dissolved organic matter in lakes - diva...
TRANSCRIPT

ACTAUNIVERSITATIS
UPSALIENSISUPPSALA
2017
Digital Comprehensive Summaries of Uppsala Dissertationsfrom the Faculty of Science and Technology 1486
Dissolved organic matter in lakes
Chemical diversity and continuum of reactivity
ALINA MOSTOVAYA
ISSN 1651-6214ISBN 978-91-554-9840-5urn:nbn:se:uu:diva-316893

Dissertation presented at Uppsala University to be publicly examined in Friessalen, EBC,Norbyvägen 14, Uppsala, Friday, 28 April 2017 at 09:15 for the degree of Doctor ofPhilosophy. The examination will be conducted in English. Faculty examiner: AssociateProfessor Colin Stedmon (Technical University of Denmark).
AbstractMostovaya, A. 2017. Dissolved organic matter in lakes. Chemical diversity and continuumof reactivity. Digital Comprehensive Summaries of Uppsala Dissertations from theFaculty of Science and Technology 1486. 44 pp. uppsala: Acta Universitatis Upsaliensis.ISBN 978-91-554-9840-5.
Dissolved organic matter (DOM) is the largest pool of organic carbon in aquatic systems and animportant component of the global carbon cycle. Large amounts of DOM are decomposed withinlakes, resulting in fluxes of CO2 and CH4 to the atmosphere. Therefore, there is a considerableinterest in understanding the controls of DOM decomposition in freshwaters. There is evidencethat in lakes intrinsic controls related to DOM composition are of primary importance, yet ourknowledge about molecular drivers of DOM degradation is limited. This thesis addresses thelink between chemical composition and reactivity of lake DOM by applying an experimentalapproach, molecular-level DOM characterization, and kinetic modeling of DOM decay.
The first study shows that photoinduced transformations and partial removal of coloredaromatic components of DOM have profound effects on DOM degradation kinetics, mediatedby the shifts in the relative share of rapidly and slowly degrading DOM fractions. Twofollowing studies estimate exponential decay coefficients for each individual molecular formulaidentified within bulk DOM. A continuous distribution of exponential decay coefficients isfound within bulk DOM, which directly corroborates the central and previously empiricallyuntested assumption behind the reactivity continuum model of DOM decay. Further, individualdecay rates are evaluated in connection to specific molecular properties. On average, highlyunsaturated and phenolic compounds appear to be more persistent than compounds with higheraromatic content (plant polyphenols and polycondensed aromatics), and aliphatic compoundsdemonstrate the highest decay rates. The reactivity of aromatics additionally increaseswith increasing nominal oxidation state of carbon. Molecular analysis further indicates thatincreasing reactivity of DOM after UV exposure is caused by disintegration of supramolecularcomplexes. Study IV shows that changes in relative proportion of terrestrial versus algalDOM control degradability of DOM through seasons. Under ice, when algal-derived DOM ismaximally depleted, DOM degradation potential converges to similarly low levels, regardless oflake type (productive or humic), and bacterial respiration primarily relies on terrestrial carbon.This suggests a general pattern of baseline metabolism across boreal lakes. I conclude thatDOM is a dynamic reactivity continuum and a tight link exists between DOM behavior andcompositional properties.
Keywords: dissolved organic matter, DOM, dissolved organic carbon, DOC, PARAFAC,reactivity continuum model, ultrahigh resolution mass spectrometry, FT-ICR-MS, organicmatter characterization, decomposition kinetics, baseline metabolism
Alina Mostovaya, Department of Ecology and Genetics, Limnology, Norbyv 18 D, UppsalaUniversity, SE-75236 Uppsala, Sweden.
© Alina Mostovaya 2017
ISSN 1651-6214ISBN 978-91-554-9840-5urn:nbn:se:uu:diva-316893 (http://urn.kb.se/resolve?urn=urn:nbn:se:uu:diva-316893)

“They worked in an institute that was concerned first and foremost with the problems of human happiness and the meaning of human life, but even in their ranks there was no one who knew for certain what happiness is and what exactly is the meaning of life. And they had accepted as a working hypothesis that happiness lies in the constant cognition of the unknown, which is also the meaning of life.”
- Strugatsky brothers
To my mom


List of Papers
This thesis is based on the following papers, which are referred to in the text by their Roman numerals.
I Mostovaya, A., Koehler, B., Guillemette, F., Brunberg, A.-K., and Tranvik, L. J. (2016) Effects of compositional changes on reactivity continuum and decomposition kinetics of lake dissolved organic matter. J. Geophys. Res. Biogeosciences, 121(7), 1733–1746.
II Mostovaya, A., Hawkes, J. A., Koehler, B., Dittmar, T., and Tranvik, L. J. (2017) The emergence of the reactivity continuum of organic matter from kinetics of a multitude of individual molecular constituents. Manuscript.
III Mostovaya, A., Hawkes, Dittmar, T., and Tranvik, L. J. (2017). Molecular determinants of dissolved organic matter reactivity in lake water. Manuscript.
IV Guillemette, F., Mostovaya, A., and Tranvik, L. J. (2017) Seasonal variability in dissolved organic carbon degradation in boreal lakes: links to composition, sources, and baseline metabolism. Manuscript.
Reprints were made with permission from John Wiley and sons.


Contents
Introduction ................................................................................................... 11Dissolved organic matter in lakes ............................................................ 11Reactivity continuum of organic matter ................................................... 12Approaches to DOM characterization ...................................................... 13
Aims of the thesis .......................................................................................... 15Methods......................................................................................................... 16
Study lakes ............................................................................................... 16DOM decomposition experiments ........................................................... 16DOM characterization .............................................................................. 17
Concentration and isotopic signatures ................................................. 17Absorbance and fluorescence .............................................................. 18Ultrahigh resolution mass spectrometry .............................................. 18
Modeling of DOM decomposition ........................................................... 19Reactivity continuum modeling ........................................................... 19Exponential modeling of individual formula time series .................... 20
Statistical analysis and specific data representation ................................. 20Results and discussion .................................................................................. 21
How do kinetics of DOM degradation respond to alterations in specific fractions of lake DOM? (Papers I and IV) ............................................... 21Does behavior of individual molecular constituents support the main premise of the RC model, namely that there is a continuous spectrum of reactivity within bulk DOM? (Paper II) ................................................... 23What specific molecular properties influence the disappearance rate of lake DOM constituents? (Paper III) ......................................................... 24How does seasonality affect DOM composition and degradation potential in lakes of differing productivity and humic content, and what factors set the limits for baseline metabolism of DOM in lakes (Paper IV)? ............ 26
Conclusions and outlook ............................................................................... 29General Summary ......................................................................................... 31Summary in Swedish (Sammanfattning) ...................................................... 33Thank you ..................................................................................................... 36References ..................................................................................................... 40


Abbreviations
AImod Modified aromaticity index DBE Double bond equivalence DOC Dissolved organic carbon DOM Dissolved organic matter EEM Excitation-emission matrix ESI Electrospray ionization FT-ICR-MS Fourier-transform ion cyclotron resonance mass
spectrometry NOSC Nominal oxidation state of carbon OM Organic matter PARAFAC PCA
Parallel factor analysis Principal component analysis
RC model Reactivity continuum model SUVA Specific UV absorbance at 254 nm


11
Introduction
Dissolved organic matter in lakes Present-day limnology is to a large extent focused on carbon biogeochemistry. This particular focus emerged from a multitude of observations and estimates, all pointing toward a significant involvement of lakes in the global carbon cycling (Cole et al. 2007, Tranvik et al. 2009). The transformation of carbon in lakes is ultimately linked to organic matter (OM), produced internally by living organisms or delivered from soil via the terrestrial runoff. In aquatic environments OM exists predominantly in dissolved and colloidal form, and a collective term “dissolved organic matter” (DOM) is used to refer to this wide range of organic compounds.
Lake water DOM is extensively involved in ecological and biogeochemical processes. It is a primary energy source for heterotrophic bacteria and an important carbon subsidy for higher trophic levels (Azam et al. 1983, Tranvik 1992, Carpenter et al. 2005, Karlsson et al. 2012). Humic organic matter of terrestrial origin encompasses structures with light-absorbing properties and compounds with acidic functional groups, which has implications for the light regime and acid-base chemistry in lakes (Lydersen 1998, Perdue and Ritchie 2003, Wetzel 2003). Due to its binding and coagulation properties DOM can influence the toxicity of metals and availability of nutrients (Perdue 1998). From the perspective of ecosystem services, high concentrations of DOM in lake water pose a challenge for drinking water treatment (Lavonen et al. 2013).
Although globally lakes occupy only 3.7% of the non-glaciated land area (Verpoorter et al. 2014), they represent disproportionally active sites for organic carbon processing (Cole et al. 2007). Among the major carbon processing pathways are microbial and photochemical mineralization of DOM, and sedimentation with subsequent storage. Boreal lakes receive particularly large amounts of allochthonous (terrestrially-derived) DOM. Despite its relative resistance to biodegradation, this terrigenous DOM is an essential resource for heterotrophic microorganisms (Hessen 1992, Tranvik 1992). As a result, most boreal lakes are net-heterotrophic, and emit substantial amounts of carbon dioxide and methane to the atmosphere (Duarte and Prairie 2005, Bastviken et al. 2011, Raymond et al. 2013).
While in the recent decades our knowledge about lake DOM advanced to a great extent, the specific controls of DOM decomposition still require

12
clarification. Heavy dependency of OM degradation on extrinsic factors has been demonstrated in soils (Schmidt et al. 2011), but in freshwaters intrinsic properties of DOM, such as molecular composition and DOM size, appear to dominate over extrinsic factors (Kellerman et al. 2015a). The composition of lake water DOM is highly diverse, and it is difficult to identify specific intrinsic drivers of DOM degradation. However, with appropriate analytical methods for DOM characterization and a realistic vision of OM decay, our understanding of the link between composition and decay of DOM can be improved.
Reactivity continuum of organic matter Microbial decomposition of OM involves hydrolysis of OM constituents by bacterial extracellular enzymes and a subsequent uptake and metabolism of hydrolysis products. However, different compounds are hydrolyzed, taken up, and metabolized by bacteria with unequal efficiency (Arnosti 2011). For example, lake water compounds produced by autotrophs, such as amino acids, carbohydrates, and low molecular weight carboxylic acids, can be mineralized within hours or days (Bertilsson and Tranvik 1998). In contrast, vascular plant-derived components of allochthonous origin, such as some lignin constituents, can resist biological degradation for weeks and years. It has been abundantly shown that different fractions of OM degrade at different rates (e.g., Westrich and Berner 1984, Amon and Benner 1996, Guillemette and del Giorgio 2011) and, for realistic description and modeling of OM decay, this fact needs to be taken into account.
Early attempts to describe OM decomposition presented OM as a homogenous pool degrading following apparent first-order kinetics (linear or exponential decay) (Olson 1963, Berner 1964). However, a growing number of observations contradicted this view (e.g., Skopintsev 1981, Pelet 1984) and discrete multi-exponential models were suggested as an alternative. In the frame of this new approach, the OM was conceptually divided into two or three pools of differing and constant reactivity (Westrich and Berner 1984), and OM decay was described as a sum of pool-specific exponential decay functions. However, since it was nearly impossible to assess the amount and reactivity of each conceptual pool empirically, and a discrete low number of pools appeared conceptually unrealistic, these models were later criticized (e.g., Middelburg 1989, Koehler and Tranvik 2015).
A more realistic approach was offered by the theory of a reactivity continuum, which views OM as a continuous spectrum of reactive constituents ranging from fast to slowly decaying (Carpenter 1981, Boudreau and Ruddick 1991, Bosatta and Ågren 1995). The corresponding statistical reactivity continuum model (RC model) assumes an infinite number of reactive components within OM, with each component

13
undergoing a first-order exponential decay. The mass loss of OM over time is calculated as an integral of the exponential decay functions, weighted by an initial probability distribution of reactivity (Boudreau and Ruddick 1991). Assuming that the probability distribution of reactivity follows a gamma distribution (Boudreau and Ruddick 1991), the RC model has only two fit parameters. The apparent decay coefficient k can be calculated at each time point. Accordingly, the RC model allows calculating the decrease in OM reactivity over time.
A number of studies have advocated the benefits of the RC model for a simple and realistic description of OM decomposition (e.g., Vähätalo et al. 2010; Koehler and Tranvik 2015). The RC model is computationally simple and, unlike its predecessors, accounts for the heterogeneity of OM. Therefore, the RC model could be a useful tool for understanding the controllers of OM decay. Moreover, the assumptions of the RC model remain so far empirically untested, which in itself is an interesting angle for a study. Specifically, it has not been directly shown before that individual DOM constituents undergo a first-order exponential decay and form a continuous spectrum of decay rates within bulk DOM.
Approaches to DOM characterization Extreme chemical complexity of DOM poses large difficulties for characterization of DOM structure, properties, and source. However, a considerable interest in understanding the links between the quality of DOM and its fate in aquatic systems prompted the advent of powerful techniques for DOM characterization. Two major groups of methods, optical characterization techniques, and ultrahigh resolution mass spectrometry, are discussed below.
Optical methods, including absorbance and fluorescence spectroscopy rely on light-absorbing properties of DOM. These are effective and accessible techniques, useful for characterization of aromatic content (e.g., specific UV absorbance SUVA, Weishaar et al. 2003; fluorescence index FI, McKnight et al. 2001), molecular weight (e.g., absorption spectral slopes, Helms et al. 2008), as well as the source and relative abundances of aromatic versus aliphatic, or humic versus protein-like fractions of DOM (various fluorescence indexes and specific fluorescent components; Fellman et al. 2010; Stedmon and Markager 2005). At present, optical techniques remain highly useful for describing and comparing quality of DOM, but have rarely been used in combination with the RC model of OM decay. While giving an idea of OM characteristics, absorbance and fluorescence do not provide detailed information about the molecules underpinning the optical properties. This gap can be bridged by ultrahigh resolution analytical techniques capable of characterizing DOM at the molecular level.

14
The applications of ultrahigh resolution mass spectrometry span across a broad field of sciences that require detailed analysis of complex environmental mixtures. Fourier transform ion cyclotron resonance mass spectrometry (FT-ICR-MS) coupled with soft ionization methods (electrospray ionization, ESI) is capable of resolving composition of DOM down to the level of individual molecular masses. FT-ICR-MS can simultaneously detect thousands of molecular masses in the sample with the accuracy of 0.1 mDa (Kim et al. 2006), and molecular formula can be assigned to each mass with good confidence (Stenson et al. 2003, Koch et al. 2005, 2007). The structural information about the detected compounds (saturation state, the oxidation state of carbon, presence of aromatic structures, etc.) can be further deduced based on their molecular formulas (Koch and Dittmar 2006, Riedel et al. 2012, Šantl-Temkiv et al. 2013).
FT-ICR-MS is extensively used for comparing the DOM between study sites (e.g., Koch et al. 2005, Kellerman et al. 2014), and untangling the effects of biological and photochemical transformations of DOM (e.g., Stubbins et al. 2010, Riedel et al. 2016). Coupling the information of molecular characteristics of DOM with the assessment of DOM reactivity can further provide important insights into the intrinsic controllers of DOM decomposition in lake water.

15
Aims of the thesis
In this PhD project, I looked into fundamental but hitherto rarely addressed patterns in the dynamics of natural organic matter, specifically focusing on a link between chemical composition and degradation kinetics of lake DOM. The papers included in this thesis aimed to address the following questions:
• How do kinetics of DOM degradation respond to alterations in specific fractions of lake DOM? (Papers I and IV)
• Does behavior of individual molecular constituents support the main premise of the RC model, namely that there is a continuous spectrum of reactivity within bulk DOM? (Paper II)
• What specific molecular properties influence the disappearance rate of lake DOM constituents? (Paper III)
• How does seasonality affect DOM composition and degradation potential in lakes of differing productivity and humic content, and what factors set the limits for baseline metabolism of DOM in lakes? (Paper IV)

16
Methods
Study lakes For the first experiment (Papers I-III) water was sampled from three brown-water lakes, similar in dissolved organic carbon (DOC)1 concentration. For the second experiment (Paper IV) water was sampled from four lakes differing in DOC concentration, water color, and productivity. All sampled lakes are located in south-central Sweden. The catchments are underlain by calcareous moraine and granitic bedrock, and dominated by coniferous forest. Water for the experiment in papers I-III was sampled in October 2013, and water for the experiment presented in paper IV was sampled once every month from February to July 2013.
DOM decomposition experiments The DOM decomposition experiments were carried out on filtered water with addition of a small inoculum (plankton net filtered aliquot from the respective lake, 5 or 1% of the volume). Subsamples for optical analysis were taken before the start of incubations. The water was incubated in sealed 40 mL vials, at 20°C and in the dark. Over several time intervals the vials were sacrificed to measure DOC. Subsamples for mass spectrometry analysis (papers II, III) were taken at the same time.
Experiment I, discussed from different angles in papers I-III, was conducted for 120 days. A complete setup included four treatments: non-manipulated lake water (Control), lake water pretreated with UV light (UV treatment), lake water pretreated with polyvinylpyrrolidone (PVP treatment), and a combination of the last two (UV-PVP treatment) (Figure 1). Manipulations with UV and PVP were conducted prior to the start of dark incubation, to alter the composition of lake water DOM through phototransformation and removal of phenolic compounds, respectively. The details about phenol-binding properties of PVP are described elsewhere (Carpenter et al. 1976, Freeman et al. 2004).
1 DOM and DOC are often used as synonyms since DOC is a main fraction of DOM. Further in the text, the term DOC is used to refer to analytically measured concentration.

17
Figure 1. The outline of DOM decomposition experiment (Papers I-III). Lake water was filtered (0.2 µm), treated by UV light or/and polyvinylpyrrolidone (PVP) and, after addition of a small inoculum (5% of the volume), transferred to 40 mL vials. The incubations were performed for 120 days, at 20°C and in the dark. Complete design included 24 time series (3 lakes × 4 treatments × 2 replicates).
Experiment II (Paper IV) was performed on water from four lakes covering a gradient in DOC, productivity, and humic content, and new incubation series were added every month upon new sampling, in order to assess the change of degradation potential of DOM through seasons. The incubations were standardized (90 days at 20°C in the dark).
DOM characterization Concentration and isotopic signatures The concentration of DOC was measured on a Sievers 900 TOC (total organic carbon) analyzer (General Electric Analytical Instruments). Dissolved inorganic carbon (DIC) concentration and δ13C isotopic signature were determined on a modified Aurora 1030W TOC analyzer (OI Analytical) connected to a Finnigan MAT Delta Plus isotope ratio mass spectrometer with a ConFlo III interface (Thermo Finnigan).
Dark incubations
120 days, 20°C
DOC
FT-ICR-MS
subsample
Inoculated
PVP
DOC
FT-ICR-MS
subsample
DOC
DOC
Filtered
UV
UV
+
PVP
Treatments
Inoculated
Inoculated
Inoculated
Optical
analysis
Optical
analysis
Optical
analysis
Optical
analysis
Control
Lake
water

18
Absorbance and fluorescence Absorbance spectra were collected on filtered water across the range of 200–600 nm with a Lambda 40 UV-visible spectrophotometer (Perkin Elmer). Absorbance values were converted into Naperian absorption coefficients, a (m−1) (Kirk 1994). Colored DOM (CDOM) was represented either by integrating a in the range of 250–500 nm (Moran et al. 2000), or as a at 440 nm (a440). Specific UV absorbance (SUVA), indicating a degree of aromaticity, was calculated by dividing the absorbance at 254 nm (A254) by DOC concentration and optical path length (Weishaar et al. 2003). A slope ratio (SR), suggestive of molecular weight, was calculated as described in Helms et al. (2008). Ratio of absorbance at 365 and 250 nm was used as an indicator of molecular size (Peuravuori and Pihlaja 1997).
Fluorescence excitation emission matrices (EEMs) were generated on filtered water using a Fluoromax-4 spectrofluorometer (Horiba Jobin Yvon), with excitation measured at 260–450 nm with 5 nm increments and emission measured between 300–600 nm with 2 nm increments. The EEMs were blank-subtracted, corrected for instrument-specific biases (Cory et al. 2010) and inner filter effects (Kothawala et al. 2013), and normalized to Raman units (Lawaetz and Stedmon 2009).
The following indexes were calculated from the corrected EEMs: fluorescence index (used to distinguish between microbial and terrestrial sources of DOM; Cory et al. 2010, McKnight et al. 2001), freshness index (indicates the contribution of recently produced DOM; Parlanti et al. 2000, Fellman et al. 2010), and humification index (an indicator of aromatic content in DOM; Zsolnay et al. 1999, Fellman et al. 2010).
The corrected EEMs were modeled with parallel factor analysis (PARAFAC) (Murphy et al. 2013) and six fluorescent components were identified by the analysis: four components associated with different types of humic-like fluorescence, one microbial humic-like component, and one tryptophan-like component.
Ultrahigh resolution mass spectrometry In the experiment discussed in papers II and III samples for mass spectrometry analysis were taken at 12 different time points from non-manipulated and UV-manipulated treatments (Figure 1). Lake water samples were analyzed on a 15-Tesla Fourier transform ion cyclotron resonance mass spectrometer (FT-ICR-MS; Bruker Daltonics) coupled to an electrospray ionization source operating in negative mode. The samples were diluted with ultrapure water to a concentration of 10 mg C L−1 and mixed with methanol in a proportion of 2:1 (final concentration 6.7 mg C L−1). Mass spectra were collected within a range of 150–2000 m/z. A reference sample of North

19
Equatorial Pacific Intermediate Water (NEqPIW) (Green et al. 2014) was run at the beginning and end of each day of analysis.
Only peaks with a signal to noise ratio greater than four were considered. Molecular formulas were assigned based on the following criteria: C ≥ O; O > (2P + S); N ≤ 4; S ≤ 4 and P ≤ 1. Contaminant peaks and peaks detected in procedural blanks were removed. S-containing, P-containing, and N>1-containing peaks were not included in our studies. Detection limit was applied based on dynamic range (see paper II for details). In each sample the intensities were normalized to the total intensity and multiplied to account for dilution, i.e., to get an estimate of the intensity in the original undiluted sample.
Atomic H/C and O/C ratios, double bond equivalence and modified aromaticity index (DBE and AImod; Koch and Dittmar 2006), and nominal oxidation state of carbon (NOSC; Riedel 2012) were calculated for each molecular formula. Compound groups were outlined based on AImod and H/C ratio, as polycondensed aromatics (AImod>0.66; Koch and Dittmar 2006), plant polyphenols (0.50<AImod≤0.66; Kellerman et al. 2015a), highly unsaturated and phenolic compounds (AImod≤0.5 & H/C<1.5; Šantl-Temkiv et al. 2013), and aliphatics (H/C≥1.5; Riedel et al. 2016).
Modeling of DOM decomposition Reactivity continuum modeling Decomposition of DOM was modeled with the gamma reactivity continuum model (RC model). The RC model assumes an infinite number of reactive species within bulk DOM, characterized by a continuous gamma distribution of decay coefficients k (each individual compound is assumed to follow single pool exponential decay). The time series of DOC mass loss were modeled as:
DOCtDOC0
= aa + t
⎛⎝⎜
⎞⎠⎟
v
where a and v are rate and shape parameters of the simulated gamma distribution of the initial decay coefficients k, respectively (Boudreau and Ruddick 1991, Koehler et al. 2012). The apparent decay coefficient kt (day−1) can be calculated for any time point as:
kt =
va + t
(Boudreau et al. 2008). Accordingly, v/a gives the initial apparent first-order decay coefficient k0 (expected value of the initial probability distribution of

20
reactivity), and the sequence of kt simulates a continuous decrease in reactivity over time.
Exponential modeling of individual formula time series Provided that the sample matrix is similar, the signal intensity of each molecular mass is essentially linearly proportional to the concentration of the corresponding isomeric mixture of compounds (Seidel et al. 2015). This fact allowed us to model mass spectrometry data according to the assumption of the RC model that each individual compound undergoes single exponential decay (Boudreau and Ruddick 1991). The time series of formula-specific intensities were modeled as:
ItI0
= e−kexpt
where kexp is an exponential decay coefficient (day−1) and It/I0 is the change in intensity over time (Olson 1963).
Statistical analysis and specific data representation For paper I statistical analysis of the effects of pretreatments (UV, PVP) on initial conditions of incubations (DOC concentration, optical characteristics) was carried out using linear mixed effect models, with treatment set as fixed effect and lake set as a random effect.
For papers I and II, the initial probability distribution of decay coefficients was presented using cumulative probability plots. These plots convey information that cannot be retrieved from otherwise more intuitive density plots or histograms, and allow evaluating the proportion of DOM associated with a specific range of initial decay coefficients k (see papers I and II for the details).
In paper III, the association of exponential decay coefficients with the groups of chemical compounds was visualized on van Krevelen diagrams in which each molecule is displayed in a plot of the atomic ratios – H/C versus O/C (Kim et al. 2003).
In paper IV, principal component analysis (PCA) was used to explore the variability in DOM characteristics (optical parameters, composition-related indexes) across seasons and trophic status of the sampled lakes. The Keeling plot method was used to evaluate the sources of respired carbon. Bacterial respiration adds CO2 of a given δ13C signature to the background DIC pool over the course of an incubation, and a linear relation exists between δ13C and the inverse DIC concentration where the intercept corresponds to isotopic signature of respired CO2 (Karlsson et al. 2007).

21
Results and discussion
How do kinetics of DOM degradation respond to alterations in specific fractions of lake DOM? (Papers I and IV) Extreme complexity of natural OM and the diversity of its interactions with the environment make it difficult to identify the principal drivers of OM degradation. The decisive role of extrinsic controls has been demonstrated in soils (Schmidt et al. 2011), but in aquatic environments composition and size of OM have been identified as primary determinants of OM fate (Kellerman et al. 2015a, Walker et al. 2016). The ideas about degradability of different fractions of freshwater DOM are nevertheless conflicting, and our understanding of how changes in DOM quality translate into changes in decomposition rates is incomplete. For example, colored aromatic compounds have long been considered the most refractory component of freshwater DOM (Tranvik 1992), but recently it was shown that colored aromatic DOM can be preferentially removed on longer time scales (Koehler et al. 2012, Weyhenmeyer et al. 2012, Kellerman et al. 2015a). Further, the DOM photoproducts are often assumed to be rather short-lived (hours-days) (Wetzel et al. 1995, Bertilsson and Tranvik 1998) and not many studies have explored whether photoproduced DOM can persist for longer time (e.g., months).
In papers I and IV we show that changes in DOM composition, whether experimentally-induced or seasonal, have profound effects on DOM decomposition kinetics.
In paper I, we manipulated DOM composition by exposing lake water to artificial UV light, and by removing phenolic compounds with PVP. While certain fractions of DOM were completely lost as a result of these pretreatments, optical analysis suggested that in the UV treatment aromatic compounds were also transformed into more biodegradable smaller fragments. Optical parameters additionally indicated that PVP effectively removed colored aromatic DOM, but did not exclusively target high molecular weight fraction of these compounds.
UV exposure resulted in significant increase in the initial decay coefficient k0, compared to non-manipulated control (Figure 2). Moreover, tracing the decrease in reactivity over time showed that the stimulating effect
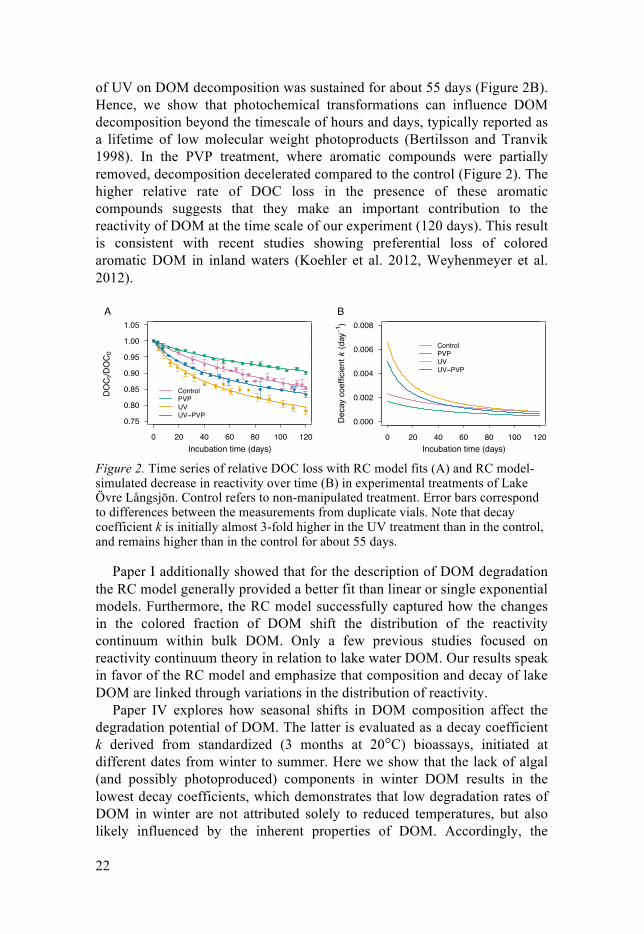
22
of UV on DOM decomposition was sustained for about 55 days (Figure 2B). Hence, we show that photochemical transformations can influence DOM decomposition beyond the timescale of hours and days, typically reported as a lifetime of low molecular weight photoproducts (Bertilsson and Tranvik 1998). In the PVP treatment, where aromatic compounds were partially removed, decomposition decelerated compared to the control (Figure 2). The higher relative rate of DOC loss in the presence of these aromatic compounds suggests that they make an important contribution to the reactivity of DOM at the time scale of our experiment (120 days). This result is consistent with recent studies showing preferential loss of colored aromatic DOM in inland waters (Koehler et al. 2012, Weyhenmeyer et al. 2012).
Figure 2. Time series of relative DOC loss with RC model fits (A) and RC model-simulated decrease in reactivity over time (B) in experimental treatments of Lake Övre Långsjön. Control refers to non-manipulated treatment. Error bars correspond to differences between the measurements from duplicate vials. Note that decay coefficient k is initially almost 3-fold higher in the UV treatment than in the control, and remains higher than in the control for about 55 days.
Paper I additionally showed that for the description of DOM degradation the RC model generally provided a better fit than linear or single exponential models. Furthermore, the RC model successfully captured how the changes in the colored fraction of DOM shift the distribution of the reactivity continuum within bulk DOM. Only a few previous studies focused on reactivity continuum theory in relation to lake water DOM. Our results speak in favor of the RC model and emphasize that composition and decay of lake DOM are linked through variations in the distribution of reactivity.
Paper IV explores how seasonal shifts in DOM composition affect the degradation potential of DOM. The latter is evaluated as a decay coefficient k derived from standardized (3 months at 20°C) bioassays, initiated at different dates from winter to summer. Here we show that the lack of algal (and possibly photoproduced) components in winter DOM results in the lowest decay coefficients, which demonstrates that low degradation rates of DOM in winter are not attributed solely to reduced temperatures, but also likely influenced by the inherent properties of DOM. Accordingly, the
●
●
●
●
●● ●
● ●
● ●
●● ●
●●
●● ●
●
● ●●
●● ●
●●
●● ●
● ● ●● ●
●●
●
●
●●
●
●
●●
●●
●
● ● ●
● ●
●●
●
●●
●
●
●●
●
● ● ●
● ● ●
● ●● ●
●Control
PVP
UV
UV−PVP
0 20 40 60 80 100 120
0.75
0.80
0.85
0.90
0.95
1.00
1.05
Incubation time (days)
DO
Ct/D
OC
0
0 20 40 60 80 100 120
0.000
0.002
0.004
0.006
0.008
De
cay c
oe
ffic
ien
t k
(day
−1)
Control
PVP
UV
UV−PVP
A B
Incubation time (days)

23
winter DOM pool was dominated by high molecular weight and aromatic fractions as compared to summer where fresher and more aliphatic compounds were found.
Does behavior of individual molecular constituents support the main premise of the RC model, namely that there is a continuous spectrum of reactivity within bulk DOM? (Paper II) Single-pool exponential and discrete multiexponential models of OM decomposition imply that OM consists of a limited number of homogenous kinetic pools (typically, 1–3). At the same time, high resolution analytical techniques show that OM consists of thousands of compounds of different structure (Koch and Dittmar 2006, Minor et al. 2012). Such diversity in structure suggests the diversity in chemical properties that goes beyond a few kinetic pools. To overcome the unrealistic visions of OM complexity, OM was conceptualized as a continuous spectrum of compounds of differing reactivity (Carpenter 1981, Boudreau and Ruddick 1991, Bosatta and Ågren 1995). This concept and the reactivity continuum model of OM decay are in the focus of paper II.
Paper II discusses the fundamentals of organic matter decomposition and investigates whether reactivity continuum theory is supported by the empirical data on rates of decomposition of individual molecular constituents of the DOM. The study is based on a subset of DOM decomposition experiment described in detail in the methods section (Figure 1).
The RC theory proposes the existence of a continuous range of reactivities within bulk OM. Accordingly, the RC model assumes that each individual compound within bulk OM decays following a single-pool exponential model (Boudreau and Ruddick 1991). To get to the compound level, we combined ultrahigh resolution mass spectrometry with kinetic modeling. The signal intensity of each molecular mass is essentially linearly proportional to the concentration of the corresponding isomeric mixture of compounds (Seidel et al. 2015). Hence, for every molecular mass with a corresponding assigned formula, we fitted the single exponential model to changes in the signal intensity over decomposition time and acquired a decay coefficient kexp.
We show that individual molecular formulas within bulk DOM indeed produce a continuous range of decay coefficients (Figure 3), as predicted by the RC model. However, the range of empirically derived exponential decay coefficients (kexp) was narrower than the range simulated with the RC modeling of the bulk DOC mass loss. The reasons for this discrepancy are discussed in detail in paper II.

24
The expected effect of phototransformations on the reactivity continuum of DOM was clearly visible in the mass spectrometry derived data. Accordingly, the distribution of decay coefficients shifted toward higher values in the UV treatment (Figure 3). This suggests that, despite the possible method limitations, our approach to the datasets was reliable. Our results further demonstrate that the reactivity continuum of lake DOM emerges from a multitude of individual components of different reactivity, which supports the main premise of the RC model of OM decomposition.
Figure 3. Distribution of exponential decay coefficients within bulk DOM. The distributions are continuous, and UV-treatment distribution is shifted toward higher decay coefficients. The data shows the treatments of Lake Ramsjön.
What specific molecular properties influence the disappearance rate of lake DOM constituents? (Paper III) Intrinsic properties of DOM have been identified as important controllers of DOM persistence in lakes (Kellerman et al. 2015a). However, the exact relationship between DOM composition and reactivity still requires clarification. The observed behavior of bulk DOM is an integrated result of many multidirectional influences and isolating the major drivers of decomposition requires a thoughtful approach.
Paper III addresses molecular determinants of DOM reactivity in lake water, and is based on the subset of DOM decomposition experiment (Figure 1) which considers non-manipulated and UV-manipulated experimental treatments. Here, using ultrahigh resolution mass spectrometry data, we characterize molecular properties of lake DOM, and model formula-specific decay to link molecular characteristics to reactivity of DOM. The decay coefficient kexp was calculated for each formula, by fitting a single
0100200300400
0.000 0.002 0.004 0.006 0.008 0.010
Num
ber o
f mol
ecul
ar fo
rmul
as Non−manipulated lake water
0100200300400
0.000 0.002 0.004 0.006 0.008 0.010
UV−manipulated lake water
Exponential decay coefficient k

25
exponential model to the relative change in peak intensities over decomposition time (120 days).
We show that kexp forms a continuous distribution within each group of compounds, i.e., both fast and slowly degrading compounds were present in each group (Figure 4). The exact character of the distributions, however, differed between the groups (Figure 4). The group of highly unsaturated and phenolic compounds, likely representing the products of vascular plant decay, was dominated by the lowest kexp. Aliphatic compounds (H/C>1.5) demonstrated the highest reactivity. Compounds with increased aromatic content (plant polyphenols and polycondensed aromatics) showed intermediate reactivity in the non-manipulated treatment (Figure 4), and high reactivity in the UV-manipulated treatment. The fact that compounds with high aromatic content were on average more reactive than less aromatic compounds (highly unsaturated and phenolic) could be considered surprising since colored humic DOM is often associated with low decay rates (Tranvik 1998). Yet, this result is in line with the recent findings suggesting that colored terrestrial DOM is removed faster than bulk DOM on longer timescales (Koehler et al. 2012, Weyhenmeyer et al. 2012).
Further, in our study the disappearance rate of compounds with high aromatic content increased with increasing nominal oxidation state of carbon. This result shows a striking similarity to the conclusions of Kellerman et al. (2015a) who, using FT-ICR-MS analysis of DOM from 109 Swedish lakes, showed that oxidized aromatic DOM is preferentially removed with increasing water residence time of the sampled lakes. The role of oxygen content and carbon oxidation state in reactivity of DOM is still debated, and we encourage further studies of this complex subject.
Molecular mass did not influence kexp in the non-manipulated treatment, but was inversely related with kexp in the UV-manipulated treatment. Between 87.7 and 97.9% of formulas identified in non-manipulated treatment were also present in the UV-manipulated treatment. Notably, the
Figure 4. Kernel density plots showing the distributions of decay coefficients kexp within different groups of compounds and “bulk” DOM (all formulas). Kernel density function is a non-parametrically estimated probability density function of kexp. The integral of each function equals 1. The data corresponds to non-manipulated treatment of Lake Lumpen.
0.000 0.002 0.004 0.006
0
200
400
600
800D
ensity
kexp (day−1
)
Aliphatics
Highly unsaturated and
phenolic compounds
Plant polyphenols
Polycondensed aromatics
All formulas

26
kexp associated with these shared formulas was often higher in the UV-manipulated treatment. The increase in reactivity primarily occurred in the groups of plant polyphenols and polycondensed aromatics, and these groups had previously been linked to supramolecular DOM complexes of higher apparent molecular weight (Kellerman et al. 2015b). We propose that increase in reactivity was triggered by the disintegration of supramolecular complexes, and a resulting diminishing influence of hydrophobic interactions, multiple hydrogen bonds and steric hindrance.
The photoproduced compounds that were formed as a result of UV pretreatment were not restricted to a particular chemical group, and did not show a systematic trend in reactivity. This demonstrates that apart from commonly described aliphatic and highly reactive compounds (Moran and Zepp 1997), photoproducts can include slowly degrading DOM capable of supporting bacterial metabolism on longer timescales.
Finally, paper III demonstrates the advantages of using ultrahigh resolution mass spectrometry techniques in studies addressing the role of molecular properties in DOM dynamics in aquatic systems.
How does seasonality affect DOM composition and degradation potential in lakes of differing productivity and humic content, and what factors set the limits for baseline metabolism of DOM in lakes (Paper IV)? The fate of DOM in freshwater systems is strongly linked to DOM composition (Kellerman et al. 2015a). The composition, in turn, is governed by the source (Guillemette et al. 2013) and history of biogeochemical transformations (Riedel et al. 2016) of DOM. Through seasons, lake water DOM undergoes compositional shifts, yet comprehensive studies of the effects of seasonality on DOM degradation are rare.
In paper IV we used standardized (3 months at 20°C) bioassays to compare degradation potential of lake DOM in a temporal gradient from winter to summer, across four lakes of differing productivity and humic content. We found that in winter under ice conditions the degradation potential of DOM (measured as decay coefficient k) was similarly low among lakes (Figure 5). Further, using the ratio between chlorophyll a and colored DOM absorbance measurement (a440) as a proxy of algal versus terrestrial influence, we found that increasing share of algal DOM from ice-melt to ice-free conditions was accompanied by increasing DOM degradation potential, and this effect was stronger in more productive lakes.

27
Analysis of isotopic signatures (δ13C) showed that under ice conditions in March, the terrestrial contribution to respiration approached 100% in all lakes. Interestingly, just one month earlier in one of the more productive lakes up to 28% of respiration was still supported by autochthonous carbon. Considering that photosynthesis under ice is limited, this result emphasizes that not all autochthonous DOM is short lived: some of it can remain in lake water from summer through autumn and fuel slow decomposition under the ice. Toward the summer, reliance of respiration on terrestrial carbon was decreasing, and this decrease was especially steep in the productive lakes. Yet, even in the more productive lake Lilla Sångaren terrestrial carbon supported about half of microbial respiration in summer, suggesting that terrestrial DOM is important for microbial metabolism even in the presence of alternative, readily available energy sources.
While some general trends, such as transition toward more aromatic DOM in winter, existed among all lakes, each lake preserved its unique compositional identity through the seasons. This was suggested by the distinct separated positions of different lakes on the PCA plot constructed from optical indices and PARAFAC components as indicators of DOM composition.
Considering that the studied lakes covered a broad gradient of productivity and humic content, the similarity in degradation potential of winter DOM among the lakes is striking. We connect this result to the concept of baseline metabolism. While low temperatures are typically thought to be responsible for the slow metabolism in winter, we here show that DOM quality is another important factor constraining utilization of carbon. There is an idea that rapid intermittent metabolism of labile compounds is underlain by a continuous slow metabolism of longer-living substrates (Tranvik 1992, McCallister and del Giorgio 2008, Solomon et al. 2013, Guillemette et al. 2013). Our results agree with this idea and we show that as algal and fresh terrestrial DOM inputs become depleted, lakes
Figure 5. Seasonal variability in the initial decay coefficient k estimated using the reactivity continuum model of DOM decay in standardized (3 months at 20oC) lake water DOM incubation experiments. The open and filled symbols correspond to the clear water and brown water lakes, respectively (four lakes in total). The shaded area denotes the period of ice cover in winter.
k (d
ay−1
)
0.000
0.001
0.002
0.003
0.004
0.005
0.006
Feb Mar Apr May Jun Jul
Ice cover

28
transition to slower metabolism, and respiration increasingly relies on terrestrial substrates. The fact that the degradation potential of winter DOM is similar among the contrasting lakes suggests the presence of a relatively invariant core of slowly decaying compounds of primarily terrestrial origin.
Finally, our results accentuate the importance of sunlight in seasonal trends of DOM degradation dynamics. Both photosynthesis and photochemical transformation of DOM cease in winter, cutting off the production of algal DOM and formation of smaller labile DOM fragments from recalcitrant aromatic predecessors (Moran and Zepp 1997). Apart from the rapidly decaying components, both algal (see above) and phototransformed (see papers I-III) DOM includes compounds with longer life times. This DOM is likely to continue its slow decomposition through early winter, extending the effects of sunlight beyond the ice-free season.

29
Conclusions and outlook
The main focus of this thesis is the relationship between composition and reactivity of lake water DOM. The main conclusions are as follows:
• Kinetics of DOM degradation respond to alterations in specific fractions of lake DOM with the dynamic shifts of reactivity continuum toward higher or lower decay rates. This was exemplified through comparison of the effects of phototransformations and partial removal of colored aromatic DOM on overall DOM degradation kinetics.
• Behavior of individual molecular constituents supports the main premise of the reactivity continuum model, namely that there is a continuous spectrum of reactivity within bulk DOM. This was demonstrated by estimating exponential decay coefficients of numerous individual molecular formulas within bulk DOM.
• Average decay rates differ between the molecular compound groups: highly unsaturated and phenolic compounds are more persistent than compounds with higher aromatic content (plant polyphenols and polycondensed aromatics), and aliphatic compounds show the highest decay rates. For aromatic compounds, reactivity additionally increases with increasing nominal oxidation state of carbon. These results derive from direct estimations of individual decay rates.
• Increased reactivity of phototransformed DOM is likely caused by disintegration of supramolecular complexes, resulting in diminishing structural constraints on decomposition. The evidence came from the detailed comparison of formulas and corresponding decay rates in non-treated versus UV-treated lake DOM.
• During transition from ice-covered to ice-free conditions growing algal influence increases degradability of lake DOM, which is especially evident in more productive lakes. In winter degradability of DOM is similarly low among different lake types, suggesting a general pattern of baseline metabolism, almost entirely supported by terrestrial carbon.
With this thesis, I contribute to understanding of the mechanisms regulating decomposition of DOM in lakes and demonstrate that intrinsic properties of DOM are important determinants of DOM decay in freshwaters. An

30
important practical outcome of this work is the comprehensive demonstration of the RC model as a useful tool facilitating realistic description of DOM decomposition. Empirical evidence of a reactivity continuum existing within bulk DOM, and even within different DOM fractions, indicates that labeling either of these fractions as ubiquitously recalcitrant or labile can be misleading. Future efforts should be directed toward a better mechanistic understanding of the links between composition and reactivity. Given the advances in analytical chemistry and the growing access to ultrahigh resolution techniques for DOM characterization, this becomes increasingly possible to achieve. My thesis constitutes a first step along this path, for the first time combining ultrahigh resolution characterization of complex natural OM with a comprehensive experimental approach and realistic kinetic modeling of OM decay.

31
General Summary
Carbon is a building block of life, but it is even more abundant in dead organic matter. Remains of plants and animals in various stages of decay, as well as by-products of metabolism, constitute the largest pool of carbon in the biosphere. While in terrestrial ecosystems most of the dead organic matter accumulates in soils, in aquatic environments it is mostly present in dissolved and colloidal form, collectively referred to as dissolved organic matter, or DOM. The focus of my thesis is DOM in lakes.
Lake DOM is extensively involved in ecological and biogeochemical processes. It is a primary energy source for heterotrophic bacteria and an important carbon subsidy for higher trophic levels. It has potential to influence light regime, acid-base water chemistry, toxicity of metals, and availability of nutrients. Boreal lakes receive particularly large amounts of DOM via terrestrial runoff. Additionally, DOM can be produced internally by autotrophs. Some DOM sinks and remains in the sediments, while other fractions are decomposed by bacteria or broken down by sunlight, resulting in fluxes of CO2 and CH4 to the atmosphere. Globally lakes occupy only 3.7% of the non-glaciated land area, yet they emit large amounts of greenhouse gases. Therefore, there is a significant interest in identifying the main drivers of DOM decomposition in lakes. It was recently shown that molecular composition and size of DOM largely determine its fate in freshwaters. However, many aspects of the relationship between DOM composition and degradability remain unclear. A primary goal of my thesis was to improve our understanding of this relationship.
In my thesis, the links between DOM composition and decay are addressed in a series of DOM decomposition experiments. Optical methods, i.e., absorbance and fluorescence spectroscopy, are used to evaluate contribution of different fractions (e.g., aromatic versus aliphatic, high molecular weight versus low molecular weight) to bulk DOM. More detailed molecular information is provided by ultrahigh resolution mass spectrometry (ESI FT-ICR-MS), which resolves composition of DOM to the level of individual molecular formula. Further, a specific model (reactivity continuum (RC) model) is used to evaluate and describe DOM decay. The RC model acknowledges the complexity of DOM as a substrate, and views DOM decomposition as an integrated decay of an infinitely large number of compounds of differing reactivity.

32
Study I of this thesis shows that photoinduced transformations and partial removal of colored aromatic components of DOM have profound effects on DOM degradation kinetics, mediated by the shifts in the relative share of rapidly and slowly degrading DOM fractions. Additionally, this study demonstrates the advantages of the RC model as a simple and informative tool for the description of DOM degradation kinetics.
Study II addresses the decay rates of individual molecular formulas and focuses on the assumptions of the RC model. This study directly demonstrates that individual DOM constituents form a continuous distribution of exponential decay rates within bulk DOM. The results support the idea that overall DOM decomposition pattern is a result of integrated decay of a multitude of DOM constituents. The outcomes of studies I and II imply that the RC model should be preferred over linear and discrete (multi)exponential models which incorrectly conceptualize DOM as one to three homogenous pools of constant reactivity.
Study III relates individual decay rates to specific molecular properties. We observed that, on average, highly unsaturated and phenolic compounds are more persistent than compounds with higher aromatic content (plant polyphenols and polycondensed aromatics), reactivity of which additionally increases with increasing nominal oxidation state of carbon. Aliphatic compounds are, on average, found to have the highest decay rates. Results of molecular analysis also indicate that increasing reactivity of DOM after UV exposure is linked to disintegration of supramolecular complexes.
Study IV shows that changes in relative proportions of terrestrial versus algal DOM reflect on degradability of DOM through seasons. Under ice, when algal-derived sources are at their lowest, DOM degradation potential is similarly low across lakes, irrespective of lake type (productive or humic), and bacterial respiration is primarily supported by terrestrial carbon. This can be viewed as a general pattern of baseline metabolism across boreal lakes.
All four studies in this thesis demonstrate that despite the general trends in reactivity attributed to particular DOM fractions (terrestrial, algal, aromatic, photoproduced) or large compound groups (aliphatic, highly unsaturated and phenolic, etc.), each fraction and group contains its own, smaller continuum of reactivity and can include both fast and slowly decaying constituents. Hence, labeling any of the large DOM fractions or groups as ubiquitously labile or recalcitrant can be misleading. This thesis additionally reflects that ultrahigh resolution mass spectrometry is a powerful technique that can be helpful in assessing how reactivity is distributed among DOM constituents and how it responds to specific composition-altering factors, e.g., UV exposure.
Overall, I conclude that bulk DOM is a dynamic reactivity continuum, and degradation patterns of DOM are, in principle, predictable from DOM compositional properties.

33
Summary in Swedish (Sammanfattning)
Grundämnet kol är en av livets viktigaste byggstenar. Det förekommer inte bara i allt levande, utan i ännu större mängder i dött organiskt material. Rester av djur och växter i olika stadier av nedbrytning utgör biosfärens största förråd av organiskt bundet kol. I ekosystem på land (terrestra ekosystem) finns det mesta av det döda organiska materialet i marken; i vatten finns det mesta i form av lösta organiska föreningar och som kolloider, dvs. som partiklar som är alltför små för att sjunka, utan håller sig kvar i vattenmassan. Tillsammans utgör dessa lösta och kolloidala organiska ämnen så kallat löst organiskt material (”dissolved organic matter”, DOM). Min avhandling handlar om DOM i insjöar.
DOM i sjöar och vattendrag är en viktig komponent i ekologiska och biogeokemiska processer. Det är den huvudsakliga energikällan för heterotrofa bakterier, dvs. bakterier som liksom djur är beroende av organisk föda. Genom att DOM är färgat påverkar det ljusklimatet i vattnet och därmed förutsättningarna för fotosyntes. Dessutom påverkar det vattenkemin, inte minst syra-bas-förhållanden, metallers toxicitet, och näringsämnens tillgänglighet. Särskilt i den boreala zonen, dvs. nordliga skogsområden, tillförs stora mängder DOM till sjöar och vattendrag genom avrinning från omgivande landområden. En mindre del av DOM är dock även ett resultat av interna processer i insjöarna, genom växtplanktons och andra gröna växters primärproduktion. En del av DOM bildar partiklar som sjunker till botten och lagras in i sediment. Andra delar av DOM bryts ned av bakterier eller av solljus, vilket leder till att koldioxid och metan bildas, som sedan lämnar vattnet och förs över till atmosfären.
Sjöar täcker en liten del av kontinenterna, endast 3.7% av det isfria landskapets yta. Trots detta avger de stora mängder växthusgaser till atmosfären, till stor del som ett resultat av nedbrytning av DOM. Det är därför viktigt att utröna vilka faktorer som driver nedbrytningen av DOM i sjöar. Det har nyligen visats att mekanismerna för hur DOM bryts ner och försvinner i sjövatten till stor del är en funktion av DOM’s molekylära sammansättning. Sambanden mellan kemiska strukturer och nedbrytbarhet är dock till stora delar okänt. Det övergripande målet med denna avhandling är att öka vår kunskap om dessa samband.
I en serie experiment avseende nedbrytning av DOM har jag undersökt hur dess sammansättning påverkar hur det bryts ned. Optiska metoder, dels spektrofotometri och dels spektrofluorometri, ger via organiska ämnens färg

34
och fluorescens övergripande information om kemisk sammansättning och molekylvikt. Jag använde dessa metoder för att undersöka förekomsten av olika fraktioner i DOM, till exempel aromatiska jämfört med alifatiska strukturer, och molekyler med hög respektive låg molekylvikt. För att få mer detaljerad information använde jag ”electrospray ionization Fourier transform ion cyclotron resonance mass spectrometry” (ESI FT-ICR-MS), en analysmetod med extremt hög upplösning, med förmåga att samtidigt identifiera och särskilja flera tusen olika komponenter som tillsammans utgör DOM, ner till nivån av enskilda molekylformler.
Parallellt med kemisk analys har jag undersökt nedbrytningsmönster hos DOM med hjälp av en modell (”reactive continuum model”, RC-modellen), som återspeglar komplexiteten i hur materialet bryts ner, utifrån teorin att det organiska materialet utgör ett kontinuerligt spektrum av ett mycket stort antal komponenter, som samtidigt var och en bryts ner exponentiellt, men med sinsemellan olika hastighet.
Den första studien i min avhandling visar att fotokemiska förändringar i DOM, som naturligt förekommer som en följd av solens ultravioletta strålning, har stor inverkan på nedbrytningsmönstren. Belysning av DOM medförde en förskjutning till högre nedbrytningshastighet. Tidigare studier har fokuserat på mycket labila föreningar som bildas fotokemiskt, och sedan bryts ner mycket snabbt av mikroorganismer, inom timmar eller dagar. Mina resultat visar att de fotokemiska förändringarna också innebär förhöjd nedbrytningshastighet under månader. Dessutom visar undersökningen att RC-modellen är ett enkelt och informativt verktyg för att beskriva DOM’s nedbrytningskinetik.
Nästa studie belyser nedbrytningen av enskilda komponenter i DOM. Experimentet utgör ett test av ett antagande bakom RC-modellen, att DOM omfattar ett stort antal olika komponenter som bryts ner exponentiellt med olika hastighet. Resultaten visar att så är fallet. De första två studierna sammantaget indikerar att RC-modellen är att föredra framför andra modeller som används för att analysera nedbrytningen av DOM, speciellt linjära modeller och exponentiella modeller som antar en eller ett fåtal homogena pooler av organiskt material med konstant nedbrytbarhet.
I den tredje delen av avhandlingen kopplar jag nedbrytningshastigheten hos individuella komponenter till deras molekylära egenskaper. Kraftigt omättade och fenoliska föreningar var överlag mer moståndskraftiga mot nedbrytning än föreningar med högt aromatiskt innehåll (växtpolyfenoler och polykondenserade aromatiska föreningar). Dessa föreningars nedbrytning var snabbare ju mer oxiderade de var. Alifatiska föreningar uppvisade de högsta nedbrytningshastigheterna. Resultatet av analysen av molekylära egenskaper indikerar också ökad nedbrytningshastighet efter belysning med UV-strålning. Därmed kan samma molekyler visa olika nedbrytbarhet beroende på hur de uppträder i den naturliga miljön. Att de är mer svårnedbrytbara före belysning kan bero på att de i större utsträckning

35
undgår nedbrytning, genom att de skyddas från nedbrytande enzymer till följd av att de är ”gömda” i kolloider eller grupper av molekyler.
I den fjärde studien följde vi DOM’s sammansättning och nedbrytningsmönster under en säsong i flera sjöar, från vinter till sommar. Vi visar att förändringar i den relativa andelen av DOM som har ursprung antingen i material från omgivande marker, eller från sjöns egen primärproduktion via bland annat planktiska algers fotosyntes, var relaterad till DOM’s nedbrytbarhet. På vintern, medan sjöarna fortfarande var islagda och tillgången till ljus var mycket låg, var andelen DOM med ursprung i alger minimal. Vid denna tid på året var nedbrytningspotentialen låg och likartad i de studerade sjöarna, oavsett deras karaktär (brunvattensjöar i skogslandskapet, jämfört med näringsrika sjöar). Bakteriernas aktivitet var under denna tid huvudsakligen beroende av organiskt material från land. Detta kan tolkas som att sjöarna under vintern uppvisar en basmetabolism, understödd av organiskt material från land med begränsad nedbrytbarhet.
Sammantaget visar min avhandling att det finns en generell trend i nedbrytbarhet och nedbrytningshastighet vid jämförelse av olika specifika fraktioner av DOM (till exempel terrestert, av algursprung, aromatiskt, eller fotokemiskt modifierat) eller större grupper av föreningar (till exempel alifatiska eller kraftigt omättade och fenoliska föreningar). Var och en av dessa grupper uppvisar samtidigt ett eget reaktivitetskontinuum, och innehåller komponenter som bryts ned både hastigt och mycket långsamt. Detta visar att breda fraktioner av organiskt material inte kan klassificeras övergripande som antingen labila eller motståndskraftiga mot nedbrytning.
Jag visar i denna avhandling att masspektrometri med ultrahög upplösning är en kraftfull metod för att karakterisera DOM, och hur fördelningen av olika delar av DOM påverkas av specifika yttre faktorer som påverkar den kemiska sammansättningen, till exempel solens UV-ljus.
En viktig slutsats är att DOM utgör ett dynamiskt kontinuum av komponenter som bryts ned i olika hastighet. Därmed är DOM’s nedbrytningsmönster principiellt förutsägbart utifrån kemiska egenskaper hos de tusentals olika kemiska formler som det är uppbyggt av.

36
Thank you
The following pages are dedicated to the people who supported and inspired me during these past years. Knowing that I’m not alone in my journey was the greatest comfort, especially at times when I didn’t really know the way.
To my supervisors and co-authors: Lars, thank you for always finding time for me, no matter how busy you are. In the interview for the PhD position you asked me “Why science?” and showed me some van Krevelen diagrams (to make me consider the alternatives, I suppose). In the following years, with your encouragement and support, I learnt to be more confident about my work, and realized that there is no reason to be scared of van Krevelens, or anything else that can be learnt and understood. You often talked of how bottom-up initiative and diversity sustain a successful group, and for me, being able to work in such an environment was just as important as the research itself. I also thank you for your kindness, patience, and sense of humor. Please know that I couldn’t have wished for a better supervisor. Thank you Thorsten, for your participation and thoughtful assistance. When it comes to mass spectrometry, we truly stand on your shoulders. Your personal advice and the expertise of your lab were an invaluable help to this PhD project. I will forever be fascinated by FT-ICR-MS and will always remember my time in Oldenburg as a unique learning opportunity. Thank you Birgit, for helping me with things that frighten us biologists the most: scripting, modeling, and statistics. I now use R with a confidence I never thought I could achieve and this, to a large extent, is your accomplishment. Your family has grown in the time of my PhD and it was great to see how happy these changes made you. Thank you for your support – it was very important to me. Anna, thanks for your kind advice, both regarding my projects and teaching. Before I went through the pain of getting my drivers license you were the one driving me to the lakes for sampling. Those lakes turned out to be goldmines of data, and as my later history at the department suggests, without you I may not have even sampled the right ones. Francois, not only you are my co-author and a colleague – I consider you my unofficial supervisor and a good friend. I was very lucky to have your support. You always asked important questions and helped me see the forest for the trees. I wish all the best to you and your family: be happy and get the most out of life! Jeff, thank you for our valuable collaboration and for taking the time to share your knowledge with me. It always amazed me how

37
quickly you respond to emails: a truly appreciated quality in our work. I hope you and Jolie continue enjoying Sweden, and also thank you for your regular Pub Quiz, where I could both have fun and feel extremely uneducated in pop culture.
To the Oldenburg team: Jutta, Helena, Katrin, Maren Z., Betty and all the other members of ICBM Marine Geochemistry Group, thanks for making my visits to Oldenburg pleasant and fruitful.
To my current and former officemates: Katrin and Matilda, thank you for making our long days in the office brighter. Upon many hours of wasted R efforts, hearing you laugh in our office was the most cheerful thing. In our work, it’s nice to be able to switch off sometimes and chat about random things. Thank you for sharing small moments like that with me. Anne, Cristian, and Núria, thanks for your good energy and friendly attitude. Anne, thanks also for the work you’ve done before my time. Your ideas and findings were a great inspiration for my project.
To all generations of PhD students: Valerie, thanks for the adventures we had together, for talking to me when I needed it, and for all those weekends we spent in the awesome city of Stockholm. Be happy and successful, my dear, and thank you for our friendship. Frieda, thank you for being a sincere, supporting friend, with a great sense of humor. Karólína, thank you for being such a good friend and for making me feel that I can always rely on you. Your hard work and enthusiasm are inspiring, and I wish that there were more people like you in this world: caring, fair-minded, and giving. Thank you so much for your support and your friendship! Máté, thanks for making me laugh. Even your sarcasm is always sweet. Thanks for your sincerity and friendship, and thanks for listening to me and showing that you care about my stupid problems. Lucas, thank you for our thought-provoking conversations and fun activities, like the vacations in Norredatorp and ice-skating to complete exhaustion. Thanks also for a great idea for my thesis cover. You are a good friend and I hope we can hang out more, despite the distance. Jovana and Hannah, what a great time we had in New Orleans and New York! Jovana, thanks for keeping an eye on me and Hannah and being the most responsible among us. You are a very strong and determined person – I always admired that about you. Hannah, thank you for being so friendly and easy-going. Your positive thinking instantly makes things better, thank you for sharing your light with all of us. Roger, thank you for being a very considerate, helpful friend. You, Christina and little Nico make an adorable family; be happy guys. Thanks Blaize, for your energy, lovely thanksgiving dinners, cool EDLA games, and countless attempts to break awkward fika silence. Monica, thanks for your friendly and graceful personality and your empathy toward others. Thanks Lorena, for your cheerful character and our small chats when you were around. Maria, thanks for being friendly and fun, I look forward to seeing you again after this long time. Inga and Domenico,

38
thanks for being good company for going out. Thanks Yinghua, for being nice and helpful, and for our hilarious efforts to communicate with the fish salesmen. Fernando, thank you for being helpful and creative, especially when it comes to writing the spex. You have a good heart and it is always a pleasure to have you around. Thanks Anastasia for your good spirit and irrepressible optimism, and for encouraging others to be more active. Thanks Martin for every interesting conversation we had on every possible topic. Thanks Annika, Marloes and Leyden for sharing the teaching responsibilities with me and for your company on many other occasions. Thanks Anna N. for managing our PhD group so well, and thanks Fabian, Xavi, and Theresa for participating in our entertaining activities.
To the department seniors, researchers, engineers and postdocs: Eva and Peter, thanks for your kind attitude and all the good efforts that you make for our department. Eva, thanks also for your sparkling sense of humor – being that funny and at the same time highly professional is a talent. Stefan, thank you for encouraging us PhDs to be brave and think bigger, and thanks for your hospitality and delicious homemade food. Gesa, thanks for the words of encouragement, your special wisdom, and your active personality. Silke, thanks for involving me in your course and giving me and other assistants an opportunity to share our research with the students. I wish you success in your work at Erken. Thanks Kurt and Karin for taking good care of the Erken station over the years. It was always a pleasure to visit the station and spend time in its nice, home-like atmosphere. Don, thanks for an interesting excursion to Malma island and for showing us all the cool equipment at Erken. Sebastian, thank you for being kind and helpful, and for providing the best music for many Limno parties. Thanks Dolly, for being this awesome person radiating empathy and warmth. Thank you for taking time to discuss my data with me – I always appreciated your ability to explain difficult things with easy language. Your graceful and easy-going character is an inspiration, and I wish tons of happiness to you and your family. Janne, thank you for every time when I thought to myself “Calm down, Janne is on his way”. I learnt a lot from you, perhaps most importantly, not to panic, because the solution can always be found as long as we act and think more like you. Christoffer, thank you for being helpful with practical things in the lab. Thanks Anna S. for your sincerity and ever-present will to help; also big thanks to Lujza for being kind and friendly with Polya – we appreciate it a lot. Thanks Andrea, Kristin, Sari, and Pablo for your cheerful personalities. Thanks Raven for being helpful and friendly. Moritz, thanks for working your R magic in the moments when I was about to give up. Thank you Sarahi, for your good spirit, interesting initiatives, and for sharing your ideas and experience with others. Simone, thanks for your kindness and warmth, and of course, for helping me to take care of the misbehaving equipment.

39
To those who first inspired me to pursue aquatic sciences: big thanks to Nikolay Kashulin, Irina Koroleva, Anatoly Lukin, Julia Sharova, Per-Arne Amundsen, Rune Knudsen, and everyone else who showed me how great it is to be a limnologist.
To my former supervisor: Emma, not only did you spark my interest in biogeochemistry, you also showed me what it means to be a good teacher and a good scientist. Thank you. I will never forget it. I also thank your husband Niklas, for his dedicated work that makes people think about the values of empathy, tolerance, and equality.
To my friends: Silke E., thank you so much for the great time I had in Oldenburg! You are one of the most helpful and selfless people I know, and I admire it very much. Dina, thanks for your friendship and your hospitality. Those days we spent together in Stockholm were really nice. I miss you and I hope we will meet again some day. Natasha, we met not so long ago, but it feels like we’ve known each other for years. Thank you for your hospitality, sense of humor, support and friendship. You are a smart and strong person, and whatever you do in the future, I know that you will succeed. Juan, thanks for your friendship and support, especially in these past weeks when you reminded me that I should sometimes leave my computer. Liliya, my sweet and loving friend, thank you for being my travelling companion and my source of inspiration. Markusha and co., thanks for our reunions that made me feel like I never left home. Andrey, thank you for your support, optimism and friendship, and for keeping our communication alive despite the distance. Julia and Ira, thank you for our small traditions and for the friendship between our families.
To my family To my grandparents: thank you for the childhood memories that I have
from your home, for biology books on your shelves, for sharing your life wisdom with me and for always putting us children first and loving us unconditionally.
To my dad: thank you for your care and love and for constantly making the compromises in the family full of females with a lot of opinions.
To my mom: thank you for being a role model of a person who always takes the high road, even if it’s not easy. Nothing measures up to the amount of help me and Polya got from you throughout our lives – I cannot even describe it with words. Thank you for your love and support, because without it I would not be where I am right now.
To Polya: thank you for patience and understanding, for bringing joy into my life, and for every minute we spend together. I love you with all my heart. You are my light, my happiness, and my hope. You are my everything.

40
References
Amon, R. M. W., and R. Benner. 1996. Bacterial utilization of different size classes of dissolved organic matter. Limnology and Oceanography 41:41–51.
Arnosti, C. 2011. Microbial extracellular enzymes and the marine carbon cycle. Annual review of marine science 3:401–425.
Azam, F., T. Fenchel, J. G. Field, J. C. Gray, L. A. Meyer-Reil, and F. Thingstad. 1983. The ecological role of water-column microbes in the sea. Marine Ecology Progress Series 10:257–263.
Bastviken, D., L. J. Tranvik, J. A. Downing, P. M. Crill, and A. Enrich-Prast. 2011. Freshwater methane emissions offset the continental carbon sink. Science 331:50.
Berner, R. A. 1964. An idealized model of dissolved sulfate distribution in recent sediments. Geochimica et Cosmochimica Acta 28:1497–1503.
Bertilsson, S., and L. J. Tranvik. 1998. Photochemically produced carboxylic acids as substrates for freshwater bacterioplankton 43:885–895.
Bosatta, E., and G. I. Ågren. 1995. The power and reactive continuum models as particular cases of the q-theory of organic matter dynamics. Geochimica et Cosmochimica Acta 59:3833–3835.
Boudreau, B. P., C. Arnosti, B. B. Jørgensen, and D. E. Canfield. 2008. Comment on “Physical model for the decay and preservation of marine organic carbon.” Science 319:1616.
Boudreau, B. P., and B. R. Ruddick. 1991. On a reactive continuum representation of organic matter diagenesis. American Journal of Science 291:507–538.
Carpenter, A., S. Siggia, and S. Carter. 1976. Separation and/or concentration of phenolic materials from dilute aqueous solutions 48:225–228.
Carpenter, S. R. 1981. Decay of heterogenous detritus: a general model. Journal of theoretical Biology 89:539–547.
Carpenter, S. R., J. J. Cole, M. L. Pace, M. Van de Bogert, D. L. Bade, D. Bastviken, C. M. Gille, J. R. Hodgson, J. F. Kitchell, and E. S. Kritzberg. 2005. Ecosystem subsidies: terrestrial support of aquatic food webs from 13C addition to contrasting lakes. Ecology 86:2737–2750.
Cole, J. J., Y. T. Prairie, N. F. Caraco, W. H. McDowell, L. J. Tranvik, R. G. Striegl, C. M. Duarte, P. Kortelainen, J. A. Downing, J. J. Middelburg, and J. Melack. 2007. Plumbing the global carbon cycle: integrating inland waters into the terrestrial carbon budget. Ecosystems 10:171–184.
Cory, R. M., M. P. Miller, D. M. McKnight, J. J. Guerard, and P. L. Miller. 2010. Effect of instrument-specific response on the analysis of fulvic acid fluorescence spectra. Limnology and Oceanography: Methods 8:67–78.
Duarte, C. M., and Y. T. Prairie. 2005. Prevalence of heterotrophy and atmospheric CO2 emissions from aquatic ecosystems. Ecosystems 8:862–870.
Fellman, J. B., E. Hood, and R. G. M. Spencer. 2010. Fluorescence spectroscopy opens new windows into dissolved organic matter dynamics in freshwater ecosystems: a review. Limnology and Oceanography 55:2452–2462.

41
Freeman, C., N. J. Ostle, N. Fenner, and H. Kang. 2004. A regulatory role for phenol oxidase during decomposition in peatlands. Soil Biology and Biochemistry 36:1663–1667.
Green, N. W., E. M. Perdue, G. R. Aiken, K. D. Butler, H. Chen, T. Dittmar, J. Niggemann, and A. Stubbins. 2014. An intercomparison of three methods for the large-scale isolation of oceanic dissolved organic matter. Marine Chemistry 161:14–19.
Guillemette, F., and P. A. del Giorgio. 2011. Reconstructing the various facets of dissolved organic carbon bioavailability in freshwater ecosystems. Limnology and Oceanography 56:734–748.
Guillemette, F., S. L. McCallister, and P. A. del Giorgio. 2013. Differentiating the degradation dynamics of algal and terrestrial carbon within complex natural dissolved organic carbon in temperate lakes. Journal of Geophysical Research: Biogeosciences 118:963–973.
Helms, J. R., A. Stubbins, J. D. Ritchie, E. C. Minor, D. J. Kieber, and K. Mopper. 2008. Absorption spectral slopes and slope ratios as indicators of molecular weight, source, and photobleaching of chromophoric dissolved organic matter. Limnology and Oceanography 53:955–969.
Hessen, D. 1992. Dissolved organic carbon in a humic lake: effects on bacterial production and respiration. Hydrobiologia:115–123.
Karlsson, J., M. Berggren, J. Ask, P. Byström, A. Jonsson, H. Laudon, and M. Jansson. 2012. Terrestrial organic matter support of lake food webs: Evidence from lake metabolism and stable hydrogen isotopes of consumers. Limnology and Oceanography 57:1042–1048.
Karlsson, J., M. Jansson, and A. Jonsson. 2007. Respiration of allochthonous organic carbon in unproductive forest lakes determined by the Keeling plot method. Limnology and Oceanography 52:603–608.
Kellerman, A. M., T. Dittmar, D. N. Kothawala, and L. J. Tranvik. 2014. Chemodiversity of dissolved organic matter in lakes driven by climate and hydrology. Nature Communications 5:1–8.
Kellerman, A. M., D. N. Kothawala, T. Dittmar, and L. J. Tranvik. 2015a. Persistence of dissolved organic matter in lakes related to its molecular characteristics. Nature Geoscience 8:454–457.
Kellerman, A. M., D. N. Kothawala, T. Dittmar, and L. J. Tranvik. 2015b. Do we have a weight problem? The discrepancy between actual and apparent molecular weight of dissolved organic matter in lakes. in Kellerman A. M. Molecular-level dissolved organic matter dynamics in lakes: constraints on reactivity and persistence. Doctoral thesis. Uppsala University.
Kim, S., R. W. Kramer, and P. G. Hatcher. 2003. Graphical method for analysis of ultrahigh-resolution broadband mass spectra of natural organic matter, the van Krevelen diagram. Analytical Chemistry 75:5336–5344.
Kim, S., R. P. Rodgers, and A. G. Marshall. 2006. Truly “exact” mass: Elemental composition can be determined uniquely from molecular mass measurement at∼ 0.1 mDa accuracy for molecules up to∼ 500Da. International Journal of Mass Spectrometry 251:260–265.
Kirk, J. T. O. 1994. Light and Photosynthesis in Aquatic Ecosystems. Cambridge University Press.
Koch, B. P., and T. Dittmar. 2006. From mass to structure: an aromaticity index for high‐resolution mass data of natural organic matter. Rapid communications in mass spectrometry 20:926–932.

42
Koch, B. P., T. Dittmar, M. Witt, and G. Kattner. 2007. Fundamentals of molecular formula assignment to ultrahigh resolution mass data of natural organic matter. Analytical Chemistry 79:1758–1763.
Koch, B. P., M. Witt, R. Engbrodt, T. Dittmar, and G. Kattner. 2005. Molecular formulae of marine and terrigenous dissolved organic matter detected by electrospray ionization Fourier transform ion cyclotron resonance mass spectrometry. Geochimica et Cosmochimica Acta 69:3299–3308.
Koehler, B., and L. J. Tranvik. 2015. Reactivity continuum modeling of leaf, root, and wood decomposition across biomes. Journal of Geophysical Research: Biogeosciences 120:1196–1214.
Koehler, B., E. Von Wachenfeldt, D. Kothawala, and L. J. Tranvik. 2012. Reactivity continuum of dissolved organic carbon decomposition in lake water. Journal of Geophysical Research: Biogeosciences 117:1–15.
Kothawala, D. N. ., K. R. . Murphy, C. A. . Stedmon, G. A. . Weyhenmeyer, and L. J. . Tranvik. 2013. Inner filter correction of dissolved organic matter fluorescence. Limnology and Oceanography: Methods 11:616–630.
Lavonen, E. E., M. Gonsior, L. J. Tranvik, P. Schmitt-Kopplin, and S. J. Köhler. 2013. Selective chlorination of natural organic matter: identification of previously unknown disinfection byproducts. Environmental Science and Technology 47:2264–2271.
Lawaetz, A. J., and C. A. Stedmon. 2009. Fluorescence intensity calibration using the Raman scatter peak of water. Applied Spectroscopy 63:936–940.
Lydersen, E. 1998. Humus and acidification. Pages 63–92 in D. Hessen and L. J. Tranvik, editors. Aquatic humic substances: Ecology and biogeochemistry. Springer.
McCallister, S. L., and P. A. del Giorgio. 2008. Direct measurement of the δ13C signature of carbon respired by bacteria in lakes: Linkages to potential carbon sources, ecosystem baseline metabolism, and CO2 fluxes. Limnology and Oceanography 53:1204–1216.
McKnight, D. M., E. W. Boyer, P. K. Westerhoff, P. T. Doran, T. Kulbe, and D. T. Anderson. 2001. Spectroflourometric characterization of dissolved organic matter for indication of precursor organic material and aromaticity. Limnology and Oceanography 46:38–48.
Middelburg, J. J. 1989. A simple rate model for organic matter decomposition in marine sediments. Geochimica et Cosmochimica Acta 53:1577–1581.
Minor, E. C., C. J. Steinbring, K. Longnecker, and E. B. Kujawinski. 2012. Characterization of dissolved organic matter in Lake Superior and its watershed using ultrahigh resolution mass spectrometry. Organic Geochemistry 43:1–11.
Moran, M. A., W. M. Sheldon, and R. G. Zepp. 2000. Carbon loss and optical property changes during long-term photochemical and biological degradation of estuarine dissolved organic matter. Limnology and Oceanography 45:1254–1264.
Moran, M. A., and R. G. Zepp. 1997. Role of photoreactions in the formation of biologically labile compounds from dissolved organic matter. Limnology and Oceanography 42:1307–1316.
Murphy, K. R., C. A. Stedmon, D. Graeber, and R. Bro. 2013. Fluorescence spectroscopy and multi-way techniques. PARAFAC. Analytical Methods 5:6557–6566.
Olson, J. S. 1963. Energy storage and the balance of producers and decomposers in ecological systems. Ecology 44:322–331.

43
Parlanti, E., K. Wörz, L. Geoffroy, and M. Lamotte. 2000. Dissolved organic matter fluorescence spectroscopy as a tool to estimate biological activity in a coastal zone submitted to anthropogenic inputs. Organic Geochemistry 31:1765–1781.
Pelet, R. 1984. A model for the biological degradation of recent sedimentary organic matter. Organic Geochemistry 6:317–325.
Perdue, E. M. 1998. Chemical composition, structure, and metal binding properties. Pages 41–61 in D. Hessen and L. J. Tranvik, editors. Aquatic humic substances: Ecology and biogeochemistry. Springer.
Perdue, E. M., and J. D. Ritchie. 2003. Dissolved organic matter in freshwaters. Pages 273-318 in J. I. Drever, editor. Surface and Ground Water, Weathering, Erosion and Soils. Treatise on geochemistry 5. Elsevier–Pergamon, Oxford.
Peuravuori, J., and K. Pihlaja. 1997. Molecular size distribution and spectroscopic properties of aquatic humic substances. Analytica Chimica Acta 337:133–149.
Raymond, P. A., J. Hartmann, R. Lauerwald, S. Sobek, C. McDonald, M. Hoover, D. Butman, R. Striegl, E. Mayorga, C. Humborg, P. Kortelainen, H. Durr, M. Meybeck, P. Ciais, and P. Guth. 2013. Global carbon dioxide emissions from inland waters. Nature 503:355–359.
Riedel, T., H. Biester, and T. Dittmar. 2012. Molecular fractionation of dissolved organic matter with metal salts. Environmental Science and Technology 46:4419–4426.
Riedel, T., M. Zark, A. V. Vähätalo, J. Niggemann, R. G. M. Spencer, P. J. Hernes, and T. Dittmar. 2016. Molecular signatures of biogeochemical transformations in dissolved organic matter from ten world rivers. Frontiers in Earth Science 4.
Šantl-Temkiv, T., K. Finster, T. Dittmar, B. M. Hansen, R. Thyrhaug, N. W. Nielsen, and U. G. Karlson. 2013. Hailstones: a window into the microbial and chemical inventory of a storm cloud. PloS one 8:e53550.
Schmidt, M. W. I., M. S. Torn, S. Abiven, T. Dittmar, G. Guggenberger, I. A. Janssens, M. Kleber, I. Kögel-Knabner, J. Lehmann, D. A. C. Manning, P. Nannipieri, D. P. Rasse, S. Weiner, and S. E. Trumbore. 2011. Persistence of soil organic matter as an ecosystem property. Nature 478:49–56.
Seidel, M., P. L. Yager, N. D. Ward, E. J. Carpenter, H. R. Gomes, A. V. Krusche, J. E. Richey, T. Dittmar, and P. M. Medeiros. 2015. Molecular-level changes of dissolved organic matter along the Amazon River-to-ocean continuum. Marine Chemistry 177:218–231.
Skopintsev, B. A. 1981. Decomposition of organic matter of plankton, humification and hydrolysis. Pages 125-177 in E. K. Duursma and R. Dawson, editors. Marine organic chemistry. Elsevier.
Solomon, C. T., D. A. Bruesewitz, D. C. Richardson, K. C. Rose, M. C. Van de Bogert, P. C. Hanson, T. K. Kratz, B. Larget, R. Adrian, B. L. Babin, C.-Y. Chiu, D. P. Hamilton, E. E. Gaiser, S. Hendricks, V. Istvánovics, A. Laas, D. M. O’Donnell, M. L. Pace, E. Ryder, P. A. Staehr, T. Torgersen, M. J. Vanni, K. C. Weathers, and G. Zhu. 2013. Ecosystem respiration: Drivers of daily variability and background respiration in lakes around the globe. Limnology and Oceanography 58:849–866.
Stedmon, C. A., and S. Markager. 2005. Resolving the variability of dissolved organic matter fluorescence in a temperate estuary and its catchment using PARAFAC analysis. Limnology and Oceanography 50:686–697.
Stenson, A. C., A. G. Marshall, and W. T. Cooper. 2003. Exact masses and chemical formulas of individual Suwannee River fulvic acids from ultrahigh resolution electrospray ionization Fourier transform ion cyclotron resonance mass spectra. Analytical Chemistry 75:1275–1284.

44
Stubbins, A., R. G. M. Spencer, H. Chen, P. G. Hatcher, K. Mopper, P. J. Hernes, V. L. Mwamba, A. M. Mangangu, J. N. Wabakanghanzi, and J. Six. 2010. Illuminated darkness: molecular signatures of Congo River dissolved organic matter and its photochemical alteration as revealed by ultrahigh precision mass spectrometry. Limnology and Oceanography 55:1467–1477.
Tranvik, L. J. 1992. Allochthonous dissolved organic matter as an energy source for pelagic bacteria and the concept of the microbial loop. Hydrobiologia:107–114.
Tranvik, L. J. 1998. Degradation of dissolved organic matter in humic waters by bacteria. Pages 259–283 in D. Hessen and L. J. Tranvik, editors. Aquatic humic substances: Ecology and biogeochemistry. Springer.
Tranvik, L. J. 2009. Dystrophy. Pages 405–410 in G. E. Likens, editor. Encyclopedia of inland waters. Academic Press.
Tranvik, L. J., J. A. Downing, J. B. Cotner, S. A. Loiselle, R. G. Striegl, T. J. Ballatore, P. Dillon, K. Finlay, K. Fortino, and L. B. Knoll. 2009. Lakes and reservoirs as regulators of carbon cycling and climate. Limnology and Oceanography 54:2298–2314.
Verpoorter, C., T. Kutser, D. A. Seekell, and L. J. Tranvik. 2014. A global inventory of lakes based on high-resolution satellite imagery. Geophysical Research Letters 41:6396–6402.
Vähätalo, A. V., H. Aarnos, and S. Mäntyniemi. 2010. Biodegradability continuum and biodegradation kinetics of natural organic matter described by the beta distribution. Biogeochemistry 100:227–240.
Walker, B. D., S. R. Beaupré, T. P. Guilderson, M. D. McCarthy, and E. R. M. Druffel. 2016. Pacific carbon cycling constrained by organic matter size, age and composition relationships. Nature Geoscience 9:888–891.
Weishaar, J. L., G. R. Aiken, B. A. Bergamaschi, M. S. Fram, R. Fujii, and K. Mopper. 2003. Evaluation of specific ultraviolet absorbance as an indicator of the chemical composition and reactivity of dissolved organic carbon. Environmental Science and Technology 37:4702–4708.
Westrich, J. T., and R. A. Berner. 1984. The role of sedimentary organic matter in bacterial sulfate reduction: The G model tested. Limnology and Oceanography 29:236–249.
Wetzel, R. G. 2003. Dissolved oganic carbon: detrital energetics, metabolic regulators, and drivers of ecosystem stability of aquatic ecosystems. Pages 455–477 in S. E. G. Findlay and R. A. Sinsabaugh Aquatic ecosystems: interactivity of dissolved organic matter. Academic Press.
Wetzel, R. G., P. G. Hatcher, and T. S. Bianchi. 1995. Natural photolysis by ultraviolet irradiance of recalcitrant dissolved organic matter to simple substrates for rapid bacterial metabolism. Limnology and Oceanography 40:1369–1380.
Weyhenmeyer, G. A., M. Fröberg, E. Karltun, M. Khalili, D. Kothawala, J. Temnerud, and L. J. Tranvik. 2012. Selective decay of terrestrial organic carbon during transport from land to sea. Global Change Biology 18:349–355.
Zsolnay, A., E. Baigar, M. Jimenez, B. Steinweg, and F. Saccomandi. 1999. Differentiating with fluorescence spectroscopy the sources of dissolved organic matter in soils subjected to drying. Chemosphere 38:45–50.


Acta Universitatis UpsaliensisDigital Comprehensive Summaries of Uppsala Dissertationsfrom the Faculty of Science and Technology 1486
Editor: The Dean of the Faculty of Science and Technology
A doctoral dissertation from the Faculty of Science andTechnology, Uppsala University, is usually a summary of anumber of papers. A few copies of the complete dissertationare kept at major Swedish research libraries, while thesummary alone is distributed internationally throughthe series Digital Comprehensive Summaries of UppsalaDissertations from the Faculty of Science and Technology.(Prior to January, 2005, the series was published under thetitle “Comprehensive Summaries of Uppsala Dissertationsfrom the Faculty of Science and Technology”.)
Distribution: publications.uu.seurn:nbn:se:uu:diva-316893
ACTAUNIVERSITATIS
UPSALIENSISUPPSALA
2017