competition in tree row agroforestry systems. 1. distribution and dynamics of fine root length and...
TRANSCRIPT

Plant and Soil227: 149–161, 2000.© 2000Kluwer Academic Publishers. Printed in the Netherlands.
149
Competition in tree row agroforestry systems. 1. Distribution anddynamics of fine root length and biomass
S.J. Livesley1,4, P.J. Gregory2 & R.J. Buresh31Ecosystems Research Group, Botany Department, The University of Western Australia, Nedlands, WA 6907,Australia.2Soil Science Department, The University of Reading, Whiteknights, P.O. Box 233, Reading, RG6 6DW,U.K. 3ICRAF, United Nations Avenue, Gigiri, P.O. Box 30667, Nairobi, Kenya.4Corresponding author∗
Received 24 September 1999. Accepted in revised form 10 July 2000
Key words:agroforestry,Grevillea robusta, maize, root competition, root length,Senna spectabilis
Abstract
Complementarity in the distribution of tree and crop root systems is important to minimise competition for re-sources whilst maximising resource use in agroforestry systems. A field study was conducted on a kaolinitic Oxisolin the sub-humid highlands of western Kenya to compare the distribution and dynamics of root length and biomassof a 3-year-oldGrevillea robustaA. Cunn. ex R. Br. (grevillea) tree row and a 3-year-oldSenna spectabilisDC.(senna) hedgerow grown withZea maysL. (maize). Tree roots were sampled to a 300 cm depth and 525 cm distancefrom the tree rows, both before and after maize cropping. Maize roots were sampled at two distances from the treerows (75–150 cm and 450–525 cm) to a maximum depth of 180 cm, at three developmental stages. The meanroot length density (Lrv) of the trees in the upper 15 cm was 0.55 cm cm−3 for grevillea and 1.44 cm cm−3 forsenna, at the start of the cropping season. The Lrv of senna decreased at every depth during the cropping season,whereas the Lrv of grevillea only decreased in the crop rooting zone. The fine root length of the trees decreased byabout 35% for grevillea and 65% for senna, because of maize competition, manual weeding, seasonal senescenceor pruning regime (senna). At anthesis, the Lrv of maize in the upper 15 cm was between 0.8 and 1.5 cm cm−3.Maize root length decreased with greater proximity to the tree rows, potentially reducing its ability to compete forsoil resources. However, the specific root length (m g−1) of maize was about twice that of the trees, so may havehad a competitive uptake advantage even when tree root length was greater. Differences in maize fine root lengthand biomass suggest that competition for soil resources and hence fine root length may have been more importantfor maize grown with senna than grevillea.
Introduction
Minimising resource competition between trees andcrops, whilst maximising the use of available re-sources, is central to improving yields and overallproductivity in agroforestry systems (Cannell et al.,1996; Ong and Black, 1995). The root system andits associated rhizosphere is the below-ground phys-ical, chemical and biological interface within whichresource uptake and competition between a crop andtree may occur. Knowledge of the spatial distribution
∗ FAX No: 61 8 9380 7925.E-mail: [email protected]
and density of tree and crop root systems, is a pre-liminary step towards gauging complementarity andunderstanding the processes that may occur (Gregory,1996; Van Noordwijk et al., 1996). The differencebetween the root length of two species growing in thesame soil volume can be used as an indication of theproportional share of the soil resources and thereforethe competitive ability of the plants (Bowen, 1985).
Root research is inherently laborious and time con-suming, particularly so with trees because of the largesoil volume exploited by their roots. The differentobjectives of previous studies have led to the use ofdifferent methodologies to assess and compare thespatial distribution of tree roots. Jonsson et al. (1988)

150
measured the vertical distribution of tree root densitiesas a means of assessing their competition potential,whereas Jama et al. (1998) compared the verticaldistribution of root length densities and nitrate con-centrations to investigate different demands by treesfor nitrogen. Rao et al. (1993) mapped tree root distri-bution with depth and distance from a tree in order toassess root mapping as a less labour intensive methodthan root sampling and to explore means of repres-enting such data graphically. Ruhigwa et al. (1992)assessed the competition potential of several tree spe-cies by combining root mapping with root lengthdensity measurement over depth and distance from atree row. Schroth and Zech (1995) added a temporalaspect by measuring root length density over depth anddistance at intervals through a growing season.
Comparison among studies is difficult because ofdifferences in the research technique, the form or unitof quantification, the size and growth stage of treesand the soil and climatic environment. As a result,the current understanding of tree root distribution, de-velopment and activity in agroforestry is fragmentary.Some generalisations about root distribution patternsare possible at the species level but the actual rootpattern of a specific tree genotype is heavily depend-ent on provenance and environmental factors, suchas soil type, management regime and climatic zone(Van Noordwijk et al., 1996). Mechanistic modellingof agroforestry systems will only be possible whena spatial, quantitative and temporal understanding oftree and crop uptake processes and interactions hasbeen collected, an important element of which is theunderstanding of the spatial distribution and dynamicsof root systems.
The general hypothesis behind this study is that atree with a greater fine root length will have a greatercompetitive effect upon an inter-planted crop. The aimof this study was to compare the spatial distributionand density of tree and crop roots before, during andafter the maize cropping season. Root length and bio-mass budgets were established to analyse the poten-tial for resource competition. This study complimentsother investigations into soil nitrogen uptake, loss andavailability (Livesley et al., 2000a) and soil water dy-namics (Livesley et al., 2000b) in these agroforestrysystems.
Materials and methods
The research was conducted at Nyabeda in westernKenya (34◦ 34′ E, 0◦ 06′ N, altitude 1330 m) duringthe long rains between March and August 1996. An-nual rainfall is about 1800 mm, which supports twocrop growing seasons. Rainfall was 903 mm between1 March and 30 July 1996. The soil is a very fine, ka-olinitic, isohyperthemic Kandiudalfic Eutrudox. Thetopsoil (0–15 cm) had a pH of 5.5 (1:2.5 soil/watersuspension), a clay content of 73% and sand contentof 8%.
The experiment was established in April 1993 withnursery-grown seedlings planted as a single row ofGrevillea robusta(grevillea) and single row ofSennaspectabilis(senna).Zea mays(maize, hybrid 512) wascropped biannually in the long rains (March–July) andthe short rains (September–January). The tree rowswere 20 m long and 12 m apart, allowing seven par-allel rows of maize to be planted 75 cm apart on bothsides of a tree row, at a within row spacing of 25 cm(53 000 plant ha−1). Grevillea was planted at a withinrow spacing of 100 cm, whilst senna was planted at awithin row spacing of 50 cm.
Grevillea was managed as a ‘tree row’ with thelower branches removed annually to reduce shading,whereas senna was managed as a ‘hedgerow’ that waspruned back to a 20 cm high stump before each crop-ping season. All prunings and leaf fall were collectedand removed from the site. On 29 February 1996,200 kg P ha−1 (triple superphosphate) was incorpor-ated into the upper 15 cm of both agroforestry sys-tems. Maize was sown on 11 March and the seedlingsthinned to their intended density on 29 March. Thefield plots were manually weeded every two weeks.The maize was harvested on 8 July, 117 days afterplanting (DAP).
Trench sampling of tree roots
Four trenches (profile walls) were prepared for eachtree row both before (March) and after (mid-July) themaize cropping season. Each profile wall was 330 cmdeep and extended, perpendicular to the tree row,between 50 and 550 cm from an individual tree. Soilsamples were collected from each profile wall by re-peatedly hammering in a metal box sampler (30× 15× 10 cm) to remove a continuous column of soil ateleven depth intervals of 0–15, 15–30, 30–45, 45–60,60–90, 90–120, 120–150, 150–180, 180–210, 210–255 and 255–300 cm, at six distances from the tree row

151
of 75, 150, 225, 300, 375 and 525 cm. This produced66 soil samples per profile wall. The upper 60 cmwas sampled in 15 cm intervals because root lengthchanges rapidly with depth in this zone.
Auger sampling of maize roots
Maize roots were sampled using a Jarret auger (10 cmdiameter) between 43 and 47 DAP at rapid stem elong-ation (45 DAP), between 72 and 75 DAP at 75% maizetasselling (tasselling), and between 93 and 96 DAP at75% silking (silking). Soil samples were taken fromwithin eight quadrats that were randomly selectedin each agroforestry system. Four quadrats spannedrows 1 and 2 at 75 cm and 150 cm from the treerow, and four spanned rows 6 and 7 at 450 cm and525 cm from the tree row. Soil samples were takento a depth of 90 cm at 45 DAP, 150 cm at tassellingand 180 cm at silking. Soil was sampled at intervalsof 0–15, 15–30, 30–45, 45–60, 60–90, 90–120, 120–150 and 150–180 cm. Within a quadrat, soil sampleswere taken from 4 intra-row and 4 inter-row locations,then samples from the same depth and row locationswere bulked together for analysis. Root parameterspresented in this paper are derived from the mean ofthese two locations. Maize roots were separated fromthe soil samples and their length and biomass weremeasured. Tree roots were also separated at tassellingto provide measurements of both maize and tree rootlength and biomass.
Root length and biomass determination
Soil samples were immediately placed in coveredbuckets, thoroughly mixed, weighed and then soakedovernight with water to help disaggregate the soil andease the separation of roots. On the following day, thesoaked soil was stirred and poured through a 0.5 mmsieve. Water was used to assist the passage of thesoil through the sieve. Roots and organic debris re-covered by the sieve were placed in plastic bags with17% acetic acid solution and stored in a refrigeratorat 5 ◦C to await sorting. Roots were separated fromorganic debris using forceps and sorted according tospecies and root diameter (≤ 2 and> 2 mm). Specieswere differentiated according to their colour, pliabilityand branching characteristics. Sorted roots were storedagain at 5◦C in plastic bags with 17% acetic acidsolution to await scanning. Prior to scanning, rootswere stained with methyl violet solution (0.1% in 10%ethanol; Harris and Campbell, 1989) and spread on aglass tray with minimal root overlap. Root length was
determined using a Delta-T root length system froman image obtained with a Hewlett Packard scanner ata resolution of 95 dots cm−1. Root length was calcu-lated using the Delta-T generic formula (Kirchoff andPendar, 1995). The scanner was regularly calibratedwith reference samples of known length and diameter.The root length density (Lrv) at each sample loca-tion was calculated using the known volume of soilsampled. After scanning, each root sample was driedat 70 ◦C for 48 h and then immediately weighed todetermine root dry weight. Soil contamination of dryroot weight was periodically measured by ashing andwas approximately 6% of fine root biomass (biomassdata not corrected).
Data analysis and presentation
The presented Lrv and root biomass data are themean of four profile samples with standard error. Thepresented Lrv profiles are weighted means of the sixsample distances from a tree row. Weighted becausethe sample distances from a tree row are not regularlyspaced. The 0–562.5 cm distance from a tree row wassubdivided into fifteen 37.5 cm sections, one weight-ing factor (w) equalled one 37.5 cm section. Eachsample distance was weighted according to the propor-tion of 0–562.5 cm it represented, 75 cm (w3), 150 cm(w2), 225 cm (w2), 300 cm (w2), 375 cm (w3) and525 cm (w3). Because Lrv decreased exponentiallywith depth, the data were log transformed (log base10) to better indicate changes at depth (Van Noordwijket al., 1996). The presented root data represent roots≤ 2 mm in diameter. Auger and box sampling do notprovide a good indication of the length or biomass ofroots> 2 mm in diameter because of their large spatialvariation and the intermittent nature of the samplingprocedure (Persson, 1989).
Contour-fill figures of Lrv (log base 10) as func-tions of soil depth and distance from the tree, wereprepared at both sample times for both species. Eachcontour-fill figure represents 66 co-ordinate pointsfrom the 11 sampled depths at six distances from thetree rows, an area on a vertical plane, 7.5–277.5 cmfrom the soil surface and 75–525 cm from the tree row.
The length (km m−2) and biomass (g m−2) of treeroots was calculated for an imaginary 36 m3 volumeof soil (1 m wide× 12 m long× 3 m deep) beforeand after maize cropping. The 36 m3 volume of soilwas subdivided into six regions, soil within 300 cmor soil between 300 and 600 cm from a tree row andsoil between 0 and 60 cm, 60 and 180 cm or 180
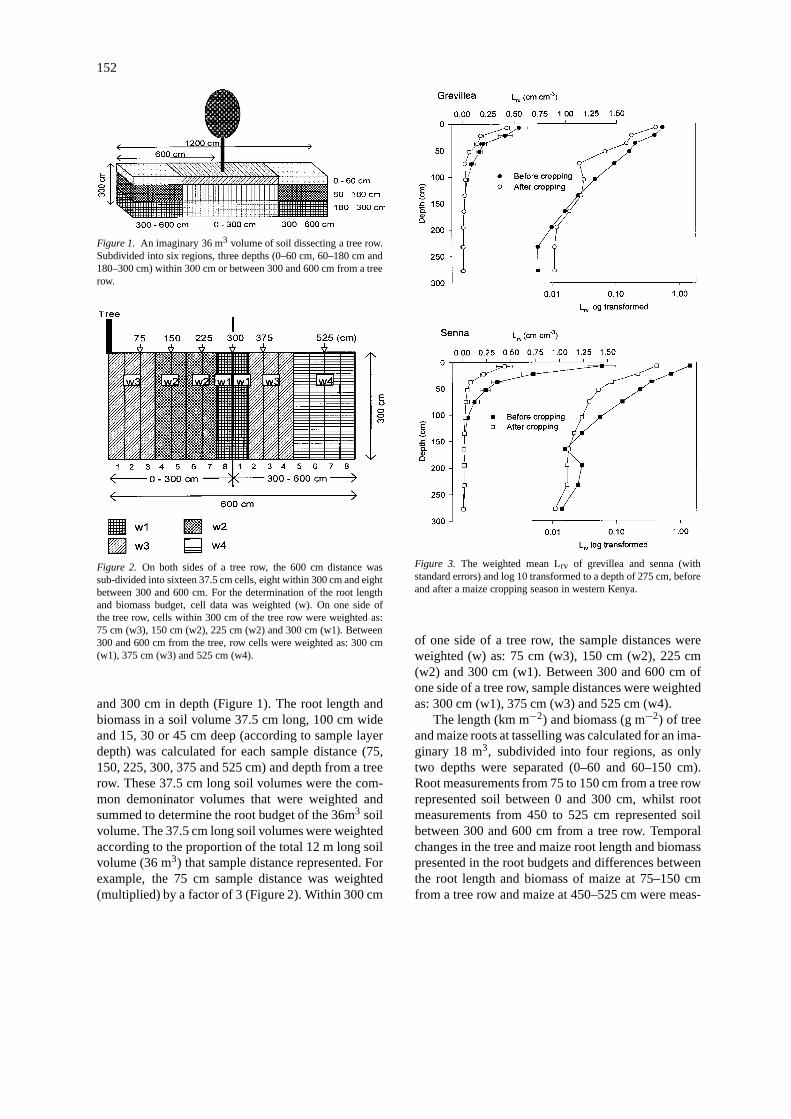
152
Figure 1. An imaginary 36 m3 volume of soil dissecting a tree row.Subdivided into six regions, three depths (0–60 cm, 60–180 cm and180–300 cm) within 300 cm or between 300 and 600 cm from a treerow.
Figure 2. On both sides of a tree row, the 600 cm distance wassub-divided into sixteen 37.5 cm cells, eight within 300 cm and eightbetween 300 and 600 cm. For the determination of the root lengthand biomass budget, cell data was weighted (w). On one side ofthe tree row, cells within 300 cm of the tree row were weighted as:75 cm (w3), 150 cm (w2), 225 cm (w2) and 300 cm (w1). Between300 and 600 cm from the tree, row cells were weighted as: 300 cm(w1), 375 cm (w3) and 525 cm (w4).
and 300 cm in depth (Figure 1). The root length andbiomass in a soil volume 37.5 cm long, 100 cm wideand 15, 30 or 45 cm deep (according to sample layerdepth) was calculated for each sample distance (75,150, 225, 300, 375 and 525 cm) and depth from a treerow. These 37.5 cm long soil volumes were the com-mon demoninator volumes that were weighted andsummed to determine the root budget of the 36m3 soilvolume. The 37.5 cm long soil volumes were weightedaccording to the proportion of the total 12 m long soilvolume (36 m3) that sample distance represented. Forexample, the 75 cm sample distance was weighted(multiplied) by a factor of 3 (Figure 2). Within 300 cm
Figure 3. The weighted mean Lrv of grevillea and senna (withstandard errors) and log 10 transformed to a depth of 275 cm, beforeand after a maize cropping season in western Kenya.
of one side of a tree row, the sample distances wereweighted (w) as: 75 cm (w3), 150 cm (w2), 225 cm(w2) and 300 cm (w1). Between 300 and 600 cm ofone side of a tree row, sample distances were weightedas: 300 cm (w1), 375 cm (w3) and 525 cm (w4).
The length (km m−2) and biomass (g m−2) of treeand maize roots at tasselling was calculated for an ima-ginary 18 m3, subdivided into four regions, as onlytwo depths were separated (0–60 and 60–150 cm).Root measurements from 75 to 150 cm from a tree rowrepresented soil between 0 and 300 cm, whilst rootmeasurements from 450 to 525 cm represented soilbetween 300 and 600 cm from a tree row. Temporalchanges in the tree and maize root length and biomasspresented in the root budgets and differences betweenthe root length and biomass of maize at 75–150 cmfrom a tree row and maize at 450–525 cm were meas-

153
Figure 4. The Lrv of grevillea and senna (log 10 transformed) withdistance (525 cm) and depth (275 cm)from the tree row before (A)and after (B) a maize cropping in western Kenya.
ured for statistical significance using a homoscedastictwo-tailedt-test. The sample number was four so thestatistical results should be viewed with caution.
Specific root lengths (SRL) expressed in m g−1
were calculated from the root biomass and lengthmeasurements of roots≤ 2 mm in diameter. The SRLof grevillea and senna was calculated before and aftermaize cropping and the SRL of maize was calculatedat 45, 75 and 95 DAP at 75–150 cm and 450–525 cmfrom the tree rows.
Results
Tree root length density before and after maizecropping
The Lrv of grevillea decreased exponentially withdepth (Figures 3 and 4). The Lrv of senna decreasedexponentially with depth but only at distances>225 cm from the hedgerow. At and within 225 cm fromthe hedgerow, Lrvdecreased exponentially between 0and 165 cm, but below 165 cm fine root length in-creased slightly before decreasing again towards thebase of the profile (Figures 3 and 4). Senna generallyhad a greater Lrv than grevillea, particularly in the sur-face layers and below 200 cm close to the hedgerow.This could be partly related to senna being plantedat twice the density of grevillea. In the upper 15 cmbefore maize planting, senna had an average Lrv of1.44 cm cm−3 (± 0.14), whereas that of grevillea was0.55 cm cm−3 (± 0.09).
The Lrv in the upper 100 cm decreased little withincreasing distance from the tree rows (Figure 4). Be-fore the maize cropping season, the Lrv of senna inthe upper 60 cm was 0.72 cm cm−3 at 75 cm from the
hedgerow, 0.93 cm cm−3 at 300 cm and 0.42 cm cm−3
at 525 cm. A similar pattern was observed for gre-villea, where the Lrv in the upper 60 cm increased from0.34 cm cm−3 at 75 cm to 0.53 cm cm−3 at 300 cm andthen decreased to 0.18 cm cm−3 at 525 cm (Figure 5).
The Lrv of both tree species decreased in the top-soil during the cropping season (Figures 3 and 4).After the maize harvest, the average Lrv of grevilleain the upper 60 cm was 0.21 cm cm−3 (± 0.03), com-pared to 0.33 cm cm−3 (± 0.02) at the start of theseason, and the comparable values for senna were0.20 cm cm−3 (± 0.01) after maize cropping and0.69 cm cm−3(± 0.03) at the start of the season.The Lrv of senna decreased during the cropping sea-son at almost every depth and distance from the treerow, whereas the Lrv of grevillea increased during thecropping season below 120 cm (Figure 3). After thecropping season, the Lrv of grevillea and senna in theupper 60 cm were< 0.30 cm cm−3 at 75–150 cmfrom the trees and decreased with increasing distancefrom the tree rows to< 0.16 cm cm−3 at a distance of525 cm. The large Lrv close to the senna tree row be-low 100 cm was still evident, though less pronounced(Figure 4).
Grevillea had about 4.9% of its total fine rootlength below 150 cm at the start of the maize crop-ping season, as compared to 6.5% for senna. Thesepercentages increased during the maize cropping sea-son to about 11.5% for grevillea and 13.8% for senna,but there was only a real increase in fine root lengthbelow 150 cm for grevillea. Tree Lrv below 150 cmwas≤ 0.025 cm cm−3 before maize cropping and≤0.040 cm cm−3 after the maize harvest.
Maize root length density during the cropping season
Generally, the Lrv of maize decreased exponentiallywith depth and Lrv was greater at 450–525 cm than at75–150 cm from the tree rows, especially at tasselling(Figure 6). At 45 DAP (Figure 6), the Lrv of maize inthe upper 15 cm was 0.57 cm cm−3at 75–150 cm fromgrevillea and 0.73 cm cm−3at 75–150 cm from senna.The Lrv of maize was 0.93 cm cm−3 at 450–525 cmfrom grevillea and 1.05 cm cm−3 at 450–525 cm fromsenna. The Lrv of maize grown with both grevilleaand senna decreased exponentially with depth to<
0.005 cm cm−3 at 60–90 cm. In the upper 15 cm, theLrv of maize grown with grevillea was significantlysmaller (p≤ 0.05) at 75–150 cm from the tree row thanthat at 450–525 cm. There was no significant distanceeffect at any depth for maize grown with the senna.
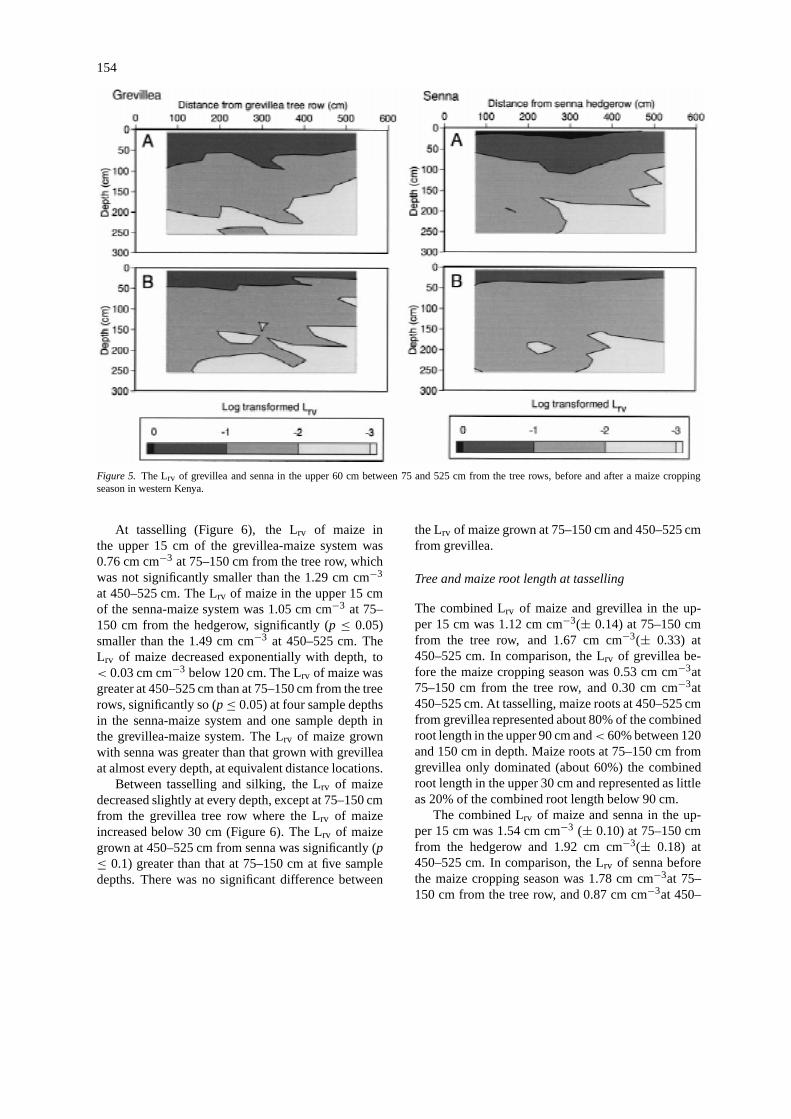
154
Figure 5. The Lrv of grevillea and senna in the upper 60 cm between 75 and 525 cm from the tree rows, before and after a maize croppingseason in western Kenya.
At tasselling (Figure 6), the Lrv of maize inthe upper 15 cm of the grevillea-maize system was0.76 cm cm−3 at 75–150 cm from the tree row, whichwas not significantly smaller than the 1.29 cm cm−3
at 450–525 cm. The Lrv of maize in the upper 15 cmof the senna-maize system was 1.05 cm cm−3 at 75–150 cm from the hedgerow, significantly (p ≤ 0.05)smaller than the 1.49 cm cm−3 at 450–525 cm. TheLrv of maize decreased exponentially with depth, to< 0.03 cm cm−3 below 120 cm. The Lrv of maize wasgreater at 450–525 cm than at 75–150 cm from the treerows, significantly so (p≤ 0.05) at four sample depthsin the senna-maize system and one sample depth inthe grevillea-maize system. The Lrv of maize grownwith senna was greater than that grown with grevilleaat almost every depth, at equivalent distance locations.
Between tasselling and silking, the Lrv of maizedecreased slightly at every depth, except at 75–150 cmfrom the grevillea tree row where the Lrv of maizeincreased below 30 cm (Figure 6). The Lrv of maizegrown at 450–525 cm from senna was significantly (p≤ 0.1) greater than that at 75–150 cm at five sampledepths. There was no significant difference between
the Lrv of maize grown at 75–150 cm and 450–525 cmfrom grevillea.
Tree and maize root length at tasselling
The combined Lrv of maize and grevillea in the up-per 15 cm was 1.12 cm cm−3(± 0.14) at 75–150 cmfrom the tree row, and 1.67 cm cm−3(± 0.33) at450–525 cm. In comparison, the Lrv of grevillea be-fore the maize cropping season was 0.53 cm cm−3at75–150 cm from the tree row, and 0.30 cm cm−3at450–525 cm. At tasselling, maize roots at 450–525 cmfrom grevillea represented about 80% of the combinedroot length in the upper 90 cm and< 60% between 120and 150 cm in depth. Maize roots at 75–150 cm fromgrevillea only dominated (about 60%) the combinedroot length in the upper 30 cm and represented as littleas 20% of the combined root length below 90 cm.
The combined Lrv of maize and senna in the up-per 15 cm was 1.54 cm cm−3 (± 0.10) at 75–150 cmfrom the hedgerow and 1.92 cm cm−3(± 0.18) at450–525 cm. In comparison, the Lrv of senna beforethe maize cropping season was 1.78 cm cm−3at 75–150 cm from the tree row, and 0.87 cm cm−3at 450–

155
Figure 6. The Lrv of maize (with standard errors) and log 10 transformed Lrv at 75–150 cm and 450–525 cm from a grevillea tree row andsenna hedgerow at 45, 75 and 95 days after planting in western Kenya.
525 cm. Maize roots at 450–525 cm from the sennahedgerow represented about 80% of the combined rootlength in the upper 45 cm and only represented< 50%below 120 cm. Maize roots at 75–150 cm from sennaonly dominated (< 70%) the combined root lengthin the upper 30 cm and represented< 40% of thecombined root length below 90 cm.
Root length and biomass budgets
Tree root length (km m−2) and root biomass (g m−2)was calculated within an imaginary soil volume, 1 mwide, 3 m deep and extending up to 6 m either side ofthe tree row (36 m3). Senna fine root length was abouttwice that of grevillea at the start of the season, butafter the cropping season the root lengths of both treespecies had decreased to similar values (Table 1).
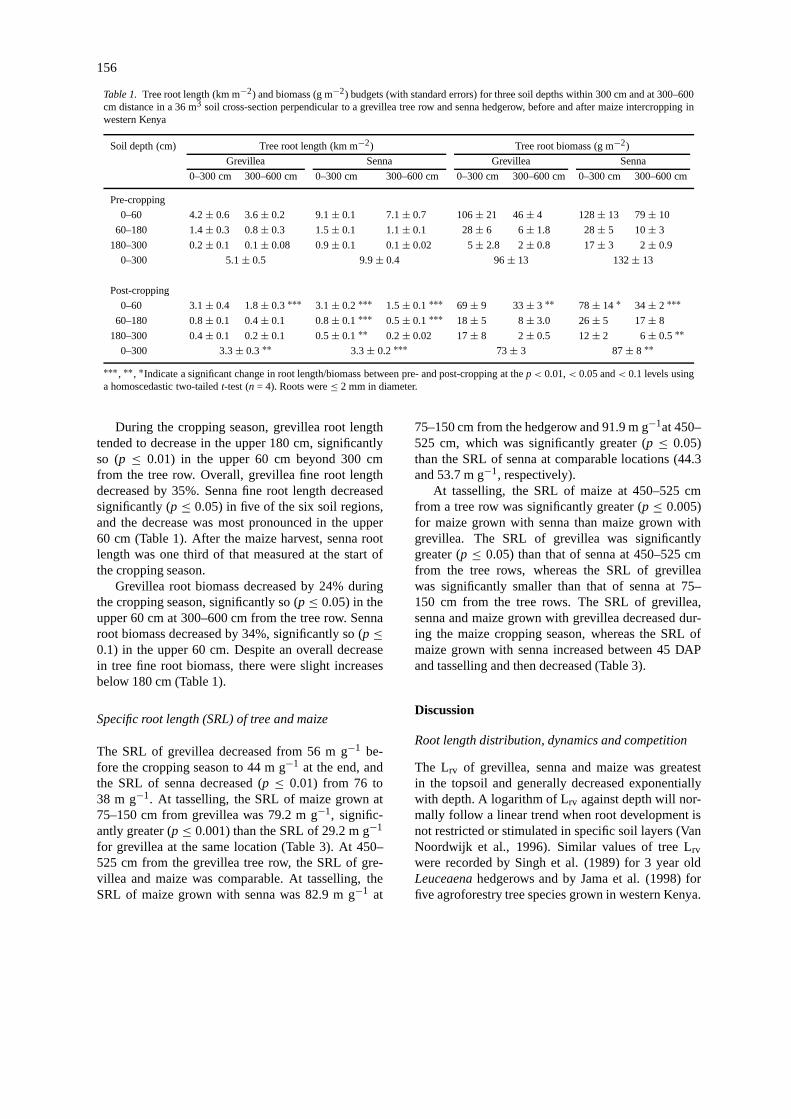
156
Table 1. Tree root length (km m−2) and biomass (g m−2) budgets (with standard errors) for three soil depths within 300 cm and at 300–600cm distance in a 36 m3 soil cross-section perpendicular to a grevillea tree row and senna hedgerow, before and after maize intercropping inwestern Kenya
Soil depth (cm) Tree root length (km m−2) Tree root biomass (g m−2)
Grevillea Senna Grevillea Senna
0–300 cm 300–600 cm 0–300 cm 300–600 cm 0–300 cm 300–600 cm 0–300 cm 300–600 cm
Pre-cropping
0–60 4.2± 0.6 3.6± 0.2 9.1± 0.1 7.1± 0.7 106± 21 46± 4 128± 13 79± 10
60–180 1.4± 0.3 0.8± 0.3 1.5± 0.1 1.1± 0.1 28± 6 6± 1.8 28± 5 10± 3
180–300 0.2± 0.1 0.1± 0.08 0.9± 0.1 0.1± 0.02 5± 2.8 2± 0.8 17± 3 2± 0.9
0–300 5.1± 0.5 9.9± 0.4 96± 13 132± 13
Post-cropping
0–60 3.1± 0.4 1.8± 0.3∗∗∗ 3.1± 0.2∗∗∗ 1.5± 0.1∗∗∗ 69± 9 33± 3 ∗∗ 78± 14 ∗ 34± 2 ∗∗∗60–180 0.8± 0.1 0.4± 0.1 0.8± 0.1∗∗∗ 0.5± 0.1∗∗∗ 18± 5 8± 3.0 26± 5 17± 8
180–300 0.4± 0.1 0.2± 0.1 0.5± 0.1∗∗ 0.2± 0.02 17± 8 2± 0.5 12± 2 6± 0.5∗∗0–300 3.3± 0.3∗∗ 3.3± 0.2∗∗∗ 73± 3 87± 8 ∗∗
∗∗∗ , ∗∗, ∗Indicate a significant change in root length/biomass between pre- and post-cropping at thep< 0.01,< 0.05 and< 0.1 levels usinga homoscedastic two-tailedt-test (n = 4). Roots were≤ 2 mm in diameter.
During the cropping season, grevillea root lengthtended to decrease in the upper 180 cm, significantlyso (p ≤ 0.01) in the upper 60 cm beyond 300 cmfrom the tree row. Overall, grevillea fine root lengthdecreased by 35%. Senna fine root length decreasedsignificantly (p ≤ 0.05) in five of the six soil regions,and the decrease was most pronounced in the upper60 cm (Table 1). After the maize harvest, senna rootlength was one third of that measured at the start ofthe cropping season.
Grevillea root biomass decreased by 24% duringthe cropping season, significantly so (p≤ 0.05) in theupper 60 cm at 300–600 cm from the tree row. Sennaroot biomass decreased by 34%, significantly so (p≤0.1) in the upper 60 cm. Despite an overall decreasein tree fine root biomass, there were slight increasesbelow 180 cm (Table 1).
Specific root length (SRL) of tree and maize
The SRL of grevillea decreased from 56 m g−1 be-fore the cropping season to 44 m g−1 at the end, andthe SRL of senna decreased (p ≤ 0.01) from 76 to38 m g−1. At tasselling, the SRL of maize grown at75–150 cm from grevillea was 79.2 m g−1, signific-antly greater (p≤ 0.001) than the SRL of 29.2 m g−1
for grevillea at the same location (Table 3). At 450–525 cm from the grevillea tree row, the SRL of gre-villea and maize was comparable. At tasselling, theSRL of maize grown with senna was 82.9 m g−1 at
75–150 cm from the hedgerow and 91.9 m g−1at 450–525 cm, which was significantly greater (p ≤ 0.05)than the SRL of senna at comparable locations (44.3and 53.7 m g−1, respectively).
At tasselling, the SRL of maize at 450–525 cmfrom a tree row was significantly greater (p ≤ 0.005)for maize grown with senna than maize grown withgrevillea. The SRL of grevillea was significantlygreater (p ≤ 0.05) than that of senna at 450–525 cmfrom the tree rows, whereas the SRL of grevilleawas significantly smaller than that of senna at 75–150 cm from the tree rows. The SRL of grevillea,senna and maize grown with grevillea decreased dur-ing the maize cropping season, whereas the SRL ofmaize grown with senna increased between 45 DAPand tasselling and then decreased (Table 3).
Discussion
Root length distribution, dynamics and competition
The Lrv of grevillea, senna and maize was greatestin the topsoil and generally decreased exponentiallywith depth. A logarithm of Lrv against depth will nor-mally follow a linear trend when root development isnot restricted or stimulated in specific soil layers (VanNoordwijk et al., 1996). Similar values of tree Lrvwere recorded by Singh et al. (1989) for 3 year oldLeuceaenahedgerows and by Jama et al. (1998) forfive agroforestry tree species grown in western Kenya.

157
Table 2. Root length (km m−2) and biomass (g m−2) budget (with standard errors) for maize, grevillea and senna at maize tasselling.Budgets represent two soil depths within 300 cm and at 300–600 cm distance in an 18 m3 soil cross-section perpendicular to a grevillea treerow and senna hedgerow intercropped with maize in western Kenya
Soil depth (cm) Maize root length (km m−2) Maize root biomass (g m−2)
Grown with grevillea Grown with senna Grown with grevillea Grown with senna
0–300 cm 300–600 cm 0–300 cm 300–600 cm 0–300 cm 300–600 cm 0–300 cm 300–600 cm
rows 1–3 rows 4–7 rows 1–3 rows 4–7 rows 1–3 rows 4–7 rows 1–3 rows 4–7
0–60 3.5± 0.7 8.0± 2.0 6.2± 1.2 9.4± 0.6 45± 9 118± 26 75± 17 108± 10
60–150 0.4± 0.2 1.2± 0.7 0.5± 0.1 1.4± 0.4 4± 2 15± 9 5± 1 11± 2
0–150 6.5± 1.3 8.7± 0.7 91± 16 99± 8
Tree root length (km m−2) Tree root length (g m−2)
Grevillea Senna Grevillea Senna
0–60 2.4± 0.4 2.1± 0.3∗∗ 3.2± 0.3∗ 2.4± 0.2∗∗∗ 92± 17 ∗ 26± 7 ∗∗ 70± 11 42± 3 ∗∗∗60–150 0.8± 0.1∗ 0.3± 0.1 0.9± 0.2∗ 0.8± 0.1 25± 4 ∗∗∗ 4± 0.3 23± 4 ∗∗∗ 19± 2 ∗∗0–150 2.8± 0.1∗∗ 3.6± 0.3∗∗∗ 73± 10 77± 8 ∗
∗∗∗, ∗∗, ∗Indicate a significant difference between tree and maize root length/biomass at thep < 0.01,< 0.05 and< 0.1 levels using ahomoscedastic two-tailedt-test (n = 4). Roots were≤ 2 mm in diameter.
Table 3. Specific root length (SRL) of maize, grevillea and senna (with standard errors)at 75–150 cmand 450–525 cm distance from a grevillea tree row and senna hedgerowintercropped with maize in western Kenya. DAP refers to days after maize planting
Treatment Distance Specific root length (m g−1
Maize Tree
45 DAP Tasselling Silking Tasselling
(75 DAP) (95 DAP) (75 DAP)
Grevillea-maize 75–150 cm 98± 18 79± 3 71± 5 29± 4
450–525 cm 84± 6 69± 5 59± 1 86± 11
Senna-maize 75–150 cm 62± 18 83± 6 72± 11 44± 2
450–525 cm 83± 6 92± 3 62± 7 54± 4
Roots were≤ 2 mm in diameter.
Similar values for maize Lrv were recorded whenmaize was grown in non-mulched tropical conditions(Schroth, 1994).
The Lrv was greater for grevillea and senna thanmaize at the start of the cropping season, but by tas-selling (the approximate time of maximum maize rootlength, Mengel and Barber, 1974) maize roots dom-inated the combined root length at 450–525 cm fromthe tree rows in the upper 120 cm. At 75–150 cm fromthe tree rows, tree roots dominated the combined rootlength in all but the upper 30 cm. Greater proximityto the tree rows reduced maize root length, potentiallydecreasing their ability to compete for resources (East-ham and Rose, 1990). Similar reductions in root lengthwith greater proximity to trees have been reported forgrass (Eastham and Rose, 1990) and sorghum (Jones
and Sinclair, 1996). Early in the cropping season, thepotential for resource competition between the treesand developing maize was widespread, and this po-tential for resource competition continued throughoutthe cropping season for maize grown close to the treerow. This potential assumes that the ability of a plantto compete for soil nutrient and water resources fromthe same soil volume is primarily proportional to itsdensity of fine root length (Bowen, 1985). Such anassumption does not take account of parameters suchas root diameter, mycorrhiza, physiological uptakecharacteristics, suberization (Schroth and Zech, 1995)and most importantly the resource demand to satisfygrowth (Gregory, 1996), all of which contribute to thecompetitive abilities of overlapping root systems.

158
The combined tree and maize Lrv did not exceed1.8 cm cm−3, less than the 5.0 cm cm−3thresholdabove which Gillespie (1989) suggested inter-rootcompetition for nitrate was likely. This threshold valuedoes not account for changes in soil type, nutrientstatus or the uptake characteristics of the species. Thesoil in this study was slightly deficient in N so thatthe critical threshold for inter-root competition may besmaller.
The Lrv of grevillea and senna decreased duringthe cropping season, especially in the upper 60 cm.The reduction in tree Lrv in the crop rooting zone(0–180 cm) may have been a response to growth ofthe maize root system and competition for resources.Concurrent growth with a competitive root system,such as maize, can reduce the growth of tree rootsin the topsoil and promote tree root growth deeper inthe profile (Eastham and Rose, 1990; Schroth, 1995).Huxley et al. (1994) observed that grevillea grownwith maize for nine cropping seasons had a smallerlateral spread and density of roots in the upper soil ascompared with grevillea grown without a crop, whereroots were found over a greater area and to greaterdepth. Concurrent growth of a competitive annual witha tree may force complementarity of their root systemdistribution and, therefore, possibly their resource ac-quisition. The large decrease in the Lrv of senna in theupper 60 cm may have been a response to the completeremoval of branches and foliage before maize plant-ing. Fine root death has been observed subsequentto shoot pruning, because of inadequate carbohydratesupply (Fownes and Anderson, 1991; Schroth, 1994).Jones and Sinclair (1996) observed greater root lengthreduction after shoot pruningProsopis juliflora thanafter pruningAcacia nilotica, and sorghum grownwith prunedP. juliflora developed a greater root lengththan sorghum grown with unprunedP. juliflora. Fineroot senescence may also have occurred in response tohoe tillage and weeding during the cropping season.Alternatively, the fine root systems may simply exper-ience seasonal cyclic growth and senescence, wherethe fine root length increases at the onset of the rains,to take advantage of the available moisture and min-eralised N, and then senesces through the rest of thecropping season (Atkinson, 1983). Srivastava et al.(1986) observed thatTectona grandishad two periodsof maximum root growth that coincided with the be-ginning of each of the bi-modal rainy seasons, and thegreatest seasonality was in the fine root fraction.
Tree root distribution as a ‘safety net’
The fine root systems of grevillea and senna extendedlaterally to> 525 cm and vertically to> 300 cm,which suggests they may be able to perform a protect-ive function against nutrient leaching (Schroth, 1994).This function would be most important early in therainy season when the maize roots are poorly de-veloped and N mineralisation and leaching are greatest(Greenland, 1958; Van Noordwijk et al., 1991). In thetopsoil, the fine root systems of grevillea and sennawere more dense at the onset of the rains than at theend of the cropping season. Under high rainfall con-ditions and low adsorption constants the retrieval andrecycling of nitrate by an extensive tree root systemmay rescue as much N as could be input by a N2-fixingtree (Van Noordwijk et al., 1991).
Less than 10% of grevillea and senna root lengthwas distributed below the crop rooting depth (180 cm),at densities< 0.04 cm cm−3. However, considerableamounts of water can be abstracted by a few roots(Gardner, 1964) and the importance of the peripheralroot system for deep water and nutrient uptake hasbeen recognised in several tropical tree species (Stoneand Kalisz, 1991). Mekonnen et al. (1997) reportedthat 15-month-old rows ofSesbania sesbanin west-ern Kenya can extract subsoil water at 200–400 cmdepth, which was not accessible to maize or nativeherbaceous vegetation.
The relatively large Lrv of senna in the subsoil(180–300 cm) and the slight increase in the Lrv ofgrevillea below 120 cm during the cropping seasonmay simply be characteristics of the rooting behaviourof these species or may be a response to maize rootcompetition in the upper soil. Similar increases inrooting density at depth have been measured forSes-bania sesban(Mekonnen et al., 1999),Dactyladenia(Acioa) barteri(Ruhigwa et al., 1992) andEucalyptuscamendulensis(Jonsson et al., 1988) tree rows. Thefrequency and persistence of increased root length atdepth is poorly understood as few studies either meas-ure below 1 m or measure the root distribution ofmature trees (Stone and Kalisz, 1991).
In contrast to other studies, the Lrv of grevilleaand senna in the upper 100 cm decreased little withincreasing horizontal distance from the tree row, bothbefore and after the cropping season. The Lrv of bothtrees was expected to decrease greatly with increas-ing distance from the tree rows (Van Noordwijk etal., 1996). This has been observed in other rootingstudies of agroforestry tree species such asAcioa bar-

159
teri (Hauser, 1993; Van Noordwijk, 1989; Ruhigwa etal., 1992),Sesbania sesban(Mekonnen et al., 1999)andGliricidia sepium(Schroth and Zech, 1995; VanNoordwijk, 1989). This apparent discrepancy withother studies may be due to the tree species, the soiltype, the management conditions, or the different tech-niques used to measure root length, particularly thefine root fraction that dominates at larger distancesfrom a tree.
Maize root length
The smaller Lrv of maize at 75–150 cm from a tree rowthan at 450–525 cm may have been caused by compet-ition for above- or below-ground resources (water, nu-trients or light) or perhaps allelopathic growth inhibi-tion. Huxley et al. (1994) observed from root mapping,that maize grown close to a grevillea hedgerow had aless dense and less extensive root distribution. Hullu-gale and Ndi (1994) observed that the root length ofcassava was smaller when grown between hedgerowsthan in monoculture. Maize grown with grevillea inthis study may have experienced above- or below-ground competition because the canopy and root sys-tem of grevillea was established and physiologicallyactive. Maize grown with senna, on the other hand pre-sumably experienced only below-ground competitionbecause the hedgerow canopy developed simultan-eously with that of the crop.
Below-ground competition is mediated indirectlythrough reductions in soil water or nutrient availab-ility, but it is very difficult to separate the effects ofallelopathy from those of resource competition. Gre-villea roots, foliage and leaf drip contain allelopathicchemicals that may inhibit growth (Anthofer, 1995).Grevillea foliage has a large C / N ratio (about 35), alarge lignin and tannin content but a small N contentof about 1.5% (Anthofer, 1998; Buresh, unpublished).In comparison, senna foliage has a large N contentof between 3.5 and 4.0% and a small lignin content(Buresh, unpublished). Maize grown within 150 cm ofgrevillea suffered late germination and poor biomassdevelopment (Livesley et al., 2000a) which may partlyhave been a result of allelopathic growth suppression.
A small greenhouse, pot study was conductedto examine the importance of allelopathy on maizegrowth. Maize was grown in pots with soil collectedfrom both the sole maize system and 1 m and 5 m dis-tances from the grevillea tree row and senna hedgerow.The soil from the grevillea and senna systems con-tained tree roots. The maize plants were supplied
with ample water, nitrogen and phosphorus. After 4weeks, the biomass of the maize shoots was compar-able between treatments and the control soil from thesole maize system.
This raised doubts about whether grevillea treeroots were allelopathically inhibiting maize develop-ment in the main experiment. It was assumed thatthe decomposition of grevillea leaf litter did not havean allelopathic effect on maize because litter fall wascollected and removed weekly. However, allelopathicchemicals could have entered the soil through exuda-tion from physiologically active roots of through leafdrip from the grevillea canopy, which was over 3 m indiameter.
Root biomass growth and loss
The decrease in grevillea and senna root length wasgreater than the decrease in root biomass during maizecropping because root senescence occurred in the finerroot fractions which provided the majority of the rootlength but little of the root biomass. The ratio of gre-villea and senna fine root length–fine root biomassdecreased because there was greater senescence in thefiner root fractions and an absence or reduction infine root formation. The decrease in the SRL of bothgrevillea and senna during the maize cropping seasonalso suggests greater senescence of fine than coarseroots. Maize produced finer roots than either tree spe-cies, investing less biomass per unit root length, asindicated by the large SRL for maize. Plants withsmall diameter roots or a large SRL tend to have agreater physiological capacity for resource uptake anda greater ability to respond to changes in their resourceenvironment (Eissenstat, 1992; Ryser and Lambers,1995). As a result, such plants may have better com-petitive efficiency (Fitter, 1994; Schroth and Zech,1995). Therefore, maize may even have a competit-ive advantage in situations where tree root length wasdominant, such as near to the tree rows.
Root biomass was not significantly greater formaize grown with senna than with grevillea, but maizetended to develop a finer root system when grown withsenna than grevillea. At tasselling, the SRL of maizegrown at 450–525 cm from senna was significantlygreater (p ≤ 0.05) than that of maize at 450–525 cmfrom grevillea. Similarly, the SRL of maize grownwith senna increased between 45 DAP and tasselling,whereas the SRL of maize grown with grevillea de-creased. These observations suggest that maize grownwith senna may have developed a finer root system to

160
better facilitate resource uptake in response to greatercompetition for soil resources. The proportional alloc-ation of biomass to a plants shoot or root system maybe strongly influenced by resource competition above-or below-ground. Maize root biomass as a proportionof total maize biomass is presented and discussed in asubsequent paper (Livesley et al., 2000a).
The large reductions in the length and biomassof tree fine roots during maize cropping may suggestopportunities for the control of tree resource uptakeand competitive ability through management practice(Jones and Sinclair, 1996). Enhanced and controlledtree root senescence may also provide an in situ sourceof organic matter and nutrients synchronised to thedemands of the crop and rhizosphere micro-organisms(Lehmann and Zech, 1998; Smucker et al., 1995).
Conclusions
Previous agroforestry root studies have indicated arapid decrease in tree root length with increasing dis-tance from a tree row (Van Noordwijk et al., 1996).The results of this study indicate that on the basis offine root length there was the potential for competitionbetween the tree row and maize crop, and this exten-ded several metres from the tree row. The fine rootlength of a tree row depends mainly on the species,planting density and management regime, and in thisstudy fine root length was shown to differ between ahigh canopy tree row and a regularly pruned hedgerow.The reduction in grevillea and senna root length dur-ing the maize cropping season may have been partly aresponse to the development of competitive maize rootsystems or, in the case of senna, the pruning regime.The causal factors were not separated, but opportunit-ies to manipulate the competitive root length of a treehave been demonstrated.
Greater proximity to a tree row reduced maizeroot length and, therefore, reduced its ability to com-pete for resources. However, the specific root lengthof maize was approximately twice that of grevilleaand senna, such that maize may have a competit-ive advantage in soil resource acquisition even insituations where its root length is proportionally in-ferior to that of the tree. Differences in maize fineroot length and biomass suggest that competition forsoil resources and hence fine root length may havebeen more important for maize grown with senna thangrevillea.
Acknowledgements
This work was financed by DFID Forestry ResearchProgramme, as a sub-contract of the AgroforestryModelling Project (R5651). The field research andlaboratory analyses were supported by a grant fromthe Swedish International Development CooperationAgency (Sida) to ICRAF. The authors thank Dr. BashirJama and the co-operation and hard work of the nu-merous field and laboratory workers at Maseno andNyabeda.
References
Anthofer J 1995 Nutritional and Allelopathic Effects of NineAgroforestry Tree Species on Wheat (Triticum aestivum) inthe Ethiopian Highlands. PhD Dissertation, Universität Ge-samthochschule Kassel, Germany.
Anthofer J, Hanson J and Jutzi S C 1998 Wheat growth as influ-enced by applications of agroforestry-tree prunings in Ethiopianhighlands. Agrofor. Syst. 40, 1–18.
Atkinson D 1983 The growth, activity and distribution of the fruittree root system. Plant Soil 71, 23–35.
Bowen G D 1985 Roots as a component of tree productivity.In At-tributes of Trees as Crop Plants. Eds. MGR Cannell and JE Jack-son pp 303–315. Institute of Terrestrial Ecology, Huntingdon,UK.
Cannell M G R, Van Noordwijk M and Ong C K 1996 The centralagroforestry hypothesis: The tree must acquire resources that thecrop would not otherwise acquire. Agrofor. Syst. 34, 27–31.
Eastham J and Rose C W 1990 Tree/pasture interactions at arange of tree densities in an agroforestry experiment. 1. Rootingpatterns. Austr. J. Agric. Res. 41, 683–695.
Eissenstat D M 1992 Costs and benefits of constructing roots ofsmaller diameter. J. Plant Nutr. 15, 763–782.
Fitter A H 1994 Architecture and biomass allocation as compon-ents of the plastic response of root systems to soil heterogeneity.In Exploitation of Environmental Heterogeneity by Plants. Eds.MM Caldwell and RW Pearcy pp 305–322. Academic Press,London, UK.
Fownes J H and Anderson D G 1991 Change in nodule and root bio-mass ofSesbania sesbanandLeaucaena leucocephalafollowingcoppicing. Plant Soil 138, 9–16.
Gardner W R 1964 Relations of root distribution to water uptakeand availability. Agron. J. 56, 41–45.
Gillespie A R 1989 Modelling nutrient flux and interspecies rootcompetition in agroforestry interplanting. Agrofor. Syst. 8, 257–265.
Greenland D J 1958 Nitrate fluctuations in tropical soils. J. Agric.Sci. 50, 82–92.
Gregory P J 1996 Approaches to modelling the uptake of water andnutrients in agroforestry systems. Agrofor. Syst. 34, 51–65.
Harris G A and Campbell G S 1989 Automated quantification ofroots using a simple image analyzer. Agron. J. 81, 935–938.
Hauser S 1993 Root distribution ofDactyladenia(Acioa) barteriandSenna(Cassia) siameain alley cropping on Ultisol. I. Im-plication for field experimentation. Agrofor. Syst. 24, 111–121.
Hulugalle N R and Ndi J N 1994 Changes in soil properties of anewly-cleared ultisol due to establishment of hedgerow speciesin alley cropping systems. J. Agric. Sci. 122, 435–443.

161
Huxley P A, Pinney A, Akunda E and Muraya P 1994 A tree/crop in-terface orientation experiment withGrevillea robustahedgerowand maize. Agrofor. Syst. 26, 23–45.
Jama B, Buresh R J, Ndufa J K and Shepherd K D 1998 Vertical dis-tribution of tree roots and soil nitrate as affected by tree speciesand phosphorous. Soil Sci. Soc. Am. J. 62, 280–286.
Jones M and Sinclair F 1996 Differences in root system responsesof two semi-arid tree species to crown pruning. Agrofor. Forum7 (2), 24–27.
Jonsson K, Fidjeland L, Maghembe J A and Hogberg P 1988 Thevertical distribution of fine roots of five tree species and maize inMorogoro, Tanzania. Agrofor. Syst. 6, 63–69.
Kirchoff G and Pendar C E 1995 Delta-T Scanner – User Manualv.2.0. Ed. Webb N. Delta-T Devices Ltd., Cambridge, UK.
Lehmann J and Zech W 1998 Fine root turnover of irrigatedhedgerow intercropping in Northern Kenya. Plant Soil 198,19–31.
Livesley S J, Gregory P J and Buresh R J 2000a Competitionbetween trees and crops in tree row agroforestry systems. 2. Dis-tribution, dynamics and uptake of soil inorganic N. Plant Soil(submitted).
Livesley S J, Gregory P J and Buresh R J 2000b Competitionbetween trees and crops in tree row agroforestry systems. 3.Distribution and dynamics of soil water. Plant Soil (submitted).
Mekonnen K, Buresh R J and Jama B 1997 Root and inorganic ni-trogen distributions in sesbania fallow, natural fallow and maize.Plant Soil 188, 319–327.
Mekonnen K, Buresh R J, Coe R and Kipleting K 1999 Rootlength and nitrate underSesbania sesban: Vertical and horizontaldistribution and variability. Agrofor. Syst. 42, 265–282.
Mengel D B and Barber S A 1974 Development and distribution ofthe corn root system under field conditions. Agron. J. 66, 341–344.
Ong C K and Black C R 1995 Complementarity in resource use inagroforestry systems.In Alley Farming – Research and Develop-ment. Eds. BT Kang, AO Osiname and A Larbi pp 73–89. IITA,Ibadan, Nigeria.
Persson H 1989 Methods of studying root dynamics in relationto nutrient cycling.In Nutrient Cycling in Terrestrial Systems.Eds. AF Harrison, P Ineson and OW Heal. pp 198–217. ElsevierScience Publishing Ltd., London.
Rao M R, Muraya P and Huxley P A 1993 Observations of sometree root systems in agroforestry intercrop situations and theirgraphical representation. Exp. Agric. 29, 183–194.
Ruhigwa B A, Gichuru M P, Mambani B and Tariah N M 1992
Root distribution ofAcioa barteri, Alchornea cordifolia, CassiasiameaandGmelina arboreain an acid Ultisol. Agrofor. Syst.19, 67–78.
Ryser P and Lambers H 1995 Root and leaf attributes accounting forthe performance of fast- and slow-growing grasses at differentnutrient supply. Plant Soil 170, 251–265.
Schroth G 1994 Above- and Below-Ground Interactions in AlleyCropping withGliricidia sepiumas Compared to Conventionaland Mulched Sole Cropping on a High Base Status Soil in theWest African Rainforest Zone. Ph.D. Dissertation, BayreutherUniversity, Germany.
Schroth G 1995 Tree root characteristics as criteria for species se-lection and systems design in agroforestry. Agrofor. Syst. 30,125–143.
Schroth G and Zech W 1995 Root length dynamics in agroforestrywith Gliricidia sepiumas compared to sole cropping in the semi-deciduous rainforest of West Africa. Plant Soil 170, 297–306.
Singh R P, Ong C K and Saharan N 1989 Above- and below-groundinteractions in alley-cropping in semi-arid India. Agrofor. Syst.9, 259–274.
Smucker A J M, Ellis B G and Kang B T 1995 Alley cropping onan Alfisol in the forest savanna transition zone: root, nutrient andwater dynamics.In Alley Farming – Research and Development.Eds. BT Kang, OA Osiname and A Larbi pp 103–121. IITA,Ibadan, Nigeria.
Srivastava S K, Singh K P and Upadhayay R S 1986 Fine rootgrowth dynamics in teak (Tectona grandis). Can. J. For. Sci. 16,1360–1364.
Stone E L and Kalisz P J 1991 On the maximum extent of tree roots.For. Ecol. Man. 46, 59–102.
Van Noordwijk M 1989 Rooting depth in cropping systems in thehumid tropics in relation to nutrient use efficiency.In NutrientManagement for Food Crop Production in Tropical Farming Sys-tems. Ed. J Van der Heide pp 157–169. Institute of Soil Fertility,Haren, The Netherlands.
Van Noordwijk M, Lawson G, Soumare A, Groot J J R and HariahK 1996 Root distribution of trees and crops competition and/orcomplementarity.In Tree Crop Interactions – A PhysiologicalApproach. Eds. CK Ong and P Huxley Chapter 9, pp 319–364.CAB International, Wallingford, UK.
Van Noordwijk M, Widianto H M and Hairiah K 1991 Old tree rootchannels in acid soils in the humid tropics: important for croproot penetration, water infiltration and nitrogen management.Plant Soil 134, 37–44.