dissolved humic substances of vascular plant origin in a coastal marine environment
TRANSCRIPT

Dissolved Humic Substances of Vascular Plant Origin in a Coastal Marine EnvironmentAuthor(s): Mary Ann Moran and Robert E. HodsonSource: Limnology and Oceanography, Vol. 39, No. 4 (Jun., 1994), pp. 762-771Published by: American Society of Limnology and OceanographyStable URL: http://www.jstor.org/stable/2838414 .
Accessed: 17/06/2014 23:55
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
American Society of Limnology and Oceanography is collaborating with JSTOR to digitize, preserve andextend access to Limnology and Oceanography.
http://www.jstor.org
This content downloaded from 188.72.126.55 on Tue, 17 Jun 2014 23:55:42 PMAll use subject to JSTOR Terms and Conditions

Limnol. Oceanogr., 39(4), 1994, 762-771 ? 1994, by the Amencan Society of Limnology and Oceanography, Inc.
Dissolved humic substances of vascular plant origin in a coastal marine environment
Mary Ann Moran and Robert E. Hodson Department of Marine Sciences, University of Georgia, Athens 30602-2206
Abstract
Bacterial decomposition of vascular plant detritus in coastal wetlands results in the conversion of particulate organic matter to dissolved form and causes the release of humic substances into the bulk dissolved organic carbon (DOC) pool. We found that 34% of the DOC accumulating during degradation of salt marsh grass (Spartina alterniflora) from coastal wetlands of the southeastern U.S. fits the definition of humic substances and that lignin is the primary source of the dissolved humic substances (66% of the total). Although marine bacterioplankton used the lignin-rich humic substances more slowly and less efficiently than other components of the DOC pool, a significant fraction (24%) of these substances was mineralized within 7 weeks. Concentrations of vascular plant biomarkers (lignin phenols) indicate that 11-75% of the dissolved humic substances on the southeastern U.S. continental shelf is from vascular plant-dominated environments. Calculations indicate that about half this material is contributed by the coastal salt marshes and half by river export. Vascular plant influence was lower in the bulk DOC pool (2-38%), indicating that terrestrially derived material is harbored preferentially in the humic substances subcomponent of marine DOC.
Solubilization of particulate organic matter by bacteria has recently been recognized as an important feature of carbon cycling in marine ecosystems and a quantitatively important route by which carbon is transferred from the particulate to the dissolved phase (Moran and Hodson 1989; Smith et al. 1992). This process appears to be especially significant in coastal salt marshes of the southeastern U.S., where as much as 40% of the carbon in Spartina al- terniflora detritus is converted to stable dis- solved intermediates during bacterial decom- position (Moran and Hodson 1989). The conversion of vascular plant detritus to dis- solved organic matter (DOM) is significant from an ecological standpoint because it pro- vides plant-derived carbon to populations of salt marsh bacteria other than those attached to the particulate detritus, including free-living bacterioplankton (Moran and Hodson 1989). Furthermore, plant-derived DOM is suscep- tible to export from the coastal salt marshes to adjacent marine environments, whereas
Acknowledgments Edward Sheppard provided technical assistance in the
lab and field. We appreciate the detailed reviews of this manuscript by John Ertel, Rick Keil, and John Hedges.
This research was funded by grants NA80AA-D0009 1 from the NOAA Office of Sea Grant and OCE 91-16450 from the National Science Foundation.
plant material in particulate form generally is not (Chalmers et al. 1 985; Williams et al. 1 992).
The ecological role of vascular plant-derived DOM, both within coastal marshes and in ma- rine ecosystems to which it has been exported, will depend on its relative biological lability among other factors. We know from previous studies that rates of utilization of DOM de- rived from Spartina detritus are highly vari- able: some components are rapidly assimilated by bacteria over time scales of hours to days, whereas other components have much longer turnover times (Moran and Hodson 1989). The refractory components are of particular inter- est because they appear to be enriched in lignin byproducts and are potential precursors of hu- mic substances (Moran and Hodson 1990b). The refractory components are also the com- ponents most likely to persist long enough to be available for export. Indeed, the export of humified, lignin-rich DOM to marine envi- ronments has been demonstrated by the iden- tification of lignin biomarkers in the humic substances of several coastal and oceanic sys- tems (Meyers-Schulte and Hedges 1986; Mo- ran et al. 199 1a,b).
Although the rates of turnover and the eco- logical roles of chemically identifiable, low molecular weight components of marine DOM have been studied for nearly three decades (Wright and Hobbie 1966; Azam and Hodson 1977), little is known about the fate of refrac-
762
This content downloaded from 188.72.126.55 on Tue, 17 Jun 2014 23:55:42 PMAll use subject to JSTOR Terms and Conditions

Coastal humic substances 763
tory, higher molecular weight dissolved organ- ics, especially marine humic substances. Ra- diocarbon dating indicates that humic substances constitute the oldest component of marine dissolved organic carbon (DOC) (Bauer et al. 1992), which suggests a largely geochem- ical, rather than ecological, role for these com- pounds in oceanic environments. Yet recent studies of humic substances in freshwater en- vironments demonstrate that some humic ma- terial is turning over on relatively short time scales and that the turnover is driven by bac- terial activity (Moran and Hodson 1 990a; Tranvik 1990; Tulonen et al. 1992). Thus, fur- ther study of the formation and turnover of dissolved humic substances in marine ecosys- tems is warranted.
In this study, we begin to assemble a more complete picture of the ecological role of vas- cular plant-derived humic substances in coast- al marine environments. We document the microbially mediated formation of dissolved humic substances from S. alterniflora detritus in coastal salt marshes of the southeastern U.S., measure rates of turnover of this material by natural marine bacterioplankton, and estimate the contribution of these coastal marshes to the dissolved humic substances pool on the continental shelf.
Methods Radiolabeling S. alterniflora lignocellu-
lose-S. alterniflora plants were collected in- tact from a salt marsh on Sapelo Island, Geor- gia, and maintained in pots under natural light. Plants were uniformly radiolabeled by peri- odically, over a period of several months, plac- ing Mylar bags over the plants and allowing 14C02 inside the bags to be incorporated into plant material. Two weeks after the final ad- dition of 14C02, aboveground portions of plants were harvested by cutting culms at sediment level. Additional S. alterniflora plants were la- beled specifically in the lignin fraction of the lignocellulose by incubating live cuttings in sterile aqueous solutions containing [U- 14C]cinnamic acid, which had been synthe- sized enzymatically from commercially avail- able [U-14C]phenylalanine. The cinnamic acid, an intermediate in the pathway of lignin bio- synthesis, was incorporated primarily into the lignin fraction of the plant material. After la-
beling, both types of plant material were dried at 50?C and ground into small particles (<425 Am in size) in a Wiley mill. Unincorporated radiolabel and labeled nonstructural compo- nents were removed from the plant material by serial extraction in boiling ethanol, ethanol- benzene [ 1: 2 (vol : vol)], and water (Benner et al. 1984), resulting in either uniformly labeled lignocellulose ([U-14C]lignocellulose; lignin, cellulose, and hemicellulose all labeled at the same specific activity) or lignin-labeled lig- nocellulose ([14C-lignin]lignocellulose; lignin fraction labeled, cellulose and hemicellulose unlabeled). These lignocellulose preparations are good models for naturally occurring Spar- tina detritus because lignocellulose accounts for the bulk (> 95%) of the remaining material after natural plant senescence in the salt marsh.
Chemical fractionation and radioanalysis of the 14C-labeled lignocellulose (according to methods of Benner et al. 1984) indicated that the label in the [U-14C]lignocellulose was dis- tributed proportionately among the major chemical fractions and that the [14CC-ig- nin]lignocellulose was substantially free of ra- diolabeled contaminants. Specific activities of the radiolabeled lignocellulose were deter- mined by combusting subsamples at 900?C in a Biological Oxidizer (Harvey Instruments) and radioassaying evolved 14CO2 after trapping in ethanolamine-amended scintillation cocktail. Specific activity of the lignin fraction of [14C- lignin]lignocellulose was calculated assuming that lignin accounts for 10% of the carbon in S. alterniflora lignocellulose (Hodson et al. 1984). Humic substance formation-Spartina lig-
nocellulose (2-2.5 g) was placed in acid-washed 1-liter flasks with 700 ml of half-strength ar- tificial seawater (Sigma Sea Salts, 20 g liter-1) prepared with autoclaved, distilled water. Three milliliters of a marine microbial inoc- ulum were added to each flask. The inoculum was obtained by concentrating microorgan- isms from 1 liter of salt marsh water into a 20- ml volume over 0.2-Am pore-size Nuclepore filters. Inorganic nutrients were added at con- centrations typical of marsh water: 10 AM N (as ammonia and nitrate) and 1 AM P. Three preparations of Spartina lignocellulose were incubated (two replicates each): [U-14C]ligno- cellulose, [14C-lignin]lignocellulose, and unla- beled lignocellulose.
This content downloaded from 188.72.126.55 on Tue, 17 Jun 2014 23:55:42 PMAll use subject to JSTOR Terms and Conditions

764 Moran and Hodson
Flasks were incubated with constant stirring and aeration to prevent accumulation of plant detritus at the bottom of the flasks and to en- sure aerobic conditions throughout the exper- iment. Elevation of flasks 1.5 cm above the surface of the stir plates minimized effects of the heat generated by continuous stirring. Aer- ation was provided by individual aquarium pumps equipped with in-line disposable filter units (Gelman Acrodisc, 0.2-Am pore-size) which were replaced weekly.
After 6 weeks, contents of the flasks were harvested by sequential filtration through Whatman GF/C glass-fiber filters and 0.2-gm pore-size Nuclepore filters. For humic sub- stance isolation and quantification, filtrate was acidified to pH 2 and pumped through 2.2- x 40-cm columns containing Amberlite XAD-8 resin. The nonhumic compounds (those pass- ing through the XAD-8 resin without adher- ing) were collected in the column effluent. The humic compounds (those concentrated on the resin) were eluted in 0.1 N NaOH and then passed through an Amberlite AGMP-50 cat- ion exchange resin to remove sodium ions.
DOC concentrations in the humic and non- humic fractions were determined by direct in- jection into a high-temperature combustion carbon analyzer (Shimadzu TOC-500). Radio- label in these fractions was determined by as- saying 1 -ml subsamples added to 10 ml of scin- tillation fluid after the pH of each was adjusted to neutral with concentrated NaOH or HCI. Recovery of radiolabel after the humic sub- stance isolation procedure averaged 81%; about half of the unrecovered label (9% of the total) was found to remain on the XAD-8 resin. The humic and nonhumic fractions were stored frozen until further use.
Humic substance utilization-Turnover of dissolved humic substances derived from Spartina lignocellulose was measured in mi- crocosms containing DOC, consisting of one labeled fraction-either humic or nonhumic (formed during the decomposition of [U- 14C]lignocellulose)-and one unlabeled frac- tion (formed during the decomposition of un- labeled lignocellulose). This approach avoided potential artifacts associated with the use of physically isolated DOC components during measurements of utilization and allowed for the possibility of bacterial cometabolism of the more refractory compounds. The final DOC mixture contained humic and nonhumic sub-
stances at their original concentrations. Six 125- ml bottles were established with the humic fraction containing the radiolabel, and six bot- tles were established with the nonhumic frac- tion containing the label. Final volume per microcosm was 25 ml, and final DOC concen- tration (labeled plus unlabeled) was 6 mg C liter-1. Three bottles of each set were inocu- lated with 1 ml of a salt marsh microbial in- oculum prepared as described above; the other three served as sterile controls. Bottles were shaken gently (to minimize aggregation of [14C]DOC) at 100 rpm in the dark at room temperature.
Experiments were terminated after 7 weeks. A 5-ml aliquot was removed from each mi- crocosm and stored at 4?C in 2% buffered formaldehyde for later bacterial counts. The remainder was passed through a 0. 22-Am pore- size Millipore filter. The filter was washed with filter-sterilized seawater to remove unincor- porated [14C]DOC, placed in 10 ml of scintil- lation cocktail, and assayed to determine ra- dioactivity incorporated into particulate material (representing incorporation into mi- crobial biomass after correction for sterile con- trols). A 1 -ml aliquot of the filtrate was acid- ified to pH 2.5 to remove inorganic C and then radioassayed to determine unincorporated [14C]DOC. The remaining filtrate (- 20 ml) was acidified to pH 2.0 and pumped through XAD-8 columns (0.8 by 12 cm). Column ef- fluent (nonhumic compounds) and column el- uant from an NaOH wash (humic compounds) were collected, concentrated by vortex evap- oration to 1 -ml volumes, dried onto glass-fiber filters, and radioassayed by combustion and trapping of 14C02 as described above. 14C bud- gets were determined and microbial mineral- ization was calculated by difference.
In these utilization experiments we were concerned about the possibility that the bio- logical availability of the DOC might be al- tered during the humic isolation procedure, for example due to pH manipulations or the loss of a fraction of the DOC on the XAD-8 resin. However, we have demonstrated previously that the sum of the bacterial growth on humic and nonhumic fractions obtained by this method equals the total growth in unfraction- ated DOC (Moran and Hodson 1990a), sug- gesting that alterations, if any, were minor.
Bacterial counts were performed on form- aldehyde-preserved samples filtered onto pre-
This content downloaded from 188.72.126.55 on Tue, 17 Jun 2014 23:55:42 PMAll use subject to JSTOR Terms and Conditions

Coastal humic substances 765
blackened 0.2-Am pore-size Nuclepore filters and stained with 0.01% acridine orange for 3 min. Numbers of bacterial cells in inoculated bottles averaged 1 x 107 ml-'; numbers of cells in uninoculated controls were indistin- guishable from those on background filters prepared with filter-sterilized distilled water instead of sample.
Lignin phenol analysis-Seawater samples (38 liters) were collected at 1-2-m depths on the continental shelf of the southeastern U.S. (between 30-32?N and 80-81?W) during a cruise on the RV Blue Fin in June 1990. Water was immediately passed through ashed, 293- mm-diameter Gelman A/E glass-fiber filters (1.6-gm approximate pore size) and acidified to pH 2 with 6 N HCI. Subsamples of water were removed and frozen for later determi- nation of DOC concentration with a Shimadzu TOC-500 carbon analyzer.
Humic substances were isolated by pumping seawater (50 ml min-') through a 2.2- x 40- cm column of Amberlite XAD-8 resin, eluting with 0.1 N NaOH, and passing the eluant through a BioRad AG-MP50 cation exchange resin (Moran et al. 1991 a). Humic materials were concentrated by rotary evaporation, freeze-dried, and stored frozen until oxidation. Carbon content of humic substances was de- termined on a Perkin-Elmer 240C CHN an- alyzer.
Lignin phenols were quantified by gas chro- matographic analysis of cupric oxide oxidation products of humic substances (Ertel et al. 1984). The oxidation procedure involved reacting 5- 30 mg of freeze-dried humic substances with alkaline cupric oxide for 3 h at 170?C. Three lignin phenols (vanillic acid, vanillin, and ace- tovanillon) were quantified on an HP 5890 gas chromatograph with an Alltech SE-30 and a J&W DB- 1701 silica capillary column, and lig- nin phenol concentrations were normalized based on vanillic acid concentrations (Moran et al. 1991 a). Ratios of the concentrations of the three phenols to one another (acid: alde- hyde; ketone: aldehyde; ketone: acid) were consistent among the samples. Reproducibil- ity of oxidation and GC analysis was 11 ?4% (Moran et al. 199 la).
Results Formation of humic DOC- Measurement
of radiolabel in dissolved organic compounds after 6 weeks of S. alterniflora decomposition
indicated that 1 .2?0. 1% (n = 2) of the carbon from particulate plant detritus and 3.6 ?0.6% (n = 2) of the carbon specifically from the lig- nin fraction of the detritus had accumulated in soluble form. Based on our previous mea- surements of the ratio of DOC accumulation to particulate plant detritus loss (Moran and Hodson 1990b), we estimate that 25% of the bulk lignocellulose was degraded during this time period, while 18% of the lignin compo- nent was degraded.
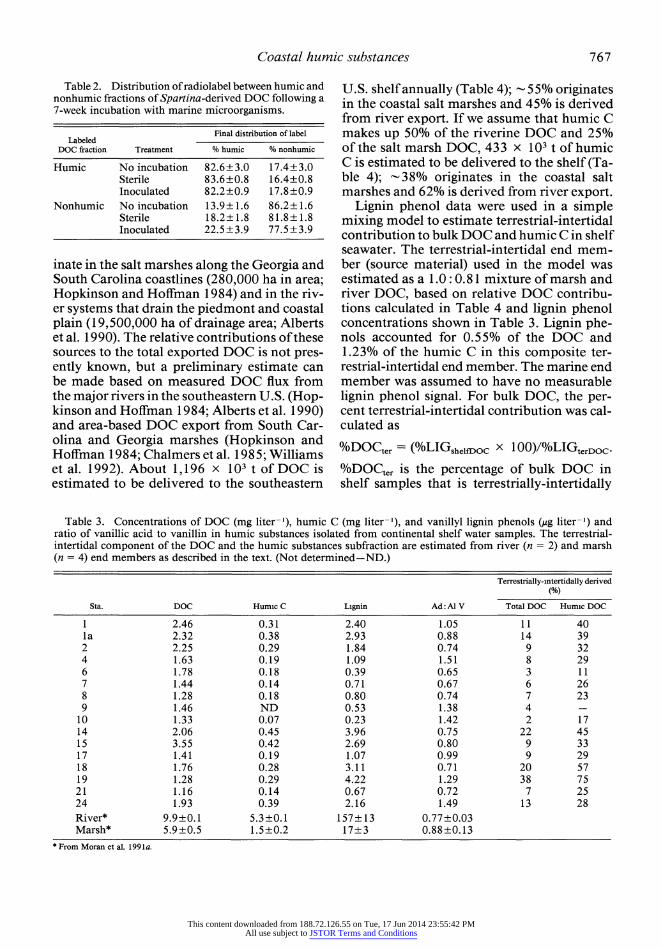
Water from the microcosms containing the newly formed radiolabeled DOC was passed through an XAD-8 resin to determine the per- cent of labeled compounds fitting the opera- tional definition of humic substances. An av- erage of 34.4% of the solubilized radiolabel was associated with the humic fraction (Fig. 1). Percent humic C was also determined based on analysis of DOC concentrations in the el- uants from the XAD-8 resin; 37.4?4.8% (n = 4) of the DOC was humic, in good agreement with the radioactivity-based measurement. From measurements of the radioactivity in the accumulated DOC and the specific activity of the uniformly labeled plant lignocellulose (68,889 dpm mg-'), we calculated that 11.7 mg of DOC accumulated per gram of initial Spartina detritus during 6 weeks of decom- position, of which 4.0 mg g- 1 were humic sub- stances.
The importance of lignin as a source ma- terial for humic DOC was suggested by the high percentage of lignin-derived DOC (73%) which fit the operational definition of humic substances (Fig. 1). Based on the specific ac- tivity of the lignin fraction of the lignin-labeled lignocellulose (55,420 dpm mg-'), we calcu- lated that lignin was the source of 2.6 mg hu- mic substances g-1 Spartina detritus, or 65% of the accumulated humic material (Fig. 1). Lignin was the source of only 12% of the non- humic DOC.
Utilization of humic DOC-Microbial up- take of Spartina-derived DOC during a 7-week period was calculated as percent label incor- porated into microbial biomass (particulate material >0.2 Am in diameter, corrected for sterile controls) plus percent label mineralized (Table 1). A total of 24.4% of the humic sub- stances fraction was used, with 0.3% incor- porated into microbial biomass and 24.1% mineralized (1. 2 ? 0.3% overall assimilation ef- ficiency). A total of 39.2% of the nonhumic
This content downloaded from 188.72.126.55 on Tue, 17 Jun 2014 23:55:42 PMAll use subject to JSTOR Terms and Conditions

766 Moran and Hodson
Nonhumic lignin-derived
Humic 79 lignin-derived
_____ 11.9 Humic 158 1 polysaccharide-derived
Nonhumic polysaccharide-derived
Fig. 1. Classification and source material for dissolved organic matter accumulating during decomposition of Spartina afterniflora from a coastal salt marsh. Values indicate the percentage of total DOC represented by each fraction.
DOC was used, with 1.6% incorporated into biomass and 37.3% mineralized (4.3?0.7% assimilation efficiency). These percentages should be considered minimum estimates, however, since some lignocellulose degrada- tion products would also have been taken up during the initial 6-week formation period, prior to the start of the utilization experiments.
[14C]DOC remaining after the 7 weeks of microbial processing was analyzed to deter- mine the distribution of radiolabel between the humic and nonhumic fractions. While 15- 20% of the label in either fraction was asso- ciated with the alternate fraction after incu- bation, this same percent interconversion was also found in sterile controls (Table 2). For DOC refractionated on an XAD-8 column without an incubation, the percent intercon- version was also similar, suggesting that the shifting of radiolabel between fractions may be the result of slight differences in the con- ditions of chromatography with the XAD-8 resin or a consequence of storage. However,
the assignment of vascular plant-derived DOC into humic or nonhumic categories is appar- ently not subject to change, due to rapid bio- logically mediated interconversions.
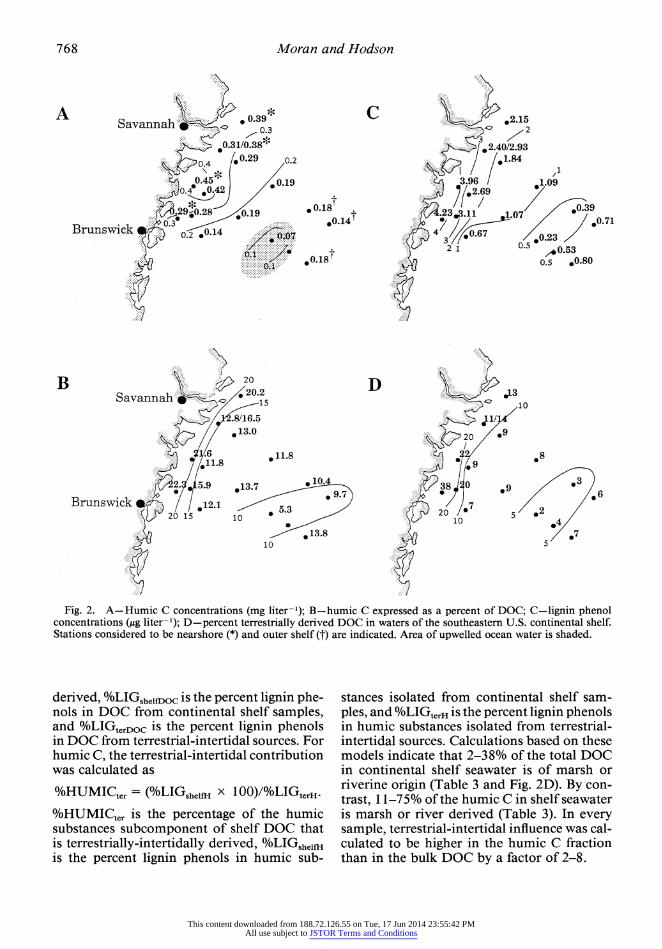
Humic substances on the continental shelf- Concentrations of humic carbon in surface seawater of the southeastern U.S. continental shelf ranged from 0.07 to 0.45 mg C liter-' (Table 3). In the nearshore region of the shelf, humic substances were present in higher con- centrations than on the outer shelf (Fig. 2A) and accounted for a greater percentage of the total DOC (19 vs. 12%; Fig. 2B). A region of seawater with humic C concentrations lower than typically found (shaded portion of Fig. 2A) was present on the midshelf. Temperature and salinity profiles in the region suggested an area of upwelled water of oceanic origin (L. R. Pomeroy pers. comm.). Lignin phenols ranged in concentrations from 0.23 to 4.2 ,ug liter-' in surface water of the shelf (Fig. 2C) and ac- counted for 0.1-0.9% of the humic C.
The lignin phenols on the shelf likely orig-
Table 1. 14C budgets for microbial utilization of DOC formed during decomposition of Spartina alterniflora detritus. Percent mineralization is calculated by difference (100 - percent recovered) for inoculated incubations. n = 3 ? 1 SD.
After 7-weeks incubation
Labeled dpm added as Percent DOC fraction Treatment [14C]DOC Filter (dpm) Filtrate (dpm) Percent recovered mineralized
Humic Sterile 8,225 213?12 8,172?283 101.9?3.3 - Inoculated 8,225 236?5* 6,007?690t 75.9?8.3 24.1
Nonhumic Sterile 13,925 252?18 13,044?129 95.5?1.0 - Inoculated 13,925 476?66t 8,263?1,212t 62.7?8.3 37.3
Measured counts in inoculated microcosms were significantly different from counts so sterile microcosms at P < 0.05 (Mann-Whitney test). t Measured counts in inoculated microcosms were significantly different from counts in sterile microcosms at P < 0.01 (Mann-Whitney test).
This content downloaded from 188.72.126.55 on Tue, 17 Jun 2014 23:55:42 PMAll use subject to JSTOR Terms and Conditions
Coastal humic substances 767
Table 2. Distribution of radiolabel between humic and nonhumic fractions of Spartina-derived DOC following a 7-week incubation with marine microorganisms.
Final distribution of label Labeled
DOC fraction Treatment % humic % nonhumic
Humic No incubation 82.6?3.0 17.4?3.0 Sterile 83.6?0.8 16.4?0.8 Inoculated 82.2?0.9 17.8?0.9
Nonhumic No incubation 13.9?1.6 86.2?1.6 Sterile 18.2?1.8 81.8?1.8 Inoculated 22.5?3.9 77.5?3.9
inate in the salt marshes along the Georgia and South Carolina coastlines (280,000 ha in area; Hopkinson and Hoffman 1984) and in the riv- er systems that drain the piedmont and coastal plain (19,500,000 ha of drainage area; Alberts et al. 1990). The relative contributions of these sources to the total exported DOC is not pres- ently known, but a preliminary estimate can be made based on measured DOC flux from the major rivers in the southeastern U.S. (Hop- kinson and Hoffman 1984; Alberts et al. 1990) and area-based DOC export from South Car- olina and Georgia marshes (Hopkinson and Hoffman 1984; Chalmers et al. 1 985; Williams et al. 1992). About 1,196 x 103 t of DOC is estimated to be delivered to the southeastern
U.S. shelf annually (Table 4); 55% originates in the coastal salt marshes and 45% is derived from river export. If we assume that humic C makes up 50% of the riverine DOC and 25% of the salt marsh DOC, 433 x 103 t of humic C is estimated to be delivered to the shelf (Ta- ble 4); 38% originates in the coastal salt marshes and 62% is derived from river export.
Lignin phenol data were used in a simple mixing model to estimate terrestrial-intertidal contribution to bulk DOC and humic C in shelf seawater. The terrestrial-intertidal end mem- ber (source material) used in the model was estimated as a 1.0: 0.81 mixture of marsh and river DOC, based on relative DOC contribu- tions calculated in Table 4 and lignin phenol concentrations shown in Table 3. Lignin phe- nols accounted for 0.55% of the DOC and 1.23% of the humic C in this composite ter- restrial-intertidal end member. The marine end member was assumed to have no measurable lignin phenol signal. For bulk DOC, the per- cent terrestrial-intertidal contribution was cal- culated as
%DOCter = (%LIGshelfDOC X l00)/%LIGterDOC.
%DOCter is the percentage of bulk DOC in shelf samples that is terrestrially-intertidally
Table 3. Concentrations of DOC (mg liter-'), humic C (mg liter-'), and vanillyl lignin phenols (,ug liter-') and ratio of vanillic acid to vanillin in humic substances isolated from continental shelf water samples. The terrestrial- intertidal component of the DOC and the humic substances subfraction are estimated from river (n = 2) and marsh (n = 4) end members as described in the text. (Not determined-ND.)
Terrestrially-intertidally derived (%)
Sta. DOC Humic C Lignin Ad: Al V Total DOC Humic DOC
1 2.46 0.31 2.40 1.05 11 40 la 2.32 0.38 2.93 0.88 14 39 2 2.25 0.29 1.84 0.74 9 32 4 1.63 0.19 1.09 1.51 8 29 6 1.78 0.18 0.39 0.65 3 11 7 1.44 0.14 0.71 0.67 6 26 8 1.28 0.18 0.80 0.74 7 23 9 1.46 ND 0.53 1.38 4 -
10 1.33 0.07 0.23 1.42 2 17 14 2.06 0.45 3.96 0.75 22 45 15 3.55 0.42 2.69 0.80 9 33 17 1.41 0.19 1.07 0.99 9 29 18 1.76 0.28 3.11 0.71 20 57 19 1.28 0.29 4.22 1.29 38 75 21 1.16 0.14 0.67 0.72 7 25 24 1.93 0.39 2.16 1.49 13 28 River* 9.9?0.1 5.3?0.1 157?13 0.77?0.03 Marsh* 5.9?0.5 1.5?0.2 17?3 0.88?0.13
* From Moran et al. 1991a.
This content downloaded from 188.72.126.55 on Tue, 17 Jun 2014 23:55:42 PMAll use subject to JSTOR Terms and Conditions
768 Moran and Hodson
A Savannah X 0.39 C *2.15 0.3 2
0.31/0.38>' *2.40/2.93
0.4 *0.29 0.2 *1.84
0.45 0 93.96 10 0o4__*0.42 t y2.69
%290.28 *01 .8.23,3.11 1.07*03 tk0.3 014t 0.71
Brunswick 402 014 00 4 *023 302
* ~~~~~~~~~2 1 .
...... 0 181 0.5 *0.80
B. ,. W20 Savannah s 1 10
. .8 /1 / *13.0 20
*11.8
2. 5.9 13.7 '10. 38 0 *9 .3
Brunswick .12.1 7 20 15 10 * 2
0 ~~~~~~~~104 9 ~~~ ~ ~~13.8 . 77
1 05
Fig. 2. A-Humic C concentrations (mg liter-'); B-humic C expressed as a percent of DOC; C-lignin phenol concentrations (,tg liter-'); D-percent terrestrially derived DOC in waters of the southeastern U.S. continental shelf. Stations considered to be nearshore (*) and outer shelf (t) are indicated. Area of upwelled ocean water is shaded.
derived, %LIGshelfDOC is the percent lignin phe- nols in DOC from continental shelf samples, and %LIGterDOC is the percent lignin phenols in DOC from terrestrial-intertidal sources. For humic C, the terrestrial-intertidal contribution was calculated as
%HUMICter = (%LIGshelfH x 100)/%LIGterH.
%HUMICter is the percentage of the humic substances subcomponent of shelf DOC that is terrestrially-intertidally derived, %LIGshelffl is the percent lignin phenols in humic sub-
stances isolated from continental shelf sam- ples, and %LIGterH is the percent lignin phenols in humic substances isolated from terrestrial- intertidal sources. Calculations based on these models indicate that 2-38% of the total DOC in continental shelf seawater is of marsh or riverine origin (Table 3 and Fig. 2D). By con- trast, 1 1-75% of the humic C in shelf seawater is marsh or river derived (Table 3). In every sample, terrestrial-intertidal influence was cal- culated to be higher in the humic C fraction than in the bulk DOC by a factor of 2-8.
This content downloaded from 188.72.126.55 on Tue, 17 Jun 2014 23:55:42 PMAll use subject to JSTOR Terms and Conditions

Coastal humic substances 769
Discussion The quantitative importance of bacterial
solubilization in the formation of DOM in aquatic environments has been demonstrated (Moran and Hodson 1989, 1990b; Smith et al. 1992). However, little is known about the chemical composition of this DOM or the eco- logical role it plays in aquatic ecosystems. In previous studies of coastal marshes of the southeastern U.S., we found evidence that the DOM formed as a result of Spartina detritus decomposition was rich in lignin byproducts and humic substances (Moran and Hodson 1989, 1990b). Uncertainty about the tropho- dynamic role of humic substances in marine environments prompted us to concentrate our attention on this chemical subcomponent of vascular plant-derived DOM.
Formation of plant-derived dissolved humic substances-Dissolved humic substances are defined in this study as those compounds which adhere to an XAD-8 resin under acidic (pH 2) conditions and back elute in base. This oper- ational definition, based on the hydrophobic- ity of humic compounds, is the most widely accepted method for identifying and isolating aquatic humic substances (Thurman 1985). A significant fraction (34%) of the dissolved ma- terial released from degrading salt marsh grass fits this definition, whether measurements are based on radioactivity isolated by XAD-8 methodology or on DOC concentrations in resin eluants. Thus, vascular plant detritus de- grading in coastal marshes can be an important source of humic substances. Furthermore, these operationally defined humic substances form quite rapidly (within a few weeks of the ini- tiation of decomposition) and therefore do not necessarily require extended biological and physical processing for their formation. In pre- vious studies, Filip and Alberts (1988) docu- mented the biologically mediated accumula- tion of humic substances from S. alterniflora detritus after 10 months of decomposition, and Fustec et al. (1989) demonstrated the forma- tion of humic substances from the lignin frac- tion of plant detritus in alluvial soils after 7 weeks. Humic substances also have been ex- tracted from partially degraded tissues of sev- eral other vascular plant species (Ertel and Hedges 1985), and they have been found in the culture medium of lignin-degrading bac- terial cultures growing on lignocellulose (Mc- Carthy et al. 1986).
Table 4. Inputs of terrestrial-intertidal DOC and hu- mic C from Georgia and South Carolina rivers and salt marshes to the southeastem U.S. continental shelf.
DOC input Humic C input
Source (103 t yr-1)
Rivers 536* (45%) 268t (62%) Marshes 660t (55%) 165? (38%)
* Average of values calculated by Hopkinson and Hoffman 1984 (626 x 103
t yr-') and Alberts et al. 1990 (445 x 103 t yr-1). t Assuming riverine DOC to be 50% humic C (Thurman 1985; Alberts et al.
1990; Moran et al. 199 la). : Estimated from Hopkinson and Hoffman 1984, Chalmers et al. 1985, and
Williams et al. 1992, assuming a 7,000-m-wide zone of salt marsh along a 400,000-m coastline (Hopkinson and Hoffman 1984). Range = 361-918 x 103 t.
? Assuming salt marsh DOC to be 25% humic C (Moran et al. 199 la).
Turnover of vascular plant-derived humic substances-The ecological role of vascular plant-derived humic substances in marine eco- systems will depend in large part on how rap- idly and efficiently the humic substances are assimilated by marine bacterioplankton. Nat- ural humic substances generally are considered to be among the most refractory components of the bulk DOC pool, although recent evi- dence from freshwater environments suggests that humic substances can support significant levels of bacterial secondary production (Mo- ran and Hodson 1990a; Tranvik 1990; Tulo- nen et al. 1992). In our experiments, the non- humic components of DOC were assimilated more readily and more efficiently by natural marine bacteria than were the humic compo- nents, although 24% of the humic substance fraction was nonetheless used in a 7-week pe- riod. These plant-derived humic materials of recent age (3 weeks average age) are apparently more labile than dissolved humic substances which have accumulated in natural environ- ments, as only a few percent (<4) of the humic substances from a South Carolina reservoir or a Georgia backwater swamp were readily as- similable by bacteria over similar time inter- vals (Moran and Hodson 1990a). Thus, sig- nificant heterogeneity of compounds included in the humic substances definition is evident. The compounds accumulating in natural en- vironments are likely to be those that are most resistant to biological attack (i.e. those having the longest turnover times), yet relatively labile compounds with ecologically relevant turn- over times are also encompassed by this broad chemical classification.
The differentially radiolabeled plant mate- rials used in these studies allowed us to deter-
This content downloaded from 188.72.126.55 on Tue, 17 Jun 2014 23:55:42 PMAll use subject to JSTOR Terms and Conditions

770 Moran and Hodson
mine the relative importance of lignin and nonlignin components of lignocellulose as source materials for humic substances. Lignin is a highly aromatic, chemically heterogeneous polymer that accounts for 10% or less of the lignocellulose in S. alterniflora (Moran and Hodson 1990a) and other species of grasses, yet it was the source material for 30% of the DOC and 65% of the dissolved humic sub- stances accumulating from degrading Spar- tina. The vascular plant polysaccharides (cel- lulose and hemicellulose) were presumably used more rapidly by bacteria; although readi- ly lost from the plant detritus during microbial degradation, a smaller proportion of polysac- charides accumulated in dissolved form. Moreover, the polysaccharide-derived com- pounds that did accumulate were much less likely to be humic matter. Lignin therefore makes a disproportionate contribution to the stable DOC accumulating from vascular plant detritus in coastal marshes.
Terrestrial-intertidal contribution to humic substances in a marine environment-The ac- cumulation of vascular plant-derived DOC in coastal marshes suggests a potential role, either ecological or geochemical, for this material in adjacent marine environments. In this study and a previous one (Moran et al. 1991 a), we found that a measurable fraction of the DOC in seawater on the southeastern U.S. shelf was of terrestrial-intertidal origin. On the basis of calculations of export from southeastern U.S. rivers and marshes (Table 4), about half the terrestrial DOC present on the shelf originates in the coastal salt marshes and half comes from river export.
Terrestrial influence is greater in the humic substances subcomponent of shelf seawater than in the bulk DOC (Table 3). This greater terrestrial component in humic substances re- sults from the mixing of humic-rich DOC in river or coastal marsh water (-50 and 25% humic substances, respectively) with the hu- mic-poor DOC in ocean water (-5-10% hu- mic material). That is, bulk DOC in terrestrial- intertidal source waters is diluted to a greater extent by bulk marine DOC than humic sub- stances in source waters are diluted by marine humic substances. Biological considerations also argue for greater terrestrial influence on marine humic substances than on bulk DOC
because natural bacterial communities appar- ently assimilate the nonhumic components of vascular plant-derived DOC more rapidly than the humic component (Table 1) and because there is no rapid biological interconversion be- tween these two pools (Table 2). Biological processing can therefore be expected to deplete nonhumic components of terrestrial-intertidal DOC relative to the humic component. The simple mixing models we used to calculate ter- restrial influence on shelf DOC do not include considerations of biologically mediated changes in DOC composition (i.e. conservative behav- ior of terrestrial material is assumed). If sig- nificant biological processing does occur and if it involves discrimination against the humic substances fraction, the disparity in terrestrial influence between bulk DOC and the humic subcomponent would be greater than that es- timated by the model.
Quantification of the terrestrial component of marine DOC and measurement of the turn- over time of terrestrial material in the marine environment are topics of special interest for reconciling existing discrepancies in the global carbon budget (Williams and Druffel 1987; Hedges 1992). Meyers-Schulte and Hedges (1986) used lignin phenol concentrations in marine humic substances to estimate that DOC in open-ocean water is 1% terrestrially de- rived. Because our results indicate greater ter- restrial influence on humic substances than on bulk DOC in marine environments, there is unlikely to be a significant but previously un- discovered pool of terrestrially derived mate- rial in the nonhumic component of marine DOC which would greatly alter this estimate. As expected, terrestrial-intertidal contribu- tions to the marine DOC pool are higher in coastal ocean environments than in the open ocean; we estimate that 2-38% (this study) and 3-36% (Moran et al. 1991a) of DOC in sea- water on the southeastern U.S. continental shelf is of terrestrial-intertidal origin. The present study also suggests that intertidal environ- ments can be an important source of DOC, and of humic C in particular, to the ocean, yet many current global carbon budgets consider only the riverine export of terrestrial DOC (Meybeck 1982; Mantoura and Woodward 1983). DOC entering marine environments from intertidal sources may be dissimilar in
This content downloaded from 188.72.126.55 on Tue, 17 Jun 2014 23:55:42 PMAll use subject to JSTOR Terms and Conditions

Coastal humic substances 771
composition, 14C age, or biological lability to riverine DOC, and these factors may influence its ecological and geochemical role in the ocean.
References ALBERTS, J. J., J. R. ERTEL, AND L. CASE. 1990. Char-
acterization of organic matter in rivers of the south- eastern United States. Int. Ver. Theor. Angew. Lim- nol. Verh. 24: 260-262.
AZAM, F., AND R. E. HODSON. 1977. Dissolved ATP in the sea and its utilization by marine bacteria. Nature 267: 696-698.
BAUER, J. E., P. M. WILLIAMS, AND E. R. M. DRUFFEL. 1992. '4C activity of dissolved organic carbon frac- tions in the north-central Pacific and Sargasso Sea. Nature 357: 667-670.
BENNER, R., A. E. MACCUBBIN, AND R. E. HODSON. 1984. Preparation, characterization, and microbial degra- dation of specifically radiolabeled 14C-lignocelluloses from marine and freshwater macrophytes. Appl. En- viron. Microbiol. 47: 381-389.
CHALMERS, A. G., R. G. WIEGERT, AND P. L. WOLF. 1985. Carbon balance in a salt marsh: Interactions of dif- fusive export, tidal deposition, and rainfall-caused erosion. Estuarine Coastal Shelf Sci. 21: 757-771.
ERTEL, J. R., AND J. I. HEDGES. 1985. Sources of sedi- mentary humic substances: Vascular plant debris. Geochim. Cosmochim. Acta 49: 2097-2107.
9 , AND E. M. PERDUE. 1984. Lignin sig- nature of aquatic humic substances. Science 223: 485- 487.
FILIP, Z., AND J. J. ALBERTS. 1988. The release of humic substances from Spartina alterniflora (Loisel.) into sea water as influenced by salt marsh indigenous micro- organisms. Sci. Total Environ. 73: 143-157.
FUSTEC, E., E. CHAUVET, AND G. GAS. 1989. Lignin deg- radation and humus formation in alluvial soils and sediments. Appl. Environ. Microbiol. 55: 922-926.
HEDGES, J. I. 1992. Global biogeochemical cycles: Prog- ress and problems. Mar. Chem. 39: 67-93.
HODSON, R. E., R. R. CHRISTIAN, AND A. E. MACCUBBIN. 1984. Lignocellulose and lignin in the salt marsh grass Spartina alterniflora: Initial concentrations and short-term post-depositional changes in detrital ma- terial. Mar. Biol. 81: 1-7.
HOPKINSON, C. S., AND F. A. HoFFMAN. 1984. The es- tuary extended-a recipient-system study of estuarine outwelling in Georgia, p. 313-330. In V. S. Kennedy [ed.], The estuary as a filter. Academic.
MCCARTHY, A. J., A. PATERSON, AND P. BRODA. 1986. Lignin solubilisation by Thermomonospora meso- phila. Appl. Microbiol. Biotechnol. 24: 347-352.
MANTouRA, R. F. C., AND E. M. S. WOODWARD. 1983. Conservative behavior of riverine dissolved organic carbon in the Severn estuary: Chemical and geological
implications. Geochim. Cosmochim. Acta 47: 1293- 1309.
MEYBECK, M. 1982. Carbon, nitrogen, and phosphorus transport by world rivers. Am. J. Sci. 282: 401-450.
MEYERS-SCHULTE, K. J., AND J. I. HEDGES. 1986. Mo- lecular evidence for a terrestrial component of organic matter dissolved in ocean water. Nature 321: 61-63.
MORAN, M. A., AND R. E. HODSON. 1989. Formation and bacterial utilization of dissolved organic carbon derived from detrital lignocellulose. Limnol. Ocean- ogr. 34: 1034-1047.
, AND . 1990a. Bacterial production on hu- mic and nonhumic components of dissolved organic carbon. Limnol. Oceanogr. 35: 1744-1756.
, AND . 1990b. Contributions of degrading Spartina alterniflora lignocellulose to the dissolved organic carbon pool of a salt marsh. Mar. Ecol. Prog. Ser. 62: 161-168.
, L. R. POMEROY, E. S. SHEPPARD, L. P. ATKINSON, AND R. E. HODSON. 1991 a. Distribution of terres- trially derived dissolved organic matter on the south- eastern U.S. continental shelf. Limnol. Oceanogr. 36: 1134-1149.
, R. J. WICKS, AND R. E. HODSON. 199 lb. Export of dissolved organic matter from a mangrove swamp ecosystem: Evidence from natural fluorescence, dis- solved lignin phenols, and bacterial secondary pro- duction. Mar. Ecol. Prog. Ser. 76: 175-184.
SMITH, D. C., M. SIMON, A. L. ALLDREDGE, AND F. AzAM. 1992. Intense hydrolytic enzyme activity on marine aggregates and implications for rapid particle disso- lution. Nature 359: 139-142.
THuRMAN, E. M. 1985. Organic geochemistry of natural waters. Nijhoff/Junk.
TRANVIK, L. J. 1990. Bacterioplankton growth on frac- tions of dissolved organic carbon of different molec- ular weights from humic and clear waters. Appl. En- viron. Microbiol. 56: 1672-1677.
TULONEN, T., K. S. SALONEN, AND L. ARVOLA. 1992. Ef- fects of different molecular weight fractions of dis- solved organic matter on the growth of bacteria, algae and protozoa from a highly humic lake. Hydrobio- logia 229: 239-252.
WILLiAMS, P. M., AND E. R. M. DRUFFEL. 1987. Radio- carbon in dissolved organic matter in the central North Pacific Ocean. Nature 330: 246-248.
WILLiAMS, T. M., T. G. WOLAVER, R. F. DAME, AND J. D. SPURRIER. 1992. The Bly Creek ecosystem study- organic carbon transport within a euhaline salt marsh basin, North Inlet, South Carolina. J. Exp. Mar. Biol. Ecol. 163: 125-139.
WRIGHT, R. T., AND J. E. HOBBIE. 1966. Use of glucose and acetate by bacteria and algae in aquatic ecosys- tems. Ecology 47: 447-464.
Submitted: 22 March 1993 Accepted. 27 October 1993
Amended: 18 November 1993
This content downloaded from 188.72.126.55 on Tue, 17 Jun 2014 23:55:42 PMAll use subject to JSTOR Terms and Conditions