distribution of protein kinase c isozymes in rat optic nerves
TRANSCRIPT

Journal of Neuroscience Research 29:379-389 (1991)
Distribution of Protein Kinase C Isozymes in Rat Optic Nerves S. Komoly, Y. Liu, H. deF. Webster, and K.-F. J. Chan Laboratory of Experimental Neuropathology , National Institute of Neurological Disorders and Stroke, National Institutes of Health, Bethesda, Maryland
Light (LM) and electron (EM) microscopic immuno- cytochemical methods were used to study the distri- bution of protein kinase C (PKC) isozymes in adult rat optic nerves. In cryostat and vibratome sections examined by LM, type I1 (p) isozyme was localized almost exclusively in the axons. In the EM, immu- noreaction products were found to associate with mi- crotubules and neurofilaments. The inner surface of axonal membranes were occasionally stained. Analy- sis of PKC isozyme composition of the optic nerves by using immunoblot techniques revealed that type I1 (p) isozyme accounted for approximately 80% of the total immunoreactivity. By contrast, type I11 (a) isozyme, which accounted for the remaining 20% of PKC, was found mainly in the astrocytes. Astrocytic processes next to blood vessels and between myeli- nated axons were stained. In the EM, immunoreac- tion products were found in the cytoplasm and along astroglial filaments. Segments of plasma membranes also were stained; but nuclei were unstained. Adult glial cells were not stained by an antibody to type I1 (p) isozyme except for the occurrence of a few punc- tate cytoplasmic densities in occasional astrocytes. Very faint or no immunostaining was observed in sec- tions treated with a monoclonal antibody to type I ( y ) isozyme. Immunoblot analyses also did not reveal this subspecies. The absence of type I (y) isozyme in optic nerves is not due to a down-regulation of the enzyme during development. In developing (5 and 11 day) rats, immunoreactivity of protein kinase C was very faint or absent. After 15 days, reaction products of both type I11 (a) and type I1 (p) isozymes were found throughout the nerve. These findings suggest that type I1 (p) isozyme may be involved in axonal trans- port whereas type I11 (a) isozyme may play a role in some astrocyte functions in mature optic nerves.
Key words: protein kinase C, isozymes, optic nerve, axons, astrocytes, glial filaments, microtubules
’
INTRODUCTION Protein kinase C (PKC) is a calcium-activated and
phospholipid-dependent serinefthreonine protein kinase
Published 1991 by Wiley-Liss, Inc.
ubiquitous in eukaryotes. In mammals, this enzyme is found in high concentrations in the central nervous sys- tem. Protein kinase C-mediated protein phosphorylation has been implicated in the regulation of a number of biological events (for review, see Nishizuka, 1986; Kik- kawa and Nishizuka, 1986; Kaczmarek, 1987; Huang, 1989). In the nervous system, protein kinase C may play important roles in neurotransmitter release, neuritogene- sis, and memory and learning (Kaczmarek, 1987; Bur- goyne, 1989; Chiarugi et al., 1989). Seven isoforms of protein kinase C (a, PI, PII, y, 6, E, 5 ) have thus far been identified by genetic approaches (Coussens et al., 1986; Nishizuka, 1988; Kikkawa et al., 1989). These isoforms are encoded by different genes with the excep- tions of PI and PII, which are derived from the same mRNA transcript by alternative splicing (Nishizuka, 1988). Protein kinase C purified from the brain contains three major isozymes termed type I, type 11, and type 111. These subspecies are encoded by cDNA corresponding to y, PI + PII, and a; respectively. Together, they con- stitute over 90% of the total protein kinase C activity in the brain (Huang and Huang, 1986).
The exact physiological significance of the various PKC isozymes is still unclear. Biochemical studies re- vealed that different isozymes have slightly different substrate specificities and requirements of activators (Kikkawa et al., 1989). Several PKC isozymes also were found to have different subcellular distributions and ex- pression during development of the brain. Theoretically, each isozyme may have distinct biological functions. This may be due to the presence of specific protein or enzyme substrates in different subcellular locations. Al-
Received September 28, 1990; revised December 13, 1990; accepted December 13, 1990.
Address reprint requests to Dr. S . Komoly, National Institutes of Health, Building 36, Room 4A-29, Bethesda, MD 20892.
Dr. K.-F.J. Chan is now at the Department of Biochemistry, Univer- sity of Hong Kong, Hong Kong.
Abbreviations used: PKC, protein kinase C; PBS, phosphate-buffered saline; HEPES N-2-hydroxyethylpiperazine-N’-ethanesulfonic acid; EGTA, [ethylenebis(oxyethylenenitrilo)tetraacetic acid; SDS, sodium dodecyl sulfate.

380 Komoly et al.
ternatively, the presence of a particular PKC isozyme in specific cell types may come as a result of unique cellular regulation of the allosteric activators and inhibitors. So far, most of the brain regions including cerebral cortex, striatum, thalamus, spinal cord, hippocampus, septum, and mesencephalon were found to contain two or more isozymes (Mochly-Rosen et al., 1987; Huang et al., 1989; Saito et al., 1988; Yoshida et al., 1988). Expres- sion of a single subspecies was observed only in a few cell types, e.g., Purkinje cells and olfactory bulb mitral cells (Hidaka et al., 1988; Young, 1988). In this report, we show that the distribution of PKC isozymes in adult rat optic nerves also is cell-specific. Type I1 (6) isozyme was found predominantly in axons whereas type I11 (a) isozyme was localized mainly in astrocytes. No signifi- cant immunoreactivity was observed for type I (y) isozyme. These results suggest that the optic nerve may be a good anatomical system for further studies of the functions and regulations, as well as the mechanisms of gene expression, of protein kinase C isozymes.
EXPERIMENTAL PROCEDURES Materials
Monoclonal antibodies to protein kinase C isozymes type I (y), type I1 (p), and type 111 (a) were obtained from Seikagaku Kogyo Co., Ltd. The antibod- ies were reconstituted to a concentration of 0.1 mgiml and stored in aliquots at -20°C. A 1:lOO dilution of these antibodies was used in both immunostaining and immunoblotting experiments. Affinity purified anti- mouse biotinylated horse IgG, normal horse serum, nor- mal mouse IgG, and the standard ABC Elite horseradish peroxidase kit were purchased from Vector Laboratories, Inc. The sera were diluted in 0.1 M phosphate-buffered saline (PBS) containing 1% bovine serum albumin and 0.1 % sodium azide. The embedding medium for frozen tissue specimens, Tissue-Tek, was from Miles Labora- tories. Nitrocellulose membranes were from Schleicher and Schuell, Inc. Casein was from Sigma Chemical Co. '251-labeled protein G was a product of Amersham Corp. Paraformaldehyde, glutaraldehyde, uranyl acetate 3,3- diaminobenzidine tetrahydrochloride (DAB), and Poly/ Bed 812 were purchased from Polysciences, Inc. Protein kinase C was purified to apparent homogeneity from the particulate fractions of rat brains essentially according to the previously described procedures (Huang et al., 1986).
Light Microscopic Immunocytochemistry Adult and developing ( 5 , 1 1, 15, 18, and 27 days)
Wistar rats were anesthetized and perfused through the heart with physiological saline followed by a fixative
containing 4% paraformaldehyde, 15% saturated picric acid, and 0.1 M PBS. The optic nerves were dissected out and postfixed overnight in the same fixative. Cryo- protection of the samples was carried out in a solution containing 0.9% (w/v) sodium chloride, 20% (w/v) su- crose, and 3% (v/v) polyethylene glycol. After 24 hr, the optic nerves were embedded in Tissue-Tek and frozen in dry ice-cold isopenthane. Serial sections of 6-12 pm were cut in a cryostat (2800 Frigocut, Reichert-Jung) and mounted on gelatin-covered slides. The samples were kept at 4°C until used.
The sections were washed in PBS containing 0.5% of Triton X-100 for 3 hr to remove the cryoprotection reagents and to permeabilize the cell membranes. Non- specific protein binding sites were blocked by incubating the specimen with horse serum. Immunoreaction with different monoclonal antibodies to protein kinase C was carried out at 4°C for 16-24 hr. After washing, the spec- imens were incubated with biotinylated anti-mouse TgG (1:300 dilution) for 60 min at room temperature. Color reaction was developed by using ABC Elite horseradish peroxidase kit with 0.05% DAB and 0.005% H,O, in PBS.
Two sets of control experiments were performed. These included ( 1 ) substitution of monoclonal anti-PKC antibodies with normal mouse IgG; and (2) preincubation of the primary antibodies with purified protein kinase C before immunostaining.
Electron Microscopic Immunocytochemistry Four adult (180 g) Wistar rats were perfused as
described above except that the fixative also contained 0.5% glutaraldehyde. The optic nerves were removed with the brain; they were postfixed overnight, and trans- ferred into PBS. Sections of 30 pm were cut with a BSK Slicer. Vibratome sections were immunostained as de- scribed for LM but without prior detergent treatment. After color development with DAB, the sections were postfixed with 0.5-1 % osmium tetroxide in the presence or absence of 0.6% potassium fenicyanide and 5% su- crose for 60 min and embedded flat in epon. Thin sections were cut with a Reichert-Jung ultramicrotome and mounted on Formvar-coated single slot grids. Every sec- ond one of the serial sections was stained with uranyl acetate for 12 min and then with lead citrate for 30 sec in a LKB 21 68 ultrastainer. The remaining sections were not stained with heavy metals. The specimens were examined in a Philips EM 410 electron microscope at 80 kV.
Immunoblotting of Protein Kinase C Isozymes Five adult Wistar rats were anesthetized and killed
by decapitation. The optic nerves were removed from the brain and immediately homogenized in 20 mM HEPES,

Protein Kinase C Isozyrnes in Optic Nerve 381
immunostaining was similar to those obtained by using a biotinylated mouse IgG or after preincubation of the monoclonal antibodies with purified rat brain protein ki- nase C.
Immunostaining of the three PKC isozymes in vi- bratome sections was similar to those observed in frozen sections. However, no immunoreactivity was found in paraffin-embedded sections. Prior delipidation of vi- bratome and frozen sections with ethanol also abolished the immunolabeling of protein kinase C by monoclonal antibodies (results not shown).
In adult rat retina, immunostaining of type I1 (p) isozyme was localized mainly in ganglion cells whereas type I11 (a ) subspecies was found in Muller cells (results not shown). There were no immunoreaction products to type I (y) isozyme. These findings are in agreement with the immunoblotting data (Yosida et al., 1988) and sup- port our observations on the distribution of protein kinase C in the optic nerves.
Immunoblot Analysis of Protein Kinase C Isozymes The composition of PKC isozymes in adult rat op-
tic nerves were quantitated by using immunoblot analy- ses. The results are summarized in Table I. Over 80% of the total PKC immunoreactivity was due to type I1 (p) isozyme. Subcellular distribution studies revealed that approximately two-thirds of this PKC subspecies were in the soluble (cytosolic) fractions. The immunoreaction of the monoclonal antibody with type I1 (p) isozyme was specific. Only one radioactive band with an apparent M , of 80,000 was observed in the soluble and the particulate fractions of the optic nerves (Fig. 2). These findings suggest that protein kinase C, in particular, the type I1 (p) isozyme, may play an important role in axonal func- tions. Type 111 (a) isozyme accounts for the remaining 20% of PKC immunoreactivity in the optic nerves (Table I). Nearly 70% of this isozyme was found in the cyto- solic fractions. By contrast, no significant immunoreac- tivity of type I (y) isozyme was observed. The absence of immunoreaction of the monoclonal antibody to type I (y) subspecies was not due to its ineffectiveness. This anti- body could recognize type I (y) isozyme in purified pro- tein kinase C preparations (data not shown).
Electron Microscopic Localization of PKC Isozymes To learn more about the putative functions of pro-
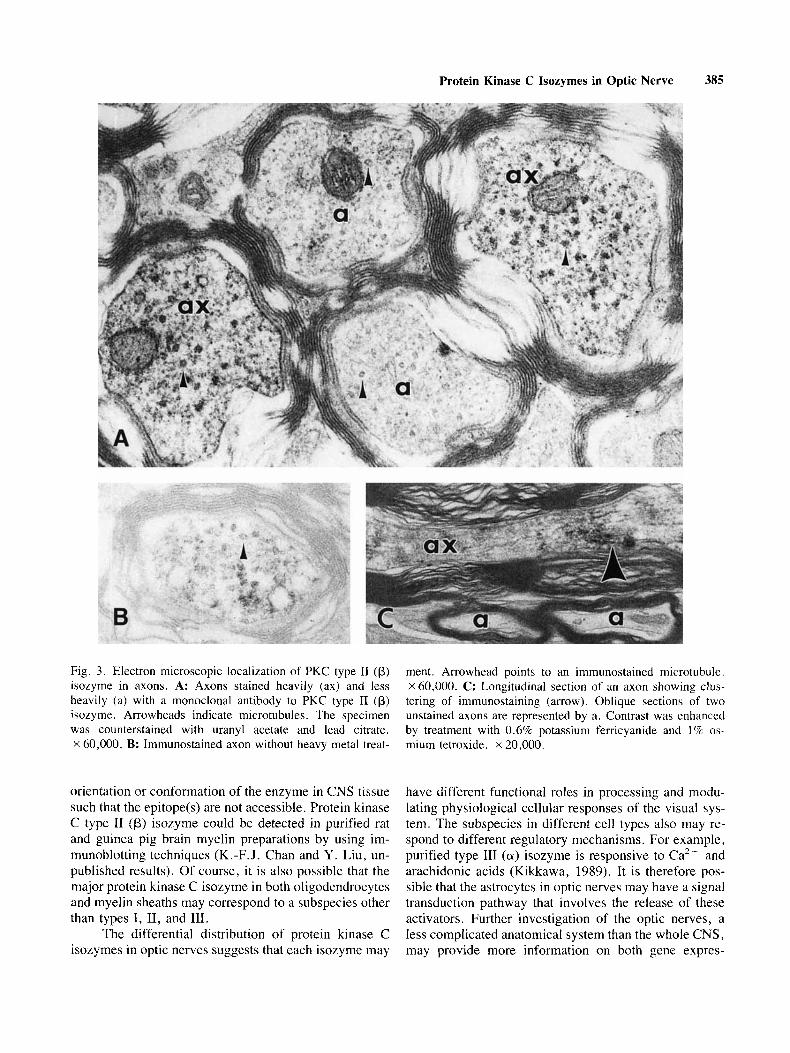
tein kinase C isozymes in optic nerves, the subcellular localization of both type I1 (p) and type 111 (a) subspecies were investigated by using electron microscopic immu- nocytochemical methods. As shown in Figure 3 , immu- noreaction products of type I1 (p) isozyme were associ- ated with microtubules in axons. The punctate staining in the cytoplasm might be due to the labeling of neurofila- ments (Fig. 3A-C). Occasionally, immunoreaction prod-
pH 7.4, 0.5 mM EGTA, 2 mM EDTA, 20 mM NaCl, and 0.5 mM dithiothreitol (buffer A). All procedures were carried out at 0-4°C. The homogenates were cen- trifuged at 15,OOOg for 30 min. The resulting supernatant fluids were saved as soluble fractions. The pellets were resuspended in buffer A and saved as particulate frac- tions.
The proteins (30 pg) were separated by using SDS- polyacrylamide gel (10%) electrophoresis and electro- blotted onto nitrocellulose membranes. The blotting buffer contained 30 mM Tris-HC1, 200 mM glycine, pH 8.3, and 20% (v/v) methanol. After washing the mem- branes with 50 mM Tris-HC1, pH 7.4, and 150 mM NaCl (TBS) containing 0.05% Tween-20, nonspecific binding sites were blocked by overnight incubation with 2% (wl v) casein. Immunoreactions of the monoclonal antibod- ies (1 pglml) with PKC isozymes types I (y), I1 (p), and I11 (a) were carried out at 25°C for 2 hr. Reaction prod- ucts were detected by incubating the membranes with
I-labeled protein G (containing 1 X lo7 cpm) in TBS for 30 min at 25°C and subsequent autoradiography. Im- munoreactivity was quantitated by both densitometric scanning of the corresponding PKC bands with a Shi- madzu CS-9000 flying-spot densitometer and gamma counting of the excised PKC-containing nitrocellulose membranes using a LKB-Wallac 1272 Clini-Gamma counter.
I 2 5
RESULTS Distribution of PKC Isozyrnes in Adult Rat Optic Nerves
To better understand the function and regulation of different protein kinase C isozymes in the brain, we searched for a well-defined anatomical system in which separate PKC subspecies are localized in different cell types. The optic nerves of adult rats were found to pos- sess these characteristics.
Light microscopic immunocytochemical studies of the cryostat sections revealed an almost exclusive local- ization of PKC type 111 (a) isozyme in the astrocytes (Fig. 1A). Immunoreaction products of a monoclonal antibody to this isozyme were found both in the cyto- plasm and in the astrocytic processes around blood ves- sels and between meylinated axons (Fig. IA,B). The nuclei were unstained. By contrast, immunoreactivity of PKC type I1 (p) isozyme was confined mainly to axons (Fig. lC,D). Most, if not all, of the axons were immu- noreactive. Adult glial cells were not stained by the monoclonal antibody to this type I1 (p) subspecies except for the occurrence of a few punctate cytoplasmic densi- ties in occasional astrocytes. Very faint or no immuno- reactivity was found in cryostat sections treated with an antibody to PKC type I (y) isozyme (Fig. lE,F). The
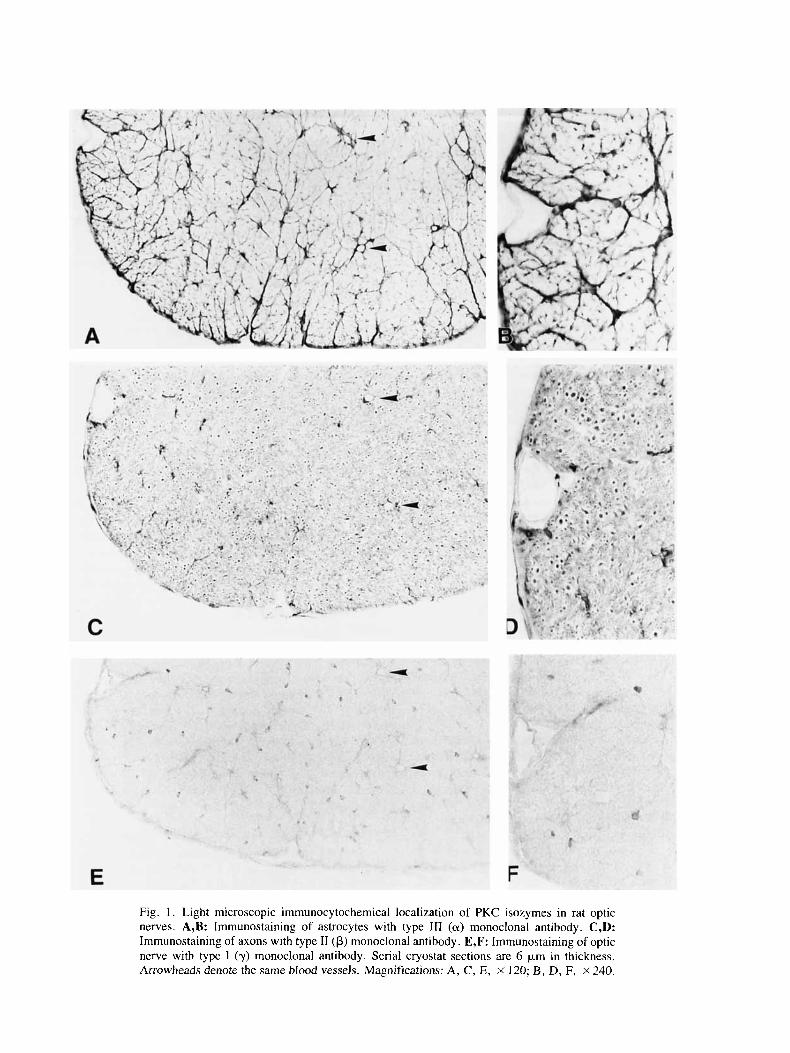
Fig. 1. Light microscopic immunocytochemical localization of PKC isozymes in rat optic nerves. A,B: Irnrnunostaining of astrocytes with type 111 (a) monoclonal antibody. C,D: Irnrnunostaining of axons with type I1 (p) monoclonal antibody. E,F: Immunostaining of optic nerve with type I (y) monoclonal antibody. Serial cryostat sections are 6 p,m in thickness. Arrowheads denote the same blood vessels. Magnifications: A, C, E, X 120; B, D, F, ~ 2 4 0 .

Protein Kinase C Isozymes in Optic Nerve 383
products were observed during the first two postnatal weeks (results not shown). Gradually, dense immuno- staining was found similar to the patterns shown in Fig- ure 1C and D. There was no positive staining with the monoclonal antibody to PKC type I (y) isozyme, regard- less of age (data not shown). This observation suggests that the failure to detect type I (y) isozyme in adult rat optic nerves is not due to a down-regulation of this sub- species during early development.
TABLE I. Distribution of Protein Kinase C Isozymes in Adult Rat Optic Nerves*
Protein kinase C isozymes (%)
I (Y) 11 (PI I11 (ff)
Total - 80.8 ? 3.3 19.2 ? 3.5 Subcellular
Soluble - 67.2 ? 7.1 69.2 & 5.8 Particulate - 32.8 ? 7.2 30.8 ? 6.4
*The composition of protein kinase C isozymes in adult rat optic nerves was determined by immunoblot assays as described under Ex- perimental Procedures. lmmunoreactivity of monoclonal antibodies to PKC isozymes type I (y), type I1 (p), and type 111 (a) was determined by using '251-1abeled protein G overlay and quantitated by densito- metric scanning of the corresponding radioactive PKC bands.
ucts were found to associate with the internal surface of the axolemma and on the outer membranes of the mito- chondria. No diffuse staining was observed in the axo- plasm. It should be noted that the intensity of the immu- nostaining could vary from axon to axon, and sometimes even within the same axon.
Immunoreaction products of PKC type 111 (a) isozyme were localized almost exclusively in astrocytes (Fig. 4). The astrocytes were identified by their ultra- structural features (Peters et al., 1991) as well as by their immunoreactivity with an antibody to glial fibrillary acidic protein (GFAP) (results not shown). Within the astrocytes, diffuse reaction products were observed in the cytoplasm (Fig. 4A). Glial fibers were stained (Figs. 4B ,C). Mitochondria were sometimes labeled. Strong immunoreactivity also was found to associate with cell membranes (Figs. 4C ,D), particularly at contact regions between perivascular astroglial processes (Figs. 4D,E). The nuclei were unstained. A few endothelial cells in the optic nerve were occasionally found to contain PKC type 111 (a ) isozyme immunoreactivity. In addition, faint re- action products were observed at tight junctions (results not shown).
No positive immunostaining of PKC type I (y) isozyme was seen in the EM.
Expression of PKC Isozymes During Development of the Optic Nerves
The distribution of the various PKC isozymes in the optic nerves of developing rats ( 5 , 1 1, 15, 18, and 27 days) was investigated by using LM immunocytochem- istry. From 5 to 15 days, reaction products of type I11 (a) subspecies were very faint or absent (Fig. 5A-C). After 15 days, immunopositive astrocytic-like cells were ob- served (Fig. 5D-F). Both the intensity and the number of stained cells were found to increase with age. The de- velopmental expression of PKC type I1 (p) isozyme in axons of rat optic nerves was similar to the type I11 (a) subspecies. Only weak and diffuse immunoreaction
DISCUSSION
Protein kinase C has been implicated in the regu- lation of a number of neural functions (Nishizuka, 1986; Coussen et al., 1986; Kaczmarek, 1987). This protein phosphorylation system also may play important roles in several pathological conditions such as ischemic brain injury (Louis et al., 1989; Olah et al., 1990) and brain edema formation (Joo et al., 1989). The exact functional roles of different protein kinase C isozymes in these and other processes still remain unclear. One problem is that many cell types in many regions of the brain express more than one subspecies. Our findings of an almost exclusive distribution of type I1 (p) isozyme in the axons and type 111 (a) isozyme in the astrocytes of adult rat optic nerves (Fig. 1) may provide a unique opportunity for further investigation of the function and regulation, and perhaps the mechanisms of gene expression, of these two isozymes.
In the EM, immunoreaction products of type 11 (p) isozyme were found to associate with microtubules and neurofilaments (Fig. 3 ) . This specific localization sug- gests that type I1 (p) isozyme may play a role in regu- lating cytoskeletal structures and axonal transport pro- cesses in axons. Indeed, microtubule-associated proteins such as MAP-2 and tau, and neurofilament proteins are putative substrates for protein kinase C (Kaczmarek, 1987; Nixon et al., 1987; Sihag et al., 1988; Pestronk et al., 1990). These proteins are phosphorylated in vivo. Immunostaining of axons with monoclonal antibodies to pII and y subspecies also have been reported in the cer- ebellum (Kose et al., 1988; Kikkawa et al., 1989; Saito et al., 1989). The functional significance of the other cytosolic type I1 (p) isozyme in axons (Fig. 2 and Table I) is less clear. In some cells, PI1 subspecies may play a role in protein processing (Kikkawa et al., 1989).
Protein kinase C type I11 (a) isozyme is more widely distributed in the astrocytes of optic nerves. Im- munoreactivity was observed in the cytoplasm as well as in association with glial filaments and cell membranes (Figs. 1 and 4). Furthermore, most, and perhaps all, of the astrocytes in the optic nerve were immunostained. These results are in agreement with the previous local- ization of PKC type I11 (a) isozyme in cultured astroglial
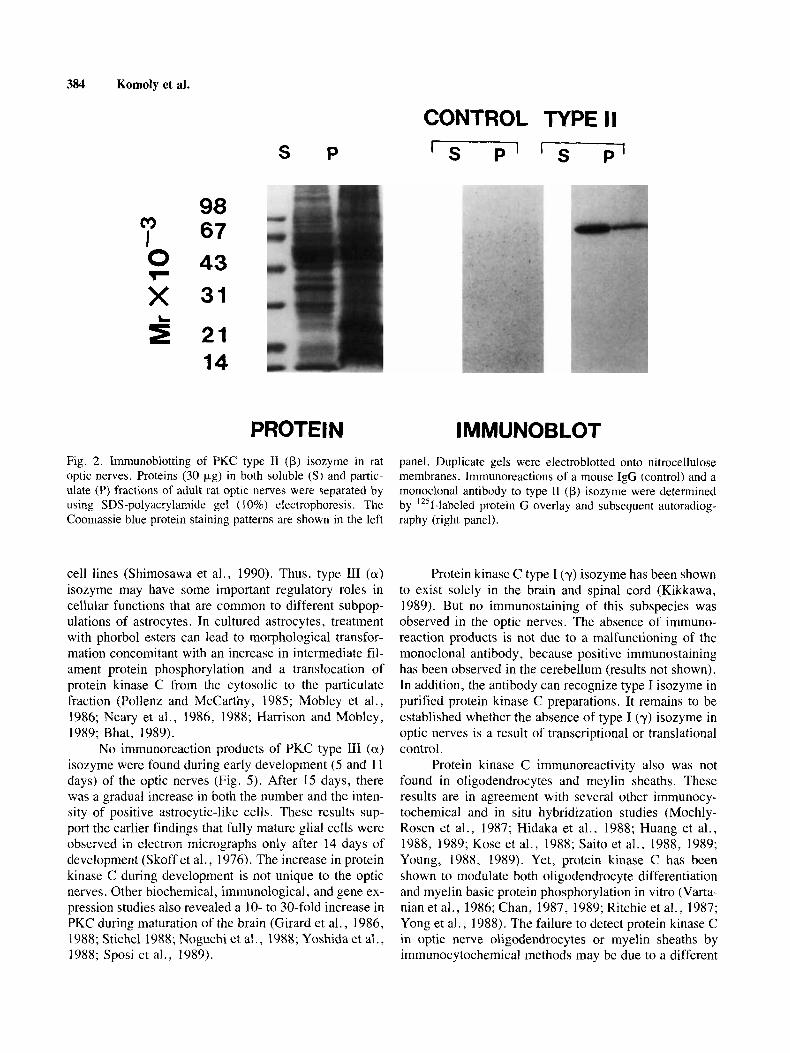
384 Komoly et al.
S P
98 c3 67 I 0 43 F
x 31
Fig. 2. Immunoblotting of PKC type I1 (p) isozyme in rat optic nerves. Proteins (30 pg) in both soluble (S) and partic- ulate (P) fractions of adult rat optic nerves were separated by using SDS-polyacrylamide gel (10%) electrophoresis. The Coomassie blue protein staining patterns are shown in the left
cell lines (Shimosawa et a]., 1990). Thus, type 111 (a) isozyme may have some important regulatory roles in cellular functions that are common to different subpop- ulations of astrocytes. In cultured astrocytes, treatment with phorbol esters can lead to morphological transfor- mation concomitant with an increase in intermediate fil- ament protein phosphorylation and a translocation of protein kinase C from the cytosolic to the particulate fraction (Pollenz and McCarthy, 1985; Mobley et al., 1986; Neary et al., 1986, 1988; Harrison and Mobley, 1989; Bhat, 1989).
No immunoreaction products of PKC type 111 (a) isozyme were found during early development (5 and 11 days) of the optic nerves (Fig. 5). After 15 days, there was a gradual increase in both the number and the inten- sity of positive astrocytic-like cells. These results sup- port the earlier findings that fully mature glial cells were observed in electron micrographs only after 14 days of development (Skoff et al., 1976). The increase in protein kinase C during development is not unique to the optic nerves. Other biochemical, immunological, and gene ex- pression studies also revealed a 10- to 30-fold increase in PKC during maturation of the brain (Girard et al., 1986, 1988; Stichel 1988; Noguchi et at., 1988; Yoshida et al., 1988; Sposi et al., 1989).
CONTROL TYPE II
' s P ' ' s P '
I MMUNOBLOT panel. Duplicate gels were electroblotted onto nitrocellulose membranes. Immunoreactions of a mouse IgG (control) and a monoclonal antibody to type I1 (p) isozyme were determined by '251-labeled protein G overlay and subsequent autoradiog- raphy (right panel).
Protein kinase C type I (y) isozyme has been shown to exist solely in the brain and spinal cord (Kikkawa, 1989). But no immunostaining of this subspecies was observed in the optic nerves. The absence of immuno- reaction products is not due to a malfunctioning of the monoclonal antibody, because positive immunostaining has been observed in the cerebellum (results not shown). In addition, the antibody can recognize type I isozyme in purified protein kinase C preparations. It remains to be established whether the absence of type I (y) isozyme in optic nerves is a result of transcriptional or translational control.
Protein kinase C immunoreactivity also was not found in oligodendrocytes and meylin sheaths. These results are in agreement with several other immunocy- tochemical and in situ hybridization studies (Mochly- Rosen et al., 1987; Hidaka et al., 1988; Huang et al., 1988, 1989; Kose et al., 1988; Saito et al., 1988, 1989; Young, 1988, 1989). Yet, protein kinase C has been shown to modulate both oligodendrocyte differentiation and myelin basic protein phosphorylation in vitro (Varta- nian et al., 1986; Chan, 1987, 1989; Ritchie et al., 1987; Yong et al., 1988). The failure to detect protein kinase C in optic nerve oligodendrocytes or myelin sheaths by immunocytochemical methods may be due to a different
Protein Kinase C Isozymes in Optic Nerve 385
Fig. 3. Electron microscopic localization of PKC type I1 (p) ment. Arrowhead points to an immunostained microtubule. isozyme in axons. A: Axons stained heavily (ax) and less X60,OOO. C: Longitudinal section of an axon showing clus- heavily (a) with a monoclonal antibody to PKC type I1 (p) tering of immunostaining (arrow). Oblique sections of two isozyme. Arrowheads indicate microtubules. The specimen unstained axons are represented by a . Contrast was enhanced was counterstained with uranyl acetate and lead citrate. by treatment with 0.6% potassium ferricyanide and 1 % os- X 60,000. B: Immunostained axon without heavy metal treat- mium tetroxide. X 20,000.
orientation or conformation of the enzyme in CNS tissue such that the epitope(s) are not accessible. Protein kinase C type I1 (p) isozyme could be detected in purified rat and guinea pig brain myelin preparations by using im- munoblotting techniques (K.-F.J. Chan and Y. Liu, un- published results). Of course, it is also possible that the major protein kinase C isozyme in both oligodendrocytes and myelin sheaths may correspond to a subspecies other than types I , 11, and 111.
The differential distribution of protein kinase C isozymes in optic nerves suggests that each isozyme may
have different functional roles in processing and modu- lating physiological cellular responses of the visual sys- tem. The subspecies in different cell types also may re- spond to different regulatory mechanisms. For example, purified type 111 (a) isozyme is responsive to Ca2+ and arachidonic acids (Kikkawa, 1989). It is therefore pos- sible that the astrocytes in optic nerves may have a signal transduction pathway that involves the release of these activators. Further investigation of the optic nerves, a less complicated anatomical system than the whole CNS, may provide more information on both gene expres-
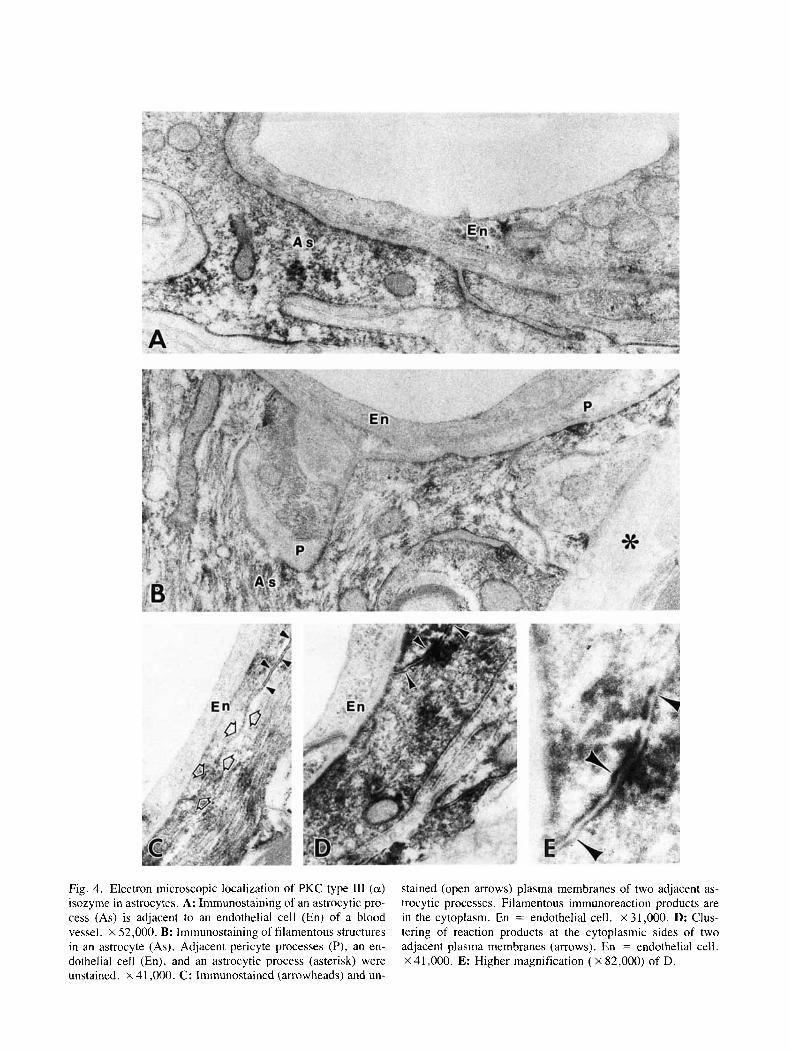
Fig. 4. Electron microscopic localization of PKC type 111 (a) isozyme in astrocytes. A: Immunostaining of an astrocytic pro- cess (As) is adjacent to an endothelial cell (En) of a blood vessel. x 52,000. B: Immunostaining of filamentous structures in an astrocyte (As). Adjacent pericyte processes (P), an en- dothelial cell (En), and an astrocytic process (asterisk) were unstained. x 41,000. C: Immunostained (arrowheads) and un-
stained (open arrows) plasma membranes of two adjacent as- trocytic processes. Filamentous immunoreaction products are in the cytoplasm. En = endothelial cell. X 31,000. D: Clus- tering of reaction products at the cytoplasmic sides of two adjacent plasma membranes (arrows). En = endothelial cell. X 41,000. E: Higher magnification ( X 82,000) of D.
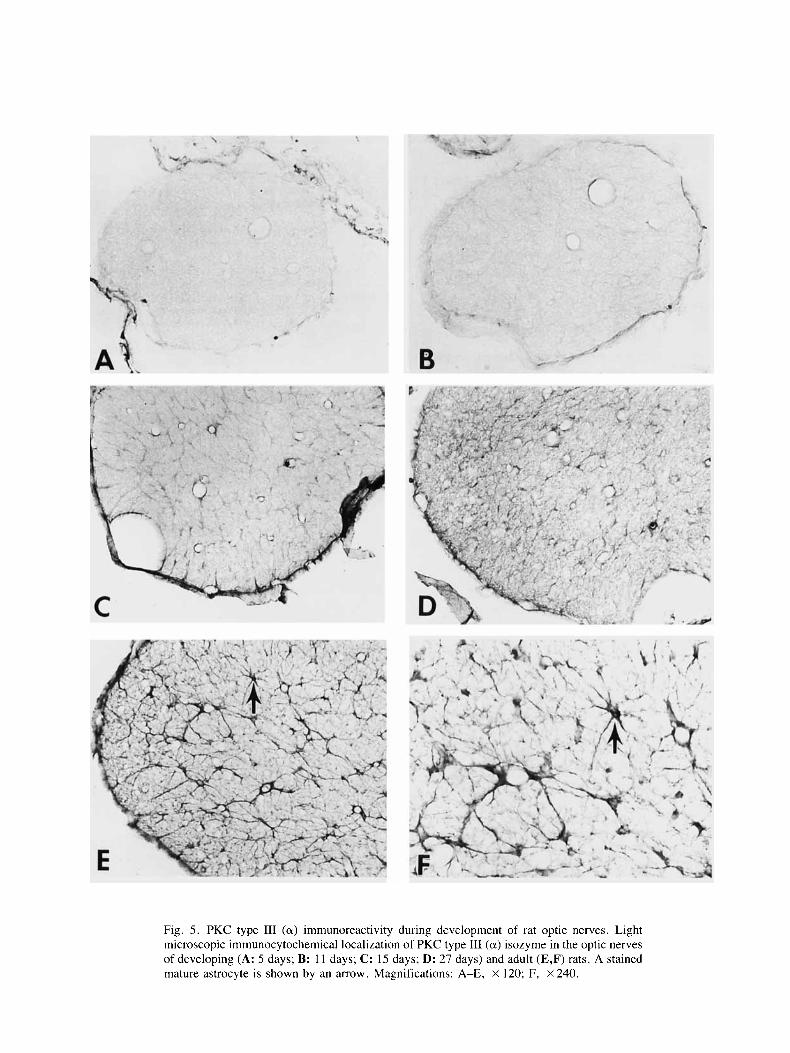
Fig. 5 . PKC type I11 (a) immunoreactivity during development of rat optic nerves. Light microscopic immunocytochemical localization of PKC type 111 (a) isozyme in the optic nerves of developing (A: 5 days; B: 11 days; C: 15 days; D: 27 days) and adult (E,F) rats. A stained mature astrocyte is shown by an arrow. Magnifications: A-E, X 120; F, X 240.

388 Komoly et al.
sion and activation of different protein kinase C iso- zymes.
ACKNOWLEDGMENTS
We wish to thank Mrs. Yoong Chang for her ex- cellent technical assistance in the EM studies.
REFERENCES Bhat NR (1989): Role of protein kinase C in glial cell proliferation. J
Neurosci Res 22:20-27. Burgoyne RD (1989): A role of membrane-inserted protein kinase C in
cellular memory? Trends Biol Sci 14:87-88. Chan, K-FJ (1987) Ganglioside-modulated protein phosphorylation in
myelin. J Biol Chem 262:2415-2422. Chan, K-FJ (1989) Effects of gangliosides on protein phosphorylation
in rat brain myelin. Neurosci Res Commun 5:95-104. Chiarugi VP, Ruggiero M, Corradetti R (1989): Oncogenes, protein
kinase C, neuronal differentiation and memory. Neurochem Int
Coussens L, Parker PJ, Rhee L, Yang-Feng TL, Chen E, Waterfield D, Francke U, Ullrich A (1986): Multiple, distinct forms of bovine and human protein kinase C suggest diversity in cellular signaling pathways. Science 233:859-866.
Girard PR, Mazzei GJ, Kuo JF (1986): Immunological quantitation of phospholipidiCa’ -dependent protein kinase and its fragments. J Biol Chem 261:370-375.
Girard PR, Wood JG, Freshci JE, Kuo JF (1988): Immunocytochem- ical localization of protein kinase C in developing brain tissue and in primary neuronal cultures. Dev Biol 126:98-107.
Harrison BC, Mobley PL (1989): Protein phosphorylation in astro- cytes mediated by protein kinase C: Comparison with phos- phorylation by cyclic AMP-dependent protein kinase. J Neu- rochem 53: 1245 -1 25 I .
Hidaka H, Tanaka T, Onada K, Hagiwara M, Watanabe M, Ohta H, Ito Y , Tsurudome M, Yoshida T (1988): Cell type specific expression of protein kinase C isozymes in the rabbit cerebel- lum. J Biol Chem 263:4523-4526.
Huang, K-P (1989): The mechanism of protein kinase C activation. Trends Neurosci 12:424-432.
Huang K-P, Huang FL ( 1986): lmmunochemical characterization of rat brain protein kinase C. J Biol Chem 261:14781-14787.
Huang K-P, Chan K-FJ, Singh TJ, Nakabayashi H, Huang FL (1986): Autophosphorylation of rat brain Ca2+-activated and phospho- lipid dependent protein kinase. J Biol Chem 261:12134- 12140.
Huang K-P, Huang FL, Nakabayashi H, Yoshida Y (1989): Expres- sion and function of protein kinase C isozymes. Acta Endo- crinol 12 1 :307-3 16.
Joo F, Tosaki A, Olah 2, Koltai M (1989): Inhibition by H-7 of protein kinase C prevents formation of brain edema in Sprague- Dawley CFY rats. Brain Res 490:141-143.
Kaczmarek, L (1987) The role of protei kinase C: in the regulation of ion channels and neurotransmitter release. Trends Neurosci 10: 30-34.
Kikkawa U , Nishizuka Y (1986): Protein kinase C. The Enzymes 17: 167-189.
Kikkawa U, Kishimoto A, Nishizuka Y (1989): The protein kinase C family: Heterogeneity and its implications. Annu Rev Biochem
1411-9.
58131-44.
C (1988): Electron microscopic localization of type I protein kinase C in rat Purkinje cells. J Neuroscience 8:4262-4268.
Louis JC, Magal E, Yavin E (1988): Protein kinase C alterations in the fetal rat brain after global ischemia. J Biol Chem 36:19282- 19285.
Mobley PL, Scott SL, CNZ EG (1986): Protein kinase C in astrocytes: A determinant of cell morphology. Brain Res 398:366-369.
Mochly-Rosen D, Basbaum AI, Koshlan DE (1987): Distinct cellular and regional localization of immunoreactive protein kinase C in rat brain. Proc Natl Acad Sci USA 84:4660-4664.
Neary JT, Norenberg LOB, Norenberg MD (1986): Calcium- activated, phospholipid-dependent protein kinase and protein substrates in primary cultures of astrocytes. Brain Res 385:
Neary JT, Norenberg LOB, Norenberg MD (1988): Protein kinase C in primary astrocyte cultures: cytoplasmic localization and translocation by phorbol ester. J Neurochem 50:1179-1184.
Nishizuka Y (1986): Studies and perspectives of protein kinase C. Science 233:305-3 12.
Nishizuka Y (1988): The molecular heterogeneity of protein kinase C and its implications for cellular regulation. Nature (London) 334:661-664.
Nixon RA, Lewis SE, Martotta CA (1987): Posttranslational modifi- cation of neurofilament proteins by phosphate during axoplas- mic transport in retinal ganglion cell neurons. J Neurosci 7: 1145-1 158.
Noguchi A, DeGuire J , Zanaboni P (1988): Protein kinase C in the developing rat liver, heart and brain. Dev Pharmaol Ther 11 : 31-43.
Olah 2, Ikeda J , Anderson WB, Joo F (1990): Altered protein kinase C activity in different subfields of hippocampus following ce- rebral ischemia. Neurochem Res 15:515-518.
Pestronk A, Watson DF, Yuan CM (1990): Neurofilament phosphor- ylation in peripheral nerve: Changes with axonal length and growth state. J Neurochem 54:977-982.
Peters A, Palay SL, Webster HdeF (1991): “The Fine Structure of the Nervous System. ” New York: Oxford University Press.
Pollenz R, McCarthy KD (1985): Regulation of intermediate filament protein phosphorykation in astroglia. Trans Amer Soc Neuro- chem 16:188.
Ritchie T, Cole R, Kim H-M, de Vellis J , Noble EP (1987): Inositol phospholipid hydrolysis in cultured astrocytes and oligodendro- cytes. Life Sci 41:31-39.
Saito N, Kikkawa U , Nishizuka Y , Yanaka C (1988): Distribution of protein kinase C-like immunoreactive neurons in rat brain. J Neurosci 8:369-382.
Saito N , Kose A, Ito H, Hosoda K, Mori M, Hirata M, Ogita K, Kikkawa U, Ono Y, Igarashi K , Nishizuka Y, Tanaka C ( I 989): lmmunocytochemical localization of PI1 subspecies of protein kinase C in rat brain. Proc Natl Acad Sci USA 86: 3409-341 3.
Shimosawa S, Hachiya T, Hagiwara M, Usuda N, Sugita K, Hidaka H (1990): Type-specific expression of protein kinase C isozymes in CNS tumor cells. Neurosci Lett 108: 11-16.
Sihag RK, Jeng AY, Nixon RA (1988): Phosphorylation of neurofil- ament proteins by protein kinase C. FEBS Lett 233:181-185.
Skoff RO, Price LD, Stocks A (1976) Electron microscopic autorad- iographic studies of gliogenesis in rat optic nerve. J Comp Neurol 169:3 13-334.
Sposi NM, Bottero L, Cossu G, Russo G , Testa U, Peschle C (1989): Expression of protein kinase C genes during otogenic develop- ment of the central nervous system. Mol Cell Biol 9:2284- 2288.
420-424.
Kose A, Saito N , Ito H, Hiroshi I, Kikkawa U, Nishizuka Y, Tanaka Stichel CC (1988): Ontogenetie changes in the level and subcellular

distribution of protein kinase C in cat visual cortex. Int J Dev Neurosci 6:341-349.
Vartanian T, Szuchet S, Dawson G, Campagnoni AT (1986): Oligo- dendrocyte adhesion activates protein kinase C-mediated phos- phorylation of myelin basic protein. Science 234: 1395-1398.
Yong VW, Sekiguchi S, Kim MW, Kim SU (1988): Phorbol ester enhances morphological differentiation of oligodendrocytes in culture. J Neurosci Res 19:187-194.
Yoshida Y, Huang FL, Nakabayashi H, Huang K-P (1988): Tissue
Protein Kinase C Isozymes in Optic Nerve 389
distribution and developmental expression of protein kinase C isozymes. J Biol Chem 263:9868-9873.
Young 111 SW (1988): Expression of three (and a putative four) protein kinase C genes in brains of rat and rabbit. J Chem Neuroanat
Young 111 SW (1989): Levels of transcripts encoding a member of the protein kinase C family in the paraventricular and supraoptic nuclei are increased by hyperosmolarity . J Neuroendocrinol
1 : 177-1 94.
1:79-82.