developmental expression patterns of candidate cofactors for vertebrate six family transcription...
TRANSCRIPT

a PATTERNS & PHENOTYPES
Developmental Expression Patterns ofCandidate Cofactors for Vertebrate Six FamilyTranscription FactorsKaren M. Neilson,1* Francesca Pignoni,2 Bo Yan,1 and Sally A. Moody1
Six family transcription factors play important roles in craniofacial development. Their transcriptionalactivity can be modified by cofactor proteins. Two Six genes and one cofactor gene (Eya1) are involved inthe human Branchio-otic (BO) and Branchio-otic-renal (BOR) syndromes. However, mutations in Six andEya genes only account for approximately half of these patients. To discover potential new causativegenes, we searched the Xenopus genome for orthologues of Drosophila cofactor proteins that interactwith the fly Six-related factor, SO. We identified 33 Xenopus genes with high sequence identity to 20 of the25 fly SO-interacting proteins. We provide the developmental expression patterns of the Xenopus ortho-logues for 11 of the fly genes, and demonstrate that all are expressed in developing craniofacial tissueswith at least partial overlap with Six1/Six2. We speculate that these genes may function as Six-interactingpartners with important roles in vertebrate craniofacial development and perhaps congenital syndromes.Developmental Dynamics 239:3446–3466, 2010. VC 2010 Wiley-Liss, Inc.
Key words: Branchio-otic syndrome; Branchio-otic-renal syndrome; placodes; craniofacial development; Eya;Groucho; Sobp; Gadd45; Six1; Six2; sine oculis; neural crest
Accepted 11 October 2010
INTRODUCTION
Placodes are patches of thickenednon-neural ectoderm that form lateralto the neural crest in the head of thevertebrate embryo. They give rise tosensory organs (olfactory epithelium,lens, auditory-vestibular organs, andlateral line), and contribute to the an-terior pituitary and cranial sensoryganglia (Webb and Noden, 1993;Schlosser and Northcutt, 2000; Bakerand Bonner-Fraser, 2001; Schlosser,2006; Moody, 2007). Placodes arisefrom a common precursor domain of
non-neural ectoderm that surrounds
the cranial portion of the neural plate,
called the preplacodal ectoderm (PPE;
Knouff, 1935; Streit, 2004, 2007;
Schlosser, 2006). Among the earliest
genes expressed throughout the PPE
are Six1, Six2, and Six4 (Esteve and
Bovolenta, 1999; Kobayashi et al.,
2000; Pandur and Moody, 2000;
Ghanbari et al., 2001; Schlosser and
Ahrens, 2004; Sato et al., 2010), sug-
gesting that they have important
roles in placodal development. The
expression of Six1 in the PPE domi-
nantly promotes a placodal fate,
whereas knock-down of Six1 or
repression of its targets results in cra-
niofacial defects in frog, chick, and
mouse (Laclef et al., 2003; Brugmann
et al., 2004; Christophorou et al.,
2009); Six2- or Six4-null mice do not
have reported craniofacial defects
(Ozaki et al., 2001; Fogelgren et al.,
2009; Self et al., 2009), but mis-
expression of Six2 leads to fronto-
nasal dysplasia (Fogelgren et al.,
2008). Two human syndromes, the
branchio-otic (BO) and branchio-oto-
1Department of Anatomy and Regenerative Biology, The George Washington University, School of Medicine and Health Sciences,Washington, DC2Department of Ophthalmology, Center for Vision Research, SUNY Upstate Medical University, Syracuse, New YorkGrant sponsor: NIH; Grant numbers: R03 HD055321, NSF IOS-0817902, NIH R01 EY1316709.*Correspondence to: Karen M. Neilson, Department of Anatomy and Regenerative Biology, The George WashingtonUniversity, School of Medicine and Health Sciences, 2300 I (eye) Street, NW, Washington, DC 20037.E-mail: [email protected]
DOI 10.1002/dvdy.22484Published online 10 November 2010 in Wiley Online Library (wileyonlinelibrary.com).
DEVELOPMENTAL DYNAMICS 239:3446–3466, 2010
VC 2010 Wiley-Liss, Inc.
Dev
elop
men
tal D
ynam
ics

renal (BOR) syndromes, which arecharacterized by craniofacial defectsand hearing loss, can be caused bymutations in SIX1, SIX5, or EYA1(Abdelhak et al., 1997; Kumar et al.,1997; Rodriguez-Soriano, 2003; Rufet al., 2004; Hoskins et al., 2007).Six1- and Eya1-null mutant mice like-wise exhibit severe defects in the de-velopment of some placodal struc-tures (Oliver et al., 1995; Vincentet al., 1997; Johnson et al., 1999; Xuet al., 1999, 2002; Laclef et al., 2003;Zheng et al., 2003; Ozaki et al., 2004).However, mutations in these genesaccount for less than half of the casesof BO or BOR, indicating that defectsin other genes contribute to thesesyndromes.
The founding member of the Sixgene family is Drosophila sine oculis(so; Cheyette et al., 1994; Serikakuand O’Tousa, 1994). In vertebrates,there are six different members of theSix gene family (Six1–6) that areexpressed in a variety of tissues dur-ing development (reviewed in Kawa-kami et al., 2000; Brugmann andMoody, 2005). Each of the differentSix proteins contains two highly con-served regions: a Six-type homeodo-main (Six-HD) that binds to DNA andan N-terminal Six domain (SD) thatcan bind with other proteins (cofac-tors) that are unable to interactdirectly with DNA (Pignoni et al.,1997; Kawakami et al., 2000; Kobaya-shi et al., 2001). These cofactorsincrease the DNA binding specificityof the Six proteins and modulate Sixfunction as either coactivators or core-pressors of transcription (Zhu et al.,2002; Tessmar et al., 2002). For exam-ple, during PPE development wheneither Eya1 or Eya2 is coexpressedwith Six1, other placode genes are up-regulated, and when Groucho is coex-pressed, neural crest and epidermalgenes are down-regulated (Brugmannet al., 2004; Christophorou et al.,2009). Eya1 and Six1 also coopera-tively regulate the onset of neurogen-esis in the placodes that give rise tothe cranial ganglia (Schlosser et al.,2008).
Searches for additional Six1 part-ners in vertebrates have met withvery limited success (Rual et al.,2005), whereas yeast 2-hybrid screensof the smaller, more manageable Dro-sophila proteome have resulted in the
identification of 25 factors that inter-act with the fly SO protein (Pignoniet al., 1997; Giot et al., 2003; Kenyonet al., 2005). Seven factors, includingEya and Groucho, are nuclear/tran-scriptional regulators, 14 are cyto-plasmic and 4 are of unknown func-tion (Table 1). However, there is verylittle information available regardingthe developmental expression of mostof these genes or their protein func-tions in conjunction with Six proteins;the exceptions are Eya, Groucho, andSOBP (Pignoni et al., 1997; Heanueet al., 1999; Ohto et al., 1999; Kobaya-shi et al., 2001; Giot et al., 2003; Ken-yon et al., 2005). To identify new,potential Six-interacting proteinsthat might have roles in vertebratecraniofacial development, we used thesequences of the Drosophila SO-inter-acting proteins to search for putativeXenopus orthologues. Herein, weidentify 33 Xenopus genes related bysequence to 20 of the Drosophila SO-interacting proteins, and demonstratethat a large number of these areexpressed during Xenopus craniofa-cial development. Thereby, thesegenes are potential candidates forregulating normal craniofacial devel-opment and being involved in cranio-facial birth defects.
RESULTS AND DISCUSSION
Drosophila SO and Xenopus
Six1 and Six2 Are Highly
Similar in the Six Domain
It is well documented that SO/Six pro-teins bind to Eya and Groucho pro-teins (Pignoni et al., 1997; Heanueet al., 1999; Ohto et al., 1999; Kobaya-shi et al., 2001; Zhu et al., 2002;Lopez-Rios et al., 2003), and we previ-ously showed in Xenopus that coex-pression of either Eya1 or Grouchocauses Six1 to have significantly dif-ferent effects on PPE/placode develop-ment (Brugmann et al., 2004). Dro-sophila SO and Xenopus Six1/Six2proteins are highly conserved throughtheir Six-HD domains (Pandur andMoody, 2000), which is consistentwith the observations that SO, Six1,and Six2 share DNA binding sitespecificity (Kawakami et al., 1996;Spitz et al., 1998; Silver et al., 2003;F. Pignoni, unpublished observa-tions). Figure 1 illustrates that SO,
Six1, and Six2 also are highly con-served throughout their protein–pro-tein interaction domains (the SD). Infact, only four amino acid substitu-tions are nonconserved; all otherchanges in sequence involve con-served or semiconserved amino acids.This suggests that these three pro-teins are likely to share cofactor bind-ing specificity.
Identification of Putative
Xenopus Orthologues of
Drosophila SO-Interacting
Proteins
In Drosophila, 25 factors that caninteract with SO have been identifiedby yeast 2-hybrid screens (Pignoniet al., 1997; Giot et al., 2003; Kenyonet al., 2005). We performed databasesearches for putative Xenopus ortho-logues and identified between one andfour genes with amino acid sequencehomology to 20 of the fly proteins (Ta-ble 1). Two members of the Eya genefamily (Eya1, David et al., 2001;Eya3, Kriebel et al., 2007), four Grou-cho-related genes (Grg4, Grg5, ESG1,ESG2; Choudhury et al., 1997; Mole-naar et al., 2000), two C15-relatedgenes (Hox11, Hox11L2; Pattersonand Krieg, 1999), and one Gadd45-related gene (Gadd45g; de la Calle-Mustienes et al., 2002) are previouslyreported in Xenopus. In addition tothese, we identified two additionalEya genes (Eya2, Eya4), three geneswith homology to SOBP, two geneseach with homology to CG5033 andCG5665, two additional Gadd45genes (Gadd45a, Gadd45b) and onegene each with homology to CG9083,CG17265, CG1218, CG1135, CG7878,CG33090, RhoGEF2, Chc, CKIIb,CG10576, VhaAC39, RhoGAP93B,and RpS27 (Table 1).
Xenopus Candidate Six-
Interacting Genes Are
Expressed in Developing
Craniofacial Tissues
Xenopus Six1 is initially expresseddiffusely in the dorsal embryonic ecto-derm, and then is highly expressedthroughout the PPE (Pandur andMoody, 2000); weak expression hasbeen noted in the early neural plateas well (Schlosser and Ahrens, 2004).
EXPRESSION OF CANDIDATE SIX COFACTORS 3447
Dev
elop
men
tal D
ynam
ics

TABLE
1.Droso
philaSO-InteractingProtein
sandTheir
Puta
tiveXenopusOrth
ologues
SO-interactingproteinsa
Protein
activityb
Interaction
confiden
cescorea
PutativeXen
opusortholog
ues
c(X
l¼
X.laevis;
Xt¼
X.trop
icalis)
Eyes
absent
Transcription
factor
0.624
Eya1(gi:13591786,Xl)
Eya2(gi:134024326,Xt)
Eya3(gi:58702065,Xl)
Eya4(X
tgen
omic
sequen
ceENSXETG0000000011
0in
Ensembldatabase)
CG15459
Unknow
n,on
lyex
pressed
intestis
andadult
infly
0.430
Iden
tified
only
inDrosophila
CG6688
Ubiquitin-dep
enden
tprotein
catabolism,PDZ
dom
ain
0.421
Iden
tified
only
inDrosophila
CG9083
Unknow
n0.415
Nov
elEnsemblpredication
(Xtgen
omic
sequen
ceENS-
XETT00000000661in
Ensembldatabase)
Grouch
oTranscription
factor
0.415
Grg4/Tle4(gi:50925318,Xl)
ESG1/Tle3(gi:50414464,Xl)
ESG2(gi:2408194,Xl)
Grg5/AES(gi:38174466,Xl)
CG17265
RNA
interferen
ce0.413
Coiledcoildom
ain
protein
ccd85c
(gi:169642663,Xt;gi:66912014,Xl)d
CG5466
Cellgrowth,protein
binding
0.406
Iden
tified
only
inDrosophila
C15
Transcription
factor
0.404
Hox
11(gi:10185813,Xl)
Hox
11L2(gi:10185811
,Xl)
CG1218
Znfinger
protein,unknow
n0.403
Sim
ilarto
C4orf27EST(gi:8579074Xl)
Pif1B
PFTAIR
E-interactingfactor
1B,protein
bind-
ing,unknow
n0.402
Iden
tified
only
inDrosophila
SOBP
SO
BindingProtein,transcription
factor
0.394
SOBP(gi:159155685,Xt)
Zincfinger
protein
198(gi:49903401,Xl)
Zincfinger
protein
LOC414497(gi:80479434,Xl)
CG4807(A
bru
pt)
Transcription
factor,BTB
dom
ain
forprotein
dim
erization
0.393
Iden
tified
only
inDrosophila
CG11
35
Forkhea
dassociateddom
ain,putativenuclea
rsignalingdom
ain
0.389
LOC100049093(gi:126631279,Xl)
CG7878
Dea
dbox
RNA
helicase
0.386
Ddx43(gi:11
2419338,Xt;gi:11
8404345,Xt)e
CG5665
Lipop
rotein
lipase
0.373
Lipase
mem
ber
H(X
tgen
omic
sequen
ceENS-
XETG00000005849ENSEMBLdatabase)
Phospholipase
A1mem
ber
A(X
tgen
omic
sequen
ceENSXETG00000018070ENSEMBLdatabase)
CG33090
Hydroxymethylglutaryl-CoA
reductase
class
I/II,bileacidglucosidase
0.356
GBA2(gi:169642398Xt)
RhoG
EF2
Rhoguaninenucleotideex
changefactor
0.331
Guaninenucleotideex
changefactor
(gi:148223172,Xl)
Chc
Clathrinhea
vych
ain,vesicle
med
iated
transp
ort
0.330
MCG80936(gi:4911
5531,Xl)
CKIIb
Casein
kinase
IIregulatory
subunit,role
inWntsignaling
0.327
Casein
kinase
IIbsu
bunit
(gi:50414548,Xl)
CG10576
Methionylaminop
eptidase
activity;proteolysis
0.325
Proliferation
-associated2G4(gi:54311186,Xl)
VhaAC39
Protontransp
ort,ATPase
0.322
ATPase
Protontransp
ortinglysosomal38kD
VO
subunit
d1(gi:27769219,Xl)
3448 NEILSON ET AL.
Dev
elop
men
tal D
ynam
ics

As individual placodes form, Six1 isexpressed in each except the lens,whereas Xenopus Six2 expression isinitially concentrated in the olfactoryand otic placodes (Pandur and Moody,2000; Ghanbari et al., 2001). At tailbud to larval stages, both are addi-tionally expressed in the cranial gan-glia, somites, hypaxial muscle precur-sors, and nephric mesoderm, and Six1expression is reported in the migrat-ing lateral line primordium. Ghanbariet al. (2001) report Six1 and Six2expression in the branchial arch mes-oderm, whereas Pandur and Moody(2000) did not observe this for Six1.To determine whether the putative
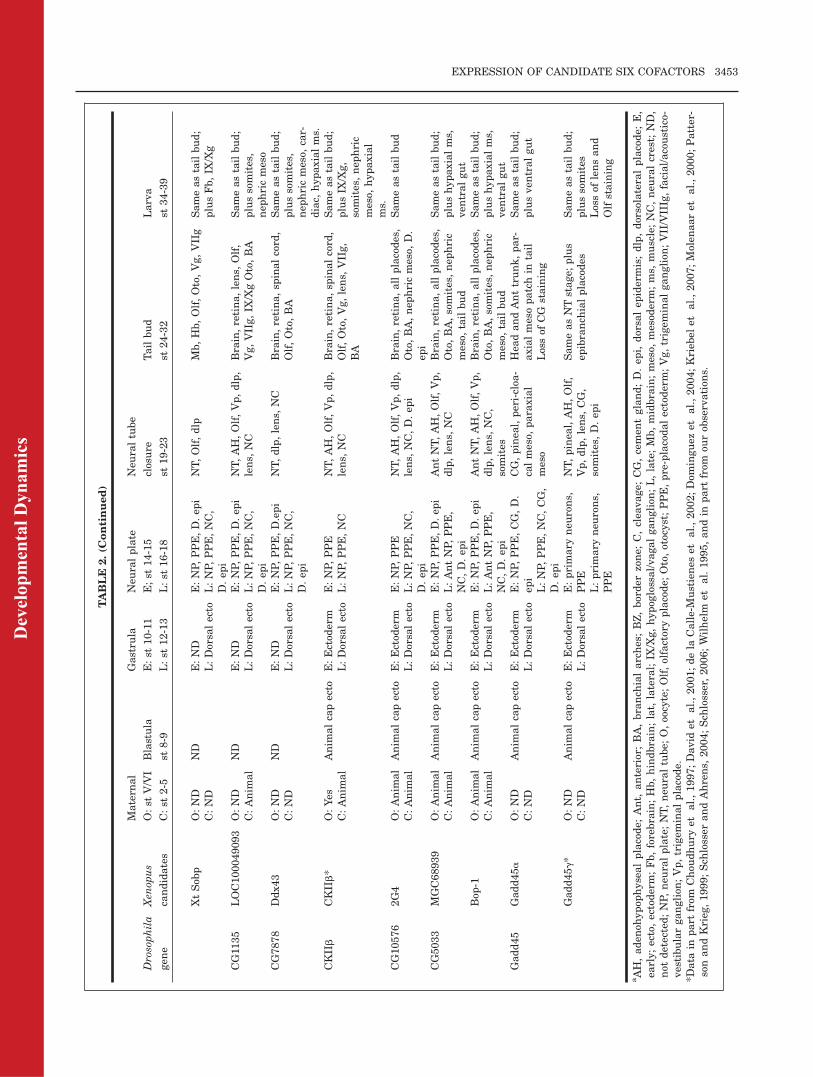
Xenopus orthologues of DrosophilaSO-interacting proteins are expressedin tissue domains that overlap withSix1/Six2 gene expression, we per-formed in situ hybridization (ISH)assays for candidates of 11 of the Dro-sophila genes. We did not study theexpression patterns of the putativeXenopus orthologues of the remainingDrosophila genes because they are:(1) very broadly expressed in nearlyall tissues in other organisms (Rho-GEF2, Chc, RpS27); (2) reportedin the Xenopus Unigene expressedsequence tag (EST) expression profileto not overlap with Six1 expression(CG33090, Unigene Str.38506;VhaAC39,Unigene Xl.77521); (3) only availableas genomic sequence in the X. tropica-lis database (Eya4, CG9083, CG5665,RhoGAP93B, Gadd45b); or (4) onlyavailable as an EST with homologyto only a very short region of theDrosophila protein (CG1218), whichreduced our confidence that this is atrue orthologue. As summarized inTable 2, we found that the candidateXenopus Six-interacting genes repre-senting 11 of the Drosophila SO-inter-acting proteins are expressed incraniofacial tissues, most in patternsthat overlap extensively with Six1/Six2 gene expression. In the followingsections, we present the expressionpattern of each of the 11 sets ofputative Xenopus orthologues, andsummarize what is known about theirexpression patterns and function infly and in other vertebrates.
Eya genes.
We identified three members of theXenopus Eya gene family (Eya1,
TABLE
1.(C
ontinued)
SO-interactingproteinsa
Protein
activityb
Interaction
confiden
cescorea
PutativeXen
opusortholog
ues
c(X
l¼
X.laevis;
Xt¼
X.trop
icalis)
RhoG
AP93B
RhoGTPase
activatorprotein
0.312
RhoG
AP93B
(Xtgen
omic
sequen
ceENS-
XETG00000007422Ensembldatabase)
CG5033
Ribon
ucleoprotein
binding,transcription
factor
0.294
HypotheticalMGC68939(gi:33416659,Xl)
Block
ofproliferation
(gi:2877251,Xl)
Gadd45
Cellcycleregulator
0.254
gadd45a(gi:27694852,Xl)
gadd45g(gi:148223518,Xl)
gadd45g(X
tgen
omic
sequen
ceENSXETT00000008029
ENSEMBLdatabase)
RpS27
Cytosolicsm
allribosom
alsu
bunit
0.202
RpS27(gi:297124,Xl)
aDroID
,TheDrosophilaIn
teractionsDatabase,Version
5.0,http://w
ww.droidb.org/Index
.jsp
bFlybase,FB2010_0
4,http://flybase.org
c IftheX.laevis
gen
ewasiden
tified
intheBLASTsearch,theX.trop
icalisgen
eis
not
listed
(exceptforCG17265,seenoted).
dBothX.laevis
andX.trop
icalisgen
esare
listed
forCG17265(ccd
85c)
because
theX.trop
icalisgen
ehasahigher
aminoacidsequen
ceiden
tity
toCG17265ov
eritsen
tire
cod-
ingregion.
eThesetw
oNCBIGen
Banken
triesforDdx4
3are
iden
ticalin
sequen
ce,andthusare
countedason
lyon
egen
ein
thetext.
EXPRESSION OF CANDIDATE SIX COFACTORS 3449
Dev
elop
men
tal D
ynam
ics

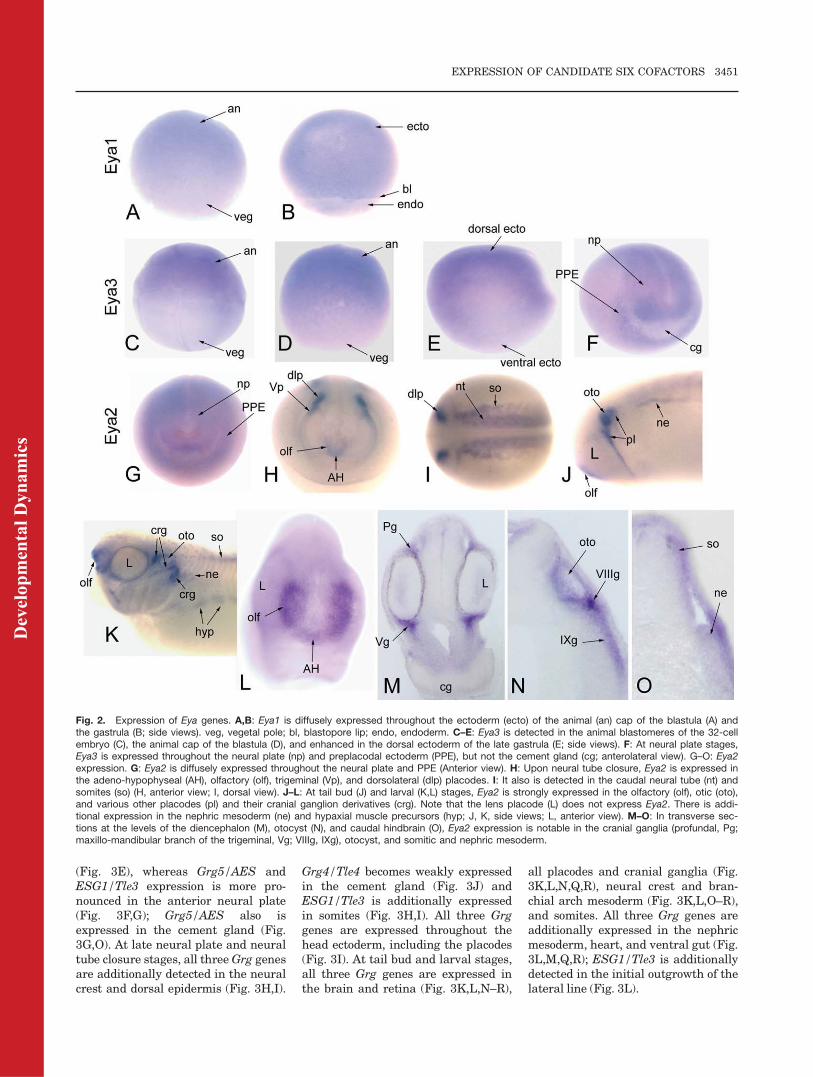
Eya2, Eya3); the Xenopus tropicalisgenomic database annotates an Eya4gene (Table 1), but no cDNA clonesare currently available. The expres-sion of Eya1 was previously reported(David et al., 2001; Schlosser andAhrens, 2004; Schlosser, 2006), and issummarized in Table 2. Our ISHanalyses confirm the reported pat-terns and additionally show weakexpression throughout the blastulaanimal cap ectoderm (Fig. 2A), earlygastrula ectoderm (Fig. 2B), dorsalectoderm of the late gastrula, andnephric mesoderm of the larva (Table2). The expression of Xenopus Eya3was previously reported (Kriebelet al., 2007). Unlike Eya1, it isdetected maternally in the animalregion of oocytes (Table 2) and animalblastomeres of cleavage stageembryos (Fig. 2C). It should be notedthat the lack of vegetal pole stainingfor these genes (and all subsequentgenes described) could result frompoor probe penetration into the yolkyendodermal cells rather than lack ofexpression. However, maternal ani-mal hemisphere expression is consist-ent with the later zygotic expressionin the ectodermal precursors in theanimal half of blastula stage embryosand throughout the embryonic ecto-derm in the early gastrula. LikeEya1, we observed that Eya3 isexpressed in the blastula animal capectoderm (Fig. 2D) and gastrula ecto-derm, most prominently on the dorsalside (Fig. 2E). In addition, Eya3 isstrongly expressed in the neural plateand PPE (Fig. 2F), anterior neuraltube, neural crest and the branchialarches derived from them, and thelens (Table 2; Kriebel et al., 2007).
Xenopus Eya2 expression has not pre-viously been reported. Maternal Eya2mRNA is not detected. Like the otherEya genes, Eya2 is weakly expressedin the blastula animal cap, early gas-trula ectoderm, and enhanced in thedorsal ectoderm of the late gastrula(Table 2). By neural plate stages, it isdetected diffusely throughout theneural plate and in the PPE (Fig. 2G).As the neural tube closes, it isdetected in the neural tube, multipleplacodes, and somites (Fig. 2H,I).During early tail bud stages, it isadditionally expressed in the otocystand nephric mesoderm (Fig. 2J), andby late tail bud/larval stages, it isadditionally expressed in the cranialganglia, epibranchial placodes, andhypaxial muscle precursors (Fig.2K–O).
These expression patterns are con-sistent with reports in other verte-brates that Eya genes are expressedin ectodermal placodes, somites,brain, nephric mesoderm, and hypax-ial muscle precursors, i.e., tissuesthat express Six genes (Xu et al.,1997, 2003; Sahly et al., 1999; Schon-berger et al., 2005; Soker et al., 2008;Ishihara et al., 2008). The develop-mental functions of Eya are well-stud-ied in Drosophila; it plays a key rolein early eye formation, head morpho-genesis, gametogenesis, and muscledevelopment (Bonini et al., 1993,1997, 1998; Pignoni et al., 1997; Baiand Montell, 2002; Fabrizio et al.,2003; Liu et al., 2009). The involve-ment of vertebrate Eya proteins incraniofacial development also is well-established. In Xenopus and chick,Eya1 and Eya2 are involved in estab-lishing the PPE domains (Brugmann
et al., 2004; Christophorou et al.,2009). Mouse Eya1 is necessary fornormal development of the ear, kid-ney, thymus, parathyroid, thyroid,and somitic muscle (Xu et al., 1999,2002, 2003; Johnson et al., 1999), andmutations in human Eya1 cause somecases of BO/BOR (Abdelhak et al.,1997). Eya4-deficient mice exhibitabnormal middle ear cavities, Eusta-chian tube dysmorphology and are amodel for heritable otitis media (Dep-reux et al., 2008); mutations inhuman Eya4 result in sensorineuralhearing loss (Wayne et al., 2001; Pfis-ter et al., 2002). However, the rolesfor mammalian Eya2 and Eya3 in cra-niofacial development have not yetbeen established, although they arerequired in other tissues (Grifoneet al., 2007; Soker et al., 2008); therealso are no reported human defectsinvolving Eya2 or Eya3 to date. None-theless, the expression of Eya2 andEya3 in the Xenopus and chick PPEand placodes, and the demonstratedability of each of the four vertebrateEya proteins to interact with Six1,Six2, and/or Six4 (Heanue et al.,1999; Ohto et al., 1999; Ikeda et al.,2002; Li et al., 2003; Schonbergeret al., 2005; Abe et al., 2009) predictthat all four Eya genes are likely toact as Six gene cofactors, perhapswith redundant roles, during cranio-facial development.
Groucho-related genes.
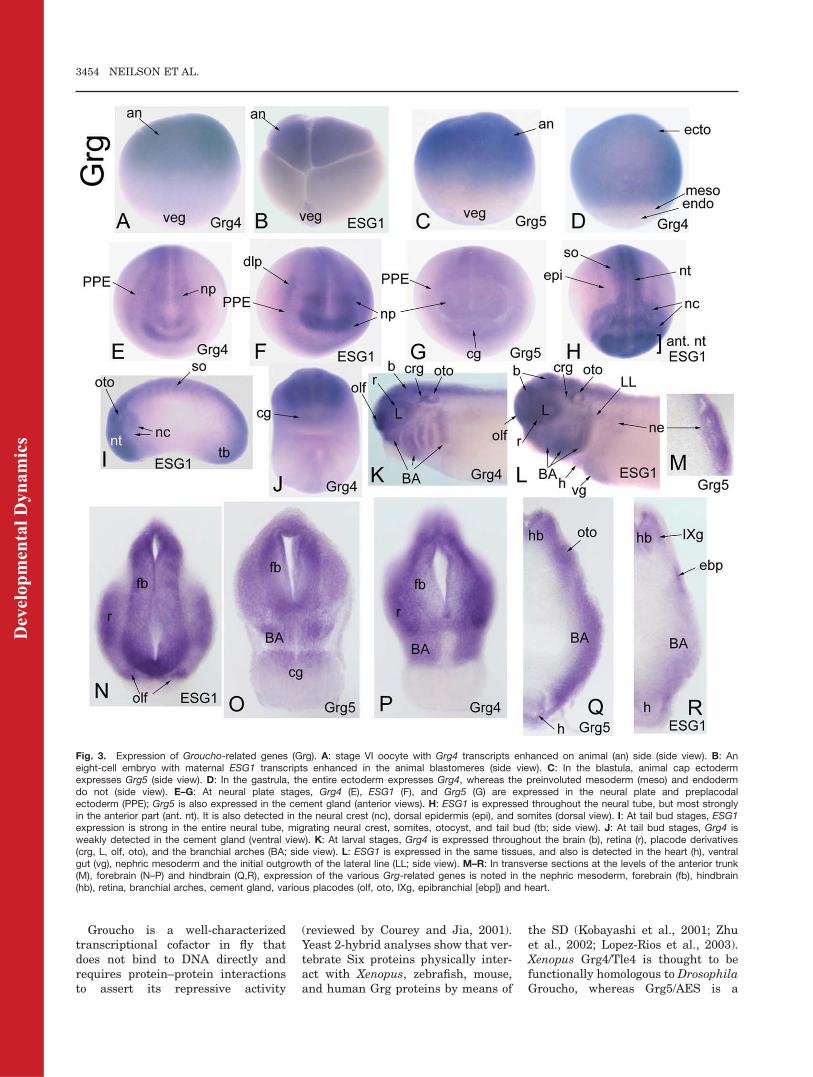
Four Groucho-related (Grg) geneshave been reported in Xenopus. ESG2is not expressed in embryos (Choud-hury et al., 1997), whereas the otherthree (Grg4/Tle4, Grg5/Aes, ESG1/Grg3/Tle3) have overlapping, but notidentical, embryonic expression pat-terns (Table 2; Fig. 3). All threemRNAs are detected by Northernblots to be maternally expressed(Choudhury et al., 1997; Molenaaret al., 2000); we confirmed this by ISHand show that each mRNA is localizedto the animal hemisphere of oocytesand animal blastomeres of cleavageembryos (Fig. 3A,B; Table 2). All threeGrg genes have enhanced expressionin the animal cap ectoderm of theblastula (Fig. 3C; Table 2) and gas-trula (Fig. 3D). At early neural platestages, Grg4/Tle4 is expressedthroughout the neural plate and PPE
Fig. 1. Fly SO, Xenopus Six1 and Xenopus Six2 proteins have a high level of amino acid iden-tity (82.5%; black boxes) throughout their Six domain (SD, aa 1-120), the protein–protein interac-tion domain. Only four amino acid substitutions are nonconserved (arrowheads). All otherchanges in sequence involve conserved (:) or semiconserved (.) amino acids. The alignment wasperformed using Clustal W version 2.0 (Larkin et al., 2007). Dm, Drosophila melanogaster; Xl,Xenopus laevis; Xt, Xenopus tropicalis.
3450 NEILSON ET AL.
Dev
elop
men
tal D
ynam
ics
(Fig. 3E), whereas Grg5/AES andESG1/Tle3 expression is more pro-nounced in the anterior neural plate(Fig. 3F,G); Grg5/AES also isexpressed in the cement gland (Fig.3G,O). At late neural plate and neuraltube closure stages, all threeGrg genesare additionally detected in the neuralcrest and dorsal epidermis (Fig. 3H,I).
Grg4/Tle4 becomes weakly expressedin the cement gland (Fig. 3J) andESG1/Tle3 is additionally expressedin somites (Fig. 3H,I). All three Grggenes are expressed throughout thehead ectoderm, including the placodes(Fig. 3I). At tail bud and larval stages,all three Grg genes are expressed inthe brain and retina (Fig. 3K,L,N–R),
all placodes and cranial ganglia (Fig.3K,L,N,Q,R), neural crest and bran-chial arch mesoderm (Fig. 3K,L,O–R),and somites. All three Grg genes areadditionally expressed in the nephricmesoderm, heart, and ventral gut (Fig.3L,M,Q,R); ESG1/Tle3 is additionallydetected in the initial outgrowth of thelateral line (Fig. 3L).
Fig. 2. Expression of Eya genes. A,B: Eya1 is diffusely expressed throughout the ectoderm (ecto) of the animal (an) cap of the blastula (A) andthe gastrula (B; side views). veg, vegetal pole; bl, blastopore lip; endo, endoderm. C–E: Eya3 is detected in the animal blastomeres of the 32-cellembryo (C), the animal cap of the blastula (D), and enhanced in the dorsal ectoderm of the late gastrula (E; side views). F: At neural plate stages,Eya3 is expressed throughout the neural plate (np) and preplacodal ectoderm (PPE), but not the cement gland (cg; anterolateral view). G–O: Eya2expression. G: Eya2 is diffusely expressed throughout the neural plate and PPE (Anterior view). H: Upon neural tube closure, Eya2 is expressed inthe adeno-hypophyseal (AH), olfactory (olf), trigeminal (Vp), and dorsolateral (dlp) placodes. I: It also is detected in the caudal neural tube (nt) andsomites (so) (H, anterior view; I, dorsal view). J–L: At tail bud (J) and larval (K,L) stages, Eya2 is strongly expressed in the olfactory (olf), otic (oto),and various other placodes (pl) and their cranial ganglion derivatives (crg). Note that the lens placode (L) does not express Eya2. There is addi-tional expression in the nephric mesoderm (ne) and hypaxial muscle precursors (hyp; J, K, side views; L, anterior view). M–O: In transverse sec-tions at the levels of the diencephalon (M), otocyst (N), and caudal hindbrain (O), Eya2 expression is notable in the cranial ganglia (profundal, Pg;maxillo-mandibular branch of the trigeminal, Vg; VIIIg, IXg), otocyst, and somitic and nephric mesoderm.
EXPRESSION OF CANDIDATE SIX COFACTORS 3451
Dev
elop
men
tal D
ynam
ics
TABLE
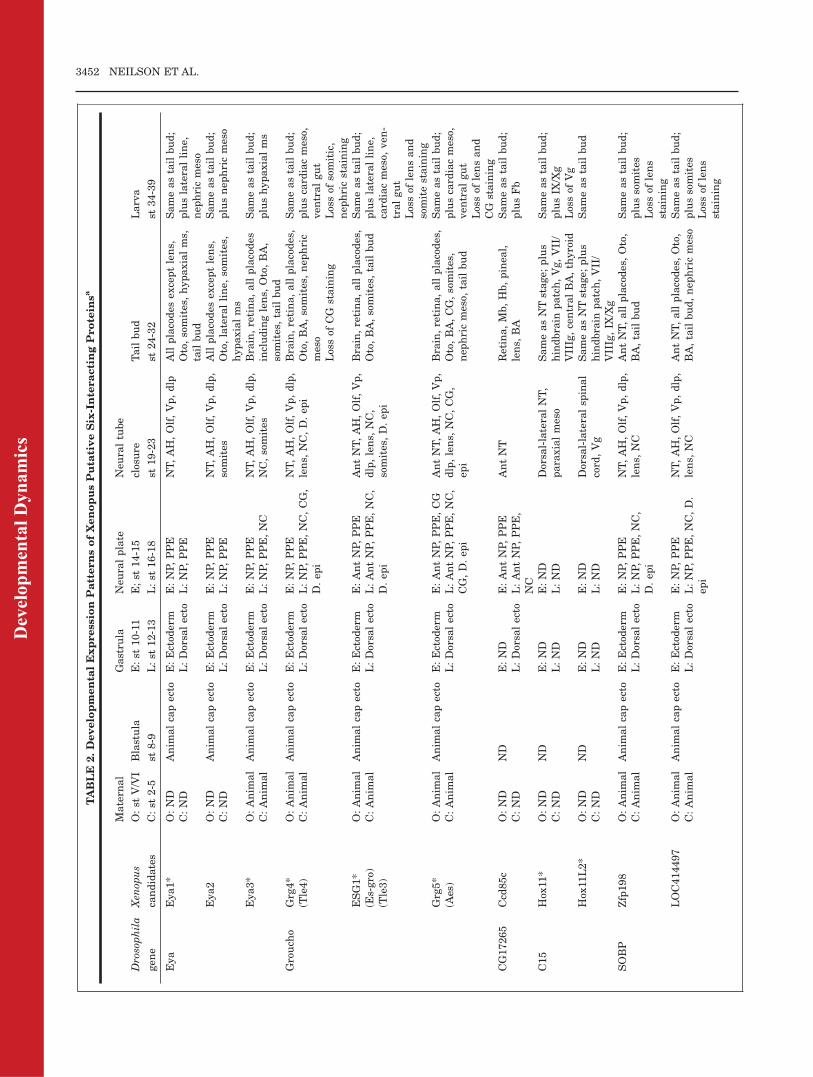
2.Developmenta
lExpression
PatternsofXenopusPuta
tiveSix-InteractingProtein
sa
Drosophila
gen
e
Xen
opus
candidates
Matern
al
O:st
V/VI
C:st
2-5
Blastula
st8-9
Gastru
la
E:st
10-11
L:st
12-13
Neu
ralplate
E;st
14-15
L:st
16-18
Neu
raltube
closure
st19-23
Tailbud
st24-32
Larva
st34-39
Eya
Eya1*
O:ND
Anim
alcapecto
E:Ectod
erm
E:NP,PPE
NT,AH,Olf,Vp,dlp
Allplacodes
exceptlens,
Oto,somites,hypaxialms,
tailbud
Sameastailbud;
pluslateralline,
nep
hricmeso
C:ND
L:Dorsa
lecto
L:NP,PPE
Eya2
O:ND
Anim
alcapecto
E:Ectod
erm
E:NP,PPE
NT,AH,Olf,Vp,dlp,
somites
Allplacodes
exceptlens,
Oto,lateralline,
somites,
hypaxialms
Sameastailbud;
plusnep
hricmeso
C:ND
L:Dorsa
lecto
L:NP,PPE
Eya3*
O:Anim
al
Anim
alcapecto
E:Ectod
erm
E:NP,PPE
NT,AH,Olf,Vp,dlp,
NC,somites
Brain,retina,allplacodes
includinglens,
Oto,BA,
somites,tailbud
Sameastailbud;
plushypaxialms
C:Anim
al
L:Dorsa
lecto
L:NP,PPE,NC
Grouch
oGrg4*
(Tle4)
O:Anim
al
C:Anim
al
Anim
alcapecto
E:Ectod
erm
L:Dorsa
lecto
E:NP,PPE
L:NP,PPE,NC,CG,
D.ep
i
NT,AH,Olf,Vp,dlp,
lens,
NC,D.ep
iBrain,retina,allplacodes,
Oto,BA,somites,nep
hric
meso
Sameastailbud;
pluscard
iacmeso,
ven
tralgut
Lossof
CG
staining
Lossof
somitic,
nep
hricstaining
ESG1*
(Es-gro)
(Tle3)
O:Anim
al
C:Anim
al
Anim
alcapecto
E:Ectod
erm
L:Dorsa
lecto
E:AntNP,PPE
L:AntNP,PPE,NC,
D.ep
i
AntNT,AH,Olf,Vp,
dlp,lens,
NC,
somites,D.ep
i
Brain,retina,allplacodes,
Oto,BA,somites,tailbud
Sameastailbud;
pluslateralline,
card
iacmeso,
ven
-tralgut
Lossof
lensand
somitestaining
Grg5*
(Aes)
O:Anim
al
C:Anim
al
Anim
alcapecto
E:Ectod
erm
L:Dorsa
lecto
E:AntNP,PPE,CG
L:AntNP,PPE,NC,
CG,D.ep
i
AntNT,AH,Olf,Vp,
dlp,lens,
NC,CG,
epi
Brain,retina,allplacodes,
Oto,BA,CG,somites,
nep
hricmeso,
tailbud
Sameastailbud;
pluscard
iacmeso,
ven
tralgut
Lossof
lensand
CG
staining
CG17265
Ccd
85c
O:ND
ND
E:ND
E:AntNP,PPE
AntNT
Retina,Mb,Hb,pinea
l,lens,
BA
Sameastailbud;
plusFb
C:ND
L:Dorsa
lecto
L:AntNP,PPE,
NC
C15
Hox
11*
O:ND
C:ND
ND
E:ND
L:ND
E:ND
L:ND
Dorsa
l-lateralNT,
paraxialmeso
SameasNTstage;
plus
hindbrain
patch,Vg,VII/
VIIIg,centralBA,thyroid
Sameastailbud;
plusIX
/Xg
Lossof
Vg
Hox
11L2*
O:ND
ND
E:ND
E:ND
Dorsa
l-lateralsp
inal
cord
,Vg
SameasNTstage;
plus
hindbrain
patch,VII/
VIIIg,IX
/Xg
Sameastailbud
C:ND
L:ND
L:ND
SOBP
Zfp198
O:Anim
al
C:Anim
al
Anim
alcapecto
E:Ectod
erm
L:Dorsa
lecto
E:NP,PPE
L:NP,PPE,NC,
D.ep
i
NT,AH,Olf,Vp,dlp,
lens,
NC
AntNT,allplacodes,Oto,
BA,tailbud
Sameastailbud;
plussomites
Lossof
lens
staining
LOC414497
O:Anim
al
C:Anim
al
Anim
alcapecto
E:Ectod
erm
L:Dorsa
lecto
E:NP,PPE
L:NP,PPE,NC,D.
epi
NT,AH,Olf,Vp,dlp,
lens,
NC
AntNT,allplacodes,Oto,
BA,tailbud,nep
hricmeso
Sameastailbud;
plussomites
Lossof
lens
staining
3452 NEILSON ET AL.
Dev
elop
men
tal D
ynam
ics
TABLE
2.(C
ontinued)
Drosophila
gen
e
Xen
opus
candidates
Matern
al
O:st
V/VI
C:st
2-5
Blastula
st8-9
Gastru
la
E:st
10-11
L:st
12-13
Neu
ralplate
E;st
14-15
L:st
16-18
Neu
raltube
closure
st19-23
Tailbud
st24-32
Larva
st34-39
XtSob
pO:ND
ND
E:ND
E:NP,PPE,D.ep
iNT,Olf,dlp
Mb,Hb,Olf,Oto,Vg,VIIg
Sameastailbud;
plusFb,IX
/Xg
C:ND
L:Dorsa
lecto
L:NP,PPE,NC,
D.ep
iCG11
35
LOC100049093
O:ND
ND
E:ND
E:NP,PPE,D.ep
iNT,AH,Olf,Vp,dlp,
lens,
NC
Brain,retina,lens,
Olf,
Vg,VIIg,IX
/XgOto,BA
Sameastailbud;
plussomites,
nep
hricmeso
C:Anim
al
L:Dorsa
lecto
L:NP,PPE,NC,
D.ep
iCG7878
Ddx43
O:ND
ND
E:ND
E:NP,PPE,D.epi
NT,dlp,lens,
NC
Brain,retina,sp
inalcord
,Olf,Oto,BA
Sameastailbud;
plussomites,
nep
hricmeso,
car-
diac,
hypaxialms.
C:ND
L:Dorsa
lecto
L:NP,PPE,NC,
D.ep
i
CKIIb
CKIIb*
O:Yes
Anim
alcapecto
E:Ectod
erm
E:NP,PPE
NT,AH,Olf,Vp,dlp,
lens,
NC
Brain,retina,sp
inalcord
,Olf,Oto,Vg,lens,
VIIg,
BA
Sameastailbud;
plusIX
/Xg,
somites,nep
hric
meso,
hypaxial
ms.
C:Anim
al
L:Dorsa
lecto
L:NP,PPE,NC
CG10576
2G4
O:Anim
al
Anim
alcapecto
E:Ectod
erm
E:NP,PPE
NT,AH,Olf,Vp,dlp,
lens,
NC,D.ep
iBrain,retina,allplacodes,
Oto,BA,nep
hricmeso,
D.
epi
Sameastailbud
C:Anim
al
L:Dorsa
lecto
L:NP,PPE,NC,
D.ep
iCG5033
MGC68939
O:Anim
al
Anim
alcapecto
E:Ectod
erm
E:NP,PPE,D.ep
iAntNT,AH,Olf,Vp,
dlp,lens,
NC
Brain,retina,allplacodes,
Oto,BA,somites,nep
hric
meso,
tailbud
Sameastailbud;
plushypaxialms,
ven
tralgut
C:Anim
al
L:Dorsa
lecto
L:AntNP,PPE,
NC,D.ep
iBop
-1O:Anim
al
Anim
alcapecto
E:Ectod
erm
E:NP,PPE,D.ep
iAntNT,AH,Olf,Vp,
dlp,lens,
NC,
somites
Brain,retina,allplacodes,
Oto,BA,somites,nep
hric
meso,
tailbud
Sameastailbud;
plushypaxialms,
ven
tralgut
C:Anim
al
L:Dorsa
lecto
L:AntNP,PPE,
NC,D.ep
iGadd45
Gadd45a
O:ND
C:ND
Anim
alcapecto
E:Ectod
erm
L:Dorsa
lecto
E:NP,PPE,CG,D.
epi
L:NP,PPE,NC,CG,
D.ep
i
CG,pinea
l,peri-cloa
-calmeso,
paraxial
meso
Hea
dandAnttrunk,par-
axialmesopatchin
tail
Sameastailbud;
plusven
tralgut
Lossof
CG
staining
Gadd45g*
O:ND
C:ND
Anim
alcapecto
E:Ectod
erm
L:Dorsa
lecto
E:primary
neu
rons,
PPE
L:primary
neu
rons,
PPE
NT,pinea
l,AH,Olf,
Vp,dlp,lens,
CG,
somites,D.ep
i
SameasNTstage;
plus
epibranch
ialplacodes
Sameastailbud;
plussomites
Lossof
lensand
Olfstaining
aAH,aden
ohypop
hysealplacode;
Ant,
anterior;BA,branch
ialarches;BZ,border
zone;
C,clea
vage;
CG,cemen
tgland;D.ep
i,dorsa
lep
idermis;dlp,dorsolateralplacode;
E,
early;ecto,ectoderm;Fb,forebrain;Hb,hindbrain;lat,
lateral;IX
/Xg,hypog
lossal/vagalganglion
;L,late;Mb,midbrain;meso,
mesod
erm;ms,
muscle;NC,neu
ralcrest;
ND,
not
detected;NP,neu
ralplate;NT,neu
raltube;
O,oocyte;Olf,olfactoryplacode;
Oto,otocyst;PPE,pre-placodalectoderm;Vg,trigem
inalganglion
;VII/VIIIg,facial/acoustico-
vestibularganglion
;Vp,trigem
inalplacode.
*Data
inpart
from
Chou
dhury
etal.,1997;David
etal.,2001;dela
Calle-Mustienes
etal.,2002;Dom
inguez
etal.,2004;Krieb
elet
al.,2007;Molen
aaret
al.,2000;Patter-
sonandKrieg
,1999;Sch
losser
andAhrens,
2004;Sch
losser,2006;Wilhelm
etal.1995,andin
part
from
ourob
servation
s.
EXPRESSION OF CANDIDATE SIX COFACTORS 3453
Dev
elop
men
tal D
ynam
ics
Groucho is a well-characterizedtranscriptional cofactor in fly thatdoes not bind to DNA directly andrequires protein–protein interactionsto assert its repressive activity
(reviewed by Courey and Jia, 2001).Yeast 2-hybrid analyses show that ver-tebrate Six proteins physically inter-act with Xenopus, zebrafish, mouse,and human Grg proteins by means of
the SD (Kobayashi et al., 2001; Zhuet al., 2002; Lopez-Rios et al., 2003).Xenopus Grg4/Tle4 is thought to befunctionally homologous to DrosophilaGroucho, whereas Grg5/AES is a
Fig. 3. Expression of Groucho-related genes (Grg). A: stage VI oocyte with Grg4 transcripts enhanced on animal (an) side (side view). B: Aneight-cell embryo with maternal ESG1 transcripts enhanced in the animal blastomeres (side view). C: In the blastula, animal cap ectodermexpresses Grg5 (side view). D: In the gastrula, the entire ectoderm expresses Grg4, whereas the preinvoluted mesoderm (meso) and endodermdo not (side view). E–G: At neural plate stages, Grg4 (E), ESG1 (F), and Grg5 (G) are expressed in the neural plate and preplacodalectoderm (PPE); Grg5 is also expressed in the cement gland (anterior views). H: ESG1 is expressed throughout the neural tube, but most stronglyin the anterior part (ant. nt). It is also detected in the neural crest (nc), dorsal epidermis (epi), and somites (dorsal view). I: At tail bud stages, ESG1expression is strong in the entire neural tube, migrating neural crest, somites, otocyst, and tail bud (tb; side view). J: At tail bud stages, Grg4 isweakly detected in the cement gland (ventral view). K: At larval stages, Grg4 is expressed throughout the brain (b), retina (r), placode derivatives(crg, L, olf, oto), and the branchial arches (BA; side view). L: ESG1 is expressed in the same tissues, and also is detected in the heart (h), ventralgut (vg), nephric mesoderm and the initial outgrowth of the lateral line (LL; side view). M–R: In transverse sections at the levels of the anterior trunk(M), forebrain (N–P) and hindbrain (Q,R), expression of the various Grg-related genes is noted in the nephric mesoderm, forebrain (fb), hindbrain(hb), retina, branchial arches, cement gland, various placodes (olf, oto, IXg, epibranchial [ebp]) and heart.
3454 NEILSON ET AL.
Dev
elop
men
tal D
ynam
ics

naturally truncated form that func-tions as a dominant-negative (Rooseet al., 1998; Molenaar et al., 2000).Xenopus Grg4/Tle4 inhibits b-catenin/Tcf-3 mediated axis duplication (Rooseet al., 1998), and enhances the tran-scriptional repressive activity ofFoxD3 required for mesoderm induc-tion (Yaklichkin et al., 2007). Mouseand human Grg4/Tle4 are expressedin brain and muscle (Koop et al.,1996); mouse and human Grg5/AESin muscle, heart, brain, and placenta(Miyasaka et al., 1993); and mouseESG1/Grg3/Tle3 in the brain, cranialganglia, olfactory and otic structures(Dehni et al., 1995; Leon and Lobe,1997). To date there is no informationregarding the functional role of verte-brate Grg cofactors in craniofacial de-velopment, but coexpression of Dro-sophila Groucho with Xenopus Six1causes repression of cranial neuralcrest and epidermal genes (Brugmannet al., 2004). The expression of Xeno-pus Grg4/Tle4, Grg5/AES, andESG1/Tle3 overlaps extensively withSix1 and Six2 in PPE and placodes;they also are expressed in cranial neu-ral crest and neural tube. Thus, thesethree Grg genes are likely to haveroles in craniofacial development.
CG17265 (ccd85c)-related genes.
We identified two putative Xenopusorthologues: Xenopus tropicalis (Xt)ccd85c and Xenopus laevis (Xl) ccd85c(Table 1). Because Xt-ccd85c showedthe highest amino acid sequence iden-tity to CG17265 over its entire codingregion, we used the Xt-ccd85c probeto study expression patterns in X. lae-vis embryos; it should be noted thatmany ISH probes can be interchangedbetween the two Xenopus species.Subsequent studies with the Xl-ccd85c probe gave identical results(data not shown). Staining is notdetected before stage 12 (Table 2).Throughout neural plate stages, theXt-ccd85c probe diffusely stains theanterior neural plate and the PPE(Fig. 4A). At neural tube closurestages, diffuse expression is detectedin the anterior neural tube, retina,otocyst, dorsal epidermis, and bran-chial arches (Fig. 4B). At tail budstages, the pineal, midbrain, andhindbrain are strongly stained andthe retina, lens, otocyst, and bran-
chial arches are weakly stained (Fig.4C). By larval stages, forebrain stain-ing intensifies (Fig. 4D,E); in someembryos expression extends into thespinal cord. Lens and retina expres-sion is maintained, but branchial archand otocyst expression is barelydetected (Fig. 4E,F). Unlike othercandidate cofactor genes, there is nodetectable expression of Xt-ccd85c innephric or somitic mesoderm (Fig.4G).
Drosophila CG17265 encodes a nu-clear protein that is thought to beinvolved in RNA interference (Dorneret al., 2006). There are no functionalstudies of this gene in Xenopus or anyother vertebrate to date. However, arelated protein, Ccd85b, was identi-fied as a binding partner of humanSix1 in the human protein–proteininteraction network (Rual et al.,2005). Xenopus ccd85c expressiononly partially overlaps those of Six1and Six2 (PPE, lens, otocyst), suggest-ing that it may have a restricted rolein craniofacial development.
C15-related genes.
C15 is assigned to Xenopus UnigeneXl.679 and shares 57.1% amino acididentity with Xenopus Hox11L2, theXenopus homolog of the mammalianT-cell leukemia homeobox 3 (Tlx3)proteins (Table 1). The expressionpatterns of Hox11L2 and the relatedgene, Hox11 (mammalian Tlx1) werepreviously described in detail (Patter-son and Krieg, 1999; summarized inTable 2). Their expression patternsare similar to those reported in mouseand chick embryos (Raju et al., 1993;Roberts et al., 1994, 1995; Loganet al., 1998; Uchiyama et al., 1999),although the prominent Hox11expression in mouse spleen was notseen in Xenopus (Patterson andKrieg, 1999). Misexpression ofHox11L2 or Hox11 is associated withsome forms of human T-cell leukemia(Hatano et al., 1991; Lu et al., 1991;Bernard et al., 2001; Ferrando et al.,2002), and their knock-down in miceresults in abnormal development ofthe spleen, colon and the ventral med-ullary respiratory center (Robertset al., 1994; Dear et al., 1995; Hatanoet al., 1997; Shirasawa et al., 2000);these studies do not report craniofa-cial defects. The expression of these
genes overlaps only minimally withSix1/Six2; however, because they areexpressed in cranial sensory gangliaand branchial arches, Hox11 andHox11L2 may have late roles in cra-niofacial development.
Sobp-related genes.
We identified two Sobp-related Xeno-pus laevis (Xl) genes (Zfp198,LOC414497) with homology in theregions designated as box2 and box3in Drosophila SOBP (Kenyon et al.,2005), and one Xenopus tropicalisgene (Xt-Sobp) with homology inboxes 1–3. Maternal mRNAs for bothXl-Sobp-related genes are detected inthe animal blastomeres of cleavageembryos (Fig. 5A; Table 2). They areexpressed in the animal cap ectodermof the blastula (Table 2), throughoutthe embryonic ectoderm at early gas-trula stages (Fig. 5B) and begin to bedorsally concentrated in the ectodermat late gastrula stages (Table 2). Incontrast, maternal Xt-Sobp mRNA isnot detected, nor is Xt-Sobp expressiondetected at blastula or early gastrulastages (Table 2). All three Sobp-relatedgenes are expressed in the neuralplate, PPE and dorsal epidermis atneural plate stages (Fig. 5C; Table 2).At neural tube closure, the two Xl-Sobp-related genes are detectedthroughout the neural tube, in all plac-odes and the neural crest (Fig. 5D; Ta-ble 2), whereas Xt-Sobp is only weaklyexpressed in the neural tube and in theolfactory and dorsolateral placodes (Ta-ble 2). Epidermal staining of all threegenes is lost at this stage. At tail budand larval stages, the two Xl-Sobp-related genes are strongly expressed inbrain and cervical spinal regions (Fig.5E,F,I,J,K), whereas the Xt-Sobp geneis expressed only in lateral patches inthe midbrain and hindbrain (Fig.5G,L). All three genes are expressed invarious placodes and placode deriva-tives, but the Xt-Sobp gene is morerestricted (Fig. 5G,L). The two Xl-Sobp-related genes are additionallyexpressed in the branchial arches (Fig.5E,F,I,J). At late larval stages, the twoXl-Sobp-related genes no longer stainthe lens (Table 2), and begin to beexpressed in the somites (Fig. 5K). Atlate larval stages, Xt-Sobp expressionexpands into the forebrain (Fig. 5H).
EXPRESSION OF CANDIDATE SIX COFACTORS 3455
Dev
elop
men
tal D
ynam
ics
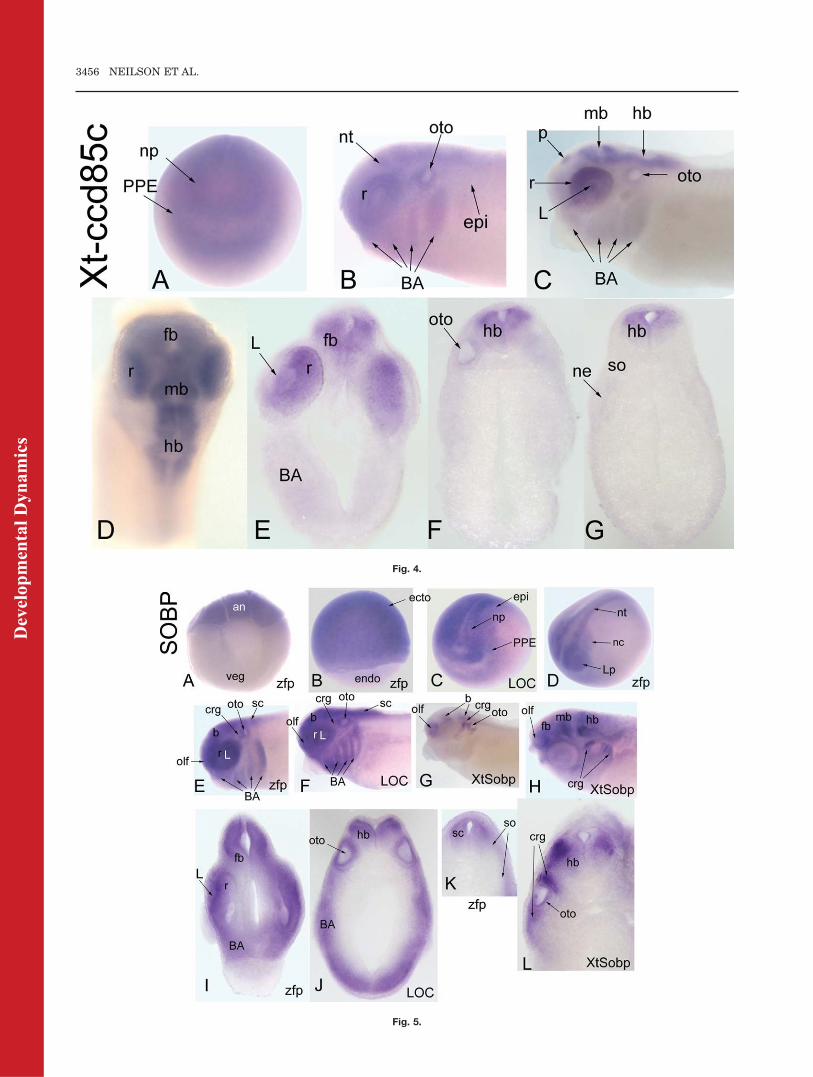
Fig. 4.
Fig. 5.
3456 NEILSON ET AL.
Dev
elop
men
tal D
ynam
ics

Drosophila SOBP is a known bind-ing partner of SO and is expressed inthe differentiating eye field (Kenyonet al., 2005). In mouse, Sobp isexpressed in the otocyst, retina, olfac-tory epithelium, trigeminal ganglion,and hair follicles, and the Jackson cir-cler mouse, which is deaf, carries a re-cessive mutation in Sobp (Chen et al.,2008). The expression of the XenopusSobp genes partially overlap with Sixgenes (PPE, placodes), and their addi-tional expression in the cranial neuralcrest, branchial arches, and anteriorneural tube suggest they will haveroles in craniofacial development.
CG1135-related gene.
Maternal LOC100049093 mRNA isweakly detected in animal blasto-meres of cleavage embryos but is notdetected by ISH at blastula or earlygastrula stages (Table 2). By late gas-trula and early neural plate stages, itis expressed throughout the dorsal ep-idermis, with enhancement in theneural plate and PPE (Table 2; Fig.6A). At neural tube closure, it isexpressed throughout the neuraltube, and diffusely in the PPE, neuralcrest and epidermis (Fig. 6B). At tailbud and larval stages, it is stronglyexpressed throughout the brain andretina in all embryos (Fig. 6C), andweakly expressed in the spinal cord ina few embryos (Fig. 6F). In addition,it is strongly expressed in placodes,cranial ganglia, and branchial arches(Fig. 6C–E), and weakly expressed in
nephric mesoderm (Fig. 6C,F). Bylarval stages, it is additionallyexpressed throughout the head meso-derm and anterior somites (Table 2).
In Drosophila, CG1135 contains aForkhead-associated domain and wasidentified as a component of the Hhsignal pathway in a large-scale RNAinterference screen (Nybakken et al.,2005). Its molecular function isunknown, but it is annotated ashaving a role in centriole replication.The Xenopus orthologue was identi-fied as a component of the origin ofreplication (Carpenter and Dunphy,1998). The human orthologue micro-spherule protein-1: (1) is part of a cen-trosomal complex that is essential forcell viability (Hirohashi et al., 2006),(2) modulates Daxx-dependent tran-scriptional repression (Lin andShih, 2002), and (3) interacts with thefragile X mental retardation proteinin polyribosomal mRNPS fromneurons (Davidovic et al., 2006).LOC100049093 expression overlapswith that of Six1/Six2 (PPE, placodes,somites, nephric mesoderm) and alsois expressed in the cranial neuralcrest, branchial arches, and neuraltube, suggesting a role in craniofacialdevelopment.
CG7878-related gene.
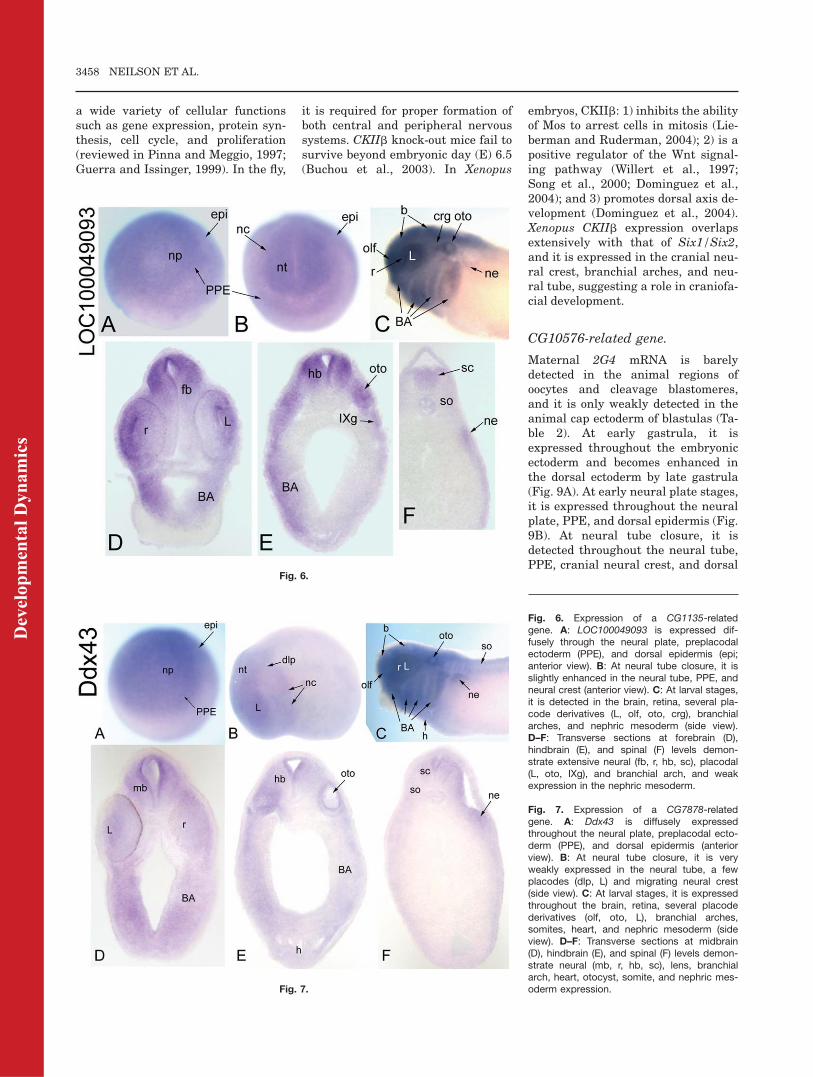
There are two entries in GenBank forXenopus tropicalis orthologues (Xt-Ddx43) of Drosophila CG7878 (Table1), but their sequences are identical.Maternal mRNA for Xt-Ddx43 is not
detected, nor is expression detected atblastula or early gastrula stages (Ta-ble 2). At neural plate stages, it isexpressed diffusely through the neu-ral plate, PPE, and dorsal epidermis(Fig. 7A). At neural tube closure, it isweakly expressed throughout theneural tube, the lens, and dorsolat-eral placodes and migrating neuralcrest (Fig. 7B). At tail bud and larvalstages, it is expressed throughout thebrain and retina and eventuallyextends into the rostral spinal cord(Fig. 7C–F; Table 2). In addition,there is staining in the olfactory pit,otocyst, and branchial arches (Fig.7C–E; Table 2). By larval stages,there is additional diffuse staining inthe head mesenchyme, somites, heart,and nephric mesoderm (Fig. 7C–F).Drosophila CG7878 is predicted to
function as an RNA helicase. Thehuman orthologue HAGE is expressedat high levels in a variety of cancersand in normal adult testes (Marte-lange et al., 2000; Mathieu et al.,2010). Xt-Ddx43 expression overlapswith that of Six1/Six2 (PPE, placo-des, somites, nephric mesoderm) andis expressed in the cranial neuralcrest, branchial arches, and neuraltube, suggesting a role in craniofacialdevelopment.
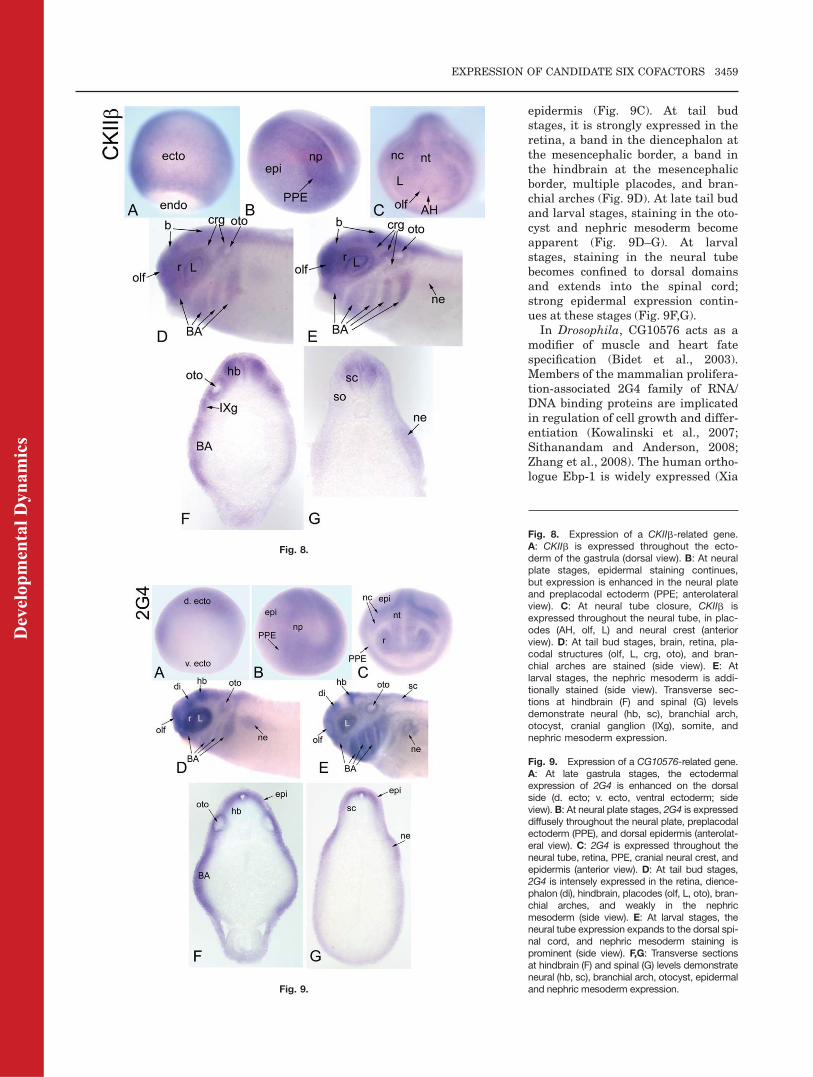
CKIIb-related gene.
We identified one Xenopus laevis generelated to Drosophila CKIIb. Previousstudies demonstrate that XenopusCKIIb is expressed maternally (Wil-helm et al., 1995) and in the animalhemispheres at cleavage and blastulastages (Dominguez et al., 2004; Table2). We detected its mRNA throughoutthe gastrula ectoderm (Fig. 8A), witha slight enhancement on the dorsalside. At neural plate stages, it isexpressed in the dorsal epidermis,neural plate, and PPE (Fig. 8B). Atneural tube closure, it is expressedthroughout the neural tube, in allplacodes and the neural crest (Fig.8C). By tail bud stages, the branchialarches are additionally stained (Fig.8D). By larval stages, there is addi-tional expression in the somites andnephric mesoderm (Fig. 8E–G), andhypaxial muscle precursors (Table 2).CKIIb is the regulatory subunit of
the pleiotropic and highly conservedprotein kinase CK2 that is involved in
Fig. 4. Expression of a CG17265-related gene, Xt-ccd85c. A: Diffuse expression of Xt-ccd85cthroughout the anterior neural plate and PPE (anterior view). B: At neural tube closure, there isdiffuse expression throughout the neural tube, retina, dorsal epidermis, otocyst, and branchialarches (side view). C: At late tail bud stages, the pineal (p), retina, midbrain, and hindbrain arestained. There also is weak staining in the lens, otocyst, and branchial arches (side view).D: Dorsal view at larval stage showing expression extending into the forebrain. E–G: Transversesections at forebrain (E), hindbrain (F), and caudal hindbrain (G) demonstrate restricted neuralexpression.
Fig. 5. Expression of SOBP-related genes. A: zfp198 mRNA is concentrated in the animal blas-tomeres at the eight-cell stage (side view). B: zfp198 is expressed throughout the ectoderm ofthe gastrula (side view). C: At early neural plate, LOC414497 is expressed throughout the neuralplate, preplacodal ectoderm (PPE), and dorsal epidermis (anterolateral view). D: At neural tubeclosure, neural crest and lens placode (Lp) expression of zfp198 becomes apparent (anterolat-eral view). E–G: At late tail bud, zfp198 (E) and LOC414497 (F) are expressed throughout thebrain, retina, placode derivatives (olf, L, crg, oto) and branchial arches, whereas Xt-Sobp (G) isexpressed in patches in the brain and in several placode derivatives (olf, crg, oto; side view).H: At larval stages, Xt-Sobp expression extends into the forebrain (side view). I–K: Transversesections at forebrain (I), hindbrain (J), and spinal (K) levels demonstrate expression of zfg198and LOC414497 in neural (fb, r, hb, sc), branchial arch, otocyst and somites. L: Transverse sec-tion at hindbrain demonstrates Xt-Sobp more restricted expression in lateral hindbrain, otocyst,and cranial ganglia.
EXPRESSION OF CANDIDATE SIX COFACTORS 3457
Dev
elop
men
tal D
ynam
ics
a wide variety of cellular functionssuch as gene expression, protein syn-thesis, cell cycle, and proliferation(reviewed in Pinna and Meggio, 1997;Guerra and Issinger, 1999). In the fly,
it is required for proper formation ofboth central and peripheral nervoussystems. CKIIb knock-out mice fail tosurvive beyond embryonic day (E) 6.5(Buchou et al., 2003). In Xenopus
embryos, CKIIb: 1) inhibits the abilityof Mos to arrest cells in mitosis (Lie-berman and Ruderman, 2004); 2) is apositive regulator of the Wnt signal-ing pathway (Willert et al., 1997;Song et al., 2000; Dominguez et al.,2004); and 3) promotes dorsal axis de-velopment (Dominguez et al., 2004).Xenopus CKIIb expression overlapsextensively with that of Six1/Six2,and it is expressed in the cranial neu-ral crest, branchial arches, and neu-ral tube, suggesting a role in craniofa-cial development.
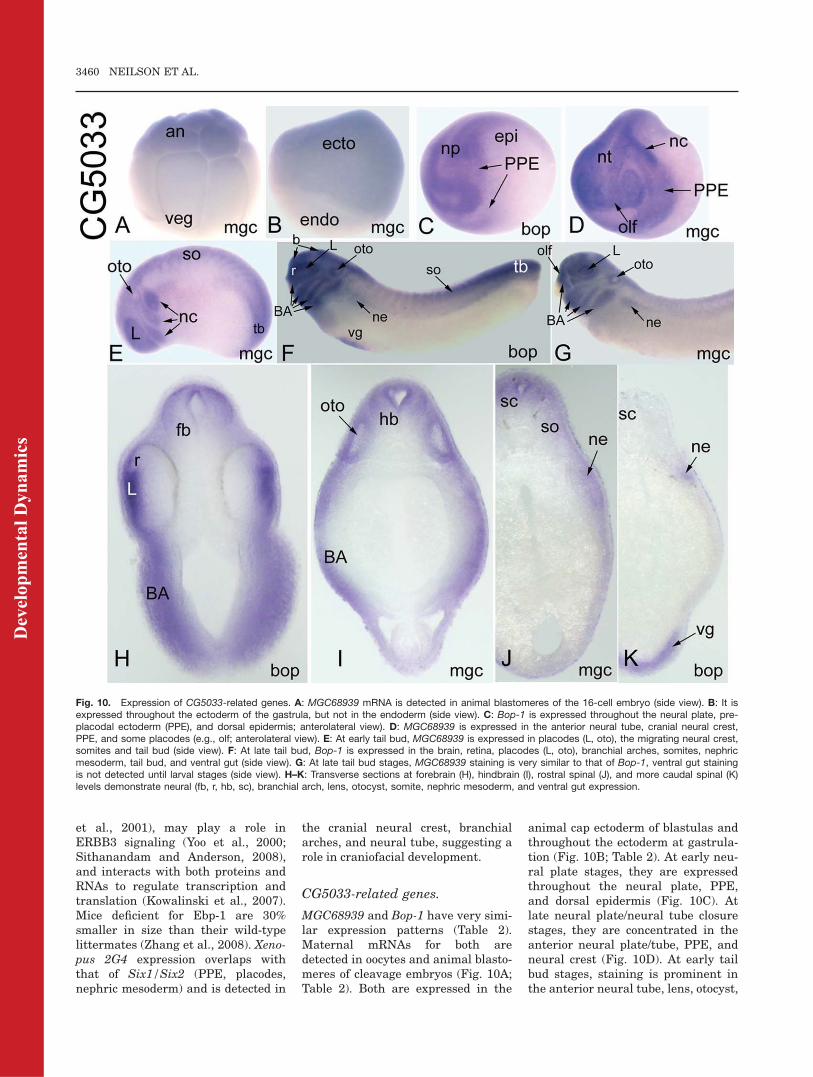
CG10576-related gene.
Maternal 2G4 mRNA is barelydetected in the animal regions ofoocytes and cleavage blastomeres,and it is only weakly detected in theanimal cap ectoderm of blastulas (Ta-ble 2). At early gastrula, it isexpressed throughout the embryonicectoderm and becomes enhanced inthe dorsal ectoderm by late gastrula(Fig. 9A). At early neural plate stages,it is expressed throughout the neuralplate, PPE, and dorsal epidermis (Fig.9B). At neural tube closure, it isdetected throughout the neural tube,PPE, cranial neural crest, and dorsal
Fig. 6.
Fig. 7.
Fig. 6. Expression of a CG1135-relatedgene. A: LOC100049093 is expressed dif-fusely through the neural plate, preplacodalectoderm (PPE), and dorsal epidermis (epi;anterior view). B: At neural tube closure, it isslightly enhanced in the neural tube, PPE, andneural crest (anterior view). C: At larval stages,it is detected in the brain, retina, several pla-code derivatives (L, olf, oto, crg), branchialarches, and nephric mesoderm (side view).D–F: Transverse sections at forebrain (D),hindbrain (E), and spinal (F) levels demon-strate extensive neural (fb, r, hb, sc), placodal(L, oto, IXg), and branchial arch, and weakexpression in the nephric mesoderm.
Fig. 7. Expression of a CG7878-relatedgene. A: Ddx43 is diffusely expressedthroughout the neural plate, preplacodal ecto-derm (PPE), and dorsal epidermis (anteriorview). B: At neural tube closure, it is veryweakly expressed in the neural tube, a fewplacodes (dlp, L) and migrating neural crest(side view). C: At larval stages, it is expressedthroughout the brain, retina, several placodederivatives (olf, oto, L), branchial arches,somites, heart, and nephric mesoderm (sideview). D–F: Transverse sections at midbrain(D), hindbrain (E), and spinal (F) levels demon-strate neural (mb, r, hb, sc), lens, branchialarch, heart, otocyst, somite, and nephric mes-oderm expression.
3458 NEILSON ET AL.
Dev
elop
men
tal D
ynam
ics
epidermis (Fig. 9C). At tail budstages, it is strongly expressed in theretina, a band in the diencephalon atthe mesencephalic border, a band inthe hindbrain at the mesencephalicborder, multiple placodes, and bran-chial arches (Fig. 9D). At late tail budand larval stages, staining in the oto-cyst and nephric mesoderm becomeapparent (Fig. 9D–G). At larvalstages, staining in the neural tubebecomes confined to dorsal domainsand extends into the spinal cord;strong epidermal expression contin-ues at these stages (Fig. 9F,G).In Drosophila, CG10576 acts as a
modifier of muscle and heart fatespecification (Bidet et al., 2003).Members of the mammalian prolifera-tion-associated 2G4 family of RNA/DNA binding proteins are implicatedin regulation of cell growth and differ-entiation (Kowalinski et al., 2007;Sithanandam and Anderson, 2008;Zhang et al., 2008). The human ortho-logue Ebp-1 is widely expressed (Xia
Fig. 8.
Fig. 9.
Fig. 8. Expression of a CKIIb-related gene.A: CKIIb is expressed throughout the ecto-derm of the gastrula (dorsal view). B: At neuralplate stages, epidermal staining continues,but expression is enhanced in the neural plateand preplacodal ectoderm (PPE; anterolateralview). C: At neural tube closure, CKIIb isexpressed throughout the neural tube, in plac-odes (AH, olf, L) and neural crest (anteriorview). D: At tail bud stages, brain, retina, pla-codal structures (olf, L, crg, oto), and bran-chial arches are stained (side view). E: Atlarval stages, the nephric mesoderm is addi-tionally stained (side view). Transverse sec-tions at hindbrain (F) and spinal (G) levelsdemonstrate neural (hb, sc), branchial arch,otocyst, cranial ganglion (IXg), somite, andnephric mesoderm expression.
Fig. 9. Expression of a CG10576-related gene.A: At late gastrula stages, the ectodermalexpression of 2G4 is enhanced on the dorsalside (d. ecto; v. ecto, ventral ectoderm; sideview). B: At neural plate stages, 2G4 is expresseddiffusely throughout the neural plate, preplacodalectoderm (PPE), and dorsal epidermis (anterolat-eral view). C: 2G4 is expressed throughout theneural tube, retina, PPE, cranial neural crest, andepidermis (anterior view). D: At tail bud stages,2G4 is intensely expressed in the retina, dience-phalon (di), hindbrain, placodes (olf, L, oto), bran-chial arches, and weakly in the nephricmesoderm (side view). E: At larval stages, theneural tube expression expands to the dorsal spi-nal cord, and nephric mesoderm staining isprominent (side view). F,G: Transverse sectionsat hindbrain (F) and spinal (G) levels demonstrateneural (hb, sc), branchial arch, otocyst, epidermaland nephric mesoderm expression.
EXPRESSION OF CANDIDATE SIX COFACTORS 3459
Dev
elop
men
tal D
ynam
ics
et al., 2001), may play a role inERBB3 signaling (Yoo et al., 2000;Sithanandam and Anderson, 2008),and interacts with both proteins andRNAs to regulate transcription andtranslation (Kowalinski et al., 2007).Mice deficient for Ebp-1 are 30%smaller in size than their wild-typelittermates (Zhang et al., 2008). Xeno-pus 2G4 expression overlaps withthat of Six1/Six2 (PPE, placodes,nephric mesoderm) and is detected in
the cranial neural crest, branchialarches, and neural tube, suggesting arole in craniofacial development.
CG5033-related genes.
MGC68939 and Bop-1 have very simi-lar expression patterns (Table 2).Maternal mRNAs for both aredetected in oocytes and animal blasto-meres of cleavage embryos (Fig. 10A;Table 2). Both are expressed in the
animal cap ectoderm of blastulas andthroughout the ectoderm at gastrula-tion (Fig. 10B; Table 2). At early neu-ral plate stages, they are expressedthroughout the neural plate, PPE,and dorsal epidermis (Fig. 10C). Atlate neural plate/neural tube closurestages, they are concentrated in theanterior neural plate/tube, PPE, andneural crest (Fig. 10D). At early tailbud stages, staining is prominent inthe anterior neural tube, lens, otocyst,
Fig. 10. Expression of CG5033-related genes. A: MGC68939 mRNA is detected in animal blastomeres of the 16-cell embryo (side view). B: It isexpressed throughout the ectoderm of the gastrula, but not in the endoderm (side view). C: Bop-1 is expressed throughout the neural plate, pre-placodal ectoderm (PPE), and dorsal epidermis; anterolateral view). D: MGC68939 is expressed in the anterior neural tube, cranial neural crest,PPE, and some placodes (e.g., olf; anterolateral view). E: At early tail bud, MGC68939 is expressed in placodes (L, oto), the migrating neural crest,somites and tail bud (side view). F: At late tail bud, Bop-1 is expressed in the brain, retina, placodes (L, oto), branchial arches, somites, nephricmesoderm, tail bud, and ventral gut (side view). G: At late tail bud stages, MGC68939 staining is very similar to that of Bop-1, ventral gut stainingis not detected until larval stages (side view). H–K: Transverse sections at forebrain (H), hindbrain (I), rostral spinal (J), and more caudal spinal (K)levels demonstrate neural (fb, r, hb, sc), branchial arch, lens, otocyst, somite, nephric mesoderm, and ventral gut expression.
3460 NEILSON ET AL.
Dev
elop
men
tal D
ynam
ics
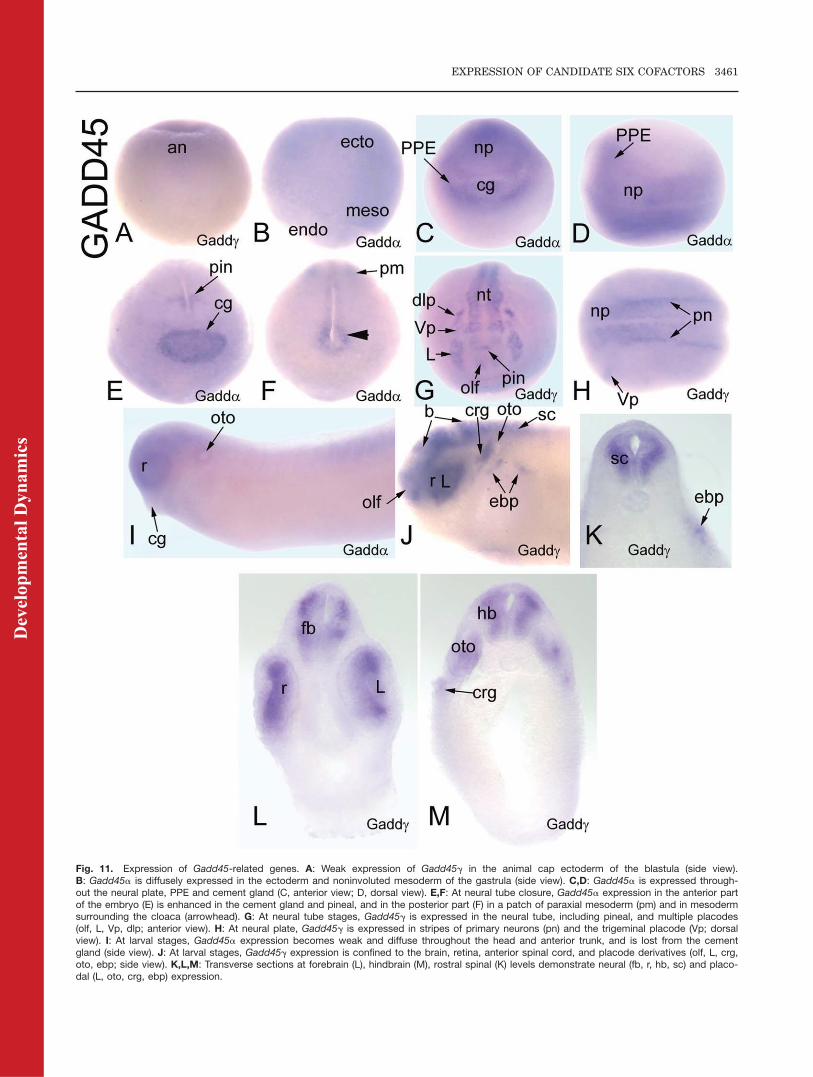
Fig. 11. Expression of Gadd45-related genes. A: Weak expression of Gadd45g in the animal cap ectoderm of the blastula (side view).B: Gadd45a is diffusely expressed in the ectoderm and noninvoluted mesoderm of the gastrula (side view). C,D: Gadd45a is expressed through-out the neural plate, PPE and cement gland (C, anterior view; D, dorsal view). E,F: At neural tube closure, Gadd45a expression in the anterior partof the embryo (E) is enhanced in the cement gland and pineal, and in the posterior part (F) in a patch of paraxial mesoderm (pm) and in mesodermsurrounding the cloaca (arrowhead). G: At neural tube stages, Gadd45g is expressed in the neural tube, including pineal, and multiple placodes(olf, L, Vp, dlp; anterior view). H: At neural plate, Gadd45g is expressed in stripes of primary neurons (pn) and the trigeminal placode (Vp; dorsalview). I: At larval stages, Gadd45a expression becomes weak and diffuse throughout the head and anterior trunk, and is lost from the cementgland (side view). J: At larval stages, Gadd45g expression is confined to the brain, retina, anterior spinal cord, and placode derivatives (olf, L, crg,oto, ebp; side view). K,L,M: Transverse sections at forebrain (L), hindbrain (M), rostral spinal (K) levels demonstrate neural (fb, r, hb, sc) and placo-dal (L, oto, crg, ebp) expression.
EXPRESSION OF CANDIDATE SIX COFACTORS 3461
Dev
elop
men
tal D
ynam
ics

migrating neural crest, somites, andtail bud (Fig. 10E). At late tail budstages, there is prominent staining inthe brain, retina, placodes, branchialarches, somites, nephric mesoderm,tail bud, and ventral gut (Fig. 10F–K). By larval stages, there is addi-tional staining in the hypaxial muscleprecursors (Table 2).
Drosophila CG5033 is predicted tofunction in ribonucleoprotein binding.Vertebrate Bop-1 was first identified asa mouse gastrula cDNA clone thatcaused morphological changes in thedeveloping Xenopus embryo in a high-through put screen (Chiao et al., 2005).Mouse Bop-1 is implicated in rRNAprocessing and ribosome biogenesisand plays a role in cell cycle progres-sion (Strezoska et al., 2000, 2002; Rohr-moser et al., 2007); deregulation ofhuman Bop-1 is associated with colo-rectal tumors (Killian et al., 2006).Xenopus CG5033-related genes areexpressed in the same tissues as Six1/Six2 (PPE, placodes, somites, nephricmesoderm), and also in the cranial neu-ral crest, branchial arches, and neuraltube, suggesting that they may haveroles in craniofacial development.
Gadd45-related genes.
Xenopus Gadd45a and Gadd45g havesimilar early expression patterns thatdiverge at neural plate stages (Table2). Maternal mRNAs for Gadd45a orGadd45g are not detected. Both areweakly expressed in the animal capectoderm of the blastula (Fig. 11A; Ta-ble 2), throughout the embryonic ecto-derm at early gastrula (Fig. 11B; Ta-ble 2) and are dorsally concentratedat late gastrulation (Table 2). At neu-ral plate stages, Gadd45a is stronglyexpressed throughout the neuralplate, PPE, and cement gland (Fig.11C,D). At neural tube closure,Gadd45a is strongly expressed in thecement gland, pineal, peri-cloacalmesoderm, and a patch of paraxialmesoderm in the posterior trunk (Fig.11E,F). In contrast, at neural platestages, Gadd45g is expressed instripes in the neural ectoderm thatcorrespond to the primary neuronsand in the trigeminal placode (Fig.11H; de la Calle-Mustienes et al.,2002), and at neural tube stages itstains discrete patches in the neuraltube including pineal, all of the placo-
des (Fig. 11G), and scattered cells inthe epidermis and somites (Table 2).At tail bud stages, Gadd45a stainingis weak and diffuse throughout thehead and anterior trunk and thecement gland staining is lost (Fig.11I); at larval stages, there is addi-tional staining in the ventral gut (Ta-ble 2). In contrast, at tail bud andlarval stages Gadd45g is stronglyexpressed in the brain, retina, ante-rior spinal cord, and several placodederivatives (Fig. 11J–M); expressionis detected in somites at late larvalstages (Table 2).
In Drosophila, Gadd45 is involvedin JNK signaling during very earlydevelopmental processes (Peretzet al., 2007) and in wound healing(Stramer et al., 2008). In humans andmouse, three Gadd45 proteins (a, b,g) act to inhibit the cell cycle and pro-mote apoptosis (Takekawa and Saito,1998); Drosophila Gadd45 shares thehighest amino acid identity with ver-tebrate Gadd45g (Peretz et al., 2007).Loss of murine Gadd45a results inphenotypes similar to p53-null mice(Hollander et al., 1999); loss of murineGadd45b interferes with activity-induced proliferation of neuronal pro-genitors (Ma et al., 2009); loss of mu-rine Gadd45g may affect the develop-ment of blood lineages (Lu et al.,2001). In Xenopus embryos, Gadd45greduces the number of dividing cellsin the neural plate, thus promotingneuronal differentiation (de la Calle-Mustienes et al., 2002). The Xenopustropicalis genome database annotatesa Gadd45b gene (Table 1), but nocDNA clones are currently available.However, in zebrafish, Gadd45b isinvolved in somite segmentation(Kawahara et al., 2005) and mouseGadd45b is required for chondrocyteterminal differentiation (Ijiri et al.,2005). The Xenopus Gadd45-relatedgenes are expressed in some of thesame tissues as Six1/Six2 (PPE, plac-odes, somites), and also in the neuraltube, suggesting that they may haveroles in craniofacial development.
CONCLUSIONS
This analysis identified 33 Xenopusgenes as putative orthologues of 20 ofthe 25 Drosophila proteins that inter-act with SO. Of these 33, we found
that 3 are not expressed in the sametissues as Six1/Six2 (ESG2, CG33090,VhaAC39), 3 are ubiquitously ex-pressed (RhoGEF2, Chc, RpS27) and 7are not currently available for ISHstudy (Eya4, CG9083, CG1218, 2 puta-tive CG5665 orthologues, RhoGAP93B,Gadd45b). However, the remainingXenopus genes, representing 11 of theDrosophila SO-interacting genes, areexpressed in craniofacial tissues, over-lapping at least partially with theexpression domains of Six1/Six2. It isnotable that the Eya, Grg, Sobp,CG1135, CG7878/Ddx43, CG10576/2G4, CG5033, and Gadd45g genesmostly closely overlapped with Six1/Six2 expression patterns in the embry-onic ectoderm, PPE, placodes, somites,and nephric mesoderm. However, theyall displayed additional expressiondomains, indicating that they likelyinteract with other transcription fac-tors, including perhaps other Six fam-ily members. Also, with the exceptionof the Eya genes, their expression inthe dorsal epidermis and PPE domainswas much more diffuse that those ofSix1/Six2. It also it notable that theexpression domains of CG17265/ccd85c, C15, and Gadd45a overlap theleast with Six1/Six2, suggesting thatthey may have more restricted func-tions in craniofacial development.Identification of additional proteins
that may interact with Six proteins ascofactors is important to fully under-stand the function of this family oftranscription factors. This is particu-larly relevant because the penetranceof BO and BOR syndromes is vari-able, and results from Six1 heterozy-gous mice suggest that there aremodifier genes that influence Six1 ac-tivity or function to modulate the mu-tant phenotype (Xu et al., 2003; Rufet al., 2004). Because mutations inSix and Eya genes only account forapproximately half of the BO andBOR patients, other genes are likelyto be involved. Our data demonstratethat a large number of genes identi-fied by sequence homology to putativepartners of fly SO are expressed invertebrate craniofacial tissues duringdevelopment. We speculate that thesemay prove to encode for new Six-interacting proteins with importantroles in vertebrate developmentalprocesses, particularly in craniofacialdevelopment.
3462 NEILSON ET AL.
Dev
elop
men
tal D
ynam
ics

EXPERIMENTAL
PROCEDURES
Bioinformatics
Putative Xenopus laevis orthologuesof the Drosophila genes encoding SO-interacting proteins were identified inBLAST searches using a translatednucleotide Drosophila query to searchthe translated nucleotide Xenopuslaevis database in the NCBI Gen-Bank. If no Xenopus laevis gene wasidentified, the BLAST search wasrepeated using the Xenopus tropicalistranslated nucleotide database. IfXenopus orthologues were not identi-fied in either of these searches, theEnsembl database was searched todetermine if Drosophila-related genesare annotated in the Xenopus tropica-lis genome. Amino acid sequenceidentity, alignment scores and E val-ues were all considered to determinethe homology between the Drosophilaand Xenopus genes. If a Xenopus lae-vis gene was identified for the candi-date SO-interacting proteins, the Xen-opus tropicalis orthologue was notlisted in Table 1. The exceptions areCG17265 (ccdc85c) and SOBP; inthese cases the X. tropicalis sequenceshowed a higher amino acid sequenceidentity to the fly orthologues overthe entire coding region than did theX. laevis sequence.
Synthesis of Antisense
RNA Probes
Some plasmids were obtained from theresearch community (Patterson andKreig, 1999; David et al., 2001; de laCalle-Mustienes et al., 2002; Burkset al., 2009). The other plasmids werepurchased from Open BioSystems. IfXenopus laevis cDNAs were availablethey were used for synthesizing probes.If not, Xenopus tropicalis cDNAs wereused and these are specifically identi-fied in the text. Digoxigenin-labeledantisense RNA probes were synthe-sized as per the manufacturer’sinstructions (Ambion Megascript kit).
Whole-Mount ISH
Xenopus laevis oocytes were surgi-cally removed from gravid females(Sive et al., 2000). Wild-type andalbino Xenopus laevis embryos wereobtained by natural mating of adult
pairs as previously described (Moody,2000). Embryos were staged accord-ing to Nieuwkoop and Faber (1994).Specimens were fixed and processedfor whole-mount ISH according tostandard procedures (Sive et al.,2000). To visualize internal expres-sion, embryos already processed forwhole-mount ISH were embedded in4% agarose and sectioned at 100 mmwith a vibratome.
ACKNOWLEDGMENTSWe thank Gerhard Schlosser (Eya1),Jose Gomez-Skarmeta (Gadd45g), IraDaar (Grg5), Paul Krieg (Hox11L2),and Betsy Pownall (Grg4) for provid-ing plasmids. We also thank LynneMied and Pallavi Mhaske for technicalassistance. K.M.N., S.A.M., and F.P.were funded by the NIH.
REFERENCES
Abdelhak S, Kalatzis V, Heilig R, CompainS, Samson D, Vincent C, Weil D, CruaudC, Sahly I, Leibovici M, Bitner-GlindziczM, Francis M, Lacombe D, Vigneron J,Charachon R, Boven K, Bedbeder P, VanRegemorter N, Weissenbach J, Petit C.1997. A human homologue of the Dro-sophila eyes absent gene underlies bran-chio-oto-renal (BOR) syndrome andidentifies a novel gene family. Nat Genet15:157–164.
Abe Y, Oka A, Mizuguchi M, Igarashi T,Ishikawa S, Aburatani H, Yokoyama S,Asahara H, Nagao K, Yamada M, Miya-shita T. 2009. Eya4, deleted in a casewith middle interhemispheric variant ofHoloprosencephaly interacts with Six3both physically and functionally. HumMutat 30:946–955.
Bai J, Montell D. 2002. Eyes absent, akey repressor of polar cell fate duringDrosophila oogenesis. Development 129:5377–5388.
Baker CV, Bronner-Fraser M. 2001. Verte-brate cranial placodes. I. Embryonicinduction. Dev Biol 232:1–61.
Bernard OA, Bisson-LeConiat M, Baller-ini P, Mauchauffe M, Della Valle V,Monni R, Nguyen Khac F, Mercher T,Penard-Lacronique V, Paturaud P,Gressin L, Hellig Rl, Daniel MT, Les-sard M, Berger R. 2001. A new recur-rent and specific cryptic translocation,t(5;14)(q35;q32), is associated withexpression of the Hox11L2 gene in Tacute lymphoblastic leukemia. Leuke-mia 15:1495–1504.
Bidet Y, Jagla T, Da Ponte JP, DastugueB, Jagla K. 2003. Modifiers of muscleand heart cell fate specification identi-fied by gain-of-function screen in Dro-sophila. Mech Dev 120:991–1007.
Bonini NM, Leiserson WM, Benzer S.1993. The eyes absent gene: geneticcontrol of cell survival and differentia-
tion in the developing Drosophila eye.Cell 72:379–395.
Bonini NM, Bui QT, Gray-Board GL, War-rick JM. 1997. The Drosophila eyesabsent gene directs ectopic eye forma-tion in a pathway conserved betweenflies and vertebrates. Development 124:4819–4826.
Bonini NM, Leiserson WM, Benzer S.1998. Multiple roles of the eyes absentgene in Drosophila. (1998). Dev Biol196:42–57.
Brugmann SA, Moody SA. 2005. Induc-tion and specification of vertebrate ecto-dermal placodes: precursors of thecranial sensory organs. Biol Cell 97:303–319.
Brugmann SA, Pandur PD, Kenyon KL,Pignoni F, Moody SA. 2004. Six1 pro-motes a placodal fate within the lateralneurogenic ectoderm by functioning asboth a transcriptional activator andrepressor. Development 131:5871–5881.
Buchou T, Vernet M, Blond O, Jensen H.H,Pointu H, Olsen BB, Cochet C, IssingerO-G, Boldyreff B. 2003. Disruption of theregulatory beta subunit of protein kinaseCK2 in mice leads to a cell-autonomousdefect and early embryonic lethality. MolCell Biol 23:908–915.
Burks PJ, Isaacs HV, Pownall ME. 2009.FGF signalling modulates transcrip-tional repression by Xenopus groucho-related-4. Biol. Cell 101:301–308.
Carpenter PB, Dunphy WG. 1998. Identi-fication of a novel 81-kDa component ofthe Xenopus origin of recognition com-plex. J Biol Chem 273:24891–24897.
Chen Z, Montcouquiol M, Caldderon R,Jenkins NA, Copeland NG, Kelley MW,Noben-Trauth K. 2008. Jxc1/Sobp,encoding a nuclear zinc finger protein,is critical for cochlear growth, cell fate,and patterning of the Organ of Corti.J Neurosci 28:6633–6641.
Cheyette BN, Green PJ, Martin K, GarrenH, Hartenstein V, Zipursky SL. 1994.The Drosophila sine oculis locus encodesa homeodomain-containing proteinrequired for the development of theentire visual system. Neuron 12:977–996.
Chiao E, Leonard J, Dickinson K, BakerJC. 2005. High-throughput functionalscreen of mouse gastrula cDNA libra-ries reveals new components of endo-derm and mesoderm specification.Genome Res 15:44–53.
Choudhury BK, Kim J, Kung HF, Li SS.1997. Cloning and developmentalexpression of Xenopus cDNAs encodingthe Enhancer of split groucho andrelated proteins. Gene 195:41–48.
Christophorou NAD, Bailey AP, HansonS, Streit A. 2009. Activation of Six1 tar-get genes is required for sensory pla-code formation. Dev Biol 336:327–336.
Courey AJ, Jia S. 2001. Transcriptionalrepression: the long and the short of it.Genes Dev 15:2786–2796.
David R, Ahrens K, Wedlich D, SchlosserG. 2001. Xenopus Eya1 demarcates allneurogenic placodes as well as migrat-ing hypaxial muscle precursors. MechDev 103:189–192.
EXPRESSION OF CANDIDATE SIX COFACTORS 3463
Dev
elop
men
tal D
ynam
ics

Davidovic L, Bechara E, Gravel M, JaglinX.H, Tremblay S, Sik A, Bardoni B,Khandjian EW. 2006. The nuclearmicospherule protein 58 is a novelRNA-binding protein that interactswith fragile X mental retardation pro-tein in polyribosomal mRNPs from neu-rons. Hum Mol Genet 15:1525–1538.
de la Calle-Mustienes E, Glavic A, Modo-lell J, Gomez-Skarmeta JL. 2002. Xirohomeoproteins coordinate cell cycle exitand primary neuron formation by upre-gulating neuronal-fate repressors anddownregulating the cell-cycle inhibitorXGadd45gamma. Mech Dev 119:69–80.
Dear TN, Colledge WH, Carlton MB, Lav-enir I, Larson T, Smith AJ, Warren AJ,Evans MJ, Sofroniew MV, Rabbitts TH.1995. The Hox11 gene is essential forcell survival during spleen develop-ment. Development 121:2909–2915.
Dehni G, Liu Y, Husain J, Stifani S. 1995.TLE expression correlates with mouseembryonic segmentation, neurogenesis,and epithelial determination. Mech Dev53:369–381.
Depreux FF. Darrow K, Conner DA,Eavey RD, Liberman MC, Seidman CE,Seidman JG. 2008. Eya4-deficient miceare a model for heritable otitis media.J Clin Invest 118:471–474.
Dominguez I, Mizuno J, Wu H, Song DH,Symes K, Seldin DC. 2004. Protein ki-nase CK2 is required for dorsal axis for-mation in Xenopus embryos. Dev Biol274:110–124.
Dorner S, Lum L, Kim M, Paro R, BeachyPA, Green R. 2006. A genome widescreen for components of the RNAipathway in Drosophila cultured cells.Proc Natl Acad Sci U S A 103:11880–11885.
Esteve P, Bovolenta P. 1999. cSix4, amember of the six gene family of tran-scription factors, is expressed duringplacode and somite development. MechDev 85:161–165.
Fabrizio JJ, Boyle JM, DiNardo S. 2003.A somatic role for eyes absent (eya) andsine oculis (so) in Drosophila spermato-cye development. Dev Biol 258:117–128.
Ferrando AA, Neuberg DS, Stauton J,Loh ML, Huard C, Raimondi SC, PulCH, Downing JR, Gilliland DG, LanderES, Golub TR Look AT. 2002. Geneexpression signatures define novel onco-genic pathways in T cell acute lympho-blastic leukemia. Cancer Cell 1:75–87.
Fogelgren B, Kuroyama MC, McBratney-Owen B, Spence AA, Malahn LE, Ana-wati MK, Cabatbat C, Alarcon VB,Marikawa Y, Lozanoff S. 2008. Misex-pression of Six2 is associated with her-itable frontonasal dysplasia and renalhypoplasia in 3H1 Br mice. Dev Dyn237:1767–1779.
Fogelgren B, Yang S, Sharp IC, HuckstepOJ, Ma W, Somoponpun SJ, CarlsonEC, Uyehara CFT, Lozanoff S. 2009.Deficiency in Six2 during prenatal de-velopment is associated with reducednephron number, chronic renal failure,and hypertension in BR/þ adult mice.Am J Renal Physiol 296:F1166–F1178.
Ghanbari H, Seo HC, Fjose A, Brandli AW.2001. Molecular cloning and embryonicexpression of Xenopus Six homeoboxgenes. Mech Dev 101:271–277.
Giot L, Bader JS, Brouwer C, ChaudhuriA, Kuang B, Li Y, Hao YL, Ooi CE,Godwin B, et al. 2003. A protein inter-action map of Drosophila melanogaster.Science 302:1727–1736.
Grifone R, Demignon J, Giordani J, NiroC, Souil E, Bertin F, Laclef C, Xu P-X,Maire P. 2007. Eya1 and Eya2 proteinsare required somatic myogenesis in themouse embryo. Dev Biol 302:602–616.
Guerra B, Issinger OG. 1999. Protein ki-nase CK2 and its role in cellular prolif-eration, development and pathology.Electrophoresis 20:391–408.
Hatano M, Roberts CW, Minden M, CristWM, Korsmeyer SJ. 1991. Deregulationof a homeobox gene, Hox11, by thet(10;14) in T cell leukemia. Science 253:79–82.
Hatano M, Aoki T, Dezawa M, Yusa S, Iit-suka Y, Koseki H, Taniguchi M, Toku-hisa T. 1997. A novel pathogenesis ofmegacolon in Ncx/Hox11L.1 deficientmice. J Clin Invest 100:795–801.
Heanue TA, Reshef R, Davis RJ, MardonG, Oliver G, Tomarev S, Lassar AB,Tabin CJ. 1999. Synergistic regulationof vertebrate muscle development byDach2, Eya2, and Six1, homologs ofgenes required for Drosophila eye for-mation. Genes Dev 15:3231–3243.
Hirohashi Y, Wang Q, Liu Q, Zhang H,Sato N, Greene MI. 2006. P78/MCRS1forms a complex with centrosomal pro-tein Nde1and is essential for cell viabil-ity. Oncogene 25:4937–4946.
Hollander MC, Sheikh MS, Bulavin DV,Lundgren K, Augeri-Henmueller L,Shehee R, Molinaro TA, Kim KE, TolosaE, Ashwell JD, Rosenberg MP, Zhan Q,Fernandez-Salguero PM, Morgan WF,Fornace AJ. 1999. Genomic instabilityin Gadd45a-deficient mice. Nat Genet23:176–184.
Hoskins BE, Cramer CH, Silvius D, ZouD, Raymond RM, Orten D, KimberlingWJ, Smith RJH, Weil D, Petit C, OttoEA, Xu PX, Hildebrandt F. 2007. Tran-scription factor SIX5 is mutated inpatients with Branchio-Oto-Renal syn-drome. Am J Hum Genet 80:800–804.
Ijiri K, Zerbini LF, Peng H, Correa RG,Lu B, Walsh N, Zhao Y, Taniguchi N,Huang XL, Otu H, Wang H, Wang JF,Komiya S, Ducy P, Rahman MU, FlavellRA, Gravallese EM, Oettgen P, Liber-mann TA, Goldring MB. 2005. A novelrole for GADD45beta as a mediator ofMMP-13 gene expression during chon-drocyte terminal differentiation. J BiolChem 280:38544–38555.
Ikeda K, Watanabe Y, Ohto H, KawakamiK. 2002. Molecular interaction and syn-ergistic activation of a promoter by Six,Eya, and Dach proteins mediatedthrough CREB binding protein. MolCell Biol 22:6759–6766.
Ishihara T, Ikeda K, Sato S, Yajima H,Kawakami K. 2008. Differential expres-sion of Eya1 and Eya2 during chick
early embryonic development. GeneExpr Patterns 8:357–367.
Johnson KR, Cook SA, Erway LC, Mat-thews AN, Sanford LP, Paradies NE,Friedman RA. 1999. Inner ear and kid-ney anomalies caused by IAP insertionin an intron of the Eya1 gene in amouse model of BOR syndrome. HumMol Genet 8:645–653.
Kawahara A, Che YS, Hanaoka R, TakedaH, Dawid IB. 2005. Zebrafish GADD45-beta genes are involved in somite seg-mentation. Proc Natl Acad Sci U S A102:361–366.
Kawakami K, Ohto H, Ikeda K, RoederRG. 1996. Structure, function andexpression of a murine homeobox pro-tein AREC3, a homologue of Drosophilasine oculis gene product, and implica-tion in development. Nucleic Acids Res24:303–310.
Kawakami K, Sato S, Ozaki H, Ikeda K.2000. Six family genes-structure andfunction as transcription factors andtheir roles in development. Bioessays22:616–626.
Kenyon KL, Li DJ Clouser C, Tran S,Pignoni F. 2005. Fly SIX-type homeodo-main proteins Sine oculis and Optixpartner with different cofactors duringeye development. Dev Dyn 234:497–504.
Killian A, Sarafan-Vasseur N, Sesboue R,Le Pessot F, Blanchard F, Lamy A, Lau-rent M, Flaman JM, Frebourg T. 2006.Contribution of the BOP1 gene, locatedon 8q24, to colorectal tumorigenesis.Genes Chromosomes Cancer 45:874–881.
Knouff RA. 1935. The developmental pat-tern of ectodermal placodes in Ranapipiens. J Comp Neurol 62:17–71.
Kobayashi M, Osanai H, Kawakami K,Yamamoto M. 2000. Expression of threezebrafish Six4 genes in the cranial sen-sory placodes and the developingsomites. Mech Dev 98:151–155.
Kobayashi M, Nishikawa K, Suzuki T,Yamamoto M. 2001. The homeobox pro-tein Six3 interacts with the Grouchocorepressor and acts as a transcrip-tional repressor in eye and forebrainformation. Dev Biol 232:315–326.
Koop KE, MacDonald LM, Lobe CG. 1996.Transcripts of Grg4, a murine groucho-related gene, are detected in adjacenttissues to other murine neurogenicgene homologues during embryonic de-velopment. Mech Dev 59:73–87.
Kowalinski E, Bange G, Bradatsch B,Hurt E, Wild K, Sinning I. 2007. Thecrystal structure of Ebp1 reveals a me-thionine aminopeptidase fold as bindingplatform for multiple interactions.FEBS Lett 581:4450–4454.
Kriebel M, Muller F, Hollemann T. 2007.Xeya3 regulates survival and prolifera-tion of neural progenitor cells withinthe anterior neural plate of Xenopusembryos. Dev Dyn 236:1526–1534.
Kumar S, Deffenbacher K, Cremers CW,Van Camp G, Kimberling WJ. 1997.Brachio-oto-renal syndrome: identifica-tion of novel mutations, molecular
3464 NEILSON ET AL.
Dev
elop
men
tal D
ynam
ics

characterization, mutation distributionand prospects for genetic testing. GenetTest 1:243–251.
Laclef C, Souil E, Demignon J, Maire P.2003. Thymus, kidney and craniofacialabnormalities in Six 1 deficient mice.Mech Dev 120:669–679.
Larkin MA, Blackshields G, Brown NP,Chenna R, McGettigan PA, McWilliamH, Valentin F, Wallace IM, Wilm A,Lopez R, Thompson JD, Gibson TJ,Higgins DG. 2007. Clustal W and Clus-tal X version 2.0. Bioinformatics 23:2947–2948.
Leon C, Lobe CG. 1997. Grg3, a murineGroucho-related gene, is expressed inthe developing nervous system and inmesenchyme-induced epithelial struc-tures. Dev Dyn 208:11–24.
Li X, Ohgi KA, Zhang J, Krones A, BushKT, Glass CK, Nigam SK, AggarwalAK, Maas R, Rose DW, Rosenfeld MG.2003. Eya protein phosphatase activityregulates Six1–Dach–Eya transcrip-tional effects in mammalian organogen-esis. Nature 426:247–254.
Lieberman SL, Ruderman JV. 2004.CK2b, which inhibits Mos function,binds to a discrete domain in the N-ter-minus of Mos. Dev Biol 268:271–279.
Lin DY, Shih HM. 2002. Essential role ofthe 58-kDa microspherule protein inthe modulation of Daxx-dependent tran-scriptional repression as revealed bynucleolar sequestration. J Biol Chem277:25446–25456.
Liu YH, Jakobsen JS, Valentin G, Amar-antos I, Gilmour DT, Furlong EE. 2009.A systemic analysis of Tinman functionreveals Eya and JAK-STAT signaling asessential regulators of muscle develop-ment. Dev Cell 16:280–291.
Logan C, Wingate RJ, McKay IJ, Lums-den A. 1998. Tlx-1 and Tlx-3 homeoboxgene expression in cranial sensory gan-glia and hindbrain of the chick embryo:markers of patterned connectivity.J Neurosci 18:5389–5402.
Lopez-Rios J, Tessmar K, Loosli F, Witt-brodt J, Bovolenta P. 2003. Six3 andSix6 activity is modulated by membersof the groucho family. Development130:185–195.
Lu B, Yu H, Chow C, Li B, Zheng W,Davis RJ, Flavell RA. 2001. GADD45g-amma mediates the activation of thep38 and JNK MAP kinase pathwaysand cytokine production in effector TH1cells. Immunity 14:583–590.
Lu M, Gong ZY, Shen WF, Ho AD. 1991.The tcl-3 proto-oncogene altered bychromosomal translocation in T cell leu-kemia codes for a homeobox protein.EMBO J 10:2905–2910.
Ma DK, Jang MH, Guo JU, Kitabatake Y,Chang ML, Pow-Anpongkul N, FlavellRA, Lu B, Ming GL, Song H. 2009.Neuronal activity-induced Gadd45b pro-motes epigenetic DNA demethylationand adult neurogenesis. Science 323:1074–1077.
Martelange V, De Smet C, De Plaaen E,Lurquin C, Boon T. 2000. Identificationon a human sarcoma of two new genes
with tumor-specific expression. CancerRes 60:3848–3855.
Mathieu MG, Linley AJ, Reeder SP, Bado-ual C, Tartour E, Rees RC, McArdleSE. 2010. HAGE, a cancer/testis anti-gen expressed at the protein level in avariety of cancers. Cancer Immun 10:2–8.
Miyasaka H, Choudhury BK. Hou EW, LiSS. 1993. Molecular cloning and expres-sion of mouse and human cDNA encod-ing AES and ESG proteins with strongsimilarity to Drosophila enhancer ofsplit groucho proteins. Eur J Biochem216:343–352.
Molenaar M, Brian E, Roose J, Clevers H,Destree O. 2000. Differential expressionof the Groucho-related genes 4 and 5during early development of Xenopuslaevis. Mech Dev 91:311–315.
Moody SA. 2000. Cell lineage analysis inXenopus embryos. In: Tuan RS, Lo CW,editors. Methods in molecular biology:developmental biology protocols. Vol.135. New York: Humana Press. p 1–17.Moody SA. 2007. Determination of pre-placodal ectoderm and sensory placo-des. In: Moody SA, editor. Principles indevelopmental genetics. New York:Elsevier. p 590–614.Nieuwkoop PD, Faber J. 1994. Normaltable of Xenopus laevis. New York: Gar-land Publishing.
Nybakken K, Vokes SA, Lin TY, McMa-hon AP, Perrimon N. 2005. A genome-wide RNA interference screen in Dro-sophila melanogaster cells for new com-ponents of the Hh signaling pathway.Nat Genet 37:1323–1332.
Ohto H, Kamada S, Tago K, Tominaga SI,Ozaki H, Sato S, Kawakami K. 1999.Cooperation of six and eya in activationof their target genes through nucleartranslocation of Eya. Mol Cell Biol 19:6815–6824.
Oliver G, Mailhos A, Wehr R, CopelandNG, Jenkins NA, Gruss P. 1995. Six3, amurine homologue of the sine oculisgene, demarcates the most anterior bor-der of the developing neural plate andis expressed during eye development.Development 121:4045–4055.
Ozaki H, Watanabe Y, Takahashi K, Kita-mura K, Tanaka A, Urase K, Momoi T,Sudo K, Sakagami J, Asano M, IwakureY, Kawakami K. 2001. Six4, a putativemyogenin gene regulator, is not essen-tial for mouse embryonal development.Mol Cell Biol 21:3343–3350.
Ozaki H, Nakamura K, Funahashi J,Ikeda K, Yamada G, Tokano H, Oka-mura HO, Kitamura K, Muto S, KotakiH, Sudo K, Horai R, Iwakura Y, Kawa-kami K. 2004. Six1 controls patterningof the mouse otic vesicle. Development131:551–562.
Pandur P, Moody S. 2000. Xenopus Six1gene is expressed in neurogenic cranialplacodes and maintained in the differ-entiating lateral lines. Mech Dev 96:253–257.
Patterson KD, Krieg PA. 1999. Hox11-family genes XHox11 and Xhox11L2 inXenopus: XHox11L2 expression is re-
stricted to a subset of the primary sen-sory neurons. Dev Dyn 214:34–43.
Peretz G, Bakhrat A, Abdu U. 2007.Expression of the Drosophila mela-nogaster GADD45 homolog (CG11086)affects egg asymmetric developmentthat is mediated by the c-jun N-termi-nal kinase pathway. Genetics 177:1691–1702.
Pfister M, Toth T, Thiele H, Haack B,Blin N, Zenner HP, Sziklai I, NurnbergP, Kupka S. 2002. A 4-bp insertion inthe eya-homologous region (eyaHR) ofEya4 causes hearing impairment in aHungarian family linked to DFNA10.Mol Med 10:607–611.
Pignoni F, Hu B, Zavitz KH, Xiao J, Garr-ity PA, Zipursky SL. 1997. The eye-specification proteins So and Eya forma complex and regulate multiple stepsin Drosophila eye development. Cell 91:881–891.
Pinna LA, Meggio F. 1997. Protein kinaseCK2 (‘‘casein kinase-2’’) and its implica-tion in cell division and proliferation.Prog Cell Cycle Res 3:77–97.
Raju K, Tang S, Dube ID, Kamel-Reid S,Bryce DM, Brietman ML. 1993. Charac-terization and developmental expres-sion of Tlx-1, the murine homolog ofHox11. Mech Dev 44:51–64.
Roberts CW, Shutter JR, Korsmeyer S.1994. Hox11 controls the genesis of thespleen. Nature 368:747–749.
Roberts CWM, Sonder AM, Lumsden A,Korsmeyer SJ. 1995. Developmentalexpression of Hox11 and specification ofsplenic cell fate. Am J Pathol 146:1089–1101.
Rodriguez-Soriano J. 2003. Brachio-oto-re-nal syndrome. J Nephrol 16:603–605.
Rohrmoser M, Holzel M, Grimm T, Mala-moussi A, Harasim T, Orban M, Pfis-tere I, Gruber-Eber A, Kremmer E,Eick D. 2007. Interdependence of Pes1,Bop1, and WDR12 controls nucleolarlocalization and assembly of the PeBoWcomplex required for maturation of the60S ribosomal subunit. Mol Cell Biol27:3682–3694.
Roose J, Molenaar M, Peterson J,Brantjes H, Moerer P, van de WeteringM, Destree O, Clever H. 1998. The Xen-opus Wnt effector XTcf-3 interacts withGroucho-related transcriptional repress-ors. Nature 395:608–612.
Rual J-F, Venkatesan K, Hao T, Hirozane-Kishikawa T, Dricot A, Li N, Berriz GF,Gibbons FD, Dreze M, Ayivi-Guede-houssou N, Klitgord N, Simon C,Boxem M, Milstein S, Rosenberg J,Goldberg DS, Zhang LV, Wong SL,Franklin G, et al. 2005. Towards a pro-teome-scale map of the human protein-protein interaction network. Nature437:1173–1178.
Ruf RG, Xu PX, Silvius D, Otto EA, Beek-mann F, Muerb UT, Kumar S, NeuhausTJ, Kemper MJ, Raymond RMJ, BrophyPD, Berkman J, Gattas M, Hyland V,Ruf EM, Schwartz C, Chang EH, SmithRJ, Stratakis CA, Weil D, Petit C, et al.2004. SIX1 mutations cause branchio-oto-renal syndrome by disruption of
EXPRESSION OF CANDIDATE SIX COFACTORS 3465
Dev
elop
men
tal D
ynam
ics

EYA1-SIX1-DNA complexes. Proc NatlAcad Sci U S A 101:8090–8095.
Sahly I, Andermann P, Petit C. 1999. Thezebrafish eya1 gene and its expressionpattern during embryogenesis. DevGenes Evol 209:399–410.
Sato S, Ikeda K, Shioi G, Ochi H, OginoH, Yajima H, Kawakami K. 2010. Con-served expression of mouse Six1 in thepre-placodal region (PPR) and identifi-cation of an enhancer for the rostralPPR. Dev Biol 344:158–171.
Schlosser G. 2006. Induction and specifi-cation of cranial placodes. Dev Biol 294:303–351.
Schlosser G, Ahrens K. 2004. Molecularanatomy of placode development inXenopus laevis. Dev Biol 271:439–446.
Schlosser G, Northcutt RG. 2000. Devel-opment of neurogenic placodes in Xeno-pus laevis. J Comp Neurol 418:121–146.
Schlosser G, Awtry T, Brugmann SA, Jen-sen ED, Neilson K, Ruan G, StammlerA, Voelker D, Yan B, Zhang C, Klym-kowsky MW, Moody SA. 2008. Eya1and Six1 promote neurogenesis in thecranial placodes in a SoxB1-dependentfashion. Dev Biol 320:199–214.
Schonberger J, Wang L, Shin JT, Kim SD,Depreux FS, Zhu H, Zon L, Pizard A,Kim JB, MacRae CA, Mungall AJ, Seid-man JG, Seidman CE. 2005. Mutationin the transcriptional coactivator Eya4causes dilated cardiomyopathy and sen-sorineural hearing loss. Nat Genet 37:418–422.
Self M, Geng X, Oliver G. 2009. Six2 ac-tivity is required for the formation ofthe mammalian pyloric sphincter. DevBiol 334:409–417.
Serikaku MA, O’Tousa JE. 1994. sine ocu-lis is a homeobox gene required for Dro-sophila visual system development.Genetics 138:1137–1150.
Shirasawa S, Arata A, Onimaru H, RothKA, Brown GA, Horning S, Arata S,Okumura K, Sasazuki T, Korsmeyer S.2000. Rnx deficiency results in congeni-tal central hypoventilation. Nat Genet24:287–290.
Silver SJ, Davies EL, Doyon L, Rebay I.2003. Functional dissection of eyesabsent reveals new modes of regulationwithin the retinal determination genenetwork. Mol Cell Biol 23:5989–5999.
Sithanandam G, Anderson LM. 2008. TheERBB3 receptor in cancer and cancergene therapy. Cancer Gene Ther 15:413–448.Sive HL, Grainger RM, Harland RM.2000. Early development of Xenopuslaevis: a laboratory manual. ColdSpring Harbor, NY: Cold Spring HarborLaboratory Press.
Soker T, Dalke C, Puk O, Floss T, BeckerL, Bolle I, Favor J, Hans W, Holter SM,Horsch M, Kallnik M, Kling E, MoerthC, Schrewe A, Stigloher C, Topp S, Gai-
lus-Durner V, Naton B, Beckers J,Fuchs H, Ivandic B, Klopstock T,Schulz H, Wolf E, Wurst W, Bally-CuifL, de Angelis MH, Graw J. 2008. Pleio-tropic effects in Eya3 knockout mice.BMC Dev Biol 8:118–136.
Song DH, Sussman DJ, Seldin DC. 2000.Endogenous protein kinase CK2 partici-pates in Wnt signaling in mammaryepithelial cells. J Biol Chem 275:23790–23797.
Spitz F, Demignon J, Porteu A, Kahn A,Concordet JP, Daegelen D, Maire P.1998. Expression of myogenin duringembryogenesis is controlled by Six/sineoculis homeoproteins through a con-served MEF3 binding site. Proc NatlAcad Sci U S A 95:14220–14225.
Stramer B, Winfield M, Shaw T, MillardTH, Woolner S, Martin P. 2008. Geneinduction following wounding of thewild-type versus macrophage-deficientDrosophila embryos. EMBO Rep 9:465–471.
Streit A. 2004. Early development of thecranial sensory nervous system: from acommon field to individual placodes.Dev Biol 276:1–15.
Streit A. 2007. The preplacodal region: anectodermal domain with multipotentialprogenitors that contribute to senseorgans and cranial sensory ganglia. IntJ Dev Biol 51:447–461.
Strezoska Z, Pestov DG, Lau LF. 2000.Bop1 is a mouse WD40 repeat nucleolarprotein involved in 28S and 5.8S rRNAprocessing and 60S ribosome biogenesis.Mol Cell Biol 20:5516–5528.
Strezoska Z, Pestov DG, Lau F. 2002.Functional inactivation of the mousenucleolar protein Bop1 inhibits multiplesteps in pre-rRNA processing andblocks cell cycle progression. J BiolChem 277:29617–29625.
Takewawa M, Saito H. 1998. A family ofstress-inducible GADD45-like proteinsmediate activation of the stress-respon-sive MTK1/MEKK4 MAPKKK. Cell 95:521–530.
Tessmar K, Loosli F, Wittbrodt J. 2002. Ascreen for co-factors of Six3. Mech Dev117:103–113.
Uchiyama K, Otsuka R, Hanaoka K.1999. cHox11L2, a Hox11 related gene,is expressed in the peripheral nervoussystem and subpopulation of the spinalcord during chick development. Neuro-sci Lett 273:97–100.
Vincent C, Kalatzis V, Abdelhak S, ChaibH, Compain S, Helias J, Vaneecloo FM,Petit C. 1997. BOR and BO syndromesare allelic defects of EYA1. Eur J HumGenet 5:242–246.
Wayne S, Robertson NG, DeClau F, ChenN, Verhoeven K, Prasad S, TranebjargL, Morton CC, Ryan AF, Van Camp G,Smith RJ. 2001. Mutations in the tran-scriptional activator Eya4 cause late-
onset deafness at the DFN10 locus.Hum Mol Genet 10:195–200.
Webb JF, Noden DM. 1993. Ectodermalplacodes: contributions to the develop-ment of the vertebrate head. Am Zool33:434–447.
Wilhelm V, Rojas P, Gatica M, AllendeCC, Allende JE. 1995. Expression of thesubunits of protein kinase CK2 duringoogenesis in Xenopus laevis. Eur J Bio-chem 232:671–676.
Willert K, Brink M, Wodarz A, Varmus H,Nusse R. 1997. Casein kinase 2 associ-ates with and phosphorylates dishev-eled. EMBO J 16:3089–3096.
Xia X, Lessor TJ, Zhang Y, Woodford N,Hamburger AW. 2001. Analysis of theexpression pattern of Ebp1, an ErbB-3-binding protein. Biochem Biophys ResCommun 289:240–244.
Xu PX, Cheng J, Epstein JA, Maas RL.1997. Mouse Eya genes are expressedduring limb tendon development andencode a transcriptional activationfunction. Proc Natl Acad Sci U S A 94:11974–11979.
Xu PX, Adams J, Peters H, Brown MC,Heaney S, Maas R. 1999. Eya1-deficientmice lack ears and kidneys and showabnormal apoptosis of organ primordia.Nat Genet 23:113–117.
Xu PX, Zheng W, Laclef C, Maire P, MaasRL, Peters H, Xu X. 2002. Eya1 isrequired for the morphogenesis of mam-malian thymus, parathyroid and thy-roid. Development 129:3033–3044.
Xu PX, Zheng W, Huang L, Maire P,Laclef C, Silvius D. 2003. Six1 isrequired for the early organogenesis ofmammalian kidney. Development 130:3085–3094.
Yaklichkin S, Steiner AB, Lu Q, KesslerDS. 2007. FoxD3 and Grg4 physicallyinteract to repress transcriptional andinduce mesoderm in Xenopus. J BiolChem 282:2548–2557.
Yoo JY, Wang XW, Rishi AK, Lessor T,Xia XM, Gustafson TA, Hamburger AW.2000. Interaction of the PA2G4 (EBP1)protein with ErbB-3 and regulation ofthis binding by heregulin. Br J Cancer82:683–690.
Zhang Y, Lu Y, Zhou H, Lee M, Liu Z,Hassel BA, Hamburger AW. 2008. Alter-ations in cell growth and signaling inErbB3 binding protein-1 (Ebp1) defi-cient mice. BMC Cell Biol 9:69–81.
Zheng W, Huang L, Wei ZB, Silvius D,Tang B, Xu PX. 2003. The role of Six1in mammalian auditory system develop-ment. Development 130:3989–4000.
Zhu CC, Dyer MA, Uchikawa M, KondohH, Lagutin OV, Oliver G. 2002. Six3-mediated auto repression and eye devel-opment requires its interaction withmembers of the Groucho-related familyof co-repressors. Development 129:2835–2849.
3466 NEILSON ET AL.
Dev
elop
men
tal D
ynam
ics