dengue and other viral hemorrhagic fevers in...
TRANSCRIPT

REVIEW
Dengue and Other Viral Hemorrhagic Fevers in India
Shally Awasthi • U. C. Chaturvedi
Received: 16 September 2011 / Accepted: 14 November 2011 / Published online: 18 January 2012
� The National Academy of Sciences, India 2012
Abstract The important viral hemorrhagic fevers (VHF)
in India spread by an arthropod vectors are Dengue fever/
Dengue haemorrhagic fever and Kyasanur forest disease.
Another VHF is caused by Hanta virus infection and for this
there is no known vector. The diagnosis of VHF is based on
typical clinical presentation and thereafter confirmed by
detection of either virus or viral RNA or by demonstration of
a rise of antibody titres against it. Treatment is by and large
symptomatic as there are no specific drugs available against
the viruses. Dengue fever has acquired epidemic magnitude
in certain parts of India and has about 10% mortality.
Research is going on for the development of effective vac-
cines against Dengue fever. The principles of primary pre-
vention of vector borne diseases is through vector control
measures, elimination of breeding sites in and around human
dwellings and personal protection against vector bite.
Keywords Hemorrhagic fevers � Dengue �Kyasanur forest disease � Virion � Hantaviruses
Introduction
Viral hemorrhagic fevers (VHF) are a distinct group of acute
viral infections which result in severe multisystem syndrome
due to wide spread damage to the vascular system. This
results in varying degree of hemorrhage, including con-
junctivitis, petechia, ecchymosis and shock, sometimes
leading to death. Hemorrhagic fever viruses belong to four
taxonomic families (Table 1), all being single-stranded
RNA viruses and possess a lipid envelope [1].
Flaviviridae
Dengue virus, Yellow fever virus, Omsk hemorrhagic fever
virus, Kyasanur forest disease virus.
Epidemiology/Geographic Distribution
Flaviviridae include Alkhurma HF virus, Kyasanur forest
disease, and Omsk HF. Alkhurma HF virus is a variant of
Kyasanur forest disease virus found in Saudi Arabia and
reported in a small number of patients since 1990s [2].
Yellow fever virus is found throughout sub-Saharan Africa
and tropical South America but its activity is intermittent and
localized [3]. The annual incidence is believed to be about
200,000 cases per year globally. Case fatality rate ranges
greatly depending on the epidemic but may reach up to 50%
in severe yellow fever cases. Dengue virus is found
throughout the tropical Americas, Africa, Australia, and
Asia. Case fatality rate for DHF is generally low 1–10%
depending on available treatment [4]. Kyasanur forest virus
is confined to Mysore state of India but spreading. Case
fatality rate is 3–5%. Omsk Hemorrhagic fever virus is still
isolated to the Omsk and Novosibirsk regions of the former
Soviet Union. Case fatality is 0.5–3%. The South American
HF has a case-infection ratio of more than 50% of those
exposed. The mortality rate is 15–30% (Table 1)1 [3, 4].
S. Awasthi (&)
Department of Pediatrics, CSM Medical University,
Lucknow, India
e-mail: [email protected]
U. C. Chaturvedi
201-Annapurna Apartments, No. 1, Bishop Rocky Street,
Faizabad Road, Lucknow 226 007, India
e-mail: [email protected] 1 http://en.wikipedia.org/wiki/Flaviviridae.
123
Proc. Natl. Acad. Sci. Sect B. Biol. Sci. (January–March 2012) 82(1):69–80
DOI 10.1007/s40011-011-0006-9

Virion Structure
The virion of the Flaviviridae is spherical to pleomorphic
and the virions are enveloped. The virions are measured
40–50 nm in diameter. An envelope and a nucleocapsid are
present in the virion of Flaviviridae. The capsid is envel-
oped with glycoprotein peplomers. The surface projections
of the capsids are made up of small spikes that are sur-
rounded by a prominent fringe. The capsid has a polyhedral
symmetry and it is round. The capsid has an inner core
protein. The core is measured 20–30 nm in diameter, and it
is isometric2 [3–5] (Fig. 1).
Genome Structure
The genome of Flaviviridae consists of a linear, positive-
sense, single stranded RNA of about 11 kb in length, and
the genome is not segmented. The genome has a 50-end
which carries a methylated nucleotide cap or genome-
linked protein. The genome of Dengue virus consists of a
single stranded, non segmented, positive sense ribonucleic
acid (RNA). The genome is translated into a single poly-
peptide which is co- and post-translationally processed by
host signalases as well as the virus encoded serine protease
into the three structural (C, prM/M, and E) and seven non-
structural proteins (NS) in the order C–prM–E–NS1–
NS2A–NS2B–NS3–NS4A–NS4BNS5 that traverse the
endoplasmic reticulum membrane. The three structural
proteins, C–M–E, are located at the amino terminus, while
NS NS–NS2a, NS2b–NS3–NS4a, NS4b, and NS5 are at the
carboxy terminus [3–5].3
Replication
Replication of Flavivirus takes place in the cytoplasm.
Flavivirus cannot replicate in the nucleus because, like
Table 1 Viruses causing haemorrhagic fevers
Family/Viruses Diseases Transmission
to humans by
Incubation
period (days)
Geographical distribution
Flaviviridae
Yellow fever virus Yellow fever Mosquito 3–6 Tropical Africa, South America
DEN Dengue HF Mosquito 3–6 Asia, Americas, Africa
KFD virus KFD Ticks 2–7 India
Bunyaviridae
Phlebovirus Rift valley fever Mosquito 2–5 Africa
Nairovirus Crimean-congo HF Tick 3–12 Europe, Asia, Africa
Hantavirus HF with renal syndrome,
hantavirus pulmonary
syndrome
Rodent 9–35 Asia, Europe, worldwide
Arenaviridae
Lassa fever virus Lassa fever Rodent 5–16 Africa
Junin virus Argentine HF Rodent 7–14 South America
Machupo virus Bolivian HF Rodent 9–15 South America
Sabia virus Brazilian HF Rodent 7–14 South America
Guanarito virus Venezuelan HF Rodent 7–14 South America
Filoviridae
Marburg virus Marburg Unknown 3–16 Africa
Ebola virus Ebola Africa
Fig. 1 Virion structure of Flaviviridae [6]
2 http://en.wikipedia.org/wiki/Flaviviridae. 3 http://en.wikipedia.org/wiki/Flaviviridae.
70 Proc. Natl. Acad. Sci. Sect B. Biol. Sci. (January–March 2012) 82(1):69–80
123

most other RNA viruses, it uses the host cell’s RNA
dependent RNA polymerase to replicate. The genome
mimics the cellular mRNA molecule in all aspects except
for the absence of the poly-adenylated (poly-A) tail. This
feature allows the virus to exploit cellular apparatus to
synthesise both structural and NS, during replication. The
cellular ribosome is crucial to the replication of the flavi-
virus, as it translates the RNA, in a similar fashion to
cellular mRNA, resulting in the synthesis of a single
polyprotein. Once translated, the polyprotein is cleaved by
a combination of viral and host proteases to release mature
polypeptide products. Nevertheless, cellular post-transla-
tional modification is dependent on the presence of a poly-
A tail; therefore this process is not host-dependent. Instead,
the polyprotein contains an autocatalytic feature which
automatically releases the first peptide, a virus specific
enzyme. This enzyme is then able to cleave the remaining
polyprotein into the individual products. One of the prod-
ucts cleaved is a polymerase, responsible for the synthesis
of a (-) sense RNA molecule. Consequently this molecule
acts as the template for the synthesis of the genomic
progeny RNA. New viral particles are subsequently
assembled. This occurs during the budding process which
is also responsible for the accumulation of the envelope
and cell lysis [3–5] (Fig. 2).4
Bunyaviridae
Rift valley fever virus, Crimean-Congo hemorrhagic fever
virus, Hantavirus.
Epidemiology/Geographic Distribution
Bunyaviruses are seen throughout Africa, the Middle East,
the Balkans, southern Russia, and western China. RVF is
found primarily in sub-Saharan Africa and was recently
isolated in Saudi Arabia and Yemen in 2000. The case
fatality rate in humans is generally around 1%. CCHF is
found in most of sub-Saharan Africa, Eastern Europe and
Asia. The case fatality rate is 30% and nosoicomial out-
breaks have been documented through exposure to infected
blood products. Approximately 1% of individuals exposed
to RVF virus become infected, but the mortality rate of
persons infected is 50%. CCHF has an infection rate of
20–100% and a fatality rate of 15–30%. Hantaviruses are
divided into two groups based on location: Old world
viruses are found in eastern Europe and eastern Asia while
new world viruses are found in North and South America
[3, 4] (Table 1).5
Virion Structure
Bunyavirus virions have a complex structure. They consist
of an envelope and three nucleocapsids. Virions are
80–120 nm in diameter. The virions vary in shape from
spherical to pleomorphic. Surface projections are 5–10 nm
long spikes which evenly coat the surface. The nucleo-
capsids are elongated and have helical symmetry
[1, 5–7]6,7,8 (Fig. 3).
Genome Structure
The bunyavirus genome is monomeric and consists of three
segments (large (L), medium (M) and small (S)). Besides,
they exhibit a pseudo-circular structure due to each seg-
ment’s complementary ends. The L segment encodes the
RNA dependent RNA-polymerase, necessary for viral
RNA replication and mRNA synthesis. The M segment
encodes the two envelope glycoproteins which project
Fig. 3 Virion structure of Bunyavirus [49]
Fig. 2 Replication cycle of Flavivirus [7]
4 http://en.wikipedia.org/wiki/Flaviviridae.
5 http://en.wikipedia.org/wiki/Bunyaviridae6 http://en.wikipedia.org/wiki/Bunyaviridae.7 http://www.virology.net/Big_Virology/BVRNAbunya.html.8 http://microbewiki.kenyon.edu/index.php/Bunyaviridae.
Proc. Natl. Acad. Sci. Sect B. Biol. Sci. (January–March 2012) 82(1):69–80 71
123

from the viral surface and aid the virus in attaching to and
entering the host cell. The S segment encodes the nucleo-
capsid protein (N). The L and M segment are negative
sense. These RNA segments are single-stranded, and exist
in a helical formation within the virion. The entire genome
is about 10,500–22,700 nucleotides long [1, 5–7].9,10,11
Replication
Replication begins with the attachment of viral proteins to
host receptors and entry of virus by endocytosis. Acidifi-
cation of endocytic vesicles causes uncoating of the virus
which is followed by fusion of viral membranes with that
of the host endosomes. Primary transcription of viral
complementary mRNA occurs and the L, M and S mRNAs
are translated. This is followed by RNA replication and
morphogenesis. Mature virions are released by fusion of
virus-containing cytoplasmic vesicles with the plasma
membrane and budding [1, 5–7]12,13,14 (Fig. 4).
In India, cases of VHF belong to the family of Flavi-
viridae, which includes Dengue virus and Kyasanur forest
disease virus. There have been recent reports of emerging
public health threats due to Hantavirus, belonging to the
family Bunyaviridae. Studies have been done in India to
look for Nairovirus which causes Crimean-Congo haem-
orrhagic fever but no conclusive evidence is found; neither
in human sera nor in the vector. Table 2 below summarizes
the reservoirs and modes of transmission of hemorrhagic
fever viruses reported from India [1, 3] (Table 3).
Dengue and Dengue Hemorrhagic Fever
Dengue has been recognized since the latter part of the 18th
century as causing epidemics in tropical and subtropical
regions globally. Some 2,500 million (2.5 billion) people
are estimated to currently be at risk of dengue in over 100
countries across the globe. It is estimated that between 50
and 100 million cases of dengue fever, 500,000 cases of
dengue hemorrhagic fever/dengue shock syndrome (DHF/
DSS) and more than 20,000 deaths from DHF/DSS occur
each year [3, 4]. Dengue virus (DEN) is a small single-
stranded RNA virus comprising four distinct serotypes
(DEN-1 to -4). These closely related serotypes of the DEN
belong to the genus Flavivirus, family Flaviviridae [8].
Humans are the only vertebrate hosts of the virus. There
is a jungle cycle between monkeys and mosquitoes, but this
plays no role in human disease (Fig. 5).
The virus is transmitted when a mosquito of the genus
Aedes bites an individual infected with DEN. The virus in
the blood of the infected individual then infects the mos-
quito and travels from the mosquito’s stomach to its sali-
vary glands where the virus multiplies. The virus is then
injected into another person, when the mosquito injects
anticoagulants that prevent blood clotting when the mos-
quito is feeding. The mosquito remains capable to transmit
dengue for its entire life. There is an enzootic dengue
transmission cycle in the forest involving Aedes mosqui-
toes and lower primates in Africa and Asia [9] (Fig. 6).
Aedes aegypti breeds in fresh water and particularly in
manmade containers such as old tyres, pot plant holders,
buckets and tree hollows in urban areas. Aedes albopictus
is a mosquito common in South East Asia and Papua New
Guinea and can also be an important vector. Other Aedes
species are involved in the enzootic monkey cycle. There is
no evidence of person to person transmission [4].
Dengue fever (break bone fever) classically presents as
an acute febrile illness. The incubation period is usually
short but varies from three to 14 days. It is extremely
debilitating with fever lasting 3–5 days, myalgia (particu-
larly backache), arthralgia, retro-orbital pain, anorexia,
gastrointestinal disturbance, rash and increased vascular
permeability. There is a high subclinical rate of milder
disease in children compared to adults and a low fatality
rate. Recovery from infection with one serotype of the
DEN results in homologous immunity but does not provide
protection against infection with other serotypes [3].15
DHF is a severe complication of DEN infection. It
occurs mainly in children and is characterized by abrupt
onset of fever, haemorrhagic phenomena and thrombocy-
topaenia. In its severest form it may result in shock (DSS),
Fig. 4 Replication cycle of Bunya virus (http://www.access
excellence.org/RC/VL/GG/influenza.php)
9 http://en.wikipedia.org/wiki/Bunyaviridae.10 http://www.virology.net/Big_Virology/BVRNAbunya.html.11 http://microbewiki.kenyon.edu/index.php/Bunyaviridae.12 http://en.wikipedia.org/wiki/Bunyaviridae.13 http://www.virology.net/Big_Virology/BVRNAbunya.html.14 http://microbewiki.kenyon.edu/index.php/Bunyaviridae.
15 http://ci.vbi.vt.edu:13001/pathinfo/pathogens/Dengue_virus_Info.
html.
72 Proc. Natl. Acad. Sci. Sect B. Biol. Sci. (January–March 2012) 82(1):69–80
123

which has a high fatality rate. The rate of death from DHF
without DSS is usually quoted at 1–5%. This is believed to
be caused by immune enhancement when a person with
dengue antibodies due to a previous infection is subse-
quently infected by a DEN of a different serotype [1–4].
Pathogenesis
After an incubation period of 4–10 days, infection by any of
the four virus serotypes can produce a wide spectrum of
illness, although most infections are asymptomatic or sub
clinical. Primary infection is thought to induce lifelong
protective immunity against the infecting serotype [10].
Individuals suffering an infection are protected from clinical
illness with a different serotype within 2–3 months of the
primary infection but there is no long-term cross-protective
immunity [8]. The DEN enters via the skin while an infected
mosquito is taking a blood meal. During the acute phase of
illness the virus is present in the blood and its clearance from
this compartment generally coincides with defervescence.
Humoral and cellular immune responses are considered to
contribute to virus clearance by the generation of neutraliz-
ing antibodies and the activation of CD4? and CD8? T
lymphocytes. In addition, innate host defense may limit
infection by the virus. After infection, serotype specific and
cross-reactive antibodies and CD4? and CD8? T cells
remain measurable for years. Plasma leakage, haemocon-
centration and abnormalities in homeostasis characterize
severe dengue. The mechanisms leading to severe illness are
not well defined. Individual risk factors determine the
severity of disease and include secondary infection, age,
ethnicity and possibly chronic diseases (bronchial asthma,
sickle cell anaemia and diabetes mellitus) [8].
Seroepidemiological studies in Cuba and Thailand con-
sistently support the role of secondary heterotypic infection
as a risk factor for severe dengue, although there are a few
reports of severe cases associated with primary infection
[10–14]. Antibody-dependent enhancement of infection has
been hypothesized [15, 16] as a mechanism to explain severe
dengue in the course of a secondary infection and in infants
with primary infections. In this model, non-neutralizing,
cross-reactive antibodies raised during a primary infection,
or acquired passively at birth, bind to epitopes on the surface
of a heterologous infecting virus and facilitate virus entry
into Fc-receptor-bearing cells. The increased number of
infected cells is predicted to result in a higher viral burden
and induction of a robust host immune response that includes
inflammatory cytokines and mediators, some of which may
contribute to capillary leakage [17–19]. During a secondary
infection, cross-reactive memory T cells are also rapidly
activated; these proliferate, express cytokines and die by
apoptosis in a manner that generally correlates with overall
disease severity [20, 21]. Host genetic determinants might
influence the clinical outcome of infection [22], though most
studies have been unable to adequately address this issue.
Laboratory evidence requires one of the following:
• Isolation of DEN from clinical material
• Detection of dengue viral RNA in clinical material
• Detection of antibodies against dengue NS1 protein
• Detection of DEN specific IgM in the serum
• A significant rise in the level of DEN specific IgG
Preventive Measures
Vaccines
The first dengue vaccine was prepared in 1945 and still no
effective vaccine is available that indicates problems in its
development. An effective dengue vaccine is a distinct
possibility because the virus causes an acute infection and
the viraemia is removed within 5 days; immunity to infec-
tion with homologous DV serotype is long lasting; and
passive transfer of virus specific antibodies are protective
against subsequent challenge with that specific viral subtype
in animal models. The major obstacles in the development of
an effective dengue vaccine are incomplete understanding
of the pathogenesis of DHF; absence of an animal model of
DHF; and pre-existing heterotypic dengue antibodies, which
are a risk factor for DHF. An effective vaccine will have to be
Table 2 Reservoirs/vectors/modes of transmission reservoirs and modes of transmission for selected hemorrhagic fever viruses found in India
Agent Reservoir Arthropod vector Modes of transmission
Dengue Humans Aedes aegypti Bite of infected mosquito
KFD virus Rodents, bats, and other small mammals;
monkeys (e. g., black-faced langur,
South Indian bonnet macaque) appear
to be amplifying hosts
Ixodid Ticks (Haemaphysalisspinigera)
Bite of infected tick
Airborne through virus-containing
aerosols in laboratory setting
Hanta virus Murid Rodents with horizontal rodent
to rodent transmission
Absent Airborne transmission from
aerosol generated from
contaminated urine, feces and
saliva of infected rodent.
Proc. Natl. Acad. Sci. Sect B. Biol. Sci. (January–March 2012) 82(1):69–80 73
123
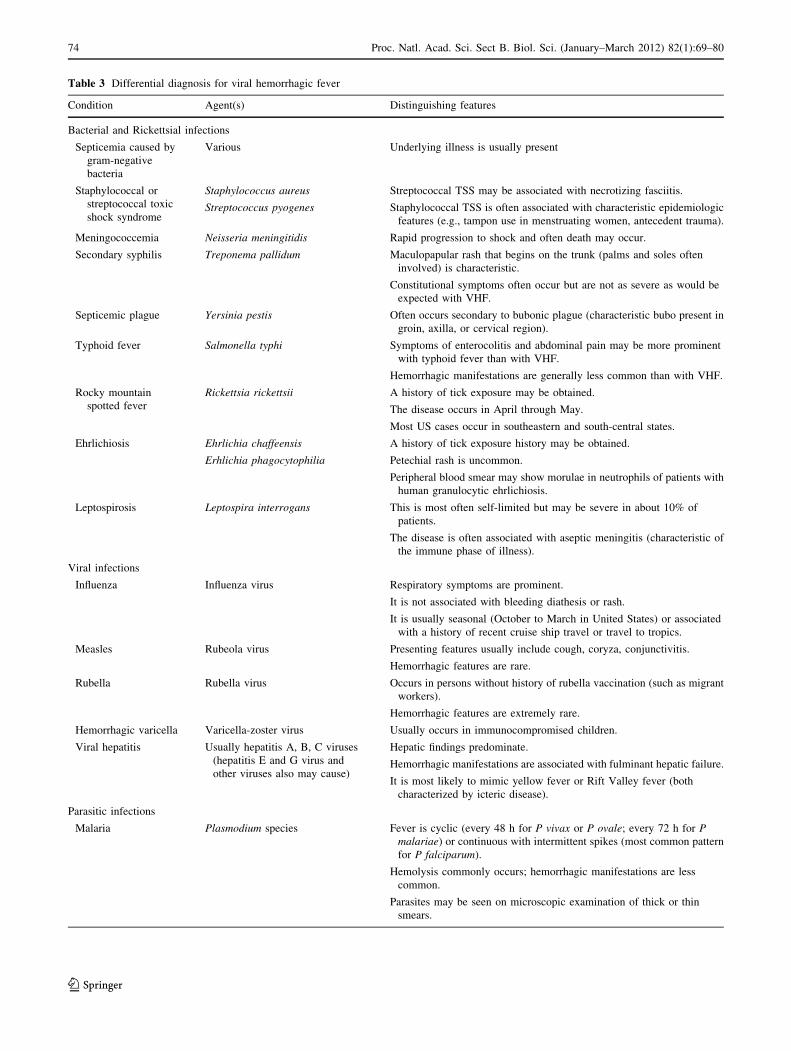
Table 3 Differential diagnosis for viral hemorrhagic fever
Condition Agent(s) Distinguishing features
Bacterial and Rickettsial infections
Septicemia caused by
gram-negative
bacteria
Various Underlying illness is usually present
Staphylococcal or
streptococcal toxic
shock syndrome
Staphylococcus aureus Streptococcal TSS may be associated with necrotizing fasciitis.
Streptococcus pyogenes Staphylococcal TSS is often associated with characteristic epidemiologic
features (e.g., tampon use in menstruating women, antecedent trauma).
Meningococcemia Neisseria meningitidis Rapid progression to shock and often death may occur.
Secondary syphilis Treponema pallidum Maculopapular rash that begins on the trunk (palms and soles often
involved) is characteristic.
Constitutional symptoms often occur but are not as severe as would be
expected with VHF.
Septicemic plague Yersinia pestis Often occurs secondary to bubonic plague (characteristic bubo present in
groin, axilla, or cervical region).
Typhoid fever Salmonella typhi Symptoms of enterocolitis and abdominal pain may be more prominent
with typhoid fever than with VHF.
Hemorrhagic manifestations are generally less common than with VHF.
Rocky mountain
spotted fever
Rickettsia rickettsii A history of tick exposure may be obtained.
The disease occurs in April through May.
Most US cases occur in southeastern and south-central states.
Ehrlichiosis Ehrlichia chaffeensis A history of tick exposure history may be obtained.
Erhlichia phagocytophilia Petechial rash is uncommon.
Peripheral blood smear may show morulae in neutrophils of patients with
human granulocytic ehrlichiosis.
Leptospirosis Leptospira interrogans This is most often self-limited but may be severe in about 10% of
patients.
The disease is often associated with aseptic meningitis (characteristic of
the immune phase of illness).
Viral infections
Influenza Influenza virus Respiratory symptoms are prominent.
It is not associated with bleeding diathesis or rash.
It is usually seasonal (October to March in United States) or associated
with a history of recent cruise ship travel or travel to tropics.
Measles Rubeola virus Presenting features usually include cough, coryza, conjunctivitis.
Hemorrhagic features are rare.
Rubella Rubella virus Occurs in persons without history of rubella vaccination (such as migrant
workers).
Hemorrhagic features are extremely rare.
Hemorrhagic varicella Varicella-zoster virus Usually occurs in immunocompromised children.
Viral hepatitis Usually hepatitis A, B, C viruses
(hepatitis E and G virus and
other viruses also may cause)
Hepatic findings predominate.
Hemorrhagic manifestations are associated with fulminant hepatic failure.
It is most likely to mimic yellow fever or Rift Valley fever (both
characterized by icteric disease).
Parasitic infections
Malaria Plasmodium species Fever is cyclic (every 48 h for P vivax or P ovale; every 72 h for Pmalariae) or continuous with intermittent spikes (most common pattern
for P falciparum).
Hemolysis commonly occurs; hemorrhagic manifestations are less
common.
Parasites may be seen on microscopic examination of thick or thin
smears.
74 Proc. Natl. Acad. Sci. Sect B. Biol. Sci. (January–March 2012) 82(1):69–80
123

tetravalent and should induce immunity to all four DV ser-
otypes simultaneously. Selection of the most promising DV
vaccine candidates rely mostly on comparing vaccine-
induced immune responses to a profile of protective immu-
nity developed from natural DV infections [23].
The dengue vaccine research focuses on the use of live
attenuated or inactivated vaccines, infectious clone-derived
vaccines, immunogens vectored by various recombinant
systems, subunit immunogens, and nucleic acid vaccines.
Serial passages of dengue viruses in PDK cells were used to
develop live attenuated vaccine. Initially, monovalent
vaccines were prepared using each of the DEN serotypes
followed by di-, tri- and tetra-valent vaccines that were
immunogenic and well tolerated by human subjects. A
chimeric YF-dengue type 2 virus vaccine was prepared,
using a recombinant cDNA infectious clone of a Yellow
fever vaccine strain (YF17D) as a backbone, into which the
PrM and envelope E genes of DEN were inserted. This
vaccine induced neutralizing antibodies in monkeys and
protected them against challenge with a wild type of DEN.
The safety as well as protective efficacy of the recombinant
DV tetravalent vaccine has been demonstrated in a monkey
challenge model. A candidate DNA vaccine expressing DEN
PrM and E proteins was developed and used for the immu-
nization of monkeys. This vaccine induces virus-neutraliz-
ing antibodies and gave partial protection against challenge
with homologous DEN. DEN grown in Vero cells was used
for the immunization of laboratory animals after inactiva-
tion, purification and concentration. The vaccine induces the
production of protective level of antibodies in monkeys.
Recombinant DNA techniques have provided the possibility
of cloning specific genes encoding protective antigens and
expressing them in other host cells, including E. coli, yeast
and insect cell systems [24].16
The question of whether any of the candidate dengue
fever vaccines will be ready by 2012 still is an open
question, although it is the declared objective of both GSK
and Sanofi Pasteur, both of which have presented timelines
compatible with licensure by 2012.17
Public Health Measures
Dengue infection can be prevented by:
• Mosquito control measures
• Personal protection measures such as long sleeves and
mosquito repellents
• Avoidance of mosquito-prone areas.
Control of Case
Isolate the patient and prevent mosquito access until fever
subsides.
Investigate the source of infection.
Control of environment
• Search for and eliminate breeding sites of Aedes ae-
gypti in the urban area.
Table 3 continued
Condition Agent(s) Distinguishing features
Acute conditions that may be associated with a bleeding diathesis
Hemolytic uremic
syndrome
Usually occurs as complication of
infection with Escherichia coliO157:H7 or other Shiga toxin-
producing E. coli
Disease involves a triad of renal involvement, thrombocytopenia, and
hemolytic anemia.
It is more common in young children.
Antecedent diarrheal illness occurs.
Hemorrhagic manifestations are uncommon, although bloody diarrhea
often occurs
Thrombotic
thrombocytopenic
purpura
May occur as complication of
infection with E. coli O157:H7
or other Shiga toxin–producing
E. coli, although may be
noninfectious
Disease includes renal involvement, thrombocytopenia, hemolytic
anemia, neurologic involvement
Hemorrhagic manifestations are uncommon
Idiopathic
thrombocytopenic
purpura
Noninfectious Low platelet count is predominant feature
Disease is generally not accompanied by severe systemic toxicity
Acute leukemia Noninfectious Peripheral blood smear shows characteristic features of leukemia.
Collagen vascular
disease
Noninfectious Acute onset of febrile illness not likely
TSS toxic shock syndrome
16 http://www.who.int/vaccine_research/diseases/vector/en/index1.html.17 http://www.who.int/vaccine_research/diseases/vector/en/index1.html.
Proc. Natl. Acad. Sci. Sect B. Biol. Sci. (January–March 2012) 82(1):69–80 75
123

• Use mosquito repellents, mosquito nets and other
methods of personal protection.
• Control Aedes aegypti near airports.
• Prevent importation of new vectors, for example Aedes
albopictus.
Individual protective measures to avoid mosquito bites
during the day should include:
• Application of insect repellents (e.g. DEET-based repel-
lent, etc.) on exposed skin: the type and concentration of
repellents depend on age and presence of pregnancy;
• Wearing appropriate clothing (long sleeves and trousers);
• The use of a mosquito net (preferably insecticide-
treated), particularly recommended for children and
viraemic people.
Infected persons should avoid mosquito bites during the
first few days of illness to prevent further dissemination of
the virus.
Local Community Measures
The vector mosquitoes mainly colonize small water col-
lections, such as flower plates and pots, barrels, used tyres,
and sometimes tree-holes and rock pools. Therefore, the
removal of these breeding sites by emptying stagnant water
around the houses is the most efficient way to avoid
mosquito proliferation.
Key Areas of Uncertainty
Key areas of uncertainty include: the lack of vaccines and
antiviral drugs, the resistance of the vector mosquitoes to
insecticides, the role and impact of social mobilisation for
control of mosquitoes, the expansion of the geographic
distribution of potential Aedes vectors, the importance of
increasing international travel, and virus circulation
worldwide [25, 26].
Kyasanur Forest Disease Virus
In 1957, several dead monkeys were noticed in the
Kyasanur forest in Shimoga district in Karnataka along
with a severe prostrating illness in some of the villagers in
the area. A similar illness has been observed in the locality
a year earlier also. A new arbovirus, antigenically, related
to the Russian spring-summer encephalitis complex of
viruses was isolated by investigators from the Virus
Research Centre, (now the National Institute of Virology)
Poona, from patients and dead monkeys. It was named the
Kyasanur forest disease (KFD) virus after the name of the
place from where the first isolations were made [3, 27, 28].
KFD a tick-borne infection, is relatively rare and found
only in one region in the southwestern part of India. KFD
virus was first recognized in 1957 when it was isolated
from a sick monkey from the Kyasanur forest in Karnataka
State, India. Rodents, monkeys, bats, and other small
mammals are the natural reservoirs. Larger animals (such
as goats, cows, and sheep) may become infected, but they
don’t have a role in transmission of the disease. Natural
infections have been identified only in several districts
of Karnataka State. Outbreaks occur periodically and are
usually signaled by epizootics in the local monkey
population [29].
Continuing deaths in monkeys and an average of
400–500 human cases have been seen annually over the
past 5 decades, commonly occurring in evergreen, semi-
evergreen, and neighboring, moist, deciduous forest areas.
An array of tick species, mainly Haemaphysalis spinigera,
acts as vector for KFDV. This species of tick is widely
Fig. 5 Enzootic transmission
cycle
Fig. 6 Life cycle of dengue [10]. Extrinsic incubation period:
incubation occurring in the vector i.e. mosquito. Intrinsic incubation
period: incubation occurring in human
76 Proc. Natl. Acad. Sci. Sect B. Biol. Sci. (January–March 2012) 82(1):69–80
123

distributed in tropical evergreen and deciduous forests of
southern and central India and Sri Lanka. KFDV has also
been isolated from 7 other species of this genus and from
Dermacentor and Ixodes ticks. This disease is transmitted
by ticks among ground birds and small mammals such as
the white-tailed rat, white-bellied rat, shrew, and bat. High
titers of virus can be obtained after experimental infection
of black-napped hares, porcupines, flying squirrels,
Malabar giant squirrels, three-striped squirrels, gerbils,
mice, long-tailed tree mice, and shrews [30].
A seasonal pattern has been noted, with most of the
cases occurring in the spring months. Outbreaks of the
disease have occurred in the area periodically since it was
first identified, but it has spread only for a few kilometers
from the original site in all these years. Human cases were
also found among persons who visited forests to collect
firewood, grass, and other forest products.
KFD has an incubation period of about 3–8 days,
manifests with sudden onset of fever with chills, headache,
conjunctivitis, myalgia and severe prostration. The disease
duration is of about 5–12 days. Some cases develop hem-
orrhages into the skin, mucosa and viscera. The major
clinical manifestations are VHF or meningo encephalitis
and case fatality rate is [30% [27–30]. The National
Institute of Virology has produced an effective killed
vaccine against KFD.
Hantaviruses
Hantaviruses causes VHF with renal syndrome (HFRS) and
hantavirus cardiopulmonary syndrome (HCPS). These are
emerging as a global cause of concern and are increasingly
being reported from India. The first Hantavirus to be cul-
tured was Thottapalayam virus from Vellore in 1964.
Serological investigation of patients with pyrexic illness
reveals anti-hantavirus IgM in about 14.7% cases. Sero-
positivity of Hantavirus in the general population is 4%
while it is much higher in persons coming in contact with
the rodents. Since there is no arthropod vector, there is
slow movement of Hantavirus species with natural rodent
to rodent transmission, which is also a rate limiting factor
to wide spreading of the infection [31, 32].
Clinical picture and severity of HFRS and HCPS
depends on the infecting species.
HFRS
The incubation period is of 1–5 weeks and the onset of
diseases is with fever and influenza like symptoms.
Hemorrhagic manifestations, if seen, are in the form of
flushing of face, infection of the conjunctiva and mucus
membranes. The disease is described in five phases:
(1) Febrile phase lasting for 3–5 days: This is character-
ized by development of acute influenza like illness,
nausea, vomiting, back and abdominal pain. Protein-
uria develops on the 4th day. During the end of this
phase flushing of the face and conjunctival suffusion
is seen.
(2) Hypotensive (shock) phase lasting for a few hours to a
few days: This is associated with thrombocytopenia
and in one-third of cases death may occur. Proteinuria
tends to persist.
(3) Oliguric phase lasting for 3–5 days: There are signs
of acute renal failure and about half the deaths occur
during this time.
(4) Diuretic phase: It occurs in those who have survived
the earlier phases and lasts from a few days to a few
weeks with progressive improvement of renal
functions.
(5) Convalescent phase: There is sudden arrest of plasma
leak with concomitant reabsorption of extravagated
plasma and fluids.
Three kinds of clinical presentations of Hantavirus
infections can occur, namely, fever with shock and multi
organ failure, fever with oliguric acute renal failure and
febrile illness without any renal failure. Thus it has to be
differentiated from influenza, viral hepatitis, streptococcal
pharyngitis, leptospirosis, dengue fever and scrub typhus.
HCPS
It occurs mostly in adults with a case fatality rate of 33%.
There is a prodromal phase with fever, headache, chills and
myalgia and can be confused with other viral infections.
The prodrome is followed by the onset of cardiopulmonary
phase with pulmonary oedema, dyspnoea and hypoxemia.
Patients with fatal illness progress to depression, respira-
tory failure and acidosis leading to fatal arrhythmias.
Common laboratory findings are presence of hemocon-
centration, thrombocytopenia and atypical lymphocytes.
HFRS and HCPS are uncommon in children [27].
Treatment and Prevention
While there are no effective antiviral drugs for the treat-
ment of Hantavirus infections, trials of ribavarin done in
China have reported reduction in case fatality rates of
HFRS. Ribavarin, however, is ineffective in the treatment
of HPS. Prevention of exposure to rodents is the best way
to avoid infection. Rodents should be avoided in human
dwellings and proper food storage practices must be
employed. Rodents as pets should be discouraged. There
are a few inactivated vaccines (Hantvax) licensed for use in
Korea but protection is short lived. Baculovirus and
Proc. Natl. Acad. Sci. Sect B. Biol. Sci. (January–March 2012) 82(1):69–80 77
123

vaccinia-expressed Hantavirus glycoproteins confer protect
in animal models. There is ongoing research on nucleic
acid vaccines against Hantavirus [33].
Differential Diagnosis
A wide range of conditions (bacterial, viral, and parasitic
infections as well as noninfectious causes) should be con-
sidered in the differential diagnosis of VHF [34]. However,
most of these conditions do not cause bleeding manifes-
tations as a primary feature and most are not likely to occur
epidemiologically as a point-source epidemic with simul-
taneous presentation of many cases. Primary agents to
consider in the differential diagnosis are outlined in the
Table 3 [24, 31].
When to Consider the Diagnosis of VHF
Most clinicians have little or no clinical experience with
the syndromes that characterize VHF; therefore, a high
index of suspicion is needed to make an accurate diagnosis.
The diagnosis of VHF should be considered for any
patient who presents with:
• Acute onset of fever (less than 3 weeks’ duration)
• Severe prostrating or life-threatening illness
• Bleeding manifestations (i.e., at least two of the
following: hemorrhagic or purpuric rash, petechiae
[particularly in nondependent areas], epistaxis, hema-
temesis, hemoptysis, blood in stool, or other evidence
of bleeding)
• No predisposing factors for a bleeding diathesis
In naturally occurring cases, an appropriate travel or
exposure history will usually be present.
Tests for Detection of Hemorrhagic Fever Virus
Infection (Available Only at Specialized Laboratories)
• Antigen detection by antigen-capture enzyme-linked
immunosorbent assay (ELISA), performed on serum or
other samples, can be used to detect most hemorrhagic
fever viruses [35–38].
• Serology (either testing an acute-phase specimen for
IgM antibody or testing paired sera) also can be used to
diagnose most VHF infections.
• Reverse transcriptase (RT)-PCR methods have been
developed for a number of hemorrhagic fever viruses
[34, 37, 39–46]. As with other rare diseases, the
positive predictive value of PCR in the absence of other
corroborating medical or epidemiologic evidence is
exceedingly low.
• Cell culture: Although this is the ‘‘gold standard’’ of
virus detection and identification, performance of cell
culture with these viruses is time consuming and
extremely dangerous, and should be performed on
suspect cases only at BSL-4 laboratories.
• Generally, hemorrhagic fever viruses can be recovered
from serum or virtually any infected tissue.
• Most hemorrhagic fever viruses will grow in Vero
and other mammalian cell lines. Passage in labora-
tory animals may increase cell culture sensitivity
[29].
Treatment
Supportive care is essential for patients with all types of
VHF and includes the following:
• Maintenance of fluid and electrolyte balance, with
hemodynamic monitoring as needed
• Mechanical ventilation, as indicated
• Dialysis, as indicated
• Steroids have not been shown to be of value; however,
because adrenal involvement may occur in VHF cases,
steroids could be considered in certain situations [47,
48].
• Anticoagulant therapies, aspirin, nonsteroidal anti-
inflammatory medications, and intramuscular injections
are contraindicated
• Appropriate therapy for secondary infections
Management of severe bleeding complications is con-
troversial. Potential therapies include
• Clotting factor concentrates
• Platelets
• Fresh frozen plasma
• Heparin for DIC
Conclusion
VHFs are important cause of morbidity and mortality in
children as well as in adults globally. Since most of VHF
are arthropod vector borne disease, with the exception of
Hanta VHF, therefore public health strategies for elimi-
nation of vector as well as such protective measures by
individuals will act synergistically to reduce incidence of
VHF. Currently there are no specific antiviral agents and
management strategy of VHF is largely symptomatic.
Further research is being done to identify efficient treat-
ment and management strategy which include use of
effective vaccines, especially against Dengue fever, since
humans are the only reservoirs. In countries like India
where VHF is endemic professionals as well as commu-
nity must be aware of its symptoms and management.
78 Proc. Natl. Acad. Sci. Sect B. Biol. Sci. (January–March 2012) 82(1):69–80
123

References
1. Jahrling P (1997) Textbook of military medicine: medical aspects
of chemical and biological warfare. In: Zajtchuk R, Bellamy RF
(eds) Viral hemorrhagic fevers. Office of the Surgeon General,
Borden Institute, Walter Reed Army Medical Center, Washing-
ton, p 59
2. Mehla R, Kumar SR, Yadav P, Barde PV, Yergolkar PN, Er-
ickson BR (2009) Recent ancestry of Kyasanur forest disease
virus. Emerg Infect Dis 15(9):1431–1437
3. Tsai TF (2000) Flaviviruses (yellow fever, dengue, dengue
hemorrhagic fever, Japanese encephalitis, St. Louis encephalitis,
tick-borne encephalitis). In: Mandell GL, Bennett JE, Dolin R
(eds) Principles and practice of infectious diseases, 5th edn.
Churchill Livingstone, New York, pp 1855–1873
4. Gubler DJ, Zaki SR (1998) Dengue and other viral hemorrhagic
fevers. In: Nelson AM, Horsburgh CR (eds) Pathology of
emerging infections 2. American Society of Microbiology Press,
Washington, pp 43–71
5. Knipe DM, Howley PM et al (2007) Fields’ virology, 5th edn.
Lippincott Williams & Wilkins, Philadelphia
6. Wagner E, Hewlett M (2003) Basic virology: replication pattern
of specific viruses. Blackwell Publishing, Malden
7. Chambers TJ, Monath TP (2003) The flaviviruses: structure
replication and evolution. California, Academic Press, pp 27–65
8. Michael B, Dayal-Drager R, Guzman M (2009) Epidemiology,
burden of disease and transmission. In dengue: guidelines for diag-
nosis. Treatment prevention and control. WHO, Geneva, pp 14–16
9. Whitehead SS, Blaney JE, Durbin AP, Murphy BR (2007) Pros-
pects for a dengue virus vaccine. Nat Rev Microbiol 5:518–528
10. Halstead SB (1974) Etiologies of the experimental dengues of
Siler and Simmons. Am J Trop Med Hyg 23:974–982
11. Halstead SB, Nimmannitya S, Cohen SN (1970) Observations
related to pathogenesis of dengue hemorrhagic fever. Yale J Biol
Med 42:311–328
12. Sangkawibha N et al (1984) Risk factors in dengue shock syn-
drome: a prospective epidemiologic study in Rayong, Thailand. I.
The 1980 out break. Am J Epidemiol 120:653–669
13. Guzman MG et al (2000) Epidemiologic studies on dengue in
Santiago de Cuba, 1997. Am J Epidemiol 152:793–799
14. Halstead SB (1993) Monograph on dengue/dengue haemorrhagic
fever. In: Thongchareon P (ed) Pathophysiology and pathogenesis
of dengue haemorrhagic fever. World Health Organization,
Regional Office for South-East Asia, New Delhi, pp 80–103
15. Halstead SB (1989) Antibody, macrophages, dengue virus infec-
tion, shock, and hemorrhage: a pathogenetic cascade. Rev Infect
Dis 11:830–839
16. Halstead SB, Heinz FX (2005) Dengue virus: molecular basis of
cell entry and pathogenesis, 25–27 June 2003, Vienna, Austria.
Vaccine 23:849–856
17. Basu A, Chaturvedi UC (2008) Vascular endothelium: the battle field
of dengue viruses. FEMS Immunol Med Microbiol 53:287–299
18. Chaturvedi UC, Agarwal R, Elbishbishi EA, Mustafa AS (2000)
Cytokine cascade in dengue haemorrhagic fever: implications for
pathogenesis. FEMS Immunol Med Microbiol 28:183–188
19. Chaturvedi UC, Shrivastava R, Tripathi RK, Nagar R (2007)
Dengue virus-specific suppressor T cells: current perspectives.
FEMS Immunol Med Microbiol 50:285–299
20. Jain A, Chaturvedi UC (2010) Dengue in infants: an overview.
FEMS Immunol Med Microbiol 59:119–130
21. Rothman AL (2009) T lymphocyte responses to heterologous
secondary dengue virus infections. Ann N Y Acad Sci 1171:
E36–E41
22. Chaturvedi UC, Nagar R, Shrivastava R (2006) Dengue and
dengue haemorrhagic fever: implications of host genetics. FEMS
Immunol Med Microbiol 47:155–166
23. Chaturvedi UC, Shrivastava R, Nagar R (2005) Dengue vaccines:
problems and prospects. Indian J Med Res 121:639–652
24. WHO (1985) Viral hemorrhagic fevers: report of a WHO expert
committee, 1984 WHO technical report series 721
25. Pialoux G, Gauzere BA, Jaureguiberry S, Strobel M (2007) Chi-
kungunya, an epidemic arbovirosis. Lancet Infect Dis 7:319–327
26. Edelman R, Tacket CO, Wasserman SS, Bodison SA, Perry JG,
Mangiafico JA (2000) Phase II safety and immunogenicity study
of live chikungunya virus vaccine TSI-GSD-218. Am J Trop Med
Hyg 62:681–685
27. Pavri K (1989) Clinical, clinicopathologic, and hematologic
features of Kyasanur forest disease. Rev Infect Dis 11:S854–S859
28. Pattnaik P (2006) Kyasanur forest disease: an epidemiological
view in India. Rev Med Virol 16:151–165
29. Dandawate CN, Desai GB, Achar TR et al (1994) Field evalua-
tion of formalin inactivated Kyasanur forest disease virus tissue
culture vaccine in 3 districts of Karnataka state. Indian J Med Res
99:152–158
30. Gritsun TS, Lashkevich VA, Gould EA (2003) Tick-borne
encephalitis. Antiviral Res 57:129–146
31. Peters CJ (2005) California encephalitis, hantavirus pulmonary
syndrome and bunyavirid hemorrhagic fevers. In: Mandell GL,
Bennett JE, Dolin R (eds) Principles and practice of infectious
diseases, 6th edn. Churchill Livingstone, New York, pp 2086–
2090
32. Peters CJ (2005) Lymphocytic choriomeningitis virus, lassa
virus, and the South American hemorrhagic fevers. In: Mandell
GL, Dolin R, Bennett JE (eds) Mandell, Douglas, and Bennett’s
principles and practice of infectious diseases, 6th edn. Churchill
Livingstone, New York, pp 2090–2096
33. Huggins JW (1989) Prospects for treatment of viral hemorrhagic
fevers with ribavirin, a broad-spectrum antiviral drug. Rev Infect
Dis 11:S750–S761
34. Drosten C, Kummerer BM, Schmitz H et al (2003) Molecular
diagnostics of viral hemorrhagic fevers. Antiviral Res 57:61–87
35. Saijo M, Niikura M, Ikegami T, Kurane I, Kurata T, Morikawa S
(2006) Laboratory diagnostic systems for ebola and marburg
hemorrhagic fevers developed with recombinant proteins. Clin
Vaccine Immunol 13:444–451
36. Saijo M, Georges-Courbot MC, Marianneau P, Romanowski V,
Fukushi S, Mizutani T et al (2007) Development of recombinant
nucleoprotein-based diagnostic systems for Lassa fever. Clin
Vaccine Immunol 14:1182–1189
37. Towner JS, Rollin PE, Bausch DG, Sanchez A, Crary SM, Vin-
cent M et al (2004) Rapid diagnosis of ebola hemorrhagic fever
by reverse transcription-PCR in an outbreak setting and assess-
ment of patient viral load as a predictor of outcome. J Virol
78:4330–4341
38. Bausch DG, Rollin PE, Demby AH, Coulibaly M, Kanu J, Conteh
AS et al (2000) Diagnosis and clinical virology of Lassa fever as
evaluated by enzyme-linked immunosorbent assay, indirect
fluorescent-antibody test, and virus isolation. J Clin Microbiol
38:2670–2677
39. Drosten C, Gottig S, Schilling S, Asper M, Panning M, Schmitz H
et al (2002) Rapid detection and quantification of RNA of ebola
and marburg viruses, lassa virus, Crimean-Congo hemorrhagic
fever virus, rift valley fever virus, dengue virus, and yellow fever
virus by real-time reverse transcription-PCR. J Clin Microbiol
40:2323–2330
40. Leroy EM, Baize S, Lu CY, McCormick JB, Georges AJ,
Georges-Courbot MC et al (2000) Diagnosis of ebola haemor-
rhagic fever by RT-PCR in an epidemic setting. J Med Virol
60:463–467
41. Gibb TR, Norwood DA Jr, Woollen N, Henchal EA (2001)
Development and evaluation of a fluorogenic 50-nuclease assay to
identify marburg virus. Mol Cell Probes 15:259–266
Proc. Natl. Acad. Sci. Sect B. Biol. Sci. (January–March 2012) 82(1):69–80 79
123

42. Gibb TR, Norwood DA Jr, Woollen N, Henchal EA (2001)
Development and evaluation of a fluorogenic 50 nuclease assay to
detect and differentiate between ebola virus subtypes Zaire and
Sudan. J Clin Microbiol 39:4125–4130
43. Sall AA, Macondo EA, Sene OK, Diagne M, Sylla R, Mondo M
et al (2002) Use of reverse transcriptase PCR in early diagnosis of
Rift Valley fever. Clin Diagn Lab Immunol 9:713–715
44. Sall AA, Thonnon J, Sene OK, Fall A, Ndiaye M, Baudez B et al
(2001) Single-tube and nested reverse transcriptase-polymerase
chain reaction for detection of Rift Valley fever virus in human
and animal sera. J Virol Methods 91:85–92
45. Vieth S, Drosten C, Lenz O, Vincent M, Omilabu S, Hass M et al
(2007) RT-PCR assay for detection of lassa virus and related Old
World arena viruses targeting the L gene. Trans R Soc Trop Med
Hyg 101:1253–1264
46. Weidmann M, Muhlberger E, Hufert FT (2004) Rapid detection
protocol for filoviruses. J Clin Virol 30:94–99
47. Abraham E, Evans T (2002) Corticosteroids and septic shock.
JAMA (Editorial) 288:886–887
48. Annane D, Sebille V, Charpentier C, Bollaert PE, Francois B,
Korach JM et al (2002) Effect of treatment with low doses of
hydrocortisone and fludrocortisone on mortality in patients with
septic shock. JAMA 288:862–871
49. Muranyi W, bahr U, Zeier M, Woude FJ (2005) Hantavirus
infection. J Am Soc Nephrol 16:3669
80 Proc. Natl. Acad. Sci. Sect B. Biol. Sci. (January–March 2012) 82(1):69–80
123