potential in tropical savanna woody species. Tree Physiology 24,1119–1127.
aird, M.A., Richards, J.H., Donovan, L.A., 2007. Nighttime stomatal conductance andtranspiration in C3 and C4 plants. Plant Physiology 143, 4–10.
hristman, M.A., Donovan, L.A., Richards, J.H., 2009. Magnitude of nighttime tran-spiration does not affect plant growth or nutrition in well-watered Arabidopsis.Physiologia Plantarum 136, 264–273.
ramer, M.D., Hoffmann, V., Verboom, G.A., 2008. Nutrient availability moderatestranspiration in Ehrharta calycina. New Phytologist 179, 1048–1057.
aley, M.D., Phillips, N.G., 2006. Interspecific variation in nighttime transpiration
and stomatal conductance in a mixed New England deciduous forest. Tree Phys-iology 26, 411–419.awson, T.E., Burgess, S.S.O., Tu, K.P., Oliveira, R.S., Santiago, L.S., Fisher, J.B., Simonin,K.A., Ambrose, A.R., 2007. Nighttime transpiration in woody plants from con-trasting ecosystems. Tree Physiology 27, 561–575.
erimental Botany 82 (2012) 37– 42
Delhon, P., Gojon, A., Tillard, P., Passama, L., 1995. Diurnal regulation of NO3−
uptake in soybean plants I. Changes in NO3− influx, efflux and utilization
in the plant during the day/night cycle. Journal of Experimental Botany 46,1585–1594.
Dodd, A.N., Salathia, N., Hall, A., Kevei, E., Toth, R., Nagy, F., Hibberd, J.M., Millar,A.J., Webb, A.A.R., 2005. Plant circadian clocks increase photosynthesis, growth,survival, and competitive advantage. Science 309, 630–633.
Gessler, A., Schneider, S., Von Sengbuch, D., Weber, P., Hanemann, U., Huber, C.,Rothe, A., Kreutzer, K., Rennenberg, H., 1998. Field and laboratory experimentson net uptake of nitrate and ammonium by the roots of spruce (Picea abies) andbeech (Fagus sylvatica) trees. New Phytologist 138, 275–285.
Geßler, A., Kreuzwieser, J., Dopatka, T., Rennenberg, H., 2002. Diurnal courses ofammonium net uptake by the roots of adult beech (Fagus sylvatica) and spruce(Picea abies) trees. Plant and Soil 240, 23–32.
Grulke, N.E., Alonso, R., Nguyen, T., Cascio, C., Dobrowolski, W., 2004. Stomata openat night in pole-sized and mature ponderosa pine: implications for O3 exposuremetrics. Tree Physiology 24, 1001–1010.
Howard, A.R., Donovan, L.A., 2007. Helianthus nighttime conductance and transpi-ration respond to soil water but not nutrient availability. Plant Physiology 143,145–155.
Howard, A.R., Donovan, L.A., 2010. Soil nitrogen does not impact nighttime waterloss in Populus. Tree Physiology 30, 23–31.
Jug, A., Hofmanlow-aSchielle, C., Makeschin, F., Rehfuess, K.E., 1999. Short-rotationplantations of balsam poplars, aspen and willows on former arable land in theFederal Republic of Germany. II. Nutritional status and bioelement export byharvested shoot axes. Forest Ecology and Management 121, 67–83.
Kupper, P., Sellin, A., 2009. The intensity of night-time water use differswithin a crown of a forest-grown Tilia cordata tree. Acta Horticulturae 846,229–235.
Kupper, P., Sõber, J., Sellin, A., Lõhmus, K., Tullus, A., Räim, O., Lubenets, K., Tulva,I., Uri, V., Zobel, M., Kull, O., Sõber, A., 2011. An experimental facility for freeair humidity manipulation (FAHM) can alter water flux through deciduous treecanopy. Environmental and Experimental Botany 72, 432–438.
Marschner, H., 2002. Mineral Nutrition of Higher Plants, 2nd edn. Academic Press,London.
Marks, C.O., Lechowicz, M.J., 2007. The ecological and functional correlates of noc-turnal transpiration. Tree Physiology 27, 577–584.
Phillips, N.G., Lewis, J.D., Logan, B.A., Tissue, D.T., 2010. Inter- and intra-specific vari-ation in nocturnal water transport in Eucalyptus. Tree Physiology 30, 586–596.
Rufty, T.W., MacKown, C.T., Israel, D.W., 1990. Phosphorus stress effects on assimi-lation of nitrate. Plant Physiology 94, 328–333.
Schjorring, J.K., 1986. Nitrate and ammonium absorption by plants growing at asufficient or insufficient level of phosphorus in nutrient solutions. In: Lambers, H.(Ed.), Fundamental, Ecological and Agricultural Aspects of Nitrogen Metabolismin Higher Plants. Martinus Nijhoff, Dordrecht.
Scholz, F.G., Bucci, S.J., Goldstein, G., Meinzer, F.C., Franco, A.C., Miralles-Wilhelm,F., 2007. Removal of nutrient limitations by long-term fertilization decreasesnocturnal water loss in savanna trees. Tree Physiology 27, 551–559.
Sellin, A., Lubenets, K., 2010. Variation of transpiration within a canopy of silverbirch: effect of canopy position and daily versus nightly water loss. Ecohydrology3, 467–477.
Tullus, A., Kanal, A., Soo, T., Tullus, H., 2010. The impact of available water content inprevious agricultural soils on tree growth and nutritional status in young hybridaspen plantations in Estonia. Plant and Soil 333, 129–145.
Van den Burg, J., 1990. Foliar Analysis for Determination of Tree Nutrient Saturation
– A Compilation of Literature Data. Institute of Forestry and Urban Ecology, DeDorschkamp, Wageningen.Wilkinson, S., Bacon, M.A.Z., Davies, W.J., 2007. Nitrate signalling to stomata andgrowing leaves: interactions with soil drying, ABA, and xylem sap pH in maize.Journal of Experimental Botany 58, 1705–1716.