color-matching, color-naming and color-memory in split-brain patients
TRANSCRIPT

Neurop~ycholog~a, Vol. 19. No. 4, pp. 523 541, 1981 Prmted in Great Bntam
OOZS-3932/Sl/O40523 l%2.00/0 t > 1981 Pergamon Press Ltd.
COLOR-MATCHING, COLOR-NAMING AND COLOR-MEMORY IN SPLIT-BRAIN PATIENTS
JERRE LEVY*
Department of Behavioral Sciences, University of Chicago, Chicago, Illinois, U.S.A
and
COLWYN TREVARTHEN
Department of Psychology, University of Edinburgh, Edinburgh, Scotland
(Rrcei~~d 19 Junuary 1981)
Abstract-Bilateral chimeric stimuli were projected tachistoscopically to split-brain patients for the purpose of assessing hemispheric asymmetries in color-naming, color-matching and memory for colors ofwell-known objects. There was a strong bias for naming colors ofstimuli projected to the left hemisphere, but some patients were also above chance in naming colors of stimuli projected to the right hemisphere. In color-matching tasks, patients varied in the direction and degree of asymmetric control of responses, and asymmetry patterns varied within patients across different tasks, with symmetry of hemispheric control increasing as task complexity increased. Both hemispheres performed well above chance in memory for colors of objects depicted in line-drawings, and the right hemisphere appeared to perform somewhat better than the left.
THE ROLES of the left and right cerebral hemispheres in color perception has been a matter of debate for some years. Loss of the ability to name colors has been reported by a number of investigators [l, 23, and GESCHWIN~ and FUSILLO [3] described a case where color-naming disability was associated with a lesion of the left-hemisphere visual areas and of the splenium of the corpus callosum, suggesting that anomia for colors results from disconnection of visual from language regions. OXBURY et al. [4] have drawn a distinction between pure linguistic disruption as a cause of anomia for colors and the disconnection syndrome described by GESCHWIND and FUSILLO [3]. In both these disorders, however, a color-naming deficit results from a left-hemisphere lesion that either damages language regions directly or isolates them from visual processing regions.
DE RENZI and SPINNLER [S] investigated subtle color discrimination not involving a verbal report, and found a significant inferiority in patients with right-hemisphere damage. SCOTTI and SPINNLER [6] observed that patients with right-hemisphere damage and visual field defects were inferior to others on the FarnsworthPMunsell 100 Hue Test, a finding consistent with CAPITANI et al.‘s [7] claim that patients with right parietal lesions have the highest error scores on this test.
These studies of brain-damaged patients suggest that defects in color naming can be due either to a pure aphasia, to disconnection of visual from linguistic processing regions, or to
*Requests for reprints should be addressed to: Jerre Levy. 5848 S. University Ave., University of Chicago. Chicago, IL 60637. U.S.A.
523

disorders in the color-perception system itself; the first two disabilities appear generally with left-hemisphere damage and the latter. with right-hemisphere damage.
Investigations of normal people support the conclusion that the two hemispheres play differing roles in color perception and naming. PENNAL [S] presented one of 12 stimuli to the left or right field at a 30 msec duration, requiring subjects to press one of 24 colored buttons. arranged in a circle, to indicate their choice of a matching color. For both latency and error data, superior performance was observed for stimuli in the left visual field (LVF) as compared to stimuli in the right visual field (RVF). DIMOND and BEAUWNT 193 failed to find hemispheric differences in voice reaction time for naming tachistoscopically presented colors, but unlike the PENNAL study. the DIMONI> and BEACJMONT colors were easily nameable and the dependent measure was a verbal response. Their results are consistent with the view that the hemispheres, though possibly using different strategies. are equally competent at discriminating and remembering nameable colors.
Support for this possibility is provided in an investigation by S~HXWT and DAVIS [IO], who measured manual reaction times. Subjects were required to identify, as rapidly as possible, one of three color-words or one of three ink colors in which a word was printed. These stimuli were projected for 150 msec in the left or right field. For both word-reading and identification of ink color, words and ink colors were either congruent (e.g., the word BLUE printed in blue ink) or incongruent (e.g. the word BLUE printed in green ink). In the ink- color identification task, no field difference appeared when words and ink colors were congruent. but incongruent stimuli induced a significantly greater increase in reaction time for the RVF than for the LVF. The field symmetry in the congruent task concurs with results of DIMOND and BEAUMWT 193 for nameable colors. The results for the incongruent task reflect, not a right-hemisphere perceptual superiority for nameable colors. but rather a greater right-hemisphere resistance to interference by color-words.
The various studies reveal a right-hemisphere superiority for color discrimination when colors are difficult to name or describe, but equality of the hemispheres when colors come from an easily nameable set. Such a nameable/non-nameable distinction is similar to that pertaining to shapes. As MILNER [I l] and others have noted, studies of brain-damaged
patients and of normal individuals indicate that the right hemisphere surpasses the left in shape discrimination and in memory for shapes that have no verbal labels or are resistant to verbal description. Many have suggested that processing strategies of the left hemisphere rely on language or language-related functions. whereas those of the right hemisphere depend on imagistic constructions closely representing the sensory experience. Thus. discrimination and memory for nameable stimuli or stimuli easy to describe would be equal for the two hemispheres, but via the use of different cognitive operations.
There are, however. certain observations of neurological patients that. taken at face value. seem incongruent with inferences drawn from other studies regarding the roles of the left and right hemispheres in color discrimination and memory. COHFS and KFLTI-R [ 121 found that aphasic patients. in selecting colors varying in hue. saturation, and brightness that represented the typical color of objects depicted in achromatic line-drawings. were equally disrupted for all three aspects of colors as compared to control subjects. confirming earliel investigations that showed poor performance of aphasics in associating colors with line- drawings [S. 13. 141 and also demonstrating that tho disability is beyond the perceptual level. at the le\-el of conceptual and abstract thinking. COW u and KF L.TI.R suggest that the deficit of aphasics is due to disruption of the abilit! to isolate and handle specific features of objects and L’onccpts. There ih much cmplrical support for the conclusion that chromatic qualities of

COLOR-MArCHING. COLOK-NAMINti AND COLOR-MEMORY IN SPLIT-BKAIN PATIENTS 525
objects are structured in the left hemisphere as specific feature characteristics, and that this structure is disordered with left-hemisphere lesions. A discrepancy arises because other findings, previously discussed, indicate that the right hemisphere has refined imagistic representations of colors, and it would be expected to have little difficulty in selecting colors of well-known objects. Either the right hemisphere is unable to use these representations for associating colors with line-drawings or left-hemisphere lesions resulting in aphasia entail a
disordering of interhemispheric regulation, precluding the right hemisphere from bringing its skills to bear on the task. In particular, the deficits observed in aphasics may be due to a retention of processing control by the damaged left hemisphere, obscuring the abilities of the right, rather than being due to a real right-hemisphere incapacity.
The possibility that, at least in some instances, the less competent hemisphere assumes control of processing and behavior, and that the tendency of a hemisphere to dominate behavioral control is not necessarily perfectly related to ability differences between the two sides of the brain has been confirmed in split-brain patients. In earlier studies, we found that the right hemisphere assumed control of behavior when a simple matching task was given, even when the left hemisphere was equally or more competent in terms of accuracy of
performance [ 151, and the left hemisphere assumed control of processing when simple words had to be read for meaning, even when the right hemisphere showed perfect accuracy on the task [16]. Further, asymmetry ofcontrol can and does shift from one hemisphere to the other as a function of processing strategies that are employed even when stimulus items, choice
items, and the motoric response remain invariant [17]. Thus, the deficit in aphasics at associating colors with line-drawings may imply abnormal
control of processing by the damaged left hemisphere, or it may imply that the right hemisphere is simply incapable of performing the task. The goals of the present investigation were to determine first, which hemisphere actually dominates control of processing in any given task; second, the degree of unihemispheric or bihemispheric control as a function of task demands; and third, whether only the left hemisphere or both hemispheres is/are competent to derive the typical colors of objects depicted in line-drawings.
We examined hemispheric asymmetry of control for the identification matching, or memory of nameable colors in split-brain patients.
METHOD SlrhifV’t Y
The subjects were the five split-brain patients. A.A., C.C.. L.B.. N.G., and R.Y.: all are epileptics who have undergone total neocommissurotomy.
A.A., although he suffered two convulsions at age 4 months associated with high fever. developed normally until age 51 when generaltzed convulsions appeared. These sometimes began in the right arm. suggesting a left- hemisphere focus. He was operated upon at age 14 in 1964. The surgery produced right cerebral swelling and a persistent spastic left leg with posittve Babinski sign. so that he prohably has significantly more right- than left- hemisphere damage. in spite of a possible left-hemisphere epileptic focus.
C.C. experienced anoxia post-partum but developed normally until age X when his school work deteriorated. At age IO. spells of speechlessness occurred. associated with rightward head-turning and. occasionally. loss of
consciousness. The hehavioral signs associated with the seizure suggest a left-hemisphere focus. The commissurotomy operation at age 13 in 1965 involved trauma to lhe left hemisphere and was followed hy muteness for 3 months. He is thought IO have greater left- than right-hemisphere damage.
L.B. weighed 5 Ih at birth. was cyanotic and remained in an lsolette for 8 days. His first convulsion occurred at age 3;. becommg progt-esstvely worse unttl he was operated upon at age 13 in 1965. The surgery went smoothly. and L.B. showed a rapid recov cry : he \!‘a< able lo speak one day after sur-ger!. His performance 1.0. is aver-aec I 1001. ;tnd his verbal I Q ~~‘11 above aver-age f I 151. The verbal I (_I of I I5 I\ prohahly an undere\timatc of L.B.‘\ verbal rc;r~~ntng ~~pacit! \IIICC he u;c IOU on ho111 the Drgit Span and 12~-~~hrnet~c wbresr~ 01’ rhe bb4IS. hut \(;I\ tv,o \tandard

deviations above the population mean on the Similarities suhtest (equivalent to an 1.0. of 130). There IS no evidence of brain damage in either cerebral hemisphere.
N.G. was premature. but developed normally until age IX when she began hacmg convulsions. She was operated upon at age 30 in 1963. Her EEG had shown left-temporal \lowing. and skull X-ray> revealed a mulberry calcificatmn, I cm in diameter. beneath the right central cortex. The postsurgical EEG WHS normal, and it has remained normal. except for a one-year period in 1972 when seizures recurred in association ulth a left tcmporal- lohc focus after a reduction III antI-se17ure medicatmn. N.G. manifesta no other sign\ of brain damage.
R.Y. had a normal dellvery and de\elopcd normally until age 13 when he was struck by a car and wa\ stuporou\ for about half an hour. At age 16. hc began having what his family descrtbed as “cpells” (probably p~vir tntrl seizures), and generalized convulsions appeared one year Inter. HIS seizure> were associated with a visual aura, suggesting a right posterior focus. but preoperative and postoperatIve EEG’s were nonlocallring and neurological examinatlonh were normal. However. conslstent with possihlllty of a right postcrmr disorder. his postsurglcal verbal I.Q. ~35 99. whereas hi\ performance I.Q. was only 79. He was operated upon in 1966 at age 43.
To summa-i/e, AA and C.C. probably have \ubstantlal asymmetric damage. /\.A. ulth more to the right hemisphere and C.C. *ith more to the left hemisphere. There I\ no evidence of damage to either side of the hraln in LB. and only Ehght local damage in N.G.; R Y. appears to habe a disorder of right posterior rcglons.
For all tests, sttmuh were exposed m a modllied Harvard 7-channel Tachlstoacopc for I SO myec. Each stmluluh card was seen as a 5 x 5 white square agamst a blue hackground, the whltc square having a luminance of 7.5 ft L. with stimuli centred in the middle of the square. A constantly illuminated tixation field of deep blue (0.2 ft L) way provided with a pin-pomt of red Ilght at the center that was allgned with the center of the stimult. Eye movements were monitored by electrooculography (EOG) during all tests. az described in a previous paper [IS].
Colored stimuli were fabricated of ‘%d-color” frosted acetate sheets chosen to obtain <p&tic hues, hrlghtnesse\ and saturations. In tests using three color\. the colorr consl\ted of red. blue-green, and orange. Matched to Munsell chips. they had the Munsell notations of h.ZR.‘4.3 11.2. 4.6B<;:4.5?+.5 and X.6YR:7.3’15.2. respectively. For tests using four color\, thecolors werepreen (3.2G4.9/1 I.1 ). red (7.OR. 4.6’13.6).mustard yellow (9.4YR; 5.7.!X.5) nnd blue (4.98/4.5,8.4). Eight color patches were chosen to represent the colors of objects shown :I$ black-outlined drawings: reddish-brown (6.1 R;2.9./7.3) representing the color of 3 fox, yellow (6.hY,‘X.5;9.5) for bananas. pink (I.1 R. 6.5/9.2) for a tlamingo, purple (O.hP/7.?;‘9.5) for grape,. green (6.7<;6.2,‘X.S) for peas, red (5.OR~3.9’15.4) for \trawherrlcq. gray (2.lY.!5.4;1.5) for an elephant. and orange (7.XYR:7..5.13.4) for an orange.
C‘hlmeric stimuli in the form of butterllle\. each umg of a dlffercnt color. were constructed having a center gray body of 0.S of visual angle: each wing extended 2 into each visual field for the upper ulng and I.5 for the lower wing. On each trial. a chimeric butterlly was tachistoscopicnlly presented and subjects were asked either to (I) point to a nonchimeric hutterlly I” free cislon matching the color of the butterOy percaved or (2) name the color of the butterlly perceived. Three dlffcrent tcsta were gl\cn: In the first. vx chimera were constructed from the three colorc red, blue-green and orange ulth three nonchlmerlc hutterllies as choices. In the \econd. I2 chimeras acre constructed from the four color\ rcd.grecn. mustard yellou.and blur. with four nonchlmerlc butterRie\ as cholcc\. In the third.24chlmeras wereconstructed.Itlentlcal to tho\c III thcfout--color tc~t.ab~,ve.cvcept that corrtr;lstlng~p~,t\. different from either wing color. were added to each wing. Thel-c v.erc 12 spotted-wing huttertlie\ 111 the choice \et Each spotted chimera. thus. contalncd all four color\ in wings and \pot\ (FIN. I )
2 ~‘oli,rccl-/o~fl1,\ I<‘\,,, A total of 36 chimcric stimuh were constructed from outlmed triangles. square,. and circle5 The outline\ wcrc
drawn in either red, blue-green. or orange ink; all chimerar differed on the left and rtght with respect to hoth COIOI and shape (Fig. 2). The sides of the square. equilateral triangle. and the diameter of the ctrcle were approximately 3 of visual angle when seen III the tachisto\cope Subject5 were asked either to (I) select from among three colored patches the color they UN. (7) select from among three black-outlmed drawing\ the form they WW. or (3) select from among nine color-outlined forms. the color and form of the qtimuluh they saw. or they ucre a\kcd to name the color. the form. or both the color and fol-m.
<‘hImeric stimuh were prepared ofthe words RED.GREEN. YELLOW and BLUE.cach word printed m an Ink color different from the color spelled on either half of the chzmcra (Fig. 3). Subject\ acre asked to (I) designate the mk color (Stroop test) or (2) designate the color-word (Reverse-Stroop test), their response\ bang either to (I ) <elect from among four colored patche\, (2) select from among four color-words prlnted in black ink, or (3) name the color.
Chlmerlc stimuli of black-outllned draulng\ of an elephant. htrawhcrrleh. an orange. pea\. honana\. ;\ fox, :I

COLOR-MAlCHING, COLOR-NAMINti AND COLOR-MEMORY IN SPLIT-BRAIN PATIENTS 527
EXAMPLE OF
CHIMERIC STIMULUS
PERCEPTUAL COMPLETION IN THE TWO HEMISPHERES
YELLOW - 0
GREEN - q BLUE -
RED - q DARK GRAY _ q
FK;. 1. An example of a chimeric stimulus from the Spotted-Wing Butterfly test, and representation of
perceptual completion by the two cerebral hemispheres.
flamingo and grapes were prepared. and presented by tachtstoscope (Fig. 4). Subjects were asked etther to (I) select from among erght colored patches the typical color of the object, (2) name the typical color of the object, (3) select from among eight pictures a matching object, or (4) name the object. When tachistoscopic testing was complete, pictures were shown tn free vision and subjects were asked to name them. Then. colors were named by the experimentor and stmjects were asked to point to the matching picture. These latter tests were run as a check that patients could. m fact. identify the ptctures and could recognize, when colors were named, which objects had these colors.
RESULTS
1. Butterfly te.st.s
(a) Numing colors. Results for the naming portion of the Butterfly tests (Table 1) show that wing colors in the RVF were generally named with a high degree of accuracy, whereas, except in a few instances, colors seen in the LVF were not named more frequently than would be expected by chance.
L.B. and A.A. were exceptions in displaying above-chance naming of colors in the LVF. For L.B., this was true on all three tests, and A.A. manifested LVF color-naming on the four- color test. Inspection of the EOG records showed that both these subjects were on fixation

528
FORM CHOICES
JEKKE LEVY and VOLWYN TKEVARIHEN
EXAMPLES OF CHIMERIC STIMULI
0
17
A
COLORED-FORM CHOICES COLOR CHOICES
FIG 2. Examples of chimeric stimuli from the Colored-Forms test and form. colored-form. and color
choices.
GREEN + YELLOW
INK COLORS
- RED
----- BLUE
FIG. 3. Examples of chlmerlc stimuli from the Stroop and Reverse-Stroop test
when LVF stimuli were named, but both displayed post-stimulus eye shifts towards the LVF when LVF colors were verbalized, consistent with CRWITZ and DAVIS’S finding [18] that post-stimulus eye shifts were significantly correlated with the region of a display correctly reported.
(b) Matching colors. On the matching portion of the test (Table 2) patients fell into three groups with respect to asymmetry patterns. A.A. had either a RVF bias (on the three-color

COLOR-MATCHING. COLOR-NAMING AND COLOR-MEMORY IN SPLIl-BRAIN PATIENTS 529
EXAMPLE OF CHIMERIC
STIMULUS
ELEPHANT FLAMINGO PEAS FOX
ORANGE GRAPES STRAWBERRIES BANANAS
Fm. 4. An example of a chimeric stimulus from the Memory Colors test and the line-drawings from which chimeric stimuli were constructed.
Table 1. Distribution of color-naming responses on the Butterfly tests
3-Color naming
4-Color naming
A.A. L.B. N.G. C.C. R.Y.
A.A. L.B. N.G. C.C.
Spotted- wing names
A.A. L.B. N.G.
LVF Error RVF Both
9
0 17 1 0 1 3 0 15 0 0 18 0 0 18 0
2 13 0 0 8 15 0 24 0 0 24 0
3 21 0 6 9 7 0 24 0
LVF*
R:F
LVFt RVFf vs “S
error error
<O.OOl R: ns o.noso2
<O.OOl R ns <O.OOl R ns t0.001 R ns
0.002 O.“o:O R <O.OOl
<O.OOl R ns <O.OOl R ns
to.001 R ns 0.066 R <O.OOl
<O.OOl R ns
< 0.001 0.063
<o.oot < 0.001 < 0.001
<O.OOl <O.OOl <O.OOl < 0.001
< 0.001 <O.OOl < 0.001
*Either exact probability or x2 tests, as appropriate, were used for a two-tailed comparison of LVF and RVF response frequencies; the comparison was restricted to trials in which only one response was given.
tSee Appendix for description of statistical techniques. The probabilities are one-tailed tests since the question of interest is whether correct responses exceed chance. Both single-response and double-response trials were counted in determining whether accuracy for a given visual field exceeded chance.
$The letter R or L indicates an asymmetry of responses in favor of the RVF or the LVF, respectively.

530 JEKKF LFVY and COLWYN ,-KEVAKrHtN
Table 2. Distribution of color matching responses on the Butterfly tests
A.A. L.B.
LVFf LVFt RVFt Hand vs v’; v’; x field
LVF Error RVF Both RVF error error effect
2 h 40 0 iO.001 R: Ilb < 0.001 II\
4x 0 0 0 < 0.00 I L rc 0.001 ns II5
3-Color N.G. 47 0 I 0 < 0.001 L < 0.00 I ns n\
matching C.c‘.( LH)$ 15 0 3 6 0.00x L < 0.001 0.00’ C.C.(RH) 2 0 IO 12 0.03X R < 0.00 1 i 0.001
c 0.00 1
R.Y.(LH) ‘1 0 15 0 ns 0.002 c: 0.001 R.Y (RH) 0 0 ‘4 0 < 0.01 R
i 0.00 I II\ < 0.001
A.A. 2x 0 19 I “S i 0.001 i 0.001 n\
4-C&r L.B. 45 0 I ’ <O.OOl L cc 0.00 I 0.037 II\
matching N.G. 44 I 3 0
c 0.001 1. < 0.001 n\ n ‘;
<‘.C.( LH J 17 0 7 0 O.OhX L / 0.001 c 0.00 I C.C.(RH) 0 0 74 0 < 0.00 I R II\ s 0.001
< 0.00 I
A.A h I? 30 0 < 0.001 R 0.004 < 0.00 I II \
Spotted L.B. 73 17 7 I < 0.02 1. c 0 001 O.DO4 ns
wing makhing N.G. ‘0 25 ? I) <O.OOl I. -: 0.001 II\ II\
(‘.<‘ 4 20 73 0 < 0.001 R II\ c-0 001 n\
IThe letter R or L designates an asymmetry in favor of RVF or LVF matches, rcqxctlvely. \‘The designation LH or RH stands for left-hand or right-hand pointing. respectively. Pointing hand ib not g~\cn
when there was a complete absence of a hand hq field interaction.
and spotted-wing tests) or hemispheric symmetry (on the four-color test). with above-chance performance for both visual fields on the four-color and spotted-wing tests. L.B. and N.G. had a LVF bias, but L.B. was also above chance for RVF matching on the four-color and spotted-wing tests. Unihemispheric control was reduced for both A.A. and L.B. from the three-color test to the spotted-wing test. Both C.C. and R.Y. showed a significant hand x field interaction. the LVF dominating with left-hand pointing and the RVF dominating with right-hand pointing. R.Y., tested only on the three-color test, also had an overall RVF bias. summing across pointing hands. and C.C. had a RVF bias on both the four-color and the spotted-wing tests.
The left-hemisphere bias for A.A. and for R.Y. may reHect asymmetric damage to their right hemispheres. However. AA. displayed no signiticant bias on the four-color test and had a predominance of responses to the LVF stimuli. Also, C.C. is likely to have more left- than right-hemisphere damage, but manifested a left-hemisphere bias on the four-color and spotted-wing tests. The direction of bias for C.C. and the variation across tasks in asymmetry patterns for A.A. are not easily interpretable on a brain-damage hypothesis.
C.C.‘s responses on the three-color test, with both left-hand and right-hand pointing. indicated possible bilateral access to colors since he produced a substantial number of double responses. Since single responses to RVF stimuli with left-hand pointing and single responses to LVF stimuli with right-hand pointing did not exceed chance, the double responses are likely to have been mediated by the right hemisphere with left-hand pointing and the left hemisphere with right-hand pointing. If so, then the ipsilateral hemisphere had access to colors,consistent with inferences that can be drawn from A.A.‘s and L.B.‘s capacities to name colors projected in the LVF.

COLOR-MATC-HINC,. COLOK-NAMING AND COLOK-MEMORY IN SPLll-BKAIN PA I IEN IS 531
2. Colored-Forms tests
(a) Naming ofcolors,forms, or color undfbrm. Color naming (Table 3) was similar to that on the Butterfly tests. Naming of RVF colors was well above chance in all subjects, but naming of LVF colors only exceeded chance for L.B. However, although LVF naming was not above chance in N.G., neither was there a significant difference in the frequency with which LVF and RVF colors were named, and the possibility that N.G., like L.B. and A.A., can occasionally name colors projected to the right hemisphere cannot be ruled out.
For form naming, all subjects were above chance in the RVF, but neither A.A. nor N.G. had a significant asymmetry between visual fields, and particularly for A.A., the possibility must be entertained that he could name forms projected in the LVF.
When both colors and forms had to be named, all patients showed a highly significant RVF naming ability, but L.B. had no asymmetry between fields and also was significantly and highly above chance at naming forms and colors projected in the LVF. Although he was not significantly above chance at LVF naming of forms when only the form had to be reported, an inspection of the distribution of L.B.‘s responses suggests the possibility that he might have been able to name LVF forms under this condition also.
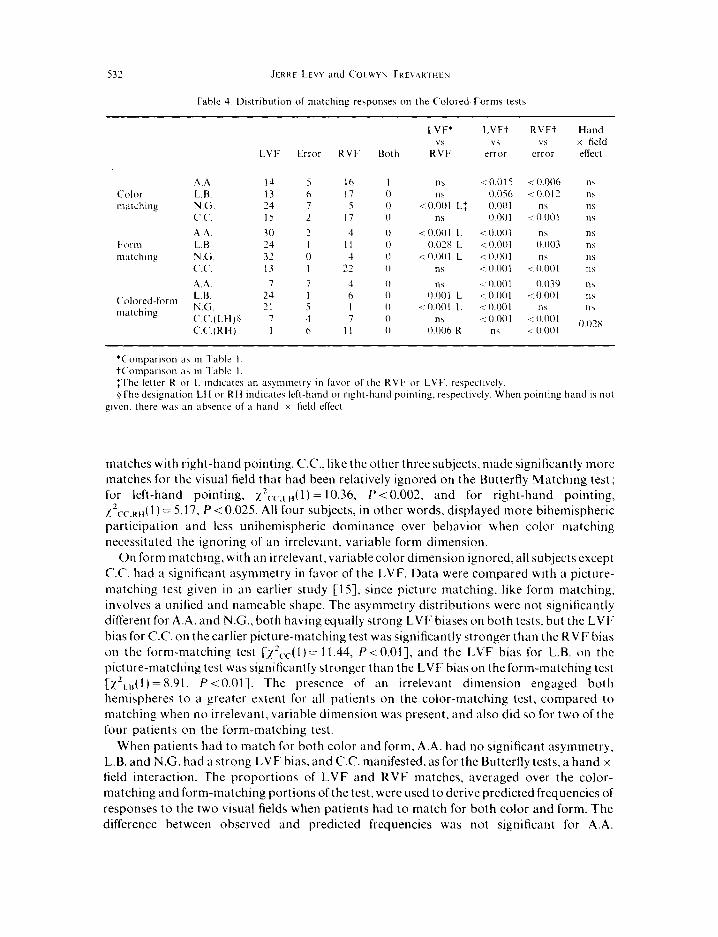
(b) Matching q/‘colors,fi,rms, or color and ,fi)rm. The most striking aspect of the color- matching data (Table 4) when the form dimension had to be ignored is the degree of symmetry of the two hemispheres in controlling responses, relative to the comparable (i.e. three-choice) Butterfly Matching Test. A.A., L.B., and N.G. all manifested significantly more bihemispheric participation for color matching of colored forms than for the three-choice Butterfly test [xzAA( 1) = 17.76. P < 0.001; x2,.8( 1) = 34.77. P < 0.001; Exact P,, = 0.0261. Since C.C. had a hand x field interaction on the three-choice Butterfly Matching test, responses for the two hands separately had to be compared. Table 4 collapses C.C.‘s responses over hands because the hand x field interaction for color matching here was not significant [x2( 1) = 2.98, P= 0.085] and went in the opposite direction from the hand x field interaction on other tests. Thus, for color matching on the Colored-Forms test, C.C. had five LVF matches and 12 RVF matches with left-hand pointing and 10 LVF matches and 7 RVF
Table 3. Distribution of naming responses on the Colored-Forms test
LVF Error RVF Both
LVF* LVFt RVFi vs “S “S
RVF error WlOr
A.A. 2 4 I? 0 0.013 R: “S
Color L.B. 7 0 4 2 ns 0.002 naming N.G. 6 2 I 0 0 ns ns
C.C. 3 0 15 0 0.008 R “S
A.A. 6 1 II 0 ns 0.062 Form L.B. 2 2 IO 4 0.039 R ns naming N.G. 4 3 9 I ns ns
C.C. 4 3 II 0 0.059 R ns
0.03x <O.OOl
0.019 < 0.00 1
0.003 0.002 0.046 0.029
Colored-form L.B. 5 0 6 7 ns <O.OOl <O.OOl naming N.G. 1 4 14 0 0.001 R ns < 0.00 1
C.C. 1 3 I4 0 0.001 R ns < 0.001
*Comparisons as in Table I. tcomparisons as in Table I. fThe letter R or L indicates a RVF or LVF asymmetry. respectively.
532 JERKF LEVY and (‘OLWYh ~KLVAKIHt.N
Table 4. Distribution of matching response5 on the Colored-Forms testa
A.A. L.U. N.G. CT.<‘.
AA L.H N.G. <CC
A.,~
Colored-form L.H.
matching N.G.
LVF Error RVF Roth
LVF* vs
RVF
14 5 I6 I Ilb 13 6 17 0 ns 74 7 5 0 < 0.001 L: I5 2 17 0 “S
30 2 4 0 i 0.00 I L
24 I I1 0 0.02x L
32 0 4 0 ~0.001 L
13 I 22 0 ns
7 7 4 0 “S 24 I h 0 0.001 L 21 5 I 0 i 0.00 I L
7 3 7 0 ns I 6 II 0 0.006 R
LVFt vs
error
10.015 0.056 0.001 0.001
-c 0.001 i 0.001 c 0.001 < 0.001
RVFt “LI
error
CI 0.006 < 0 0 I‘?
n\ i 0.00 1
“h 0.003 n\
< 0.001
0.039 < 0.001
n\ 8 0.001 c 0.001
Hand x field effect
‘C‘ompwwn as m Table I. tComparlwn as in Table I. :The letter R or L indicates an asymmetry in favor of the RVF or LVF. ~respect~ccly $The dwgnation LH or RH indutes M-hand or right-hand pointing, respecllvely. When pointinp hand IS not
given. there was an absence of a hand x field effect.
matches with right-hand pointing. C.C., like the other three subjects, made significantly more matches for the visual field that had been relatively ignored on the Butterfly Matching test; for left-hand pointing, x2 c,-,,H( 1) = 10.36, P < 0.002. and for right-hand pointing, ~~,-c.~~,( 1) = 5.17, P < 0.025. All four subjects, in other words, displayed more bihemispheric
participation and less unihemispheric dominance over behavior when color matching necessitated the ignoring of an irrelevant, variable form dimension.
On form matching, with an irrelevant, variable color dimension ignored, all subjects except C.C. had a significant asymmetry in favor of the LVF. Data were compared with a picture- matching test given in an earlier study [IS], since picture matching, like form matching, involves a unified and nameable shape. The asymmetry distributions were not significantly different for A.A. and N.G., both having equally strong LVF biases on both tests. but the LVF bias for C.C. on the earlier picture-matching test was significantly stronger than the RVF bias on the form-matching test [~2~.c(l)= 11.44, P<O.OI], and the LVF bias for L.B. on the picture-matching test was significantly stronger than the LVF bias on the form-matching test [x’,,,,( I ) = X.91, P < O.Ol]. The presence of an irrelevant dimension engaged both hemispheres to a greater extent for all patients on the color-matching test, compared to matching when no irrelevant, variable dimension was present. and also did so for two of the four patients on the form-matching test.
When patients had to match for both color and form, A.A. had no significant asymmetry, L.B. and N.G. had a strong LVF bias, and C.C. manifested, as for the Butterfly tests, a hand x field interaction. The proportions of LVF and RVF matches, averaged over the color- matching and form-matching portions of the test. were used to derive predicted frequencies of responses to the two visual fields when patients had to match for both color and form. The difference between observed and predicted frequencies was not significant for A.A.

COLOR-MAlCHING. COLOR-NAMINti AND COLOR-MEMORY IN SPLII-BRAIN PATIENTS 533
[xzAA( 1) = 0.07, P > 0.803 nor for the left hand of CC. [x~,--,~~ (Ya,e’s correc,ed, (1) = 1.56, P>O.20], but was significant for L.B. xzLs(l)= 7.00, P<O.Ol], for N.G. [‘2Nc(I)=7.68, P<O.Ol], and for the right hand of C.C. [I’ CC,RH( 1) = 8.33, P < O.Ol]. (On form matching, C.C. made six LVF matches and 12 RVF matches with the left hand and 7 LVF matches and 10 RVF matches with the right hand.) A.A. and the left hand of C.C. showed just as much bihemispheric participation when matching for color and form as for matching each separately, with the other dimension being irrelevant, but L.B., N.G. and the right hand of C.C. manifested greater asymmetry and less bihemispheric engagement when matching for both color and form than for matching each separately. Task complexity per se, evidently, was the only important factor in inducing bihemispheric engagement for A.A. and for the left hand of C.C., but for L.B., N.G. and the right hand of C.C., the presence of an irrelevant. variable dimension that had to be ignored was apparently important in engaging both hemispheres.
3. Stroop and Reverse-Stroop tests
(a) Naming of co/or-words or ink colors. Only two patients, L.B. and N.G., were tested on the Stroop and Reverse-Stroop tests, and their results were practically identical and were combined (Table 5). For the naming of either color words, or for the naming of ink colors, there was a strong RVF bias, and naming of LVF stimuli occurred in only one instance on the color-word portion and in one instance on the ink-color portion, not more frequently than would be expected by chance. The ink-color naming task contrasts with L.B.‘s results for naming on the Butterfly tests and for naming of color and form on the Colored-Form test; in these latter cases, he showed above-chance naming of LVF colors. Possibly, naming of ink colors when an incongruent word was spelled required a greater allocation of attention by the left hemisphere than for other color tasks, precluding awareness of LVF colors. As KINSBOURNE [19] has suggested, perceptual biases for one or the other side of space may be magnified when a hemisphere is activated by task demands.
(b) Matching of color-words or ink colors. When patients were required to read stimulus words for meaning or when they were required to read a choice word for designation of either a stimulus word or ink color, a strong RVF bias emerged. However, on the color-word task when choices were words printed in black ink, the LVF was also matched at an above-chance level, possibly because matching could be mediated by the form invariant between stimulus and choice, with the meaning of the word ignored. In an earlier study, L.B. and N.G. had a strong LVF bias for simple matching of words [16]. On the ink-color identification task where choices were words printed in black ink, the LVF stimuli were also matched at an above-chance level. Given the strong color-matching bias of L.B. and N.G. for the LVF, even the necessity to read choice words did not totally inhibit manifestation of LVF matching.
A highly significant LVF bias appeared on the ink-color identification task when choices consisted of colored patches, but matches for the RVF were performed well above chance also. As for color matching on the Colored-Forms test, an irrelevant, variable form dimension was present, resulting in bihemispheric engagement. If the color-matching performance of L.B. and N.G. on the Colored-Forms test is used to derive a predicted distribution of LVF and RVF matches on the ink-color identification task with colored
patches as choices, predicted and observed frequencies do not differ [x2( 1) = 2.05, P > 0.151. In contrast, if predicted frequencies are derived from the comparable (i.e. four-choice) Butterfly Matching test, predicted and observed frequencies differ significantly [Exact P<O.OOl]. The analyses show that the degree of bihemispheric engagement for L.B. and

534 J~KK~ LI VY and C‘OIV+YN TKCVAKIHI r\,
Table 5. Distributions of naming and matchmg response\ on the Reverse-Stroop :md Stroop tests
Task
Color word identification Point-to-color Point-to-word Name color word
Ink color identilication Point-to-color Point-to-word Name ink color
LVF Error
4 71 x 3: I lb
50 z 3 Y
I 0
RVF- Both
LVF*
R;\F
LVl- I\
error
hl 0 < 0.00 I R “\ 40 0 i 0.001 R 0.009 22 0 < 0.00 I R 11,
IS I** -c 0.001 L ‘. 0.00 I 53 ,** 4 0.001 R 0 045
23 0 ( 0.00 I R 11,
Hand x licld effect
i 0.001 nh c 0.00 I n\ < 0.001
Data itre the comhincd results of L.B. ;md N.G. who did not differ m their pattern\ of rcyx,n\e\ *Statistical tests were performed :is described in Tahlc I : dcsignatlonj of R and L indlutc a\ynmetrle\ ofthc RVI,
and LVF, respectively. tlnk color of LVF matched SIX times: RVF; once $Ink color of LVF matched twice; RVF once #Ink color of LVF named. Color word of LVF matched once; RVl- once.
’ Color word of LVF matched twice. **Double responses were produced by L.B Ofthe I I errorsmadeforcolor-word identilicatlon.nine werefor ink color\ofthc LVF:md only two for inhcolor\
of the RVF (Exact P = 0.033. one-talled).
N.G. was comparable on the color-matching portion of the Colored-Forms test and on the ink-color-matching portion of the Stroop test, with colored patches as choices, and that on both tests, unihemispheric dominance over matching performance was diminished as compared to the Butterfly tests.
(c) Nature rferrors. Too few errors were made on the ink-color identification task to permit a meaningful analysis, but of the 11 errors made on the color-word identification task, 9 were errors giving ink colors for the LVF and only two were errors giving ink colors for the RVF (Exact P =0.033, l-tailed), an outcome that is not unexpected given the strong LVF bias of L.B. and N.G. for color-matching. Evidently, this bias resulted in intrusion of the right hemisphere for the color-word identification task. The opposite intrusion, where words projected to the left hemisphere interfere with ink-color identification, was demonstrated by SCHMII)T and DAVIS [IO] in normal people, reflecting the well-known Stroop interference effect. In the present case, a Reverse-Stroop interference effect appeared in individuals having a strong right-hemisphere bias for the matching of nameable colors. Whether such a Reverse- Stroop interference would appear in people having no bias for the matching of nameable colors or a left-hemisphere bias is unknown.
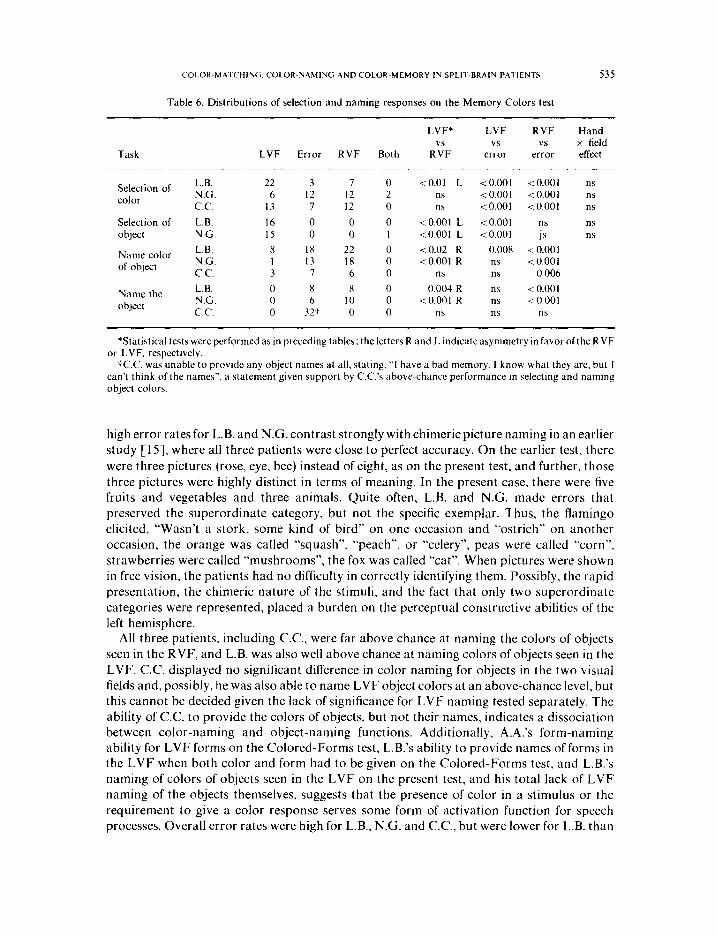
(a) Naminy c$color.s urd ohjccts. When subjects were asked merely to name the object tachistoscopically flashed, all correct responses were for RVF stimuli, but error rates were high (Table 6). C.C. was totally incapable of providing object names, saying “I have a bad memory. I know what they are, but I can’t think of the names.” The anomia for C.C. and the
COLOR-MATCHING, COLOR-NAMING AND COLOR-MEMORY IN SPLIl-BRAIN PATIENTS 535
Table 6. Distributions of selection and naming responses on the Memory Colors test
Task
LVF* LVF RVF Hand “S “S x field
LVF Error RVF Both R:F error error effect
Selection of L.B. 22 3 7 0 <O.Ol L <O.OOl <O.OOl ns color N.G. 6 12 12 2 ns < 0.001 < 0.001 ns
C.C. 13 7 12 0 ns <O.OOl <O.OOl ns
Selection of L.B. 16 0 0 0 <O.OOl L < 0.001 ns “S
object N.G. 15 0 0 I <O.OOl L <O.OOl js ns
Name color L.B. 8 18 22 0 <0.02 R 0.008 c 0.001 of object N.G. I 13 18 0 <O.OOl R ns <O.OOl
C.C. 3 7 6 0 ns ns 0.006
Name the L.B. 0 8 8 0 0.004 R ns < 0.001 object N.G. 0 6 10 0 <O.OOl R ns <O.OOl
C.C. 0 32t 0 0 ns ns ns
*Statistical tests were performed as in preceding tables; the letters R and L indicate asymmetry in favor ofthe RVF or LVF. respectively.
tC.C. was unable to provide any object names at all, stating, “I have a bad memory. I know what they are, but I can’t think of the names”, a statement given support by C.C.‘s above-chance performance in selecting and naming object colors.
high error rates for L.B. and N.G. contrast strongly with chimeric picture naming in an earlier study [1.5], where all three patients were close to perfect accuracy. On the earlier test, there
were three pictures (rose, eye, bee) instead of eight, as on the present test, and further, those three pictures were highly distinct in terms of meaning. In the present case, there were five fruits and vegetables and three animals. Quite often, L.B. and N.G. made errors that preserved the superordinate category, but not the specific exemplar. Thus, the flamingo elicited, “Wasn’t a stork, some kind of bird” on one occasion and “ostrich” on another occasion, the orange was called “squash”, “peach”, or “celery”, peas were called “corn”, strawberries were called “mushrooms”, the fox was called “cat”. When pictures were shown in free vision, the patients had no difficulty in correctly identifying them. Possibly, the rapid presentation, the chimeric nature of the stimuli, and the fact that only two superordinate categories were represented, placed a burden on the perceptual constructive abilities of the left hemisphere.
All three patients, including C.C., were far above chance at naming the colors of objects seen in the RVF, and L.B. was also well above chance at naming colors of objects seen in the LVF. C.C. displayed no significant difference in color naming for objects in the two visual fields and, possibly, he was also able to name LVF object colors at an above-chance level, but this cannot be decided given the lack of significance for LVF naming tested separately. The ability of C.C. to provide the colors of objects, but not their names, indicates a dissociation between color-naming and object-naming functions. Additionally, A.A.‘s form-naming ability for LVF forms on the Colored-Forms test, L.B.‘s ability to provide names of forms in the LVF when both color and form had to be given on the Colored-Forms test, and L.B.‘s naming of colors of objects seen in the LVF on the present test, and his total lack of LVF naming of the objects themselves, suggests that the presence of color in a stimulus or the requirement to give a color response serves some form of activation function for speech processes. Overall error rates were high for L.B., N.G. and C.C., but were lower for L.B. than

536 JFKKF Lr VY and COL.HYN TKFVAK~HFN
on the object-naming portion of the test. In free vision, patients identified colors of the eight objects correctly.
(b) Motchirzg c~f‘colors trrld objects. Only L.B. and N.G. were tested for simple object matching; both showed a complete LVF bias, and no errors were made. The LVF bias is congruent with their performance in a previous study on a picture-matching task [lS]. The absence of errors shows that the high error rates observed when object naming was tested could not have been due to simple perceptual problems. At the least, we can conclude that patients could perceive the stimuli sufficiently well to produce errorless matching. It should be noted, also, that the object-matching portion of the test was given prior to the object- naming portion for L.B. and N.G., so that by the time the naming task was presented, they had had the benefit of experience on object matching.
All three patients showed highly significant above-chance performance with both hemispheres for matching colors to line-drawings, and only L.B. had a significant asymmetry (favoring the right hemisphere). Error rates were lower on the color-matching portion of the test than on the color-naming portion of the test. For matching, there were three errors out of 32 responses for L.B. (9”,,). 12 errors out of 34 responses for N.G. (35’Z,,), and seven errors out of 32 responses for C.C. (22”J, as compared to 38”,, errors for L.B., 41 “,, errors for N.G., and 44’,;, errors for C.C. when they named colors of line-drawings. The increase in errors for naming as compared to matching was significant for the group of three patients [x2(3)= 10.62, P<O.O2]. Also, on the matching task, L.B. had the lowest error rate and the
greatest degree of LVF bias, whereas N.G. had the highest error rate and the greatest degree of RVF bias. The (I priori probability that patients should be ordered on error rates in accordance with the degree of LVF bias is 116. The pattern of results suggests that, although both hemispheres are capable of matching colors to line-drawings at an above-chance level. the right hemisphere is more accurate than the left. and that to the extent that the left hemisphere assumes control of processing, error rates increase.
DISCUSSION
All patients displayed good ability to name colors seen by the left hemisphere, confirming the results of studies in the introduction of this paper. L.B. and A.A. were also able to name colors projected to the right hemisphere, and on the color-naming portion of the Colored- Forms test, N.G. may have been able at times to name LVF colors. In addition, form naming of LVF stimuli was achieved by A.A. in the form-naming portion of the Colored-Forms test, for L.B. when both color and form had to be named in this test, by L.B. when colors had to be provided for line-drawings, and possibly, by C.C. in this latter test. Verbalizations of LVF stimuli were confined to conditions when colors were present either in the stimulus or the response. No LVF naming was observed for object identification, a condition where no chromatic variable was present. The mechanisms whereby verbalization of LVF stimuli is accomplished in split-brain patients are unknown, but the data suggest that the presence of color in the stimulus or the necessity to name a color activates speech processes. The ability of C.C. to name colors of objects, while being unable to supply their names, also indicates a special association between chromatic qualities and speech. Conceivably, brainstem systems, capable of mediating information between hemispheres, and supplying arousal input to the brain, are activated selectively by color in stimuli or by the necessity to name colors.
In simple color matching, with no other variable dimensions present, patients typically showed a strong asymmetry in favor of one hemisphere or the other, but the direction and

COLOR-MATCHING. COLOR-NAMING AND COLOR-MEMORY IN SPLIT-BRAIN PATIENTS 537
degree of the asymmetry varied between patients and within patients across different conditions. Thus, for the simplest task (three-choice Butterfly test), A.A. had a strong RVF bias, and L.B. and N.G. had strong LVF biases. C.C. showed equally strong biases in opposite directions as a function of pointing hand, the left hemisphere dominating matching when the right hand was used, the right hemisphere dominating matching when the left hand was used. R.Y. had an overall RVF bias, but this was diminished with left-hand pointing. As task complexity increased, the magnitude of asymmetric control decreased. This was observed for A.A. and L.B. even on the spotted-wing test, where matches to LVF stimuli exceeded chance for A.A. and matches to RVF exceeded chance for L.B., in contrast to their performance on the three-choice Butterfly test.
On the color-matching portion of the Colored-Forms test, where a variable, irrelevant form dimension was present, all patients showed bihemispheric engagement, in contrast to the results of the three-choice Butterfly test. This was also seen on the Stroop test when colored patches were provided as choices. The asymmetry in behavioral control of C.C. and L.B. was reduced on the form-matching portion of the Colored-Forms test, when the irrelevant, varying dimension was color, as compared to picture matching in an earlier study [ 151. When both color and form had to be matched., and neither dimension was irrelevant, A.A. and the left hand of CC. manifested as much bihemispheric engagement as when an irrelevant dimension had to be ignored, but L.B., N.G. and the right hand of C.C. showed a significant increase in asymmetry. For these latter, the presence of an irrelevant dimension that had to be ignored was, apparently, of importance in inducing both hemispheres to participate in the task.
The change in hemispheric usage over tasks shows, first, the lability of the hemispheres for matching nameable colors. The hemisphere that dominates and controls behavior in one circumstance does not necessarily do so in other circumstances. It is not unreasonable to suppose that shifts in control between hemispheres reflect changes in strategies applied as task demands change. Second, the nature of changes observed shows that unihemispheric control is diminished with increases in task complexity, regardless of which hemisphere had dominated responding in the simpler task. We suggest that as task complexity increases, the capacity of a hemisphere to retain absolute control over processing decreases, and the other side of the brain is encouraged to collaborate in solving the problem at hand. If so, this could have implications for hemispheric cooperation in normal people. While either hemisphere alone in the normal brain might tend to assume control of well-practiced, habitual functions within its domain of specialization, both hemispheres might be called into play for cognitively complex, nonhabitual, and creative processes. The relative normality of split-brain patients in everyday life is likely to derive from the fact that most requirements of day-to-day living depend on overlearned responses where extensive, redundant contextual support is provided.
The findings from the Stroop and Reverse-Stroop tests support inferences from other investigations. There was a strong left-hemisphere dominance over behavior whenever naming responses had to be given or words had to be read for meaning, either as the stimulus or as the choice. When the response entailed pointing to a colored patch, L.B. and N.G., the only patients tested on this task, manifested the same LVF bias for ink-color identification that they had shown on the color-matching portion of the Colored-Forms test and of less magnitude than produced on the Butterfly tests. Intrusion errors appeared when patients had to identify the color-word, reAecting intrusion of ink colors projected to the right hemisphere. This Reverse-Stroop interference effect seems analogous to the Stroop interference effect,

538 JFKKE LFVY and COLWYIV TKEVAKIHEN
where the word interferes with the identification of ink color. In the SCHMIT and DAVIS [ 101 study of normal people, there was a left-hemisphere asymmetry for Stroop interference, consistent with the left-hemisphere superiority for word identification that they observed. They found no evidence of hemispheric asymmetry for Reverse-Stroop interference: both hemispheres showed an increase in reaction time of equal magnitude for identification of color words printed in incongruent ink colors, as compared to the condition where they were printed in congruent ink colors. Given their observations, however, of equal reaction times for identification of ink colors when the carrier word was congruent (635 msec for the RVF and 634 msec for the LVF), one would not expect an asymmetry for Reverse-Stroop interference. In their sample of subjects, the hemispheres were symmetric for matching of nameable colors, in contrast to L.B. and N.G. who manifested a strong LVF bias. It would be of interest to determine whether normal individuals having an asymmetry for the matching of nameable colors display a correlated asymmetry in Reverse-Stroop interference.
The results from the Memory Colors test show clearly that both hemispheres are capable of matching colors to line-drawings, and the lower error rates for the matching portion of the test as compared to the naming portion of the test, as well as the association between error rates and the degree of bias for right-hemisphere matching, suggest that the right hemisphere is somewhat more competent than the left. The difference in errors for matching vs naming cannot be due merely to the presence of a choice set in the former case since we counted as correct any naming response that could be an appropriate color for the object depicted in the line-drawing; naming responses, in other words, had more freedom for some objects than did matching responses with respect to what could be counted as correct. The evidence for better performance of the right hemisphere is consistent with what has been inferred regarding the nature ofcolor representations in the two hemispheres. If object colors are represented in the left hemisphere in terms of elaborate conceptual structures and in the right hemisphere in terms of sensory-rich images, almost any familiar object would provide access to its color by the right hemisphere, but depending on various experientially derived associations, the left hemisphere may or may not have object colors easily available. This interpretation would predict that color-matching performance by the right hemisphere should be less variable from person to person than color-matching performance by the left hemisphere. This was the case in the PEN:~AI. [X] study, both for colors presented in the lower half of the visual ticld [F( 124,124) = 1.53, P = 0.0251 and for colors presented in the upper half of the visual field [ E‘( 124.124) = I .49. P < 0.051.
Our data from the Memory Colors test arc consistent with studies showing that the right hemisphere has access to good imagistic representations and that the left hemisphere has a good ability to derive feature characteristics of objects from the conceptual structure it possesses. Our results arc also concordant with findings that color representation for the right hemisphere is at least as good as that of the left. and better for colors resistant to verbal description. The dcticits of aphasic patients with left-hemisphere lesions in matching colors with line-drawings [S. 12 141 can be interpreted as reflecting abnormal interhemispheric regulation, with processing control retained by the damaged left side of the brain, preventing the right hemisphere from manifesting its abilities in matching colors to objects. This interpretation could also explain why dcnsc receptive aphasias occasionally occur with left- hemisphere lesions when receptive disorders of such an extreme magnitude are not observed in the right hemisphere of split-brain patients.
It is reasonable to infer. on the basis ofour data and those of others, that the approximately equal competence of the left and right hcmisphcres to discriminate and remember namcahle

COLOK-MArCHINti. COLOK-NAMtNcj AND COLOK-MEMORY IN SPLIT-BKAIN PATIEN IS 539
colors is an equality of performance only, that the nature of color representations and of the strategies used in processing by the two sides of the brain differ. We suggest that variations observed among split-brain patients in asymmetry patterns reflect differences in the ways they integrate color information, differences that not only characterize different individuals, but that appear within the same person as task demands change. Both sides of the human brain appear to have an amazing aptitude for solving problems with which they are confronted in most circumstances. each using whatever strategies and processes that are incorporated in it. The differences between the hemispheres seem to reside predominantly in the nature of multipurpose processes used, and differences in performance are likely to occur only when special and rather artificial tasks are presented that are fundamentally insusceptible to solution by one or the other set of laterally differentiated functions.
Ack~~/edyemrnr.~~~ This research was conducted in the laboratory of Professor ROGER W. SPERRY at the
California Institute of Technology with support from the Frank P. Hixon Fund and by the U.S. Public Health Service Grant MH 03372 01 to Professor SP~KRY. Preparation of this report was supported by a Spencer Foundation Grant and by a U.S. Public Health Service Biomedical Grant, PHS 5 SO7 RR 07029 13, to Jt RKI
Lf v’v. The subjects were patients of Drs. pHtI.ff’ Vtxif f. of the California College of Medicine and JOSI PH &)(a \ of the Ross Loos Medical Group, Los Angeles. We are indebted to Professor SPERKY for the opportunity to carry out OUI
I.
2. 3.
4. 5.
6.
7.
x. 9.
10.
1 I.
12.
13.
14.
15.
16.
17.
IX.
19.
studies in his laboratory and with-his support
REFERENCES
KfNseotlRNf . M. and WARRfNciToN, E. Observations on colour agnosia. J. Nrurd Nrurosurg. f?\ychiuf. 27, 296 299, 1964. Cancrtt.r:v, M. Acquired anomalies of colour perception of central origin. Bruin 88, 711 724. 1965. Gt:sc.tLwtNt), N. and Ft;stt.t o, M. Color-naming defects in association with alexia. Awh. Nrurol. 15, 137 146, 1966. Osetlav. J. M., OXWRY, S. M. and HtIMPffREY, N. K. Varieties of colour anomia. Bruin 92, 847 X60, 1969. DI- RtY(t, E. and SPINNLFK. H. Impaired performance on color tasks in patients with hemispheric damage. Cortrv 3, 194 217. 1967. Sr.orsr, G. and SPINNLI R, H. Colour imperception in unilateral hemisphere-damaged patients. J. Nertrol..
Ncwosury., P.$ychiut. 33, 22 28, 1970. CAIVIA\I, E., S(x)-rst, G., and StJthixt.f R. H. Color impairment in pattents with focal excisions of the cerebra1 hemispheres. Nr,urop.s~ch~~/f~~iu 16, 491 496, 197X. PI NUAI., B. E. Human cerebral asymmetry in color discrimination. Nruropsychok~~yiu 15, 563 568, 1977. DIMONI), S. and Bi AIIHOVT, G. Hemisphere function and color naming. J. r-up. P.qho/. %, X7 91. 1972. SCIIMIT, V. and DAVIS, R. The role of hemispheric specialization in the analysis of Stroop stimuli. Acru /?~~~ho/.
38, 149 15x, 1974. MII Nt-n, B. Brain mechanisms suggested by studtes of temporal lobes. In Bruin Mechanisms Un&r/yiny Spwch ud Lanquuye, F. L. DARI.I Y (editor). Grune & Stratton, New York, 1967.
Cortru, R. and Kt l.Tl R, S. Cognitive impairment of aphasics in a colour-to-picture matching task. Corre\- 15, 235 245, 1979. DI Rt.N/t, E., F~AGIONI, P...%oI rr,G., and SIVNNI f K, H. Impairment in associating colour to form.concomitant with aphasia. Bruin 95, 293 304, 1972. BASSO, A., FA(,I IOUI. P., and SPIYNLfK. H. Non-verbal colour impairment of aphasics. Nrurop.\~c.ho/ogicr 14, 1X3 193, 1976. LI vt, J.. TKI.\ ,\K IIII h, C.. and St+ant. R. W. Perception of bilateral chimeric figures following hemispheric deconnexion. Bruin 95, 61 78, 1972. Lt VI. J. and TKI \AK~ ttt:N, C. Perceptual. semantic and phonetic aspects of elementary language processes in split-brain patients. Bruin 100, 105 118, 1977. LI VY, J. and TKI vAKmf N. C. Metacontrol of hemispheric function in human split-brain patients. J. r.rp Psycho/.: Hum. Pcrcrpt. Prrlr,rtn. 2, 299 312, 1976. CKOLIT/, H. F. and DAVIS, W. Tendencies of eye movement and perceptual accuracy. J. cup. Psycho/. 63, 495 498, 1962.
KtNsaot~aNt . M. The cerebral basis of lateral asymmetries m attention. .4cru t?syctrot. 33, 193 201, 1970.

540 JI:KKL LEVY and COLWYN TKEVAKIHEN
APPENDIX
We considered three approaches for calcuiatmg whether performance is above chance for a given visual field. with the likelihood of falsely accepting the null hypothesis progreqslvely decreasmg through the three methods, A, B, and C. Method B was the procedure chosen for the analyses in this paper since It provides conservatism in acceptmg the null hypothesis without an unreasonably high likelihood of its being falsely rejected. As a simple example, consider a three-choice task where one choice conforms to the LVF stimulu?. one to the RVF stimulus, and one choice is erroneous. Suppose that in 24 trials, LVF responses are made IO times, RVF responses are made 12 times, and erroneous responses are made twice. The question, III this example. oh whether LVF choices occur slgnlficantly more often than chance would allow.
In method A, we absume that the right hemisphere always controls responses; thus. that any response other than one matching the LVF stimulus IS an error. Under this assumption,chance predicts that of 24 responses, eight would conform to the LVF stimuluh and I6 would represent other responses (eight RVF and eight errors). For the example. we calculate %‘(I) =0.75. P =0.3X44, two-talled, or :=().X7. P=O 1922. one-talled, and we conclude that LVF responses were not above chance. Method A was rejected because of the unreasonable assumption that the hemisphere under consideration always controls responding through all trials. regardless of whether “other” responses are true errors or responses conforming to the stimulus of the other hemisphere
Method C allocates erroneous responses according to the proportion of LVF and RVF responses In the present example, the LVF contributes IO/22 = 45”,, of all correct responses. and we assume that (0.45)(2)=0.91 errors should be attributed to the right hemisphere. There are, therefore. IO.91 responses under consideration. and assuming that correct and erroneous responses are equlprobablc. chance predicts 5.45 LVF responses and 5.45 errors. Ciiven !O LVF and 0.91 erroneous responses. n’( I ) = 7.60, P = 0.005X. two-talled. or ; = 2.76, P = 0.0029, one- tailed, and we conclude that LVF response% occurred more often than chance would allow. The allocation of error\ to the hemisphere5 In method C might be overly generous in that. at least theoretlcally. the hemisphere contributing fewer correct responses I\ more subject to error than the hemispherecontrihutlng morecorrect response\. Method C was rejected because the probabdlty of falsely rejcctlng the null hypothesis seemed too high.
In method B, we reason that RVF responses. LVF- rc<ponscs. and errors all ha\e a chance prohablhty of I 3: 1.e. that. compared to each other. LVF- rcsponscs and error, arc equiprobable. With RVF responses excluded from consideration. chance predicts that of the I2 remaining responses. SIX WIII he LVF response\ and \IX WIII he error\. Given IO LVF responses and two error\, /‘( I ) = 5 33. P= O.OZOX. two-tailed. or z = _.. ’ 31, P= 0.0104. one-talled. and we conclude that LVt‘responscs occurred more often than chance would predict. The conservatism of method B 1’1 the assumption that all erroneou, responses arc committed by the hemisphere uho\c performance I\ being evaluated. (When performance of the RVF is assessed. all erroneous responses are attributed to the left hemisphere.) The non-conservatlvc assumption of method B (and of method C) IS that none of the rchponsc, to the visual field not under consideration are under control of the hemisphere whose performance IS hcmg assessed. Method B war chosen as the procedure for calculatmg prohabihties that performance for agIven visual field was abate chance \~nce It seems to represent a reasonable balance between the overly conservative assumptionc ofmethod A and the overly generous assumptions of method C. Method B also ha5 the \ lrtuc ofpermlttmg double-response trials to he summed wsith single-response trials. Thus, If in 16 trials, there are eight RVF smglc responses. SIX LVF smgle rc\pon\e\. two errors, and four doubly-correct responses, we hove I7 correct RVF respomeh. IO correct LVF rc\ponses. and tv,o errors. and we compute probahllltley for the two visual lields, under method B. as hcfore.
A one-tailed statistical test was used to assess whether pcrformnncc in a given visual Geld was above chance \ince the null hypothesis IS that correct responses do not exceed chance. 4 two-talled test would assc:Ss whether calrlro correct responses were above chance 01’ whether erroneou, rcqponses were above chance. A two-tailed test was u\cd to compare the distributmns of correct responses between visual fields since. m this case. WC are slmultaneou\ly interested in an asymmetry in favor of either visual field.
When expected frequencies were less than 4 5. exact prohabdlty tests. using the blnomlal expanhlon. were adopted, Thus. for cxamplc, on the apotted-wing Butterfly test, there were I2 possible choices. In ahsessmg the chance that color-matching for A.A. in the LVF was Ggnificant (Table 2). we note that this patlent produced six correct LVF responses and I7 crrorlr. Under method B. the choice matching the RVF is cxcludcd from consideration, and the right hemisphere has a I/ I I chance of being correct and IO.!1 I chance of being wrong. The expected frequency of correct response5 out of a total of IX trials under consideration IS (1. I I ) (I 8) = I 64. far too small to permit use of the I’ statistic. Therefore. the hinomlal probahillty for pcrformancc as good as that ohscrvcd or better was employed. representing the sum of probabllltle< for 6:1X correct through IX IX correct, 1.~‘. (lX564)(l’ll)” (lo/ll)‘2+(31824)(11.11)~ (lO,ll)“+ (l)(I.‘Il)‘fi (10,1I)“=0.004.

COLOR-MATCHING. COLOK-NAMING AND COLOR-MEMORY IN SPLIT-BRAIN PATIENTS 541
RQsumi! : Oes stimulus chimeriques bilatkaux Qtaient projetes au tachlstascope
B des malades split-brain dans le but d'Qtablir les asym6tries h&misp!Sriques
dans la dBnomination, l'appariement et la &moire des couleurs d'objets familiers.
On constatait un biais important dans la denomination des couleurs des stimulus
projstes B l'h8misphSre gauche mais quelques malddes Btaient aussi au-dessus du
niveau de chance dans la dBnomination des cauleurs du stimulus projet B l'hgmis-
phke droit. Dans les tkhes d'appariement de couleurs. les malades diffkaient
dans la direction et le degr6 de contr5le asym6trique des r&ponses : les types
d'asymetrie varlaient avec les malades selon les diffkrentes tbches. l'asymetrie
du contr6le h6mlsph6rique augmentant avec la complexit de la tache. Les 2 hemis-
ph$i-es avaient des performances bien au-dessus du niveau de chance pour la m&moire
de5 couleurs d'objets dessines au trait et l'hQmisph6re droit semblait avair des
performances quelque peu meilleures que le gauche.
Zusammenfassung
Bilaterale chimsrische Stimuli wurden tachistoskopisch Split-Brain-
Patienten dargeboten mit dem Ziel, hemisphsrische Asymmetrien beim
Farbenennen, Farbenzuordnen und fiir das GedZchtnis fiir Farben van ver-
trauten Objekten herauszufinden. Es gab einen starken Effekt fiir das Benennen
van Farben, wenn die Stimuli in die linke Hemisphlre projiziert warden, aber
einige Patienten zeigten such iiberzufXllige Leistungen beim Farbbenennen,
wenn die Stimuli in die rechte Hemisphare kamen. Bei den Farbzuordnungs-
aufgaben variierteh die Leistungen der Patienten, und die Muster der Asymmetrie
variierten iiber unterschiedliche Aufgaben bei ein und demselben Patienten. Mit
zunehmender Komplexitat der Aufgaben nahm die Symmetric der hemisphtiischen
Kontrolle zu. Beide Hemisphsren waren iiberzuf%llig gut im GedLchtnis fiir die
Farben van Objekten, die als Strichzeichnungen dargeboten wurden, die rechte
Hemisphlre schien der linken etwas iiberlegen zu win.