dissipation of polycyclic aromatic hydrocarbons (pahs) in soil microcosms amended with mushroom...
TRANSCRIPT

at SciVerse ScienceDirect
Soil Biology & Biochemistry 47 (2012) 191e197
Contents lists available
Soil Biology & Biochemistry
journal homepage: www.elsevier .com/locate/soi lb io
Dissipation of polycyclic aromatic hydrocarbons (PAHs) in soil microcosmsamended with mushroom cultivation substrate
Xuanzhen Li a,b,1, Yucheng Wu a,1, Xiangui Lin a,*, Jing Zhang a, Jun Zeng a
a State Key Laboratory of Soil and Sustainable Agriculture, Institute of Soil Science, Chinese Academy of Sciences, 71 East Beijing Road, Nanjing 210008, ChinabCollege of Biological and Environmental Engineering, Zhejiang University of Technology, 18 Chaowang Road, Hangzhou 310032, China
a r t i c l e i n f o
Article history:Received 8 October 2011Received in revised form28 November 2011Accepted 2 January 2012Available online 12 January 2012
Keywords:BiostimulationFungiLaccasePolycyclic aromatic hydrocarbonMicrocosm
Abbreviations: MCS, mushroom cultivation substrhydrocarbon; DGGE, denaturing gradient gel electhydrocarbon degraders.* Corresponding author. Tel.: þ86 25 86881589; fax
E-mail address: [email protected] (X. Lin).1 These authors contributed equally to this work.
0038-0717/$ e see front matter � 2012 Elsevier Ltd.doi:10.1016/j.soilbio.2012.01.001
a b s t r a c t
The potential of mushroom cultivation substrate (MCS) in bioremediation was examined in polycyclicaromatic hydrocarbon (PAH)-contaminated soil. After a 60-day incubation, 32.9% dissipation of the 15studied PAHs was observed in MCS-amended microcosms, with anthracene, benzo(a)pyrene and ben-zo(a)anthracene being the most degradable PAHs. MCS significantly increased the abundance andchanged the community compositions of bacteria, fungi and aromatic hydrocarbon degraders. Twospecies belonging to the Sordariomycetes of the Ascomycota were enriched in all MCS-treated soilsamples, and coupled with the unique changes in the PAH profile, this implies the involvement oflaccase-like enzymes. Limited improvement was observed after adding Pleurotus ostreatus, possiblybecause of its poor colonization of the soil. In addition, alfalfa appeared to antagonize the bioremediationeffects of MCS. The results of this study suggest that MCS can be a cost-effective and green biostimulationagent, thereby providing support for the development of MCS-based biostimulation ofPAH-contaminated soil.
� 2012 Elsevier Ltd. All rights reserved.
1. Introduction
Polycyclic aromatic hydrocarbons (PAHs) are a group of organiccompounds with two or more fused benzene rings and have beenidentified as one of the major persistent organic pollutants. Bothnatural and anthropogenic sources, such as forest fires, volcaniceruptions and fossil fuel combustions, contribute to the amount ofPAHs present in the environment. PAHs are routinely found in soilsthroughout the world at various concentrations, posing potentialrisks to human and ecological health owing to their toxicity andcarcinogenic properties (Joseph, 1997).
To date, many techniques are available to reduce PAHs in soil(Gan et al., 2009). One of promising strategies is bioremediation,which is often carried out by introducing allochthonous strains(so-called bioaugmentation) normally isolated or enriched in thelaboratory to enhance the PAH degradation activity in soil. Previousstudies have mainly focused on bacterial bioaugmentation, butfungi are attractive options in this process (Harms et al., 2011).Many ligninolytic fungi, mostly wood rot fungi, are potent PAH
ate; PAH, polycyclic aromaticrophoresis; AHDs, aromatic
: þ86 25 86881000.
All rights reserved.
degraders (Baldrian et al., 2000). The ability of fungi to degradePAHs may be related either to their cytochrome P-450 enzymes, orto the extracellular ligninolytic enzymes, including lignin peroxi-dase, manganese peroxidase and laccase (Cerniglia, 1997). Thepotential of wood rot fungi in soil remediation has been evaluatedwidely in recently years (Baldrian, 2008; Harms et al., 2011).
There are still some limitations on fungal bioaugmentation. Oneof the most important considerations is the colonization capacity offungi in soil. Once introduced to the soil, allochthonous fungi areexposed to a hostile environment with less available nutrients thantheir natural habitats, as well as competition with native commu-nities (Baldrian, 2008). As such, biostimulation (i.e., the addition ofnutrients to promote the activity of indigenous microorganisms)has been identified as an alternative method to bioaugmentation(Baldrian, 2008).
Many substrates can stimulate the fungal degradation of organicpollutants in soil. One of the commonly used substrates is spentmushroom compost, which has shown potential for soil bioreme-diation (Lau et al., 2003). Spent mushroom compost provides bulknutrients for indigenous soil microflora and contains considerablemicrobial activity (Ribas et al., 2009), and thus combines theadvantages of both biostimulation and bioaugmentation. However,spentmushroomcompostmight not be readily available at all times.In contrast, mushroom cultivation substrate (MCS), which containsnormal, inexpensive agricultural byproducts and has a similar

Table 1Experimental treatments.
Treatment Abbreviation MCS P. ostreatus Alfalfa
Control CK e e e
Substrate (MCS)amendment
S þ e e
Substrate and P. ostreatus SP þ þ e
Substrate, P. ostreatusand alfalfa
SPA þ þ þ
Alfalfa A e e þ
X. Li et al. / Soil Biology & Biochemistry 47 (2012) 191e197192
nutrient composition to spent mushroom compost, is a candidatesubstrate for the biostimulation of PAH-contaminated soil.
Plants are usually introduced to enhance PAH removal bystimulating indigenous microbes in the soil (Olson et al., 2007). Forexample, alfalfa increased the abundance of PAH degraders as wellas the dissipation of hydrocarbons in soil (Phillips et al., 2006). Itshould be noted that the combination of multiple strategies addscomplexity to themicrobial degradation of PAHs in soil. As such, thesuccession of microbial communities in soil receiving bio-stimulation, bioaugmentation and/or phytoremediation providesinsight regarding the mechanisms used to degrade pollutants.
In this study, we examined the effects of one type of MCS on soilmicrobial communities and PAH degradation in aged, contami-nated soil on a laboratory scale. At the same time, combinations ofMCS, fungal inoculation (Pleurotus ostreatus) and/or plantation ofalfalfa were compared in terms of PAH dissipation and microbialeffects. The aims of this study were (1) to evaluate the potential ofbiostimulation with MCS in soil remediation and (2) to provideinsight into the possible mechanisms responsible for the degrada-tion of PAHs in soil.
2. Materials and methods
2.1. Soil
Contaminated soil was collected from a petroleum gas stationthat has operated for more than 20 years inWuxi, Jiangsu Province,China (30�3601400N, 120�2803300E). The physicochemical propertiesof the soil have been detailed previously (Li et al., 2010a). In total,the concentration of 15 EPA priority PAHs (except acenaphthene) inthe soil was 14 556 mg kg�1, which is almost 37-fold higher than theaverage value in other soils from this area according to a soil survey(Ping et al., 2007). After sampling, the soil was air-dried, sieved(2 mm) and stored at 4 �C in the dark.
2.2. Mushroom cultivation substrate
The MCS used in this study consisted of crushed corncob (60%),wheat bran (30%), cattle manure (7%), sucrose (1%), superphos-phate (1%), urea (0.5%) and gypsum (0.5%). The MCS was wellmixed, adjusted to 65% moisture with distilled water and sterilizedat 121 �C for 1 h.
2.3. Fungus
The wood rot fungus P. ostreatus was acquired from the culturecollection of our laboratory and maintained on potato-dextroseagar slants. For soil remediation, MCS was inoculated withP. ostreatus and incubated in flasks at 30 �C for 20 days. The myceliaand substrate were mixed well and used as the fungal inoculum.
2.4. Plant
Seed of alfalfa (Medicago sativa L.) were acquired from NanjingAgriculture University Seed Company. The seeds were treated with10% hydrogen peroxide for 10 min and rinsed with sterile distilledwater. After germination and growth for 7 days in moist perlite, theseeds were transplanted to separate pots.
2.5. Soil microcosms
Microcosms consisted of 2 kg of air-dried soil in plastic pots, andthe moisture of soil was adjusted to 50% water holding capacity(WHC) with sterile distilled water at the beginning of the incuba-tion. Five treatment groups, including the control, were established
in triplicate as shown in Table 1. The MCS with or withoutP. ostreatus was spiked into soil at a concentration of 5% (w/w).
After a 60-day incubation in a green house, a 50-g subsamplewas removed from each soil microcosm and stored at �20 �C formolecular analysis. The remaining soil was used to enumerate themicrobes or for a chemical analysis. Soil organic carbon wasdetermined using the K2Cr2O7 oxidation-reduction titrationmethod. Total nitrogen was measured following the Kjeldahlmethod. The pHwas determined with a soil/KCl mixture (Lu, 2000).
2.6. Extraction and determination of soil PAHs
Soxhlet extraction was performed to retrieve PAHs from soil asdescribed previously (Li et al., 2010a). Briefly, PAHs from 2 g ofair-dried soil sample were extracted using 60 ml of dichloro-methane in a Soxhlet apparatus for 24 h. The extracts wereconcentrated with a rotary evaporator and purified with a chro-matography column filled with activated silica gel. The concentra-tions of 15 EPA priority PAHs were determined using an Ultra FastLiquid Chromatograph system (UFLC-20, Shimadzu, Japan), asdescribed previously (Li et al., 2010a). All PAH concentrations arepresented on the basis of soil dry weight.
2.7. Enumeration of soil microorganisms
Soil bacteria and fungi were enumerated at the end of theincubation using cultivation-based plate counting methods.Aromatic hydrocarbon degraders (AHDs) were evaluated usinga miniaturized most probable number (MPN) method in 96-wellmicroplates with five replicates per dilution (Wrenn and Venosa,1996). Briefly, phenanthrene, anthracene, fluorene and dibenzo-thiophene were added collectively as the sole carbon sources tosupport the proliferation of AHDs. Serially diluted samples wereinoculated into the wells, and the microplates were incubated at28 �C for 3 weeks. Color development in the wells due to theaccumulation of partial oxidation products of aromatic substrateswere considered positive for the presence of AHDs. Published MPNtables were used to determine the MPN.
2.8. DNA extraction and polymerase chain reaction (PCR)amplification
DNA was extracted from approximately 0.5 g of fresh soil usingthe FastDNA SPIN kit for soil (MP Biomedicals) following themanufacturer’s instructions. The quality and quantity of DNA werechecked using a NanoDrop spectrophotometer (NanoDrop Tech-nologies Inc., Wilmington, USA).
Bacterial 16S rRNA and fungal 18S rRNA genes were amplifiedwith primer sets 341F-GC/518R (Muyzer et al., 1993) and FR1-GC/FF390 (Vainio and Hantula, 2000), respectively. The 25-ml PCRreaction mixtures contained 2.5 ml of 10� PCR buffer, 0.25 mM ofeach primer, 200 mM of each deoxyribonucleoside triphosphate,1.5 U of Taq DNA polymerase, and 0.5 ml of soil DNA. The PCR was
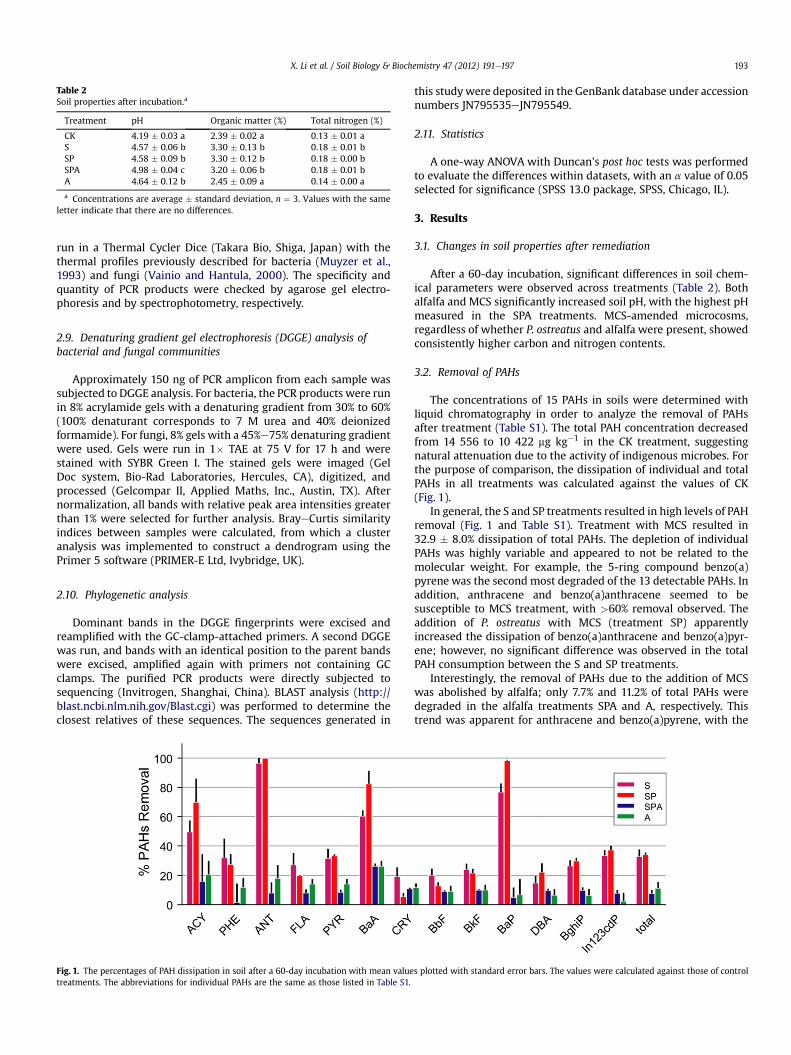
Table 2Soil properties after incubation.a
Treatment pH Organic matter (%) Total nitrogen (%)
CK 4.19 � 0.03 a 2.39 � 0.02 a 0.13 � 0.01 aS 4.57 � 0.06 b 3.30 � 0.13 b 0.18 � 0.01 bSP 4.58 � 0.09 b 3.30 � 0.12 b 0.18 � 0.00 bSPA 4.98 � 0.04 c 3.20 � 0.06 b 0.18 � 0.01 bA 4.64 � 0.12 b 2.45 � 0.09 a 0.14 � 0.00 a
a Concentrations are average � standard deviation, n ¼ 3. Values with the sameletter indicate that there are no differences.
X. Li et al. / Soil Biology & Biochemistry 47 (2012) 191e197 193
run in a Thermal Cycler Dice (Takara Bio, Shiga, Japan) with thethermal profiles previously described for bacteria (Muyzer et al.,1993) and fungi (Vainio and Hantula, 2000). The specificity andquantity of PCR products were checked by agarose gel electro-phoresis and by spectrophotometry, respectively.
2.9. Denaturing gradient gel electrophoresis (DGGE) analysis ofbacterial and fungal communities
Approximately 150 ng of PCR amplicon from each sample wassubjected to DGGE analysis. For bacteria, the PCR products were runin 8% acrylamide gels with a denaturing gradient from 30% to 60%(100% denaturant corresponds to 7 M urea and 40% deionizedformamide). For fungi, 8% gels with a 45%e75% denaturing gradientwere used. Gels were run in 1� TAE at 75 V for 17 h and werestained with SYBR Green I. The stained gels were imaged (GelDoc system, Bio-Rad Laboratories, Hercules, CA), digitized, andprocessed (Gelcompar II, Applied Maths, Inc., Austin, TX). Afternormalization, all bands with relative peak area intensities greaterthan 1% were selected for further analysis. BrayeCurtis similarityindices between samples were calculated, from which a clusteranalysis was implemented to construct a dendrogram using thePrimer 5 software (PRIMER-E Ltd, Ivybridge, UK).
2.10. Phylogenetic analysis
Dominant bands in the DGGE fingerprints were excised andreamplified with the GC-clamp-attached primers. A second DGGEwas run, and bands with an identical position to the parent bandswere excised, amplified again with primers not containing GCclamps. The purified PCR products were directly subjected tosequencing (Invitrogen, Shanghai, China). BLAST analysis (http://blast.ncbi.nlm.nih.gov/Blast.cgi) was performed to determine theclosest relatives of these sequences. The sequences generated in
Fig. 1. The percentages of PAH dissipation in soil after a 60-day incubation with mean valuetreatments. The abbreviations for individual PAHs are the same as those listed in Table S1.
this studywere deposited in the GenBank database under accessionnumbers JN795535eJN795549.
2.11. Statistics
A one-way ANOVA with Duncan’s post hoc tests was performedto evaluate the differences within datasets, with an a value of 0.05selected for significance (SPSS 13.0 package, SPSS, Chicago, IL).
3. Results
3.1. Changes in soil properties after remediation
After a 60-day incubation, significant differences in soil chem-ical parameters were observed across treatments (Table 2). Bothalfalfa and MCS significantly increased soil pH, with the highest pHmeasured in the SPA treatments. MCS-amended microcosms,regardless of whether P. ostreatus and alfalfa were present, showedconsistently higher carbon and nitrogen contents.
3.2. Removal of PAHs
The concentrations of 15 PAHs in soils were determined withliquid chromatography in order to analyze the removal of PAHsafter treatment (Table S1). The total PAH concentration decreasedfrom 14 556 to 10 422 mg kg�1 in the CK treatment, suggestingnatural attenuation due to the activity of indigenous microbes. Forthe purpose of comparison, the dissipation of individual and totalPAHs in all treatments was calculated against the values of CK(Fig. 1).
In general, the S and SP treatments resulted in high levels of PAHremoval (Fig. 1 and Table S1). Treatment with MCS resulted in32.9 � 8.0% dissipation of total PAHs. The depletion of individualPAHs was highly variable and appeared to not be related to themolecular weight. For example, the 5-ring compound benzo(a)pyrene was the second most degraded of the 13 detectable PAHs. Inaddition, anthracene and benzo(a)anthracene seemed to besusceptible to MCS treatment, with >60% removal observed. Theaddition of P. ostreatus with MCS (treatment SP) apparentlyincreased the dissipation of benzo(a)anthracene and benzo(a)pyr-ene; however, no significant difference was observed in the totalPAH consumption between the S and SP treatments.
Interestingly, the removal of PAHs due to the addition of MCSwas abolished by alfalfa; only 7.7% and 11.2% of total PAHs weredegraded in the alfalfa treatments SPA and A, respectively. Thistrend was apparent for anthracene and benzo(a)pyrene, with the
s plotted with standard error bars. The values were calculated against those of control
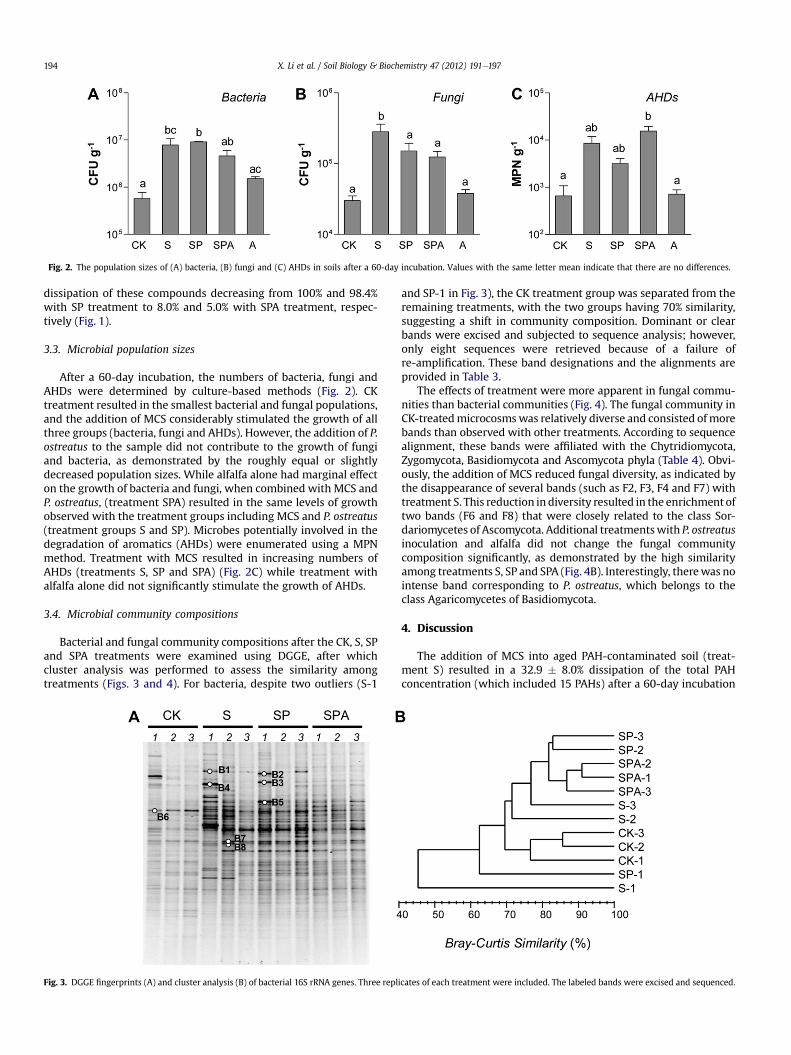
Fig. 2. The population sizes of (A) bacteria, (B) fungi and (C) AHDs in soils after a 60-day incubation. Values with the same letter mean indicate that there are no differences.
X. Li et al. / Soil Biology & Biochemistry 47 (2012) 191e197194
dissipation of these compounds decreasing from 100% and 98.4%with SP treatment to 8.0% and 5.0% with SPA treatment, respec-tively (Fig. 1).
3.3. Microbial population sizes
After a 60-day incubation, the numbers of bacteria, fungi andAHDs were determined by culture-based methods (Fig. 2). CKtreatment resulted in the smallest bacterial and fungal populations,and the addition of MCS considerably stimulated the growth of allthree groups (bacteria, fungi and AHDs). However, the addition of P.ostreatus to the sample did not contribute to the growth of fungiand bacteria, as demonstrated by the roughly equal or slightlydecreased population sizes. While alfalfa alone had marginal effecton the growth of bacteria and fungi, when combined with MCS andP. ostreatus, (treatment SPA) resulted in the same levels of growthobserved with the treatment groups including MCS and P. ostreatus(treatment groups S and SP). Microbes potentially involved in thedegradation of aromatics (AHDs) were enumerated using a MPNmethod. Treatment with MCS resulted in increasing numbers ofAHDs (treatments S, SP and SPA) (Fig. 2C) while treatment withalfalfa alone did not significantly stimulate the growth of AHDs.
3.4. Microbial community compositions
Bacterial and fungal community compositions after the CK, S, SPand SPA treatments were examined using DGGE, after whichcluster analysis was performed to assess the similarity amongtreatments (Figs. 3 and 4). For bacteria, despite two outliers (S-1
Fig. 3. DGGE fingerprints (A) and cluster analysis (B) of bacterial 16S rRNA genes. Three repl
and SP-1 in Fig. 3), the CK treatment group was separated from theremaining treatments, with the two groups having 70% similarity,suggesting a shift in community composition. Dominant or clearbands were excised and subjected to sequence analysis; however,only eight sequences were retrieved because of a failure ofre-amplification. These band designations and the alignments areprovided in Table 3.
The effects of treatment were more apparent in fungal commu-nities than bacterial communities (Fig. 4). The fungal community inCK-treatedmicrocosmswas relatively diverse and consisted of morebands than observed with other treatments. According to sequencealignment, these bands were affiliated with the Chytridiomycota,Zygomycota, Basidiomycota and Ascomycota phyla (Table 4). Obvi-ously, the addition of MCS reduced fungal diversity, as indicated bythe disappearance of several bands (such as F2, F3, F4 and F7) withtreatment S. This reduction in diversity resulted in the enrichment oftwo bands (F6 and F8) that were closely related to the class Sor-dariomycetes of Ascomycota. Additional treatmentswith P. ostreatusinoculation and alfalfa did not change the fungal communitycomposition significantly, as demonstrated by the high similarityamong treatments S, SP and SPA (Fig. 4B). Interestingly, therewas nointense band corresponding to P. ostreatus, which belongs to theclass Agaricomycetes of Basidiomycota.
4. Discussion
The addition of MCS into aged PAH-contaminated soil (treat-ment S) resulted in a 32.9 � 8.0% dissipation of the total PAHconcentration (which included 15 PAHs) after a 60-day incubation
icates of each treatment were included. The labeled bands were excised and sequenced.
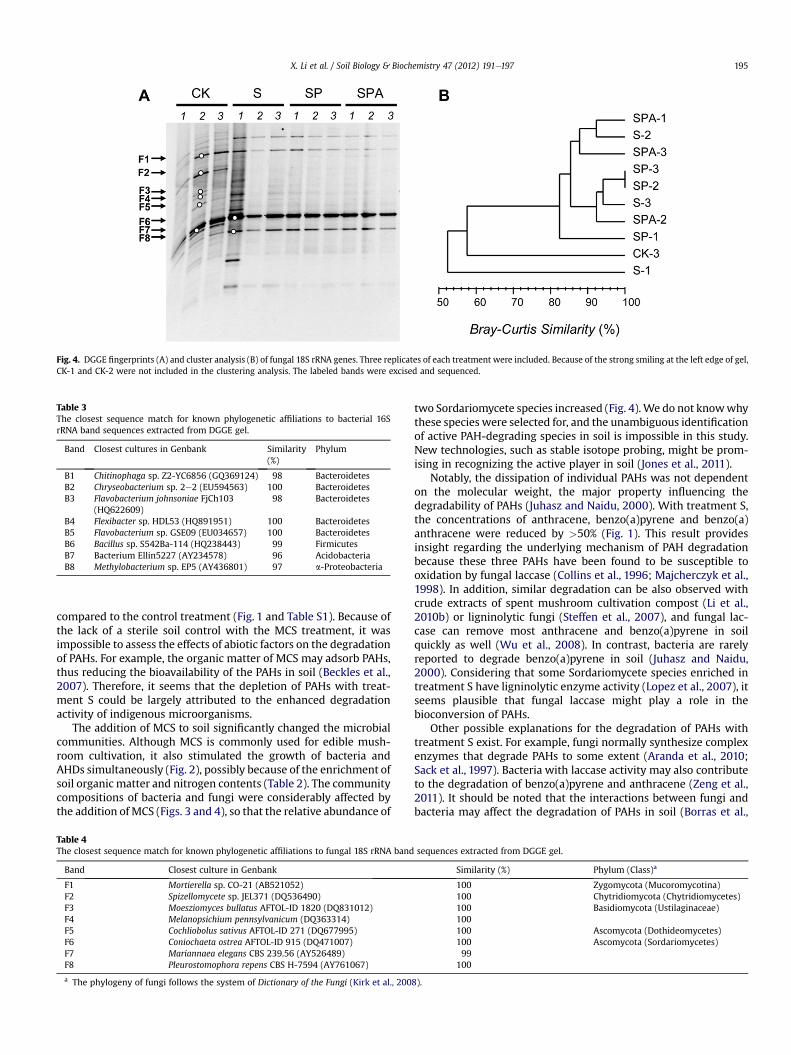
Fig. 4. DGGE fingerprints (A) and cluster analysis (B) of fungal 18S rRNA genes. Three replicates of each treatment were included. Because of the strong smiling at the left edge of gel,CK-1 and CK-2 were not included in the clustering analysis. The labeled bands were excised and sequenced.
Table 3The closest sequence match for known phylogenetic affiliations to bacterial 16SrRNA band sequences extracted from DGGE gel.
Band Closest cultures in Genbank Similarity(%)
Phylum
B1 Chitinophaga sp. Z2-YC6856 (GQ369124) 98 BacteroidetesB2 Chryseobacterium sp. 2e2 (EU594563) 100 BacteroidetesB3 Flavobacterium johnsoniae FjCh103
(HQ622609)98 Bacteroidetes
B4 Flexibacter sp. HDL53 (HQ891951) 100 BacteroidetesB5 Flavobacterium sp. GSE09 (EU034657) 100 BacteroidetesB6 Bacillus sp. S542Ba-114 (HQ238443) 99 FirmicutesB7 Bacterium Ellin5227 (AY234578) 96 AcidobacteriaB8 Methylobacterium sp. EP5 (AY436801) 97 a-Proteobacteria
X. Li et al. / Soil Biology & Biochemistry 47 (2012) 191e197 195
compared to the control treatment (Fig. 1 and Table S1). Because ofthe lack of a sterile soil control with the MCS treatment, it wasimpossible to assess the effects of abiotic factors on the degradationof PAHs. For example, the organic matter of MCS may adsorb PAHs,thus reducing the bioavailability of the PAHs in soil (Beckles et al.,2007). Therefore, it seems that the depletion of PAHs with treat-ment S could be largely attributed to the enhanced degradationactivity of indigenous microorganisms.
The addition of MCS to soil significantly changed the microbialcommunities. Although MCS is commonly used for edible mush-room cultivation, it also stimulated the growth of bacteria andAHDs simultaneously (Fig. 2), possibly because of the enrichment ofsoil organic matter and nitrogen contents (Table 2). The communitycompositions of bacteria and fungi were considerably affected bythe addition of MCS (Figs. 3 and 4), so that the relative abundance of
Table 4The closest sequence match for known phylogenetic affiliations to fungal 18S rRNA band
Band Closest culture in Genbank
F1 Mortierella sp. CO-21 (AB521052)F2 Spizellomycete sp. JEL371 (DQ536490)F3 Moesziomyces bullatus AFTOL-ID 1820 (DQ831012)F4 Melanopsichium pennsylvanicum (DQ363314)F5 Cochliobolus sativus AFTOL-ID 271 (DQ677995)F6 Coniochaeta ostrea AFTOL-ID 915 (DQ471007)F7 Mariannaea elegans CBS 239.56 (AY526489)F8 Pleurostomophora repens CBS H-7594 (AY761067)
a The phylogeny of fungi follows the system of Dictionary of the Fungi (Kirk et al., 200
two Sordariomycete species increased (Fig. 4).We do not knowwhythese species were selected for, and the unambiguous identificationof active PAH-degrading species in soil is impossible in this study.New technologies, such as stable isotope probing, might be prom-ising in recognizing the active player in soil (Jones et al., 2011).
Notably, the dissipation of individual PAHs was not dependenton the molecular weight, the major property influencing thedegradability of PAHs (Juhasz and Naidu, 2000). With treatment S,the concentrations of anthracene, benzo(a)pyrene and benzo(a)anthracene were reduced by >50% (Fig. 1). This result providesinsight regarding the underlying mechanism of PAH degradationbecause these three PAHs have been found to be susceptible tooxidation by fungal laccase (Collins et al., 1996; Majcherczyk et al.,1998). In addition, similar degradation can be also observed withcrude extracts of spent mushroom cultivation compost (Li et al.,2010b) or ligninolytic fungi (Steffen et al., 2007), and fungal lac-case can remove most anthracene and benzo(a)pyrene in soilquickly as well (Wu et al., 2008). In contrast, bacteria are rarelyreported to degrade benzo(a)pyrene in soil (Juhasz and Naidu,2000). Considering that some Sordariomycete species enriched intreatment S have ligninolytic enzyme activity (Lopez et al., 2007), itseems plausible that fungal laccase might play a role in thebioconversion of PAHs.
Other possible explanations for the degradation of PAHs withtreatment S exist. For example, fungi normally synthesize complexenzymes that degrade PAHs to some extent (Aranda et al., 2010;Sack et al., 1997). Bacteria with laccase activity may also contributeto the degradation of benzo(a)pyrene and anthracene (Zeng et al.,2011). It should be noted that the interactions between fungi andbacteria may affect the degradation of PAHs in soil (Borras et al.,
sequences extracted from DGGE gel.
Similarity (%) Phylum (Class)a
100 Zygomycota (Mucoromycotina)100 Chytridiomycota (Chytridiomycetes)100 Basidiomycota (Ustilaginaceae)100100 Ascomycota (Dothideomycetes)100 Ascomycota (Sordariomycetes)99
100
8).

X. Li et al. / Soil Biology & Biochemistry 47 (2012) 191e197196
2010). As such, the mechanisms of PAH degradation by MCS bio-stimulation merit further study.
Two additional bioremediation treatments appeared to be lessefficient than biostimulation with MCS. Compared with treatmentS, the addition of P. ostreatus (treatment SP) to soil improved thedegradation of several PAHs at nonsignificant levels while the totalPAH concentrations with treatments S and SP were roughly equal(Table S1). The colonization of allochthonous microbial species insoil is often complicated by many factors and is a major concern inthe application of bioaugmentation (Baldrian, 2008). P. ostreatushas been implicated in PAH depletion in some studies (Byss et al.,2008; Novotný et al., 1999); however, in this study, P. ostreatusmight not be a strong competitor in soil, demonstrated by the factthat no dominant band in DGGE profiles could be identified as thiswhite rot fungus (Fig. 4 and Table 4).
The effect of MCS on the dissipation of PAHs was hindered byalfalfa (Treatment SPA, Table S1 and Fig. 1), although the presenceof alfalfa has been reported to considerably decrease PAHs in soil(Teng et al., 2011). Most interestingly, there was no significantdifference inmicrobial population sizes (Fig. 2), and nearly identicalfungal community compositions were observed between treat-ments SP and SPA (Fig. 4). Therefore, the competition for bulknutrients between plants and microbes may be largely negligible,as further evidenced by the similar levels of carbon and nitrogen inboth treatments (Table 2). Interestingly, the addition of alfalfa tosamples increased soil pH from 4.58 with treatment SP to 4.98 withtreatment SPA (Table 2), as demonstrated in a previous study(Donegan et al., 1999). The optimal pH range for laccase activity isbetween 3.5 and 4.5 (Li et al., 2010a), which might be an explana-tion for the decreased dissipation of benzo(a)pyrene, anthraceneand benzo(a)anthracene in soil microcosms of treatment SPA.Moreover, the effects of rhizodeposition (the release of organiccompounds from plant roots into soil) on PAH degraders appear tobe complicated (Meng and Zhu, 2010), and the plant metabolitesmay hinder the biodegradation of PAH (Qiu et al., 2004). Morestudies are needed to understand the interaction between the rootsof alfalfa and soil microbial communities.
5. Conclusions
The current study tested the potential of a mixed substrate usedfor mushroom cultivation (MCS) in the bioremediation of aged,PAH-contaminated soil. Three PAHs, anthracene, benzo(a)pyreneand benzo(a)anthracene, were most susceptible to degradation,which is consistent with the PAH degradation characteristics offungal laccase. Other strategies were less effective than or inhibi-tory on biostimulation, suggesting complexity of the regulation ofPAH degrader activity. The results of this study demonstrate thatMCS is a promising biostimulation agent for the bioremediation ofPAH-contaminated soils.
Acknowledgments
We thank Junhua Wang for the technical assistance in soilanalysis. This work was financially supported by grants fromMinistry of Science and Technology of China (to X. Lin), Interna-tional Foundation for Science (to Y. Wu), Natural Science Founda-tion of China (to J. Zhang) and Natural Science Foundation ofZhejiang Province (to X. Li).
Appendix. Supplementary material
Supplementary material associated with this article can befound, in the online version, at doi:10.1016/j.soilbio.2012.01.001.
References
Aranda, E., Ullrich, R., Hofrichter, M., 2010. Conversion of polycyclic aromatichydrocarbons, methyl naphthalenes and dibenzofuran by two fungal perox-ygenases. Biodegradation 21, 267e281.
Baldrian, P., der Wiesche, C.I., Gabriel, J., Nerud, F., Zadrazil, F., 2000. Influence ofcadmium and mercury on activities of ligninolytic enzymes and degradation ofpolycyclic aromatic hydrocarbons by Pleurotus ostreatus in soil. Applied andEnvironmental Microbiology 66, 2471e2478.
Baldrian, P., 2008. Wood-inhabiting ligninolytic basidiomycetes in soils: ecologyand constraints for applicability in bioremediation. Fungal Ecology 1, 4e12.
Beckles, D.M., Chen, W., Hughes, J.B., 2007. Bioavailability of polycyclic aromatichydrocarbons sequestered in sediment: microbial study and model prediction.Environmental Toxicology and Chemistry 26, 878e883.
Borras, E., Caminal, G., Sarra,M., Novotny, C., 2010. Effect of soil bacteria on the abilityof polycyclic aromatic hydrocarbons (PAHs) removal by Trametes versicolor andIrpex lacteus from contaminated soil. Soil Biology & Biochemistry 42, 2087e2093.
Byss, M., Elhottova, D., Triska, J., Baldrian, P., 2008. Fungal bioremediation of thecreosote-contaminated soil: influence of Pleurotus ostreatus and Irpex lacteus onpolycyclic aromatic hydrocarbons removal and soil microbial communitycomposition in the laboratory-scale study. Chemosphere 73, 1518e1523.
Cerniglia, C.E., 1997. Fungal metabolism of polycyclic aromatic hydrocarbons: past,present and future applications in bioremediation. Journal of IndustrialMicrobiology & Biotechnology 19, 324e333.
Collins, P., Kotterman, M., Field, J., Dobson, A., 1996. Oxidation of anthracene andbenzo[a]pyrene by laccases from Trametes versicolor. Applied and Environ-mental Microbiology 62, 4563e4567.
Donegan, K.K., Seidler, R.J., Doyle, J.D., Porteous, L.A., Digiovanni, G., Widmer, F.,Watrud, L.S., 1999. A field study with genetically engineered alfalfa inoculatedwith recombinant Sinorhizobium meliloti: effects on the soil ecosystem. Journalof Applied Ecology 36, 920e936.
Gan, S., Lau, E.V., Ng, H.K., 2009. Remediation of soils contaminated with polycyclicaromatic hydrocarbons (PAHs). Journal of Hazardous Materials 172, 532e549.
Harms, H., Schlosser, D., Wick, L.Y., 2011. Untapped potential: exploiting fungi inbioremediation of hazardous chemicals. Nature ReviewsMicrobiology 9,177e192.
Jones, M.D., Crandell, D.W., Singleton, D.R., Aitken, M.D., 2011. Stable-isotopeprobing of the polycyclic aromatic hydrocarbon-degrading bacterial guild ina contaminated soil. Environmental Microbiology 13, 2623e2632.
Joseph, S., 1997. Review of the estrogenic and antiestrogenic activity of polycyclicaromatic hydrocarbons: relationship to carcinogenicity. Chemosphere34, 835e848.
Juhasz, A.L., Naidu, R., 2000. Bioremediation of high molecular weight polycyclicaromatic hydrocarbons: a review of the microbial degradation of benzo[a]pyrene. International Biodeterioration & Biodegradation 45, 57e88.
Kirk, P.M., Cannon, P.F., Minter, D.W., Stalpers, J.A., 2008. Dictionary of the Fungi,10th ed. CAB International, Wallingford, UK.
Lau, K.L., Tsang, Y.Y., Chiu, S.W., 2003. Use of spent mushroom compost tobioremediate PAH-contaminated samples. Chemosphere 52, 1539e1546.
Li, X., Lin, X., Yin, R., Wu, Y., Chu, H., Zeng, J., Yang, T., 2010a. Optimization of laccase-mediated benzo[a]pyrene oxidation and the bioremedial application in agedpolycyclic aromatic hydrocarbons-contaminated soil. Journal of Health Science56, 534e540.
Li, X., Lin, X., Zhang, J., Wu, Y., Yin, R., Feng, Y., Wang, Y., 2010b. Degradation ofpolycyclic aromatic hydrocarbons by crude extracts from spent mushroomsubstrate and its possible mechanisms. Current Microbiology 60, 336e342.
Lopez, M.J., Vargas-Garcia, M.D., Suarez-Estrella, F., Nichols, N.N., Dien, B.S.,Moreno, J., 2007. Lignocellulose-degrading enzymes produced by the ascomy-cete Coniochaeta ligniaria and related species: application for a lignocellulosicsubstrate treatment. Enzyme and Microbial Technology 40, 794e800.
Lu, R., 2000. Analytical Methods for Soil and Agrochemistry. China AgriculturalScience and Technology Press, Beijing.
Majcherczyk, A., Johannes, C., Hüttermann, A., 1998. Oxidation of polycyclicaromatic hydrocarbons (PAH) by laccase of Trametes Versicolor. Enzyme andMicrobial Technology 22, 335e341.
Meng, L., Zhu, Y.-G., 2010. Pyrene biodegradation in an industrial soil exposed tosimulated rhizodeposition: how does it affect functional microbial abundance?Environmental Science & Technology 45, 1579e1585.
Muyzer, G., de Waal, E.C., Uitterlinden, A.G., 1993. Profiling of complex microbialpopulations by denaturing gradient gel electrophoresis analysis of polymerasechain reaction-amplified genes coding for 16S rRNA. Applied and Environ-mental Microbiology 59, 695e700.
Novotný, �C., Erbanová, P., �Sa�sek, V., Kubátová, A., Cajthaml, T., Lang, E., Krahl, J.,Zadra�zil, F., 1999. Extracellular oxidative enzyme production and PAH removalin soil by exploratory mycelium of white rot fungi. Biodegradation 10, 159e168.
Olson, P.E., Castro, A., Joern, M., DuTeau, N.M., Pilon-Smits, E.A.H., Reardon, K.F.,2007. Comparison of plant families in a greenhouse phytoremediation study onan aged polycyclic aromatic hydrocarbon-contaminated soil. Journal ofEnvironmental Quality 36, 1461e1469.
Phillips, L.A., Greer, C.W., Germida, J.J., 2006. Culture-based and culture-independent assessment of the impact of mixed and single plant treatmentson rhizosphere microbial communities in hydrocarbon contaminated flare-pitsoil. Soil Biology & Biochemistry 38, 2823e2833.
Ping, L.F., Luo, Y.M., Zhang, H.B., Li, Q.B., Wu, L.H., 2007. Distribution of polycyclicaromatic hydrocarbons in thirty typical soil profiles in the Yangtze River Deltaregion, east China. Environmental Pollution 147, 358e365.

X. Li et al. / Soil Biology & Biochemistry 47 (2012) 191e197 197
Qiu, X.J., Reed, B.E., Viadero, R.C., 2004. Effects of flavonoids on C-14 7,10-benzoa pyrene degradation in root zone soil. Environmental Engineering Science 21,637e646.
Ribas, L.C.C., de Mendonca, M.M., Camelini, C.M., Soares, C.H.L., 2009. Useof spent mushroom substrates from Agaricus subrufescens (syn. A. blazei,A. brasiliensis) and Lentinula edodes productions in the enrichment ofa soil-based potting media for lettuce (Lactuca sativa) cultivation: growthpromotion and soil bioremediation. Bioresource Technology 100,4750e4757.
Sack, U., Hofrichter, M., Fritsche, W., 1997. Degradation of polycyclic aromatichydrocarbons by manganese peroxidase of Nematoloma frowardii. FemsMicrobiology Letters 152, 227e234.
Steffen, K.T., Schubert, S., Tuomela,M., Hatakka, A., Hofrichter,M., 2007. Enhancementof bioconversion of high-molecular mass polycyclic aromatic hydrocarbons incontaminated non-sterile soil by litter-decomposing fungi. Biodegradation 18,359e369.
Teng, Y., Shen, Y.Y., Luo, Y.M., Sun, X.H., Sun, M.M., Fu, D.Q., Li, Z.G., Christie, P., 2011.Influence of Rhizobium meliloti on phytoremediation of polycyclic aromatichydrocarbons by alfalfa in an aged contaminated soil. Journal of HazardousMaterials 186, 1271e1276.
Vainio, E.J., Hantula, J., 2000. Direct analysis of wood-inhabiting fungi usingdenaturing gradient gel electrophoresis of amplified ribosomal DNA. Myco-logical Research 104, 927e936.
Wrenn, B.A., Venosa, A.D., 1996. Selective enumeration of aromatic and aliphatichydrocarbon degrading bacteria by a most-probable-number procedure.Canadian Journal of Microbiology 42, 252e258.
Wu, Y., Teng, Y., Li, Z., Liao, X., Luo, Y., 2008. Potential role of polycyclic aromatichydrocarbons (PAHs) oxidation by fungal laccase in the remediation of an agedcontaminated soil. Soil Biology & Biochemistry 40, 789e796.
Zeng, J., Lin, X.G., Zhang, J., Li, X.Z., Wong, M.H., 2011. Oxidation of polycyclicaromatic hydrocarbons by the bacterial laccase CueO from E. coli. AppliedMicrobiology and Biotechnology 89, 1841e1849.