differentiation of mouse embryonic stem cells into insulin-secreting cells in vitro
TRANSCRIPT

JOURNAL OF MEDICAL COLLEGES OF PLA
www.elsevier.com/locate/jmcpla
Journal of Medical Colleges of PLA 26 (2011) 1–12
Differentiation of mouse embryonic stem cells into insulin-secreting cells in vitro
Sui Jing1, 2, Jiang Fangxu2, Shi Bingyin1* 1Department of Endocrinology, First Affiliated Hospital, Medical College, Xi’an Jiaotong University, Xi’an 710061,
China 2Center for Diabetes Research, Western Australian Institute for Medical Research, Centre for Medical Research,
University of Western Australia, Perth 6000, Australia
Received 26 December 2010; accepted 21 January 2011
Abstract
Regenerative medicine, including cell-replacement strategies, may have an important role in the treatment of type 1 diabetes which is associated with decreased islet cell mass. To date, significant progress has been made in generating insulin-secreting β cells from pluripotent mouse embryonic stem cells (ESCs).The aim of this study is to explore the potential of regulating the differentiation of ESCs into pancreatic endocrine cells capable of synthesizing the pancreatic hormones including insulin, glucagon, somatostatin and pancreatic polypeptide under proper conditions. Undifferentiated ES cell line was stably transfected with mouse RIP-YFP plasmid construction in serum-free medium using Lipofectamine™ 2000 Reagents. We tested pancreatic specific gene expression and characterized these ESC-derived pancreatic endocrine cells. Most of these insulin-secreting cells co-expressed many of the phenotypic markers characteristic of β cells such as insulin1, insulin2, Islet1, MafA, insulinoma-associated antigen 1 (IA1) and so on, indicating a similar gene expression pattern to adult islet β cells in vivo. Characterization of this population revealed that it consisted predominantly of pancreatic endocrine cells that were able to undergo pancreatic specification under the appropriate conditions. We also demonstrated that zinc supplementation mediated up-regulation of insulin-secreting cells as an effective inducer promoted the development of ESC-derived diabetes therapy. In conclusion, this work not only established an efficient pancreatic differentiation strategy from ESCs to pancreatic endocrine lineage in vitro, but also leaded to the development of new strategies to derive transplantable islet-replacement β cells from embryonic stem cells for the future applications of a stem cell based therapy of diabetes.
Keywords: Embryonic stem cells; Pancreatic differentiation; Insulin-secreting cells; Transcription factors; Zinc
1. Introduction
Diabetes mellitus affects 4%–5% of the world’s population and is the most common
* Corresponding author.
E-mail address: [email protected] (Shi B.)

2 Sui Jing et al. / Journal of Medical Colleges of PLA 26 (2011) 1–12
metabolic disorder in humans. The number of people with diabetes is predicted to exceed 350 million by 2010 [1]. Type 1 diabetes mellitus is characterized by autoimmune destruction of β cells by the cellular and humoral immune system in the pancreatic islets of Langerhans which leads to impaired insulin secretion and subsequently to hyperglycemia. Daily injection of insulin is the current treatment for the disease. Transplantation of islets represents a potential cure; however, the limited supply of donor pancreatic tissue and the necessity of chronic immunosuppression make this cell therapy approach inadequate to meet the global diabetes population demand for treatment. A potential source for endocrine cells is embryonic stem cells (ESCs). Embryonic stem cells are pluripotent cells derived from the inner cell mass (ICM) of the mammalian blastocyst and can differentiate into all cell types including pancreatic insulin-secreting cells, offers a perspective of success of cell-replacement therapy for diabetes.
It has been reported by many groups [2–7] that ESCs can be induced to differentiate into insulin-secreting cells in vitro, however, these methods have various limitations and suggest that there are still many challenges to using ESC-derived populations for clinical therapy. These challenges may be met by using new reagents or new combination of different reagents to induce the differentiation of ESCs. Therefore, much better understanding of the mechanisms that regulate expansion and differentiation of embryonic stem cells is necessary. Zinc plays an important role in all processes of insulin trafficking, i.e., synthesis, storage [8] and secretion [9]. Zinc can be transported by zinc transporters from cytoplasm into insulin secretary vesicles in which insulin is stored as a solid hexamer bound with two Zn2+ before secretion. The behavior of proinsulin and insulin in the presence of zinc suggests it plays an important role in insulin’s production in the β cell for the vast majority of animal species. Proinsulin hexamer is converted readily into the
insulin hexamer and forms a zinc containing hexamer soon after its synthesis and this organization of the molecule is maintained through all the subsequent processes [10]. In addition, the zinc ions enhance proinsulin’s solubility and render insulin insoluble. Zinc ions also appear to play an important role in the microcrystalline character of the precipitated insulin granule [10]. However, the role of zinc in the ESC-derived pancreas development is still unclear. The present study was thus undertaken to explore the potential of regulating the differentiation of pluripotent ESCs into pancreatic endocrine cells capable of synthesizing the pancreatic hormones including insulin, glucagon, somatostatin and pancreatic polypeptide under proper conditions and determine the effects of zinc supplementation on the induction of ESC-derived endocrine cells.
2. Materials and methods 2.1. In vitro cell culture and differentiation procedure
In order to maintain undifferentiated mouse ES cell line ES W9.5 (a kind gift from the Walter and Eliza Hall Institute of Medical Research, Australia) in an embryonic state, they were cultured on gelatin-coated γ-irradiated primary mouse embryonic fibroblasts (MEFs) feeder layers in Dulbecco’s modified Eagle medium (DMEM, glucose 4.5 g/L) supplemented with 15% Knockout serum replacement, 1% nonessential amino acid, 1% nucleotide, 0.05 mmol/L β-mercaptoethanol, 1% vol/vol penicillin/streptomycin and 1000 U/ml leukemia inhibitory factor (LIF, CHEMICON). Cells were passaged with 0.05% trypsin-EDTA every 3 or 4 d onto freshly seeded (3×104 cells/cm2) γ-irradiated MEFs and cultured in a humidified chamber containing a 10% CO2/air mixture at 37 ℃. Typical splitting ratios were between 1:2 and 1:6, depending on the density of the starting culture. ESCs at passages 16–33 were used in this study. All

Sui Jing et al. / Journal of Medical Colleges of PLA 26 (2011) 1–12 3
other reagents were purchased from Invitrogen (Gibco) unless stated otherwise.
For differentiation with our differentiation strategy, ES W9.5 (subclone 1) cells were seeded at a density of 2×104 cells/cm2 at passages 16–33, and cultured for 3–4 d until a confluent flat layer of undifferentiated cells was formed. These cells were differentiated towards insulin-secreting cells using a 5-stage differentiation protocol. At the start of each differentiation procedure, the trypsinized ESCs were plated onto 6-well plate with 200 ng/ml MatrigelTM (BD Biosciences) without LIF and feeder layers in DMEM (glucose 1 g/L) medium. First, 50 ng/ml activin A (R&D) was used to induce definitive endoderm (DE) formation. Second, on day 5 the differentiated endoderm cells were treated with 10 μmol/L retinoic acid (RA) [11] and 10 ng/ml fibroblast growth factor 10 (FGF10) [12] (R&D) in DMEM (glucose 4.5 g/L) medium supplemented with 1% B27 supplement to induce posterior foregut (PF) formation. Third, on day 9 10 μmol/L RA, 10 μmol/L cyclopamine [13] (CYC), 5 μmol/L Notch (γ-secretase) inhibitor DAPT [14] (Sigma), 50 ng/ml β cell trophic glucagon-like peptide-1 receptor agonist exendin 4 (Ex4) [15] and various concentrations of zinc chloride (ZnCl2) (0, 0.1, 0.5 and 1 mmol/L) were added on the differentiation of foregut cells to induce pancreatic endoderm (PE) specialization, which was assessed by RT-PCR. Fourth, on day 16 the differentiated pancreatic cells were treated with 10 μmol/L RA, 5 μmol/L DAPT and 50 ng/ml exendin 4 to induce islet progenitor cells (IP). Fifth, 5 mmol/L Nicotinamide (Nic) [l6] and exendin 4 were used on the differentiation of insulin-secreting cells. The various factors combination during the differentiation experiments is displayed in Fig. 1.
2.2. Plasmid constructs and transfection of ES cells with RIP-YFP plasmid
Undifferentiated ES W9.5 cells were stably transfected with mouse rat insulin promoter-yellow
fluorescent protein (RIP-YFP) plasmid construction (a kind gift from the Walter and Eliza Hall Institute of Medical Research, Australia) in serum-free medium using Lipofectamine™ 2000 Reagents according to the manufacturer’s instructions. The RIP gene is placed above the neomycin resistance gene, thus permitting neomycin resistance when RIP is activated. The rat insulin promoter was used to control the expression of enhanced yellow fluorescence protein in mouse embryonic stem cells during their differentiation towards insulin- secreting cells. After 2 d, transfected cells were cultured in our standard medium and selected by growing with 500 µg/ml G418 neomycin to select RIP-YFP positive cells. These cells were cultured
Fig. 1. Sketch of ESCs differentiation protocol. First, undifferentiated ESCs were first cultured in DMEM (glucose 1 g/L) containing 50 ng/ml activin A for 4 d. Second, the differentiated DE cells were further induced with 10 μmol/L RA and 10 ng/ml FGF10 in DMEM (glucose 4.5 g/L) with 1% B27 supplement for another 4 d. Third, the differentiated PF cells were exposed to RA, 10 μmol/L CYC, 5 μmol/L DAPT and 50 ng/ml Ex4 in DMEM (glucose 4.5 g/L) with 1% B27 supplement to induce pancreatic endoderm specialization on day 12–15. Fourth, the pancreatic endoderm cells were treated with RA, DAPT and Ex4 in DMEM (glucose 4.5 g/L) with 1% B27 supplement to induce islet progenitor cells on day 20–25. Fifth, 5 mmol/L Nic and 50 ng/ml Ex4 were used on the differentiation of insulin-secreting cells on day 25–30.
Insulin-secretingmESCs DE PF PE IP cells
Day 0 4 8 12–15 20–25 25–30
Activin A
RA
+FGF10
RA
+CY
C+D
APT
+Ex4
RA
+DA
PT+Ex4
Nic+Ex4
4 Sui Jing et al. / Journal of Medical Colleges of PLA 26 (2011) 1–12
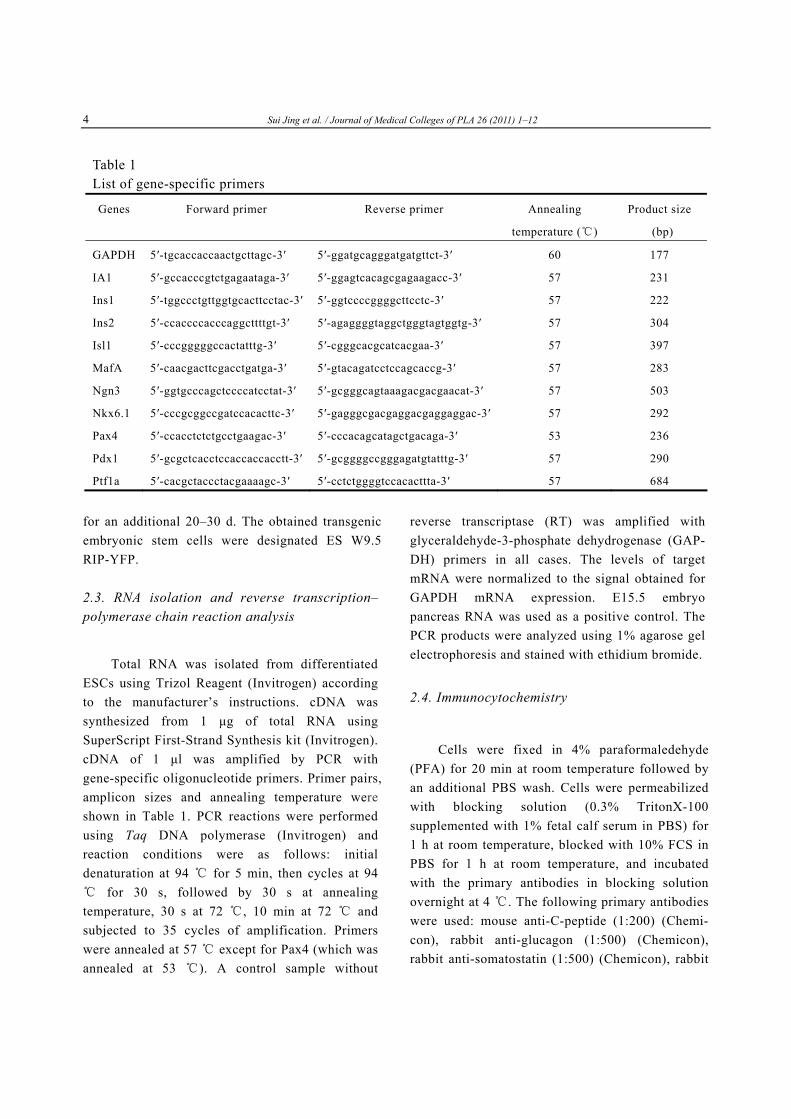
Table 1 List of gene-specific primers
Genes Forward primer Reverse primer Annealing Product size
temperature (℃) (bp)
GAPDH 5′-tgcaccaccaactgcttagc-3′ 5′-ggatgcagggatgatgttct-3′ 60 177
IA1 5′-gccacccgtctgagaataga-3′ 5′-ggagtcacagcgagaagacc-3′ 57 231
Ins1 5′-tggccctgttggtgcacttcctac-3′ 5′-ggtccccggggcttcctc-3′ 57 222
Ins2 5′-ccaccccacccaggcttttgt-3′ 5′-agaggggtaggctgggtagtggtg-3′ 57 304
Isl1 5′-cccgggggccactatttg-3′ 5′-cgggcacgcatcacgaa-3′ 57
for an additional 20–30 d. The obtained transgenic embryonic stem cells were designated ES W9.5 RIP-YFP.
2.3. RNA isolation and reverse transcription– polymerase chain reaction analysis
Total RNA was isolated from differentiated ESCs using Trizol Reagent (Invitrogen) according to the manufacturer’s instructions. cDNA was synthesized from 1 μg of total RNA using SuperScript First-Strand Synthesis kit (Invitrogen). cDNA of 1 μl was amplified by PCR with gene-specific oligonucleotide primers. Primer pairs, amplicon sizes and annealing temperature were shown in Table 1. PCR reactions were performed using Taq DNA polymerase (Invitrogen) and reaction conditions were as follows: initial denaturation at 94 ℃ for 5 min, then cycles at 94
for ℃ 30 s, followed by 30 s at annealing temperature, 30 s at 72 , 10 min at 72℃ and ℃
subjected to 35 cycles of amplification. Primers were annealed at 57 except for ℃ Pax4 (which was annealed at 53 ). A control sample without℃
reverse transcriptase (RT) was amplified with glyceraldehyde-3-phosphate dehydrogenase (GAP- DH) primers in all cases. The levels of target mRNA were normalized to the signal obtained for GAPDH mRNA expression. E15.5 embryo pancreas RNA was used as a positive control. The PCR products were analyzed using 1% agarose gel electrophoresis and stained with ethidium bromide.
2.4. Immunocytochemistry
Cells were fixed in 4% paraformaledehyde (PFA) for 20 min at room temperature followed by an additional PBS wash. Cells were permeabilized with blocking solution (0.3% TritonX-100 supplemented with 1% fetal calf serum in PBS) for 1 h at room temperature, blocked with 10% FCS in PBS for 1 h at room temperature, and incubated with the primary antibodies in blocking solution overnight at 4 . The following primary℃ antibodies were used: mouse anti-C-peptide (1:200) (Chemi- con), rabbit anti-glucagon (1:500) (Chemicon), rabbit anti-somatostatin (1:500) (Chemicon), rabbit
397
MafA 5′-caacgacttcgacctgatga-3′ 5′-gtacagatcctccagcaccg-3′ 57 283
Ngn3 5′-ggtgcccagctccccatcctat-3′ 5′-gcgggcagtaaagacgacgaacat-3′ 57 503
Nkx6.1 5′-cccgcggccgatccacacttc-3′ 5′-gagggcgacgaggacgaggaggac-3′ 57 292
Pax4 5′-ccacctctctgcctgaagac-3′ 5′-cccacagcatagctgacaga-3′ 53 236
Pdx1 5′-gcgctcacctccaccaccacctt-3′ 5′-gcggggccgggagatgtatttg-3′ 57 290
Ptf1a 5′-cacgctaccctacgaaaagc-3′ 5′-cctctggggtccacacttta-3′ 57 684

Sui Jing et al. / Journal of Medical Colleges of PLA 26 (2011) 1–12 5
anti-polypeptide (1:500) (Chemicon). The following day, cells were washed three times with PBS and then incubated for 1 h at room temperature in the dark with the secondary antibody in PBS, washed for 3 times again, and then counterstained with 4’,6-diamidino-2-phenylindole (DAPI) (1:1 000) (Sigma) for 5 min. The following secondary antibodies were used: goat anti-mouse TEXAS RED (1:200) (Vector), goat anti-rabbit FITC (1:200). Images of immunofluorescently stained cells were taken on an Olympus IX71 U-RFL-T fluorescence microscope and DPController software was used. Images were edited in Adobe Photoshop version 7.0. For quantification of positive cells, cells in 5 different randomly chosen images from 2 separate experiments were calculated. Based on these calculations, mean±SEM was determined.
3. Results 3.1. Directed pancreatic differentiation of mouse embryonic stem cells
We have developed a 5-step protocol which was optimized primarily in the mouse ES W9.5 cell line for differentiation of ESCs to pancreatic islet-like insulin-secreting cells (Fig. 1). We focused first on generating definitive endoderm (DE), followed by posterior foregut (PF), pancreas endoderm (PE), islet progenitors(IP) and, finally, insulin-secreting cells. Most of the living cells were epithelial-like and many small cell clusters appeared and formed small cluster-like constructions (Fig. 2).
To test whether the mouse ESCs differentiated into pancreatic endocrine cells, we detected the expression of several marker genes in induced epithelial-like cells at short intervals throughout the course of differentiation according to our previous study by RT-PCR. When the differentiated PF cells were exposed to RA, cyclopamine, DAPT and exendin 4, the cells rapidly began to express high levels of Pdx1, one of the earliest genes expressed
in the pancreatic lineage, and associated with progressive up-regulation of the early pancreatic epithelial markers, including basic helix-loop-helix (bHLH) transcription factor Ptf1a [17], paired domain transcription factors Pax4 and homeodomain proteins Nkx6.1 (Fig. 3). The co- expression of these genes appeared to closely imitate the timed expression of these factors during this decisive phase of pancreatic development. In order to get some relevant estimation of the magnitude of Pdx1-expression in differentiated ESCs, related gene expression pattern was compared to mouse E15.5 embryo pancreas tissue.
3.2. Pancreatic differentiation of ES W9.5 cells results in RIP activation and identification
To facilitate the identification of insulin- secreting cells and to track these cells, we have developed a marker gene construct: fusing the rat insulin promoter gene to the yellow fluorescent protein gene. This gene-trapping construct was transfected into undifferentiated ESCs to trace differentiation into insulin-secreting cells. YFP expression recapitulated endogenous RIP ex- pression when RIP was induced by a list of factors according to our differentiation procedure. The subsequent YFP expression was identified in RIP positive cells, YFP expressing cells appeared biased to an pancreatic endocrine fate. This cell line enables one to identify distinct developmental stage of pancreatic endocrine lineage based on YFP expression induced by RIP.
In our study, YFP expression was identified by fluorescent microscopy during the differentiation procedure (Fig. 4).This gene appeared initially in the small cluster epithelial cells of the pancreatic buds on embryonic day 20 (E20) , until around E26, when its expression became stronger and stronger and restricted to pancreatic islet insulin-secreting cells. We observed that YFP positive cells could survive until E35. The result strongly indicated that our differentiat ion s trategy is eff icient and

6 Sui Jing et al. / Journal of Medical Colleges of PLA 26 (2011) 1–12
Fig. 4. Mouse ESCs differentiation towards pancreatic endocrine insulin-secreting cells by expression of RIP-YFP in mouse embryonic stem cells derived cellpopulation. ESCs were grown in differentiation mediaand differentiated to RIP-YFP expressing cells according to our differentiation strategy. YFP positive cells appeared initially in the small cluster epithelialcells of the pancreatic buds on embryonic day 20(E20), until around E26, when its expression becomesstronger and stronger and restrict to
pancreatic endocrine fate was identified in vitro. Most of the YFP positive cells appeared scattered in the cell clusters, demonstrated that ESC-derived insulin-secreting cells differentiated better in the cell clusters than in single cells. It seems that differentiation events within cell clusters appear to mimic early embryogenesis. Intracellular interactions, surrounded with the nutrients and growth factors, may be fundamental to this differentiation process. The above analyses highlight the power of transgene reporters in defining distinct developmental stages during ESCs cell differentiation. When analyzing pancreatic endocrine insulin-secreting cells induction and differentiation from ESCs, this reporter line is feasible and powerful.
Fig. 2. Morphological analysis of ESC colonies on MEFs and the differentiated ESC-derived insulin- secreting like cells at the final differentiation stage. A: Phase-contrast images of an undifferentiated ESC colonies at day 3 after passaging. B: ESC-derived differentiated insulin-secreting like cells in the center of the colonies on day 26 (Scale bars: 50 µm).
A
B
pancreatic isletinsulin-secreting cells. YFP positive cells couldsurvive until E35. A: E20; B: E22; C: E26; D: E35. Scale bars: 50 µm.
A B
C D
1 2 3 4
Fig. 3. RT-PCR analysis of gene expression in
Pdx1-expressing pancreas endoderm by adding various
concentrations of ZnCl2. Total differentiated cells
derived from designated populations were procured and
gene expression analyzed. 1–4: 4 separate culture
experiments by adding 0.1, 0.5, 1 and 0 mmol/L o
Pdx1
Ptf1a
Pax4
Nkx6.1
Ngn3
GAPDH
f
ZnCl2. The GAPDH gene was used as a housekeeping-
ene standard. g

Sui Jing et al. / Journal of Medical Colleges of PLA 26 (2011) 1–12 7
3.3. Zinc promotes differentiation of mouse embryonic stem cells into insulin-secreting cells in vitro
Zinc plays an important role in all processes of insulin trafficking, i.e., synthesis, storage [8], and secretion [9]. Study of the developmental biology literature leads us to test the roles of various concentrations of ZnCl2 on the differentiation of mouse embryonic stem cells into insulin-secreting cells in vitro. Activation of Pdx1 is considered a prerequisite for pancreatic differentiation in vitro and should precede the progressive expression of more mature markers of the endocrine lineage, including Ptf1a, Nkx6.1, Pax4, and Ngn3. After foregut endoderm cultures are exposed to RA, CYC, DAPT and exendin 4, there is a rapid induction of many important transcription factors which have been shown to be essential for correct specification of the pancreatic anlagen and to be important in pancreatic endoderm development, including Pdx1, Ptf1a, Nkx6.1 and Pax4. The coordinate expression of these genes appears to closely imitate the timed expression of these factors during this decisive phase of pancreatic development.
We checked the role of zinc chloride on the differentiation of pancreatic endocrine lineage by adding 4 reasonable concentrations( 0, 100 and 500 μmol/L, and 1 mmol/L) to the differentiation medium every 2–3 d. Compared to the negative control which was absence of zinc chloride (0 μmol/L), the cells adding 100 μmol/L ZnCl2 and 500 μmol/L ZnCl2 showed higher gene expression of pancreatic markers such as Pdx1, Ptf1a, Nkx6.1, Pax4, and Ngn3 (Fig. 3), while cells adding 1 mmol/L ZnCl2 showed poorer gene expression. Furthermore, among the 4 different concentrations, 500 μmol/L ZnCl2 gave the highest gene expression and showed the potential during the ESC-derived pancreas differentiation. This indicates that ZnCl2
maybe not directly cause or affect the differentiation of pancreatic endoderm cells, but
shows great benefit. Therefore, zinc supplemen- tation can promote the differentiation of ESCs to pancreatic endocrine lineage and promote the pancreatic gene expression by adding the proper concentrations at the proper time. 3.4. In vitro characterization of ESC-derived RIP-expressing insulin-secreting cells
The ideal characteristics of embryonic stem cell-based differentiation to pancreatic β cells depends on ideal characteristics of islet insulin- secreting cells for their ability to express pancreatic islet genes and proteins. Using immunofluorescence and RT-PCR, we systematically characterized ESC-derived RIP-expressing insulin-secreting cells. The transcription factors expressing profile closely matches that of committed pancreatic endocrine development in the embryos according to our differentiation protocol. To characterize the generated insulin-secreting cells at this stage, we first examined gene expression of islet β cell specific marker genes such as Ngn3, Insulin1, Insulin2, Islet1, IA1 and MafA which were clearly detected by RT-PCR (Fig. 5) and consistent with the hormone expression pattern of mature islets and referred to as pancreatic endocrine cells. In order to get some relevant estimation of the magnitude of the insulin-secreting cells in differentiated ESCs, related gene expression pattern was compared to mouse E15.5 embryo pancreas. The crucial transcription factors Insulin 1, Insulin 2, Islet 1, IA1 and MafA were highly up regulated and co-expressed (Fig. 5), which confirmed their pancreatic endocrine islet β cells characteristics. Our ESCs derived insulin-secreting cells co-express MafA similarly to the insulin-expressing cells of the 13.5-week fetal human pancreas. Therefore, the specific expression pattern of these important pancreatic hormones and transcription factors strongly indicated that pancreatic endocrine islet-like cells had been obtained in vitro at this

8 Sui Jing et al. / Journal of Medical Colleges of PLA 26 (2011) 1–12
stage. The majority of the RIP-expressing cells co-expressed β cell markers, reflecting initial commitment to the endocrine islet lineage.
To determine whether there were insulin positive or C-peptide positive cells, in addition to the gene expression by RT-PCR, the differentiation process was also characterized at the level of protein expression by immunocytochemistry using anti-C-peptide, anti-glucagon, anti-somatostatin and anti-poly-peptide antibodies. C-peptide is a genuine marker of de novo insulin production. Glucagon, somatostatin and poly-peptide were checked as negative markers of β cell development on the premise that they were hormones produced exclusively within the islets. In our study ESC-derived insulin-secreting cell clusters exhibited superior expression of c-peptide, a by-product of de novo insulin synthesis. During the
last stage, after about 20 d of differentiation, hormone-expressing cells appeared. Several days later, hormone expression became more intense and cells expressing each of the four pancreatic endocrine hormones were detected. C-peptide immunoreactive cells were detected since E20 (Fig. 2). In addition to pancreatic endocrine cells, somatostatin-expressing cells and poly-peptide- expressing cells were found in areas of pancreatic epithelium and endocrine cell differentiation. C-peptide immunoreactive cells were in greater numbers than glucagon-immunoreactive cells (Fig. 6). We observed extensive regions of cells expressing C-peptide, glucagons, somatostatin and poly-peptide across the culture slides. Most C-peptide expressing cells were found scattered within the regions of negative pancreatic epithelium cells while a small part of positive cells formed cell clusters in the central area of the slides by chamber slides. Taken together, these data suggests that we have generated cell populations with pancreatic identity from mouse embryonic stem cells.
4. Discussion
ESCs are considered as an important source for cell therapy targeted to several diseases, including diabetes. To date, significant progress has been made in deriving insulin-secreting β-like cells from embryonic stem cells. However, the cells are not fully differentiated, and there is a long way to go before they could be used as a replenishable supply of insulin-secreting β-cells for transplantation. The shortage of transplantable pancreatic islets for diabetes treatment leads us to focus on generating renewable sources of insulin-secreting β cells from pluripotential ESCs. Recent successes on cellular reprogramming [18–20] now make it possible to generate disease models from patient-specific ESCs or induced pluripotent stem cells.
Fig. 5. Gene expression analysis for the determination ofinsulin-secreting cells. Insulin-secreting cells expressedseveral pancreas endocrine specific transcription factorsaccording to our differentiation strategy. cDNA samplesprepared from E15.5 mouse embryo pancreas tissueserved as positive controls for pancreatic gene expression. The GAPDH gene was used as a housekeeping-gene standard.
E15.5 NC
PAN
Ngn3
Insulin1
Insulin2
Islet 1
IA1
MafA
GAPDH

Sui Jing et al. / Journal of Medical Colleges of PLA 26 (2011) 1–12 9
C-P
EP
DA
PI
ME
RG
E
Fig. 6. Immunofluorescence analysis for ESC-derived insulin-secreting cells. ESC-derived insulin-secreting cells were stained with C-peptide, glucagon, somatostatin and polypeptide antibodies. Control staining with secondary antibodies showed negative results (data not shown). Abbreviations: DAPI, 4',6'-diamidino-2-phenylindole; C-P, C-peptide; GLUC, glucagons; SST, somatostatin; P-P, polypeptide. Scale bars: 50 µm
P-P
DA
PI
ME
RG
E
SST
D
APT
M
ER
GE
G
LU
C
DA
PI
ME
RG
E

10 Sui Jing et al. / Journal of Medical Colleges of PLA 26 (2011) 1–12
Mouse embryonic stem cells were first isolated in 1981 [21] and have great potential for cell replacement therapy. Differentiation of ESCs in vitro progresses through a cascade of gene activation and cell type specification events that closely mirror the pathway of definitive endoderm and pancreas formation in the mammalian embryo. We have focused on characterizing the insulin-secreting endocrine cells because this cell type is most critical for developing a diabetes cell therapy. We have described a five-stage protocol for efficiently differentiating ESCs to endocrine hormone expressing cells through a series of endodermal intermediates resembling those that occur during pancreatic development in vivo. We characterized the differentiation process at the RNA and protein levels using RT-PCR and immuno- fluorescence. As apoptotic cells can take up exogenous insulin from the culture medium, we measured both insulin as well as C-peptide mRNA to demonstrate de novo synthesis of insulin [22]. C-peptide-positive cells were assessed by immuno- cytochemistry.
The pancreatic and duodenal homeobox gene1 Pdx1 is a regional endoderm marker whose expression marks the dorsal and ventral pancreatic buds, as well as a part of the stomach and duodenal endoderm [23]. Within the mouse embryonic foregut, pancreatic fate is first specified by the expression of Pdx1 in narrow patches of dorsal and ventral endoderm [24, 25]. By late gestation, Pdx1 expression is extinguished in the pancreatic ductal epithelium and retained predominantly in insulin- expressing β cells. In mice homozygous for a Pdx1 loss-of-function mutation, the development of the pancreas is blocked at a very early stage, the pancreas fails to form and results in extreme hyperglycemia and perinatal death [26]. Thus, activation of Pdx1 is considered a prerequisite for pancreatic differentiation in vitro and should precede the progressive expression of more mature markers of the endocrine lineage, including Ngn3, Nkx6.1, Pax4, and Isl1 [25]. We observed Pdx1+
pancreatic endoderm cells appeared as early as day 12 of differentiation. After foregut endoderm cultures are exposed to RA, CYC, DAPT and exendin 4, there is a rapid induction of many important transcription factors which have been shown to be essential for correct specification of the pancreatic anlagen and to be important in pancreatic endoderm development, including Pdx1, Ptf1a, Nkx6.1, Pax6 and Pax4. The coordinate expression of these genes appears to closely imitate the timed expression of these factors during this decisive phase of pancreatic development.
Ngn3, the earliest islet cell-specific trans- cription factor in embryonic development, is required for development of all islet endocrine cell types and is normally expressed in a subset of pancreatic progenitors marking commitment to the endocrine lineage. In the embryo, Ngn3 is expressed in endocrine progenitors [27]. Ngn3 mRNA is first detected in the dorsal pancreatic epithelium at embryonic day 9 (E9.0) in the mouse, as the pancreas develops, the number of Ngn3+ cells increases and peaks at E15.5 [28]. Ngn3 is expressed in all endocrine progenitors19, initiating a cascade of transcription-factor expression that controls endocrine cell differentiation. The critical transcription factor genes include Nkx2.2, Nkx6.1, Pax4, Pax6 and Isl1. The nascent endocrine cells migrate from the branched epithelium into the surrounding mesenchyme to form the islets of Langerhans. Islets comprise five endocrine cell types: α, β, δ, PP and ε cells, which produce the hormones glucagon, insulin, somatostatin, pan- creatic polypeptide and ghrelin. Once formed, the hormone-expressing endocrine cells undergo further differentiation to a mature functional state, which, for a β-cell, involves the ability to release insulin in response to elevated glucose. There are 2 nonallelic insulin genes (insulin I and II) expressed in multiple sites in mice during development. Neuronal cells express only insulin II, whereas pancreas expresses both insulin I and II [29]. Thus, insulin I, when combined with insulin II and other lineage markers,

Sui Jing et al. / Journal of Medical Colleges of PLA 26 (2011) 1–12 11
can be used to trace the development of pancreatic β cells. In our study, after the differentiated islet progenitor cells were exposed to Nicotinamide and exendin 4, the cells rapidly began to express high levels of Ngn3, Insulin1, Insulin2, IA1 and MafA, expression of this combination of genes is indicative of insulin- expressing cells specialization.
Since it was shown in 1934 that zinc is a component of insulin crystals [30], a relationship between zinc and diabetes mellitus has been proposed. From then on, numerous studies trying to elucidate the role of zinc in diabetes mellitus have been conducted with the aim of identifying new causal mechanisms and new therapeutical options [31]. Zinc is an essential trace element crucial for the function of more than 300 enzymes and it is important for cellular processes like cell division and apoptosis and seems to exert insulin-like effects by supporting the signal transduction of insulin [31]. Zinc is one of the essential trace elements for eukaryotes and has numerous physiological functions [32]. Zinc deficiency was found to increase the risks of diabetes and diabetic complications [33]. We have demonstrated that zinc supplementation in embryonic stem cell line can significantly induce insulin-expressing cells differentiation, suggesting that induction of insulin synthesis by zinc supplementation may be a potential approach for obtaining transplantable cell-replacement β cells from embryonic stem cells for diabetes cell therapy.
We have demonstrated that ESCs can efficiently differentiate into insulin-secreting endocrine cells through a series of endodermal intermediates that are similar to those that occur in vivo. The results of our study are similar in many respects to previous studies in the mouse and human ES cell lines [34, 35]. However, some differences are also observed. These differences could be due to the very different design of the study or to the differences between mouse and human ES cell lines. Our study showed that zinc was an effective inducer for gene and
protein expression of insulin-secreting cells derived from ESCs. However, there was lack of direct evidence for the role of zinc induced ESC-derived insulin-secreting cells. Future studies will focus on exploring the exact role of zinc during the development of pancreas and try to improve the efficiency of ESCs differentiation.
References 1. Shi Y, Hou L, Tang F, et al. Inducing embryonic
stem cells to differentiate into pancreatic beta cells by a novel three-step approach with activin A and all-trans retinoic acid. Stem Cells, 2005; 23 (5): 656–662.
2. Rajagopal J, Anderson WJ, Kume S, et al. Insulin staining of ES cell progeny from insulin uptake. Science, 2003; 299 (5605): 363.
3. Roche E, Sepulcre P, Reig JA, et al. Ectodermal commitment of insulin-producing cells derived from mouse embryonic stem cells. FASEB J, 2005; 19 (10): 1341–1343.
4. Soria B. In-vitro differentiation of pancreatic beta-cells. Differentiation, 2001; 68 (4–5): 205–219.
5. Lumelsky N, Blondel O, Laeng P, et al. Differentiation of embryonic stem cells to insulin- secreting structures similar to pancreatic islets. Science, 2001; 292 (5520): 1389–1394.
6. D'Amour KA, Bang AG, Eliazer S, et al. Production of pancreatic hormone-expressing endocrine cells from human embryonic stem cells. Nat Biotechnol, 2006; 24 (11): 1392–1401.
7. Kroon E, Martinson LA, Kadoya K, et al. Pancreatic endoderm derived from human embryonic stem cells generates glucose-responsive insulin-secreting cells in vivo. Nat Biotechnol, 2008; 26 (4): 443–452.
8. Nicolson TJ, Bellomo EA, Wijesekara N, et al. Insulin storage and glucose homeostasis in mice null for the granule zinc transporter ZnT8 and studies of the type 2 diabetes-associated variants. Diabetes, 2009; 58 (9): 2070–2083.
9. Chimienti F, Devergnas S, Favier A, et al. Identification and cloning of a beta-cell-specific zinc transporter, ZnT-8, localized into insulin secretory granules. Diabetes, 2004; 53 (9): 2330–2337.
10. Emdin SO, Dodson GG, Cutfield JM, et al. Role of

12 Sui Jing et al. / Journal of Medical Colleges of PLA 26 (2011) 1–12
zinc in insulin biosynthesis. Some possible zinc- insulin interactions in the pancreatic β-cell. Diabetologia, 1980; 19(3): 174–182.
11. Duester G. Retinoic acid synthesis and signaling during early organogenesis. Cell, 2008; 134 (6): 921–931.
12. Hart A, Papadopoulou S, Edlund H. Fgf10 maintains notch activation, stimulates proliferation, and blocks differentiation of pancreatic epithelial cells. Dev Dyn, 2003; 228 (2): 185–193.
13. Mfopou JK, Bouwens L. Hedgehog signals in pancreatic differentiation from embryonic stem cells: revisiting the neglected. Differentiation, 2008; 76 (2): 107–117.
14. Shen CN, Marguerie A, Chien CY, et al. All-trans retinoic acid suppresses exocrine differentiation and branching morphogenesis in the embryonic pancreas. Differentiation, 2007; 75 (1): 62–74.
15. Noguchi H. Stem cells for the treatment of diabetes. Endocr J, 2007; 54 (1): 7–16.
16. Vaca P, Berná G, Araujo R, et al. Nicotinamide induces differentiation of embryonic stem cells into insulin-secreting cells. Exp Cell Res, 2008; 314 (5): 969–974.
17. Kawaguchi Y, Cooper B, Gannon M, et al. The role of the transcriptional regulator Ptf1a in converting intestinal to pancreatic progenitors. Nat Genet, 2002; 32 (1): 128–134.
18. Takahashi K, Tanabe K, Ohnuki M, et al. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell, 2007; 131 (5): 861–872.
19. Yu J, Vodyanik MA, Smuga-Otto K, et al. Induced pluripotent stem cell lines derived from human somatic cells. Science, 2007; 318 (5858): 1917–1920.
20. Park IH, Zhao R, West JA, et al. Reprogramming of human somatic cells to pluripotency with defined factors. Nature, 2008; 451 (7175): 141–146.
21. Martin GR. Isolation of a pluripotent cell line from early mouse embryos cultured in medium conditioned by teratocarcinoma stem cells. Proc Natl Acad Sci USA, 1981, 78 (12): 7634–7638.
22. Sipione S, Eshpeter A, Lyon JG, et al. Insulin expressing cells from differentiated embryonic stem cells are not beta cells. Diabetologia, 2004; 47 (3): 499–508.
23. Shiraki N, Yoshida T, Araki K, et al. Guided differentiation of embryonic stem cells into
Pdx1-expressing regional-specific definitive endoderm. Stem Cells, 2008; 26 (4): 874–885.
24. Ashizawa S, Brunicardi FC, Wang XP. PDX-1 and the pancreas. Pancreas, 2004; 28 (2):109-120.
25. Jensen J. Gene regulatory factors in pancreatic development. Dev Dyn, 2004; 229 (1): 176–200.
26. Offield MF, Jetton TL, Labosky PA, et al. PDX-1 is required for pancreatic outgrowth and differentiation of the rostral duodenum. Development, 1996; 122 (3): 983–995.
27. Gu G, Dubauskaite J, Melton DA. Direct evidence for the pancreatic lineage: NGN3+ cells are islet progenitors and are distinct from duct progenitors. Development, 2002; 129 (10): 2447–2457.
28. Lee CS, De León DD, Kaestner KH, et al. Regeneration of pancreatic islets after partial pancreatectomy in mice does not involve the reactivation of neurogenin-3. Diabetes, 2006; 55(2): 269–272.
29. Ku HT, Zhang N, Kubo A, et al. Committing embryonic stem cells to early endocrine pancreas in vitro. Stem Cells, 2004; 22 (7): 1205–1217.
30. Scott DA. Crystalline insulin. Biochem J, 1934; 28 (4):1592–1602.
31. Jansen J, Karges W, Rink L. Zinc and diabetes-clinical links and molecular mechanisms. J Nutr Biochem, 2009; 20 (6): 399–417.
32. Song Y, Wang J, Li XK, et al. Zinc and the diabetic heart. Biometals, 2005; 18 (4): 325–332.
33. Soinio M, Marniemi J, Laakso M, et al. Serum zinc level and coronary heart disease events in patients with type 2 diabetes. Diabetes Care, 2007; 30 (3): 523–528.
34. Planas R, Carrillo J, Sanchez A, et al. Gene expression profiles for the human pancreas and purified islets in Type 1 diabetes: new findings at clinical onset and in long-standing diabetes. Clin Exp Immunol, 2010; 159 (1): 23–44.
35. Yoshida T, Murata K, Shiraki N, et al. Analysis of gene expressions of embryonic stem-derived Pdx1-expressing cells: Implications of genes involved in pancreas differentiation. Dev Growth Differ, 2009; 51 (4): 463–472.
(Editor Wu Tao)