detection of infectious centers in c57bl/ka lymphoid cell...
TRANSCRIPT

[CANCER RESEARCH 40, 544-549. March 1980]0008-5472/80/0040-0000$02.00
Detection of Infectious Centers in C57BL/Ka Lymphoid Cell PopulationsInfected in Vitro by the Radiation Leukemia Virus1
Jacques Boniver,2 Abain Decbève,Obivera J. Finn, Cyril Honsik, Miriam Lieberman, and Henry S. Kaplan3
Cancer Biology Research Laboratory, Department of Radiology, Stanford University Medical Center, Stanford, California 94305
ABSTRACT
The cocubtivation of nonproducer bymphoma cells derivedfrom a radiation-induced bymphomaof the C57BL/Ka mousewith cultures of lymphoid cell populations from the thymus,spleen, and marrow of the same strain 48 hr after their infectionby the C57BL/Ka leukemia viruses permits the detection ofinfectious centers in these cultures. A quantitative assay isdescribed which allows the estimation in bymphoidcell subpopubationsof the numbers of target cells susceptible to productiveinfection by the thymotropic and leukemogenic viruses ofC57BL/Ka mice in vitro. This assay should greatly facilitate theidentification and characterization of such target cells.
INTRODUCTION
RadLV,4 derived from radiation- and virus-induced thymicbymphomasof CS7BL/Ka mice (2), and RadLV/LTC, the virusproduced by lymphoma cell lines in vitro (3, i 7), are both L+in this strain of mice (2, 3, 7). They are also designated as T+because virus expression occurs selectively in the subcapsulararea of the thymus after i.v. inoculation into C57BL/Ka hosts(7, 8). RadLV and RadLV/LTC can be assayed in vivo on thebasis of their thymotropic and leukemogenic activity (2, 3, 5).They can also be assayed in vitro due to their ability to act ashelper viruses in the muninesarcoma virus focus assay (1, 1i)and to their ability to replicate in nonproducer lymphoma celllines of C57BL/Ka origin (19). We now present an indirectcocultivation assay which reliably leads to the detection ofinfectious centers in cultures of lymphoid cell populations fromthe thymus, spleen, and marrow of C57BL/Ka mice afterinfection and cultivation of such cells in vitro. From a practicalstandpoint, the present procedure greatly facilitates in vitrostudies on the identification and characterization of the targetbymphoid cell populations susceptible to productive infectionby the T+L+ viruses of CS7BL/Ka mice.
MATERIALS AND METHODS
Mice. Four-week-old female CS7BL/Ka (FV@1bb)inbred mice
I This work was supported by Contract NOl -CP-71 052 from the National
Cancer Institute, NIH, Department of Health, Education, and Welfare, and by giftsto the Jerry M. Tenney Memorial Fund for Cancer Research.
2 Holder of Public Health Service International Fellowship 1 F05 TW02546; on
leave of absence from the Department of Pathology, University of Liege, Belgium.Laureate of the Braconnier-Lamarche Foundation, Liege, Belgium.
3 To whom requests for reprints should be addressed.
4 The abbreviations used are: RadLV, radiation leukemia virus; L+ , leukemo
genic; T+. thymotropic; XC, Rous sarcoma virus-transformed rat cells; MEM,minimum essential medium; FCS, fetal calf serum; Dulbeccos phosphatebuffered saline (0.8% NaCI solution containing, per liter, 0.10 g CaCI2,0.20 gKCI, 0.20 g KH,P04. 0.iO g MgCb,.6H,O, and 2.i6 g NaH2PO4.7H20); RPMIMedium 1640, RosweblPark Memorial Institute Tissue Culture Medium 1640; IF,indirect immunofluorescence; CVA, cytoplasmic viral antigen.
Received June 29, 1979; accepted November 12, 1979.
and NIH/Swiss (Fv-1―)random-bred mice were obtained fromour colony.
Viruses. RadLV/VL3, a T+L+ virusbiologicallyand senobogicalby identical to RadLV (3), was obtained from the culturefluids of the BL/VL3 cell line derived from a RadLV-inducedCS7BL/Ka thymic lymphoma (i 7). The RadLV/VL3 preparationused in these experiments had a titer of 2.5 x i O@infectiousunits/mb as estimated by the immunofluorescence assay invivo (5).
BL/Ka(B) is a cloned B-fibrotnopic virus isolated fromCS7BL/Ka fibrobbastcultures (2). This virus is nonthymotropicand nonleukemogenic (2, 4). The isolate used in these expeniments had a titer of 2.5 x i 06 infectious units/mb as estimatedby immunofluorescence (6) and reverse XC assay (26) in vitro.
Cells. The BL/RL12-NPlymphomacell linewas derivedfroma radiation-induced C57BL/Ka thymoma. These cells do notproduce infectious viruses and are negative for the presenceof cytoplasmic or membrane viral antigens, as analyzed byimmunofluorescence and radioimmunocompetition assay.They are permissive for infection by both RadLV/VL3 (i 7) andBL/Ka(B) (i 8). The BL/RL12-NP cell line is maintained inEagle's MEM supplemented with i 0% FCS, penicillin (100units/mb), and streptomycin (i 00 @g/ml).
Cell Suspensionand Short-Term Cultures. Thymusesandspleens were minced with scissors in PBS (Grand Island Biological Co., Grand Island, N.Y.). The minced tissues werepassed through a nylon screen and suspended in PBS with 5%FCS. Bone marrow cell suspensions were obtained by flushingthe cells out of mouse femurs with 5% FCS-PBS using a syringeand a 25-gauge needle. After counting, the thymus, spleen,and marrow cell suspensions were washed twice in 5% FCSPBS and resuspended at a concentration of 2.5 x i O@cebbs/ml in RPMI Medium 1640 containing 5% FCS.
Infection Procedure and Culture Conditions for the BL/RL12-NPCocultivatlon Assay. Unless otherwise stated, thymus, spleen, and marrow cell suspensions were first incubatedin RPMI medium i 640 containing 5% FCS and DEAE-dextran(25 pg/mb) (30) for 15 mm and centrifuged at 1000 rpm at 4°for i 0 mm. The cells were then resuspended at a concentrationof 2.5 x i 07/ml in 1 ml RPMI Medium 1640-5% FCS containing pobybrene(4 @tg/mb)(29) and mixed with 1 ml of undilutedRadLV/VL3 in a S-mbplastic tube before incubation for 2 hr at370 in a 5% CO2 incubator. The tubes were shaken every 1@
mm to prevent cell sedimentation. After infection, the cells werespun at 1000 rpm at 4°for i 0 mm and resuspended in RPMIMedium i 640 supplemented with i 0% heat-inactivated FCS,antibiotics, 2 mM L-glutamine, i mM sodium pyruvate, 1%nonessential amino acids, and 50 j@M2-mercaptoethanob. Thecells were plated in 35-mm dishes (Lux Scientific Corporation,Newbury Park, Calif.) and maintained in culture at 37°in a 5%CO2 incubator.
In a typical cocultivation assay, the live cells were counted
544 CANCERRESEARCHVOL. 40
on March 11, 2019. © 1980 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from
50 60 70
Target ce!!s for !nfection by RadLV
by the trypan blue exclusion test 48 hr after infection. The cellsuspensions were then spun down for i 0 mm at 1000 rpm inthe cold and resuspended in Eagle's MEM with i 0% FCS andantibiotics to obtain a final concentration of 1 x 1O@cells/mb.These suspensions were then irradiated with 5000 A in a 137Csirradiator (Mark I, Model 30; Shepherd and Assoc., Glendale,Calif.). Serial dilutions of the individual thymus, spleen, andmarrow cell preparations were made in 1 ml of medium andthen cocultivated, in triplicate, in 35-mm dishes with 2 x i O@BL/RL12-NP cells suspendedin 1 ml of Eagle's MEM supplemented with 10% FCS and antibiotics. These cocultunes, inwhich only the unirradiated BL/RL12-NP indicator cells survive,were passaged every 3 days at a concentration of 2 x i O@celbs/35-mm dish. At 3 and 10 days after cocultivation, theywere analyzed by IF for the presence of CVA (6). In someinstances, the BL/RL12-NP cocubtivation assay was performedin microtiter plates (Falcon Microtest II Tissue Culture Plate3040, Lid 304i ; Falcon Plastics, Los Angeles, Calif.) in which0.i ml of various lymphoid cell concentrations was mixed with0.1 ml of BL/RL12-NP cells at a concentration of 2 x i O@cells/ml.
RESULTS
Detection of Infectious Centers by Cocubtivation of ThymicLymphocyte Cultures with BL/RL1@-NPLymphoma Cells.Preliminary experiments were first designed to determinewhether virus production by lymphoid cells could be detectedby cocultivation in vitro with BL/RL12-NP indicator cells. Cellsuspensions containing from i to 100 IF-positive thymocytesobtained from a CS7BL/Ka mouse thymus 3 weeks after
intrathymic inoculation of 106 infectious units of RadLV/VL3were mixed with normal thymocytes obtained from a normalweanbingC57BL/Ka mouse to yield total cell suspensions withconcentrations ranging from i 02 to i 0@cells in 1 ml of tissueculture medium. These samples were irradiated with 5000 Aand mixed in 35-mm dishes with i ml of a suspension containing 2 x i O@BL/RL12-NP cells. Triplicate dishes for eachexperimental point were then passaged 3 times and maintainedin culture for a total of i 0 days, at which time the BL/AL12-NPindicator cells were processed for detection of CVA by IF. Thedata (not shown) of these experiments indicate that this procedure permits the detection of 1 virus-positive cell in i 02 toi 06 thymocytes. A 5- to 10-fold decrease of sensitivity wasobserved when thymocyte populations larger than i O@celbs/ml were mixed with the indicator cells. This could be explainedby the observation that the growth of BL/RL12-NP cells issignificantly reduced when the ratio of bymphoidcells to BL/RL12-NPcells is greater than 5.
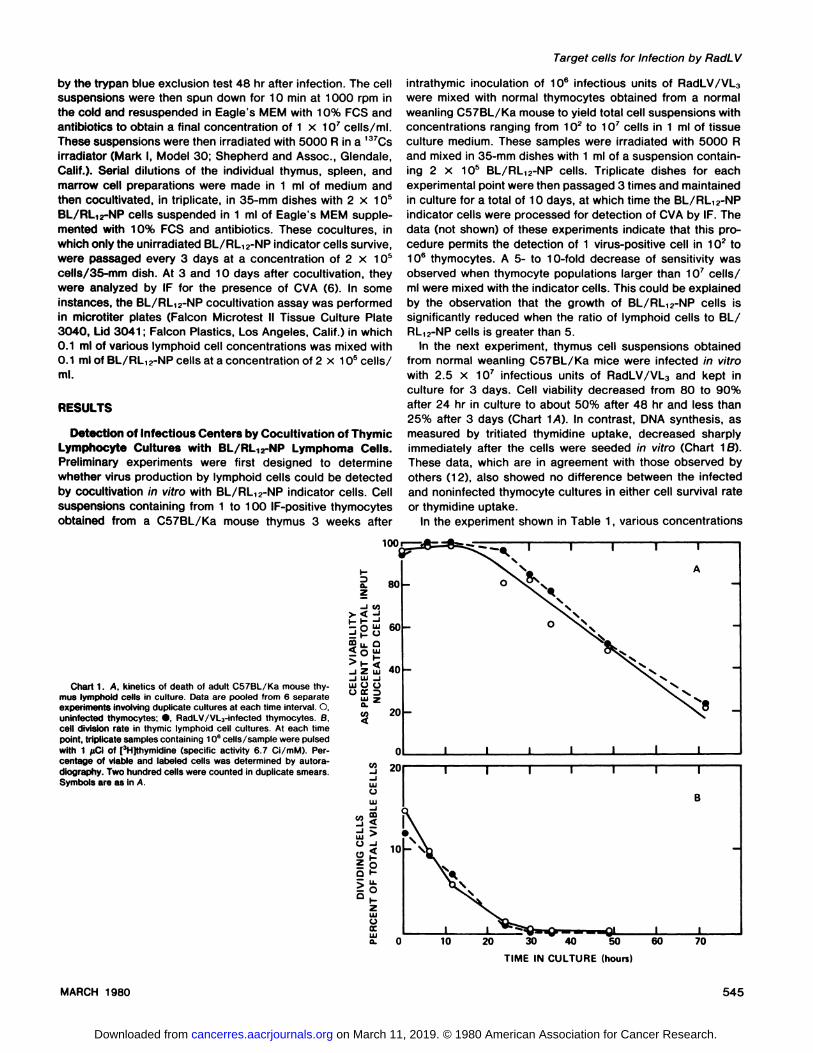
In the next experiment, thymus cell suspensions obtainedfrom normal weanling C57BL/Ka mice were infected in vitrowith 2.5 x i 0@infectious units of RadLV/VL3 and kept inculture for 3 days. Cell viability decreased from 80 to 90%after 24 hr in culture to about 50% after 48 hr and less than25% after 3 days (Chart iA). In contrast, DNA synthesis, asmeasured by tnitiated thymidine uptake, decreased sharplyimmediately after the cells were seeded in vitro (Chart i B).These data, which are in agreement with those observed byothers (i 2), also showed no difference between the infectedand noninfected thymocyte cultures in either cell survival rateor thymidine uptake.
In the experiment shown in Table 1, various concentrations
2L@ j I I I I I I
1O\k@
w—-w.-20 30 40
I I I
60- 1@@
@ A:80 - 0
40-
20 -
I I In I 1 1 1
a.z-“I,
>I_<_iZw-1w-iwOC.)
U)4
U)-1-IwC)w-1
..i<
Q..J
01——U.-00@_
zwC)a:wa.
Chart1. A, kineticsof deathof adultCS7BL/Kamousethymus lymphoid cells in culture. Data are pooled from 6 separateexperimentsinvolvingduplicateculturesat eachtimeinterval.0,uninfected thymocytes; •,RadLv/vL3-infected thymocytes. B,cell division rate In thymic lymphoid cell cultures. At each timepoint, triplicate samples containing 10°cells/sample were pulsedwith 1 @clof [3Hjthymidine (specific activity 6.7 Ci/mM). Percentage of viable and labeled cells was determined by autoradiography. Two hundred cells were counted in duplicate smears.Symbols are as In A.
0 10
TIME IN CULTURE (hours)
MARCH 1980 545
on March 11, 2019. © 1980 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

Virus detection in thymocyte cultures at intervals after infection of various numbers of thymocyteswithRadLV/VL3Presence
of virus at following time intervals between infection and cocultivationof thymocytes with BL/RL,@,-NPcellsbi
day 2 days 3 days 4daysInfec-
Infec- Infec- InfecNo. of thymo- Infec- tion + Infec- tion + Infec- tion + Infec- tion+cytes
at time of tion 5000 tion 5000 tion 5000 tion 5000infectiona only R only R only R onlyR5x107
+ + + — + — ——1x i07 + + + — — — ——1x106
+ + + — — — ——1x i05 + + — — — — ——1x io@ + + — — — — ——1x i03 + + — — — — ——None
(virus only) + + — — — — ——a
Thethymocytecultureswereinfectedwith2.5 x 1O@infectiousunitsofRadLV/VL3,andone-halfofthecultures were then immediately irradiated with 5000R.bAt each time interval, the infected cultures were irradiated with 5000 R and then cocultivated with2x
i 0' BL/RL,2-NP cells. The presence of IF-positive cells in the indicator cell cultures was determined 10
J. Boniver et a!.
Table 1
days after cocultivation.
of thymic lymphocytes from weanbing C57BL/Ka mice wereseeded in 35-mm dishes in 1 ml tissue culture medium andincubated with an equal volume of a AadLV/VL3 preparationcontaining 2.5 x 1O@infectious particles. Some culture disheswere seeded with virus alone. After the virus infection procedune was completed, one-half of the cell cultures for eachexperimental point were irradiated with 5000 A in order toprevent cell division and active virus replication. The irradiatedand unirradiated cultures were then returned to the incubatonand maintained in culture for 1 to 4 days. At each time interval,3 dishes/point were irradiated with 5000 A. Then 0.1 ml of aBL/AL12-NP cell suspension containing 2 x 106 cells/mb wasadded to each culture and to the dishes containing virus alone.The BL/RL12-NP indicator cells were examined by IF 10 dayslater. The IF positivity observed in the indicator cells plated atthe 1-day interval is attributed either to free virus present in theculture fluid or to vinions passively absorbed on carrier cells,since the BL/RL12-NP cells became infected in both the irnadiated and unirradiated cultures as well as in dishes whichcontained only virus (Table 1). In accordance with a previousreport from this laboratory (25), RadLV/VL3 maintained inculture medium for 2 days was no longer infectious. BL/RL12-NP indicator cells added 48 hr after infection and irradiation ofthe thymocyte cultures did not reveal viral antigens, whereasthese cells became IF positive when added to infected nonirradiated thymus lymphocyte cultures at the 48-hr interval,which strongly suggests that the detection of virus can beattributed to the presence of infectious centers in such cultures.
We next proceeded to exclude the possibility that passiveabsorption of viral particles by canniencells might be responsible for infection of the detector cells at the 48-hr interval in thecocubtivation assay. Since FV-1 restriction is not related tovirus absorption on penetration but rather involves an intracebbubarevent (15, 28), CS7BL/Ka thymocytes (Fv-1@)and NIH/Swiss thymocytes (Fv- 1fl@@)were both infected in vitro withRadLV/VL3 (which is B-tropic) and then assayed in the cocubtivation assay with BL/RL12-NP detector cells. The differentiation restriction system was also used in this experiment. Asdemonstrated earlier in this laboratory (2, 4), normal thymocytes from weanling mice can undergo productive infection byT+ virus but not by the fibnotropic-ecotnopic isolates from
strain CS7BL/Ka. Therefore, CS7BL/Ka thymocytes were alsoinfected with BL/Ka(B), a B-tropic fibrotnopic-ecotropic virus.As in the previous experiment, indicator cells were added toculture dishes that only contained either RadLV/VL3 on BL/Ka(B) and to AadLV/VL3-infected C57BL/Ka thymocyte cubtuneswhich had been irradiated immediately after infection. Asa final control, some RadLV/VL3-infected CS7BL/Ka thymuslymphocyte cultures were treated with trypsin and washedthoroughly immediately before cocultivation with the incubatorcells in order to remove any viral particles which might havebeen carried passively on their cell membranes.
The data presented in Table 2 clearly argue against thepossibility of passive absorption. The indicator cells became IFpositive only in the culture dishes that contained dividing unirradiated CS7BL/Ka thymocytes that had been infected withRadLV/VL3, a virus of the appropriate Fv-1 and cytotypictropism. Treatment with trypsin and washing did not preventinfection of the indicator cells 48 hr after infection. In earlierexperiments, the ability of trypsin tneatmentto remove passivelyadsorbed vinions from the cell surface was documented; infection of BL/RL12-NP indicator cells by the BL/Ka(B) virus passively adsorbed to the surface of thymocytes was preventedwhen the thymocytes were first treated with trypsin (data notshown). Viral antigens did not appear in indicator cells cocubtivated with NIH/Swiss thymocytes infected with RadLV/VL3,CS7BL/Ka thymocytes infected with BL/Ka(B), or CS7BL/Kathymocytes irradiated after infection with RadLV3. These expeniments (Tables 1 and 2) indicate that thymic lymphocytecultures infected with RadLV/VL3 concentrations greaten than1 x 106 infectious units/mb produce at beast1 surviving infectious center/S x 1O@thymic lymphoid cells after 2 days (assuming 50% survival of 1 x 106 thymocytes) and 1 infectiouscenter in 1.5 x 1o@cells after 3 days in culture (assuming 23%survival of a starting population of S x 10@'thymocytes). At 4days, when only 10% of cultured thymus cells were still viable,no infectious centers were detected, even in cultures whichoriginally contained more than 10 million cells (Table 1). 5everal attempts to infect lymphoid cell cultures with RadLV/VL3at a multiplicity of infection of less than 0.1 consistently failedto produce infectious centers detectable in this assay. Finally,as observed earlier in this laboratory with permanently infected
546 CANCERRESEARCHVOL. 40
on March 11, 2019. © 1980 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from
ThymocytesVirus typeAdditional treatment%
of IFpositivedetectorcells 1Scell dou
blings afterplatingNoneRadLV/VL3aNone0NoneBL/Ka(B)8None0C57BL/Ka
(Fv-1@RadLV/VLNone90C578L/Ka
(Fv-l@RadLV/VLTrypsinCC57BL/Ka
(Fv-1@RadLV/VLlrradiation'0C5TBL/Ka
(Fv-l@BL/Ka(B)None0NIH/Swiss
(Fv-l―t'RadLV/VLNone0
Table3Numberof infectious centers detected by the BL/RL,2-NP cocultivation assay
at 48 hr in cultures of RadLv/vL3-infected cell suspensions from thymus,marrow, and spleens of weanling C57BL/Kamice%
of IF-posi- No.of infectiouscenters/Cell types tive cells8 total no. ofcellsNormal
thymocytes <0.1 1/3 X 10@-2x 1O'@'Corticoresistant
thymocytesC NDd i /3 X 10@Normal
marrow cells <0. 1 1/ 1 x 1O@-1 x1Normal
spleen cells <0. i i / 1 x 10'
Target cells for Infection by RadLV
Table 2Virus detection by BL/RL,2-NP detector cells plated 48 hr after inoculation of
culture dishes with virus, cells, or both,0
/0//
//
,,
0...-@@ 0•
1a RSdLV/VLO(5x 10°infectiousunits/mi)andBL/Ka(B)(i x 106infectious
units/mI) were maintained at 37°In 2 ml of tissue culture medium for 2 days in35-mm tissue culture dishes prior to the addition to these dishes of 2 x 10'indicator cells in 0.1 ml of medium.
b@ (1 O@)were incubated with the concentration of virus isolatesindicated In Footnote a and maintained in culture for 2 days prior to irradiationwith 5000 R and addition of 2 x 10' Indicator cells in 0.1 ml of medium.
C Two-day-old cultures were treated with 0.05% trypsin and EDTA for S mmand thoroughly washed before the addition of the indicator cells.
d Thymocyte cultures were irradiated at a dose of 5000 R immediately after2 hr of incubation with RadLV/vL3.
fibroblasts (26) and BL/VL3 cells in the XC cell assay (17),irradiation with 5000 R did not prevent BL/VL3 cells fromacting as infectious centers in the BL/RL12-NP cocultivationassay (data not shown), indicating that, once a cell is activelyreplicating virus, irradiation immediately prior to cocultivationdoes not prevent it from acting as an infectious center eventhough the cell is no longer able to divide.
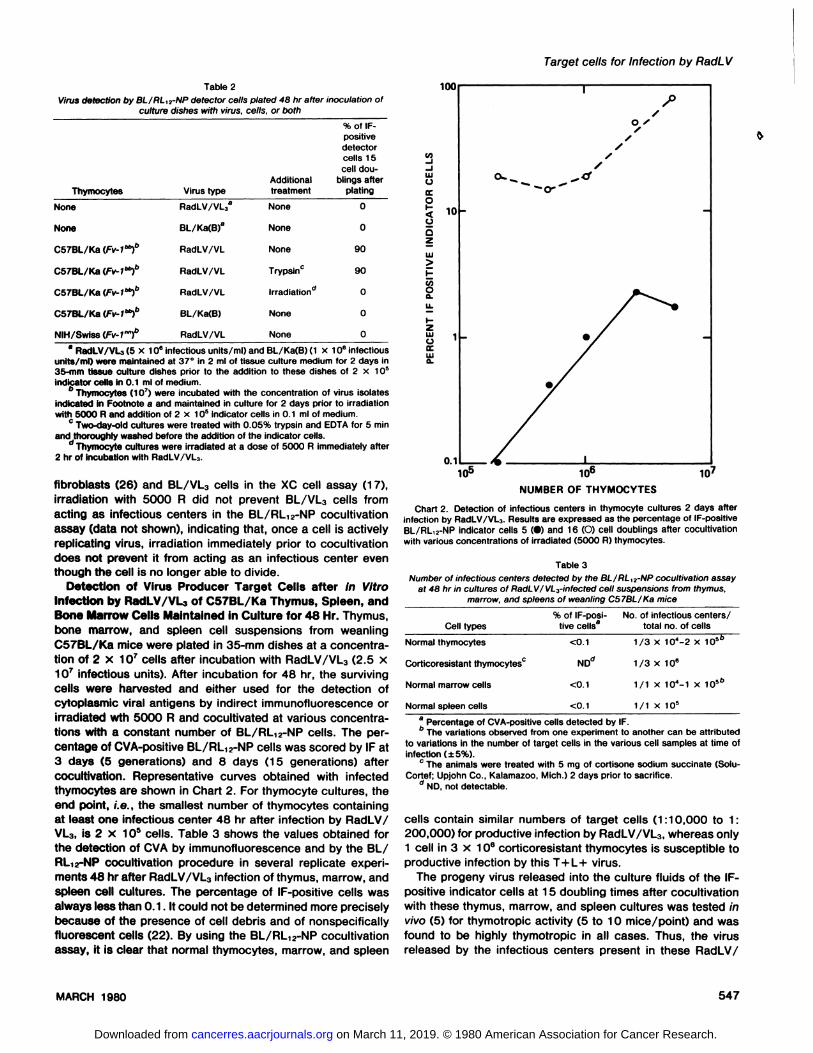
Detection of Virus Producer Target Cells after In VitroInfection by RSdLV/VL of C57BL/Ka Thymus, Spleen, andBone Marrow Cells Maintained in Culture for 48 Hr. Thymus,bone marrow, and spleen cell suspensions from weanbingC57BL/Ka mice were plated in 35-mm dishesat a concentration of 2 x 1O@cells after incubation with RadLV/VL3 (2.5 x10@infectious units). After incubation for 48 hn, the survivingcells were harvested and either used for the detection ofcytoplasmic viral antigens by indirect immunofluorescence orirradiated wth 5000 R and cocubtivated at various concentrations with a constant number of BL/RL12-NP cells. The percentage of CVA-positive BL/RL12-NP cells was scored by IF at3 days (5 generations) and 8 days (15 generations) aftercocultivation. Representative curves obtained with infectedthymocytes are shown in Chart 2. For thymocyte cultures, theend poInt, i.e., the smallestnumberof thymocytescontainingat least one infectious center 48 hr after infection by RadLV/VL3, @82 x 105 cells. Table 3 shows the values obtained forthe detection of CVA by immunofluorescence and by the BL/RL1@-NPcocultivationprocedure in several replicate expeniments 48 hr after RadLV/VL3 infection of thymus, marrow, andspleen cell cultures. The percentage of IF-positive cells wasalwayslessthan0.1 . It couldnotbe determinedmorepreciselybecause of the presence of cell debris and of nonspecificallyfluorescent cells (22). By using the BL/RL12-NP cocubtivationassay, it is clear that normal thymocytes, marrow, and spleen
U)-I@1
wC)a:04C)0z
>I-U)0a.U.
I-zwC)a:wa.
0.1i06
NUMBEROFTHYMOCYTES
Chart 2. Detection of infectious centers in thymocyte cultures 2 days afterInfection by RadLV/VL3. Results are expressed as the percentage of IF-positiveBL/RL12-NP indicator cells 5 (•)and 16 (0) cell doublings after cocultivatlonwith various concentrations of irradiated (5000 R) thymocytes.
a Percentage of CVA-positive cells detected by IF.
b The variations observed from one experiment to another can be attributed
to variations in the number of target cells in the various cell samples at time ofinfection (±5%).
CThe animals were treated with S mg of cortisone sodium succinate (SoluCortef; Upjohn Co., Kalamazoo, Mich.) 2 days prior to sacrifice.
d ND. not detectable.
cells contain similar numbers of target cells (1:10,000 to 1:200,000) for productive infection by RadLV/VL3, whereas only1 cell in 3 x 10°corticoresistant thymocytes is susceptible toproductive infection by this T+L+ virus.
The progeny virus released into the culture fluids of the IFpositive indicator cells at 1S doubling times after cocultivationwith these thymus, marrow, and spleen cultures was tested invivo (5) for thymotropic activity (S to 10 mice/point) and wasfound to be highly thymotropic in all cases. Thus, the virusreleased by the infectious centers present in these RadLV/
547MARCH1980
on March 11, 2019. © 1980 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

J. Boniver et a!.
VL3-infected cultures retains this biological property of theparental virus.
DISCUSSION
The isolation, identification, and characterization of targetcells for infection and transformation by RadLV has been ofgreat interest to laboratories studying the pathogenesis ofthymic lymphomas in C57BL/Ka mice. Previous experimentshave demonstrated that lymphoid cells isolated from the spleenand marrow of weanbings, the thymus of neonates, and thefetal liver of C57BL/Ka mice can be infected in vitro withRadLV and can then give rise to thymomas of donor originwhen inoculated into CS7BL x BALB/c F1 hosts immediatelyafter infection (16, 21). Other in vivo experiments have shownthat i.v. inoculation of RadLV into young adult C57BL/Ka miceresults in virus production by thymocytes present in the subcapsular region of the thymus (8). Such studies would begreatly facilitated by the availability of an entirely in vitro assaywhich would allow the cultivation of CS7BL lymphoid cells fromvarious tissues, infection of these cell populations by T+L+viral isolates from that strain, detection of the virus producercells in such cultures, and finally the isolation and charactenization of these target cells. The XC assay and the direct assayof viral antigens in bymphoidcultures by immunofluorescencehave been of little use for the detection and reliable quantitationof cells which have been productively infected among the totalcell population of cultivated lymphocytes. Indeed, it has beendemonstrated that virus-rebated antigens can appear in thecytoplasm of infected thymic lymphocytes concomitantly withcell death (22) and that virus-producing cells present in RadLVinduced lymphomas in vivo do not always induce plaque formation in the rat XC cell assay in vitro (17). The BL/RL12-NPcocultivation assay 48 hr after infection of the target lymphoidcell population by a T+L+ virus is thus the first quantitativeassay by which productive in vitro infection can be detected inthymus, bone marrow, and spleen cell populations maintainedin short-term culture.
The data clearly show that infection of the indicator cells isinitiated by virus-producing cells (infectious centers) whichhave undergone one or 2 divisions. Infection is not attributableto free virus surviving in the culture fluids or to virus particlespassively adsorbed on cell membranes 48 hr after virus inoculation. The assay can detect a single infectious center among102 to 106 bymphoid cells, provided that a ratio of infectiousparticles to total cell number greaten than 0.1 is used in theinfection procedure. It can thus be concluded that any singlecell producing AadLV-Iike virus in the culture will be detectedby the assay, as a consequence of the remarkable permissiveness of BL/AL12-NP cells for infection by AadLV (19).
In the experiments described in this report, infectivity hasbeen examined in thymus, spleen, and bone marrow culturesunder conditions in which many of the cells were rapidly dying.Cell growth parameters, which are better documented for thethymocyte cultures than for the bone marrow and spleen cubtures, show that DNA synthesis occurs only during the initial24 hr of a thymocyte culture. Since dividing thymocytes havean average doubling time of 10 to 12 hr (23), one can assumethat thymocytes could have undergone a maximum of 2 celldivisions during the cultivation period. Therefore, under optimalconditions, one could expect that each target cell for productive
infection by RadLV could give rise to a maximum of 4 progenycells after 24 hr in culture (at the end of the period of cellreplication), and thus to 4 infectious centers detectable by thecocubtivation assay performed 48 hr after the initiation of thecultures. However, because of the high level of cell death inthis culture system, it is likely that the number of infectiouscenters derived from each target cell ranges between 1 and 4at the time of the assay.
It would be more convenient to use an in vitro system inwhich the pattern of cell proliferation could be controlled andquantitated accurately. However, to our knowledge, there is noefficient way to maintain the proliferation of immature bymphoidpopulations in vitro. For example, cocultivation of thymocyteswith thymus neticuboepithelial cells does not significantly improve cell survival, although it induces thymocyte maturation(24).@On the other hand, mitogens and T-ceblgrowth factorsrapidly modify the differentiation patterns of prothymocytesand thymocytes in vitro (9, 14, 27) and thus select for matureT-ceblsubpopulations, which would not necessarily retain thesame susceptibility to infection by RadLV. Preliminary expeniments have shown that the use of such growth factors doesnot increase the number of infectious centers detectable inshort-term cultures of thymocytes after infection with RadLV.
At the present time, therefore, the in vitro system describedin this report seems to be the only available method for estimating the frequency of target cells permissive for productiveinfection by RadLV in lymphoid or hematopoietic cell popubations, although the kinetic limitations discussed above must bekept in mind. For example, if the true abundance of target cellsin a thymocyte culture were 1/3 x 1o@,then, considering theabove-cited estimate that such target cells could give rise tofrom 1 to 4 progeny cells at 48 hr, the cocubtivation assaymight yield frequency estimates ranging from between 1 infectious center/3 x 1O@to 1 infectious center/i .2 x 1O@cells. Itshould be noted that the experiments actually gave somewhatlower estimates (1 infectious center/2 x 1O@to 1/8 X I 0@).Ifsimilar calculations are applicable to normal bone marrowcultures, the numbers of bone marrow cells susceptible toAadLV infection in vitro would range between 1/1 x 1O@and1/4 x 10@ cells.
It is of interest that the estimates thus obtained from these invitro studies are in good agreement with previous in vivoobservations which indicated that 1 in 2 to 4 x 1O@thymuscells and 1 in 1.5 x 1O@to 2 x 10@bone marrow cells couldserve as target cells for RadLV in CS7BL/Ka mice (4, 16). Inlight of these small numbers of primary target cells for RadLVinfection, one can now readily explain the earlier observationthat it requires at beast4 days after RadLV inoculation into micebefore CVA-positive cells can be detected (7, 8). Indeed, theseestimates have been recently confirmed in vivo; using the BL/RL,2-NP cocubtivation assay, it was found that as few as 1 in1o@to 1 in 1O@thymocytes produce detectable virus 2 daysafter direct intrathymic injection of RadLV, in excellent agreement with the above cited in vitro and in vivo data.6 Conversely,these results are incompatible with a published claim by Haas
5 J. Boniver, A. Declève, M. Dailey, C. Honsik. M. Lieberman, and H. S.
Kaplan. Macrophage and lymphocyte-free thymus reticulo-epithelial cell cultures:establishment and functional influence on T-Iymphocyte maturation and transformation in vitro, submitted for publication.
6 J@Boniver, M. Travis, A. Declève, and H. S. Kaplan, unpublished results.
548 CANCERRESEARCHVOL. 40
on March 11, 2019. © 1980 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

Target cells for Infection by RadLV
factor-containing supernatants on the surface antigens of murine thymocytes. Cell. Immunol. 44: 252—261, 1979.
10. Haas, M., and Hilgers, J. In vitro infection of lymphoid cells by thymotropicradiation leukemia virus grown in vitro. Proc. NatI. Acad. Sci. U. S. A., 72:3546—3550,1975.
11. Hahn, G. N., Declève,A., Lieberman, M., and Kaplan, H. S. Focus formationby a murine sarcoma-leukemia virus complex. I. Theoretical analysis. J.Virol., 5: 432-436, 1970.
12. Haran-Ghera, N., Rubio, N., Leef, F., and Goldstein, G. Characteristics ofpreleukemia cells induced in mice. Cell. Immunol., 37: 308—314,1978.
13. Hopper, K., and Shortman, K. The differentiation of T lymphocytes. III. Thebehaviour of subpopulations of mousethymus cells in short-term cell culture.Cell. Immunol., 25: 256-273, 1976.
14. lrlé,C., Piguet, P. F., and Vassali, P. In vitro maturation of immaturethymocytes into immunocompetent T cells in the absence of direct thymicinfluence. J. Exp. Med., 148: 32—45,1978.
15. Jolicoeur, P., and Baltimore, D. Effect of Fv-i gene product on proviral DNAformation and integration in cells infected with murine leukemia viruses.Proc. NatI. Acad. Sci. U. S. A., 73: 2236-2240, i976.
i 6. Kaplan, H. S., and Lieberman, M. The role of lymphoid and haematopoietictarget cells in viral lymphomagenesis of CS7BL/Ka mice. II. Neoplastictransformation of bone marrow-derived cells in the thymic microenvironment.Blood Cells, 2: 301—317, 1976.
17. Lieberman, M., Declève,A., Finn, 0. J., Ricciardi-Castagnoli, P., Boniver,J., and Kaplan, H. S. Establishment, characterization, and virus expressionof cell lines from radiation- and from virus-induced lymphomas of CS7BL/Ka mice. mt. j. Cancer, 24: 168—177,1979.
18. Lieberman, M., Declève,A., IhIe, J. N., and Kaplan, H. S. Rescue of athymotropic, leukemogenic C-type virus from cultured non-producer lymphoma cells of strain C57BL/Ka mice. Virology, 97: 12—21, 1979.
i 9. Lieberman, M., Declève,A., and Kaplan, H. S. Rapid in vitro assay forradiation leukemia virus. Virology, 90: 274—278,1978.
20. Lieberman, M., and Kaplan, H. S. Leukemogenic activity of filtrates fromradiation-induced lymphoid tumors in mice. Science (Wash. D. C.), 130:387-388, 1959.
21. Lieberman, M., and Kaplan, H. S. Lymphoid tumor induction by mousethymocytes infected in vitro with radiation leukemia virus. NatI. Cancer Inst.Monogr., 22: 549-564, 1966.
22. Lonai, P., Declève,A., and Kaplan, H. S. Spontaneous induction of endogenous murine leukemia virus-related antigen expression during short-termin vitro incubation of mouse lymphocytes. Proc. NatI. Acad. Sci. U. S. A.,71: 2008—2012,1974.
23. Metcalf, D. The nature and regulation of lymphopoiesis In the normal andneoplastic thymus. In: G. Wolstenholme and R. Porter (eds.), Thymus:Experimental and Clinical Studies. Ciba Foundation Symposium, pp. 242—263. Baltimore: The Williams & Wilkins Co., 1966.
24. Mosier, D. E., and Pierce, C. W. Functional maturation of thymic lymphocytepopulations in vitro. J. Exp. Med., 136: 1485—1500, 1972.
25. Niwa, 0., Declève,A., Kojola, J., and Kaplan, H. S. Biological and serologicalcharacterization of the C-type RNA viruses isolated from the C57BL/Kastrain of mice. I. Influence of some chemicals and physical agents on thequantitation of murine viruses in vitro. In: J. F. Duplan (ed), Radiationinduced Leukemogenesis and Related Viruses, pp. 213—228.Amsterdam:Elsevier/North Holland Biomedical Press, 1977.
26. Niwa, 0., Declève,A., Lieberman, M., and Kaplan, H. S. Adaptation ofplaque assay methods to the in vitro quantitation of the radiation leukemiavirus. J. Virol., 12: 68—73,1973.
27. Gillis, S., Union, N. A., Baker, P. E., and Smith, K. A. The in vitro generationand sustained culture of nude mouse cytolytic T lymphocytes. J. Exp. Med.,149: i460-1476, 1979.
28. Sveda, M. M., and Soeiro, R. Host restriction of Friend leukemia virus:synthesis and integration of the provirus. Proc. Nati. Acad. Sci. U. S. A., 73:2356-2360, 1976.
29. Toyoshima, K., and Vogt, P. Enhancement and inhibition of avian sarcomaviruses by polyanions and polycations. Virology, 38: 4 14—426,i 969.
30. Vogt, P. K. DEAE-dextran: enhancement of cellular transformation inducedby avian sarcoma virus. Virology, 33: 175—i77, i 967.
549
and Hibgers that 80% of CS7BL thymocytes are productivelyinfected by RadLV after 48 hr in culture (10). This finding,which has not been confirmed in our laboratory (19), is inaddition not supported by the growth kinetics of thymic bymphocytes in culture (Ref. 13; this paper) onby studies on the invivo replication of RadLV in CS7BL mice (4, 5, 7, 8).
The numbers of infectious centers detected by the BL/RL12-NP cocultivation assay in thymus, bone marrow, and spleencultures are compatible with the hypothesis that the normaltarget cell for infection and possibly for transformation byRadLV belongs to a restricted subpopulation which might be,as suggested by previous in vivo studies, a bone marrow orspleen prothymocyte (12, 16) or a subset of the subcapsularblast cell population in the thymus (8). The present availabilityof the assay described in this report allows one to test thishypothesis by using techniques of enrichment for variouslymphoid cell populations from marrow, spleen, on thymusbefore in vitro infection with RadLV/VL3. Also, it will be possibleto analyze the interaction of RadLV with non-T-cebbpopulations.These experiments, which are now in progress in our laboratony, will hopefully provide us with enriched target cell populations for RadLV infection; these populations could then betested for neoplastic transformation either in vivo, using thebioassay described by Liebenman and Kaplan (16, 21), or invitro by cultivation in the presence of thymus reticuboepithebialcells and/or growth factors.
REFERENCES
1. Declève,A., Lieberman, M., Hahn, G., and Kaplan, H. S. Focus formation bya murine sarcoma-leukemia virus complex. II. Quantitative aspects of theinteraction between radiation leukemia virus and its murine sarcoma viruspseudotype In strain CS7BL mouse embryo cells. J. Virol., 5: 437-445,1970.
2. Declève,A., Lleberman, M., IhIe, J. N., and Kaplan, H. S. Biological andserological characterization of radiation leukemia virus (RadLV). Proc. NatI.Aced. Sd. U. S. A., 73: 4675-4679, 1976.
3. Declève,A., Lieberman, M., lhle, J. N., Rosenthal, P. N., Lung, M. L., andKaplan, H. S. Physicochemical, biological, and serological properties of aleukemogenic virus Isolated from cultured RadLV-induced lymphomas ofC57BL/Ka mice. Virology. 90: 23-35, 1978.
4. Declève.A., Ueberman, M., and Kaplan, H. S. In vivo interaction betweenRNA viruses Isolated from the C57BL/Ka strain of mice. Virology, 8 1: 270-283. 1977.
5. Declève.A., Ueberman, M., Niwa, 0.. and Kaplan, H. S. Rapid in vivo assaytechnique for murine leukemia viruses. Nature (Lond.), 252, 79—81, 1974.
6. Declève,A., Niwa, 0., Hilgers, J., and Kaplan, H. S. An improved MuLVImmunofluorescence assay. Virology, 57: 491—502,1974.
7. Declève,A., Sato, C.. Lieberman, M., and Kaplan, H. S. Selective thymiclocalization of munne leukemia-virus related antigens in C57BL/Ka miceafter inoculation with radiation leukemia virus. Proc. NatI. Acad. Sci. U. S.A.. 71: 3124—3128,1974.
8. Declève.A.. Travis, M., Weissman, I. L., Lieberman, M., and Kaplan, H. S.Focal infection and transformation in vivo of thymic cell subclasses by athymotropic murine leukemia virus. Cancer Res., 35: 3585—3595,1975.
9. DiSabato, G., Moraes. E. K., and Alton, M. Effect of thymocyte stimulating
MARCH 1980
on March 11, 2019. © 1980 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

1980;40:544-549. Cancer Res Jacques Boniver, Alain Declève, Olivera J. Finn, et al.
by the Radiation Leukemia Virusin VitroPopulations Infected Detection of Infectious Centers in C57BL/Ka Lymphoid Cell
Updated version
http://cancerres.aacrjournals.org/content/40/3/544
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/40/3/544To request permission to re-use all or part of this article, use this link
on March 11, 2019. © 1980 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from